Introduction
Gastric cancer (GC) remains a gastrointestinal
cancer with high global incidence and mortality (1). Despite major advances in perioperative
chemotherapy and immunotherapy, the overall survival (OS) of
patients with advanced disease remains poor. The widespread
development of chemoresistance poses a major clinical challenge.
Previous studies have revealed that radiotherapy and platinum-based
and other chemotherapy drugs can trigger therapy-induced senescence
(TIS) by inducing DNA damage or replication stress (2,3). As an
inherent cellular stress response, TIS has a dual role in cancer
treatment. Senescent cells enter stable cell cycle arrest,
suppressing tumor proliferation. Conversely, residual senescent
cells can secrete senescence-associated secretory phenotype (SASP)
factors, remodel the tumor immune microenvironment, foster immune
evasion, and potentially drive tumor recurrence (4). Consequently, a promising ‘one-two
punch’ strategy has gained traction, which aims to trigger
senescence and then specifically to clear or reprogram (5,6).
At the molecular level, the PI3K/AKT pathway is
commonly dysregulated in GC, driving tumor cell proliferation and
viability, regulating metabolic reprogramming, and contributing to
resistance to cytotoxic drugs (7,8).
Therefore, this is regarded as an important mechanism of treatment
resistance and disease recurrence in GC. PI3K/AKT, as the ‘master
switch’ of tumor metabolism, controls glucose and lipid metabolism,
macromolecule biosynthesis and autophagy (9). Moreover, senescent cells exhibit
unique vulnerability to PI3K/AKT inhibition (10,11),
providing a potential opportunity for subsequent ‘senolysis’
interventions. Metabolic reprogramming, particularly the balance
reconstruction of oxidative phosphorylation (OXPHOS) and
glycolysis, regulates TIS (12,13).
Clinically, platinum-based regimens are vital for
the first-line treatment of GC. Previous studies indicate that
platinum drugs can trigger cellular senescence in various tumor
models, accompanied by cell cycle arrest and enhanced SASP
secretion (14,15). The stress response nuclear receptor
NR4A1 has emerged as a remarkable molecular bridge, linking these
chemotherapeutic stimuli to the senescent phenotype. As a cellular
stress sensor, NR4A1 coordinates critical signaling and metabolic
adaptations and its ability to regulate pathways, including
PI3K/AKT, makes it a credible mediator of chemotherapy-induced
senescence. Consequently, simultaneously exploiting NR4A1-mediated
PI3K inhibition to drive metabolic reprogramming during
chemotherapy-induced senescence may amplify the antitumor effect of
senescence and create opportunities for combining senolytic or
immunotherapy strategies.
This investigation demonstrated that under
chemotherapy-induced stress, the NR4A1-PI3K/AKT axis drives
metabolic reprogramming and promotes the acquisition and
maintenance of the senescence state in GC cells. Clarifying this
molecular mechanism may provide actionable therapeutic targets for
optimizing the ‘one-two punch’ sequential treatment strategy. This
may offer novel approaches to overcome chemotherapy resistance and
prevent GC recurrence.
Materials and methods
Cell culture and senescence
induction
AGS and MKN45 human GC cell lines were acquired from
the Procell Life Science & Technology Co., Ltd. and maintained
in RPMI-1640 containing 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.) at 37°C with 5% CO2 condition. Senescence was
induced by treating AGS cells with 2.5 µM oxaliplatin (OXA) and
MKN45 cells with 5 µM OXA for 48 h.
Cell Counting Kit-8 (CCK-8) assay
To determine the 50% inhibitory concentration and GC
cells were plated in 96-well plates (3.0×103 cells/well)
and treated with a gradient of OXA concentrations: 0, 1.25, 2.5, 5,
10, 20, 40 and 80 µM. After 48 h, 10 µl CCK-8 reagent (Beyotime
Institute of Biotechnology) was added per well, followed by a 1 h
incubation in the dark at 37°C. Absorbance at 450 nm was recorded
with a microplate reader (EPOCH-SN; BioTek Instruments, Inc.).
Senescence-associated β-galactosidase
(SA-β-Gal) staining
Cellular senescence was assessed with a commercial
kit (Beyotime Institute of Biotechnology). Cells were rinsed with
phosphate-buffered saline (PBS), fixed, incubated with SA-β-Gal
staining solution overnight at 37°C without CO2, and
then washed with PBS. Images were captured using an optical
microscope (IX73; Olympus Corporation), and the percentage of
SA-β-Gal-positive cells was quantified.
Western blotting
GC cells were lysed using RIPA buffer (Beijing
Solarbio Science & Technology Co., Ltd.) with protease
inhibitors (Beyotime Institute of Biotechnology). Protein
concentration was then measured with a BCA kit (Beyotime Institute
of Biotechnology). Proteins (20 µg per lane) were resolved by
10–15% SDS-PAGE and then transferred onto PVDF membranes
(MilliporeSigma). Membranes were blocked with 5% non-fat milk at
room temperature (RT) for 2 h, washed with TBST (0.1% Tween-20).
They were then incubated with primary antibodies overnight at 4°C
and subsequently with secondary antibodies for 1 h at RT. Protein
band was developed with ECL reagent (Beyotime Institute of
Biotechnology), captured with an Amersham Image Quant 800 imaging
system (Amersham; Cytiva), and analyzed for gray-level. Antibody
details were listed in Table
SI.
Immunofluorescence
Cells (1.0×105) were fixed with 4%
paraformaldehyde, blocked with 3% BSA (Wuhan Servicebio Technology
Co., Ltd.) for 30 min at RT. Cells were then incubated overnight at
4°C with a primary antibody against phospho-H2A.X (1:100; cat. no.
GB111841; Wuhan Servicebio Technology Co., Ltd.), followed by
incubation with a Cy3-conjugated goat anti-rabbit IgG secondary
antibody (1:300; cat. no. GB21303; Wuhan Servicebio Technology Co.,
Ltd.) for 50 min at RT. Nuclei were visualized by DAPI (2.0 µg/ml)
counterstaining for 10 min. Images were captured using a Nikon
Eclipse C1 fluorescence microscope (Nikon Corporation). Antibody
details were listed in Table
SI.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated and reverse transcribed into
cDNA using commercial kits (Tiangen Biotech Co., Ltd.) according to
the manufacturer's instructions. RT-qPCR was conducted with SYBR
Green Master Mix (Tiangen Biotech Co., Ltd.) under the following
thermocycling protocol: 95°C for 2 min, 95°C for 5 sec, 60°C for 10
sec and 72°C for 15 sec for 40 cycles. Relative gene expression was
calculated using the 2−ΔΔCq method (16). Primers were listed in Table SII.
Experimental design for multi-omics
analysis
AGS cells were divided into two experimental groups:
An untreated control group and an oxaliplatin-induced senescent
group. For transcriptomic, proteomic and untargeted metabolomics
analyses, four independent biological replicates were included for
each group (n=4 per group).
Transcriptomic sequencing
Total RNA was isolated with TRIzol reagent (Beyotime
Institute of Biotechnology). RNA concentration was measured using a
NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Inc.).
RNA samples with a concentration ≥10 ng/µl and a total amount ≥1 µg
were considered qualified for subsequent library construction. RNA
integrity was verified by 1% RNase-free agarose gel electrophoresis
stained with GelRed. cDNA libraries were constructed with the Hieff
NGS® Ultima Dual mode mRNA Library Prep Kit (cat. no.
12309ES; Shanghai Yeasen Biotechnology Co., Ltd.) and were then
amplified by PCR. Library concentration was evaluated using the
DNA1000 assay Kit (cat. no. 5067-1504; Agilent Technologies, Inc.),
and the final library loading concentration was 3 ng/µl. Sequencing
was performed on an Illumina NovaSeq X Plus platform with a
paired-end 150 bp strategy by Gene Denovo Biotechnology Co.,
Ltd.
Next, using DESeq2 software, differentially
expressed genes (DEGs) were selected based on the criteria of |fold
change| ≥2 and an adjusted P<0.05. Volcano plots and heatmaps
were plotted with the online Omicsmart platform (http://www.omicsmart.com). Kyoto Encyclopedia of Genes
and Genomes (KEGG) enrichment analysis was performed on DEGs.
Proteomic analysis
Following protein extraction and enzymatic
digestion, the peptides were separated by an UltiMate 3000 liquid
chromatography system (Thermo Fisher Scientific, Inc.) and analyzed
using a timsTOF Pro2 mass spectrometer from Bruker Daltonics. DIA
proteomics data were analyzed using Spectronaut 18 (Biognosys AG)
under default parameters. The ideal extraction window was
dynamically determined by the Spectronaut using iRT calibration and
gradient stability. A false discovery rate threshold of 1% was
applied at both the precursor and protein levels. Peptides passing
this filter were quantified using the MaxLFQ algorithm to generate
normalized protein group abundances. Mass spectrometric data were
acquired in DIA mode using diaPASEF. Differentially expressed
proteins were defined by a |fold change| >1.2 and a
Benjamini-Hochberg adjusted P<0.05.
Untargeted metabolomics
The samples were mixed with
methanol/acetonitrile/H2O (2:2:1, v/v/v). Next, mixtures
were sonicated twice on ice for 30 min each and centrifuged at
14,000 × g (20 min, 4°C). The supernatant was collected and
evaporated under vacuum. Dried extracts were redissolved in an
acetonitrile aqueous solution, centrifuged, and the supernatant was
collected for injection. Separation was conducted on an Agilent
1290 Infinity UHPLC system interfaced with an AB Sciex TripleTOF
6600 Q-TOF mass spectrometer, and quality control was conducted
using QC samples. Data from the positive (pos) and negative (neg)
ion modes were processed independently. Raw data were transformed
into the mzML format utilizing ProteoWizard. Subsequent processing,
including peak alignment, retention time correction, and extraction
of peak area, was carried out with XCMS software.
Metabolite identification and data preprocessing
were then performed; the steps were as follows: Features with
missing values exceeding 50% were removed; remaining missing values
were filled using k-nearest neighbor imputation; and features with
a relative standard deviation greater than 50% were filtered out.
Significantly altered metabolites were selected with the variable
importance in projection (VIP) score ≥1 in OPLS-DA model, followed
by filtering with a univariate t-test (P<0.05).
Gene expression profiling interactive
analysis (GEPIA2) survival analysis
The prognostic significance of NR4A1 expression in
GC was evaluated via the GEPIA2 platform (http://gepia2.cancer-pku.cn/), which combines RNA-seq
data from The Cancer Genome Atlas (TCGA) and the Genotype-Tissue
Expression projects (17). For the
OS analysis, TCGA-stomach adenocarcinoma (STAD) cases were
segregated into ‘high’ and ‘low’ NR4A1 expression groups according
to the median transcripts per million values as the cut-off.
Survival outcomes between groups were compared using Kaplan-Meier
analysis with the log-rank (Mantel-Cox) test. The hazard ratio
along with 95% confidence intervals was calculated.
Cell transfection
The short hairpin RNA (shRNA) NR4A1 lentiviral
vector was constructed by Shanghai GeneChem Co., Ltd., using the
pFU-GW backbone. The sequences for the shRNAs are as follows: A
non-targeting control (sh-NC, 5′-TTCTCCGAACGTGTCACGT-3′) and three
NR4A1-specific sequences (sh-NR4A1#1 sense:
5′-TGGTGAAGGAAGTTGTCCGAA-3′, antisense:
5′-TTCGGACAACTTCCTTCACCA-3′; sh-NR4A1#2 sense:
5′-GATTGACAGTATCCTGGCCTT-3′, antisense:
5′-AAGGCCAGGATACTGTCAATC-3′; sh-NR4A1#3 sense:
5′-CTCCTTCAAGTTCGAGGACTT-3′, antisense:
5′-AAGTCCTCGAACTTGAAGGAG-3′).
GC cells plated in 6-well plates were exposed to
viral particles in medium containing 5 µg/ml polybrene at a
multiplicity of infection value of 20. Following incubation under
standard conditions (37°C, 5% CO2), medium was replaced
with a fresh complete medium and cultured the cells for another 48
h. The lentiviral transduction was conducted over 72 h.
Successfully transduced cells were selected with 2.0 µg/ml
puromycin for 48 h, followed by an additional 48 h recovery period
in fresh complete medium prior to subsequent experiments.
Lactate content assay
Following the instructions of a commercial lactate
content detection kit (Wuhan Servicebio Technology Co., Ltd.), for
lactate assay, L-lactic acid was oxidized by a specific lactate
oxidase to pyruvate and H2O2. In the presence
of peroxidase and chromogens, a colored dye was produced, which was
measured photometrically at 546 nm. The absorbance of the dye,
measured photometrically, was directly proportional to the L-lactic
acid concentration. The results were normalized according to the
total protein content.
ATP content assay
Following the instructions of the ATP content
detection kit (Wuhan Servicebio Technology Co., Ltd.), ATP assay
was based on the principle that luciferase catalyzes the
chemiluminescence reaction of luciferin under the energy provided
by ATP, and within a certain range, the luminous intensity was
directly proportional to the ATP content. The results were
normalized based on the cell count.
Statistical analyses
Statistical analyses and data visualization were
performed using GraphPad Prism software (v9.5.0; Dotmatics) and
shown as the mean ± standard deviation (SD). Significance was
determined using an unpaired two-tailed Student's t-test or one-way
analysis of variance, followed by Tukey's multiple comparisons
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Oxaliplatin induces senescence in GC
cells
To verify whether OXA induce senescence in GC cells,
an in vitro model was established using AGS and MKN45 cells
lines. Briefly, cells were assigned to control (Ctrl) and OXA
groups. CCK-8 assays revealed that OXA reduced AGS and MKN45 cell
viability in a concentration-dependent manner (Fig. S1A and B). Based on these results,
AGS cells were treated with 2.5 µM OXA, while MKN45 cells were
treated with 5 µM OXA, both for 48 h. Senescence was assessed
through SA-β-Gal staining, western blotting, immunofluorescence
detection of γ-H2AX, and qPCR analysis of SASP factors.
SA-β-Gal staining detects cellular senescence by
measuring the SA-β-Gal activity at pH 6.0, which hydrolyzes X-Gal
to generate a blue product (18).
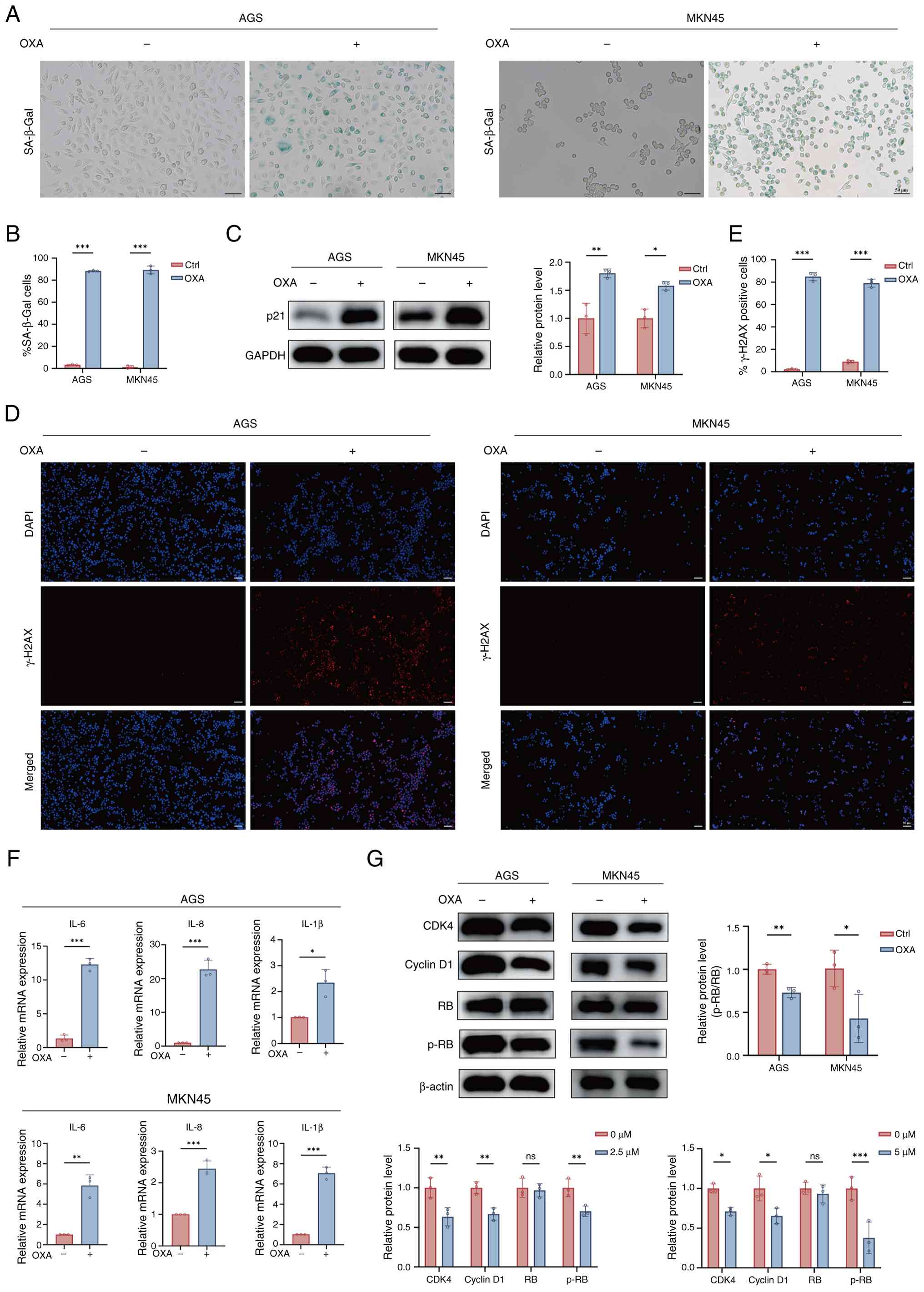
As presented in Fig. 1A, compared
with the Ctrl group, OXA group displayed a greater percentage of
SA-β-Gal-positive GC cells, along with enlarged and flattened
morphology. Quantitative analysis confirmed that the average
SA-β-Gal positivity rates were 88.3% in AGS cells from the OXA
group and 89.32% in MKN45 cells from the OXA group (Fig. 1B).
As a cyclin-dependent kinase inhibitor, p21 is a key
mediator of cellular senescence (19). Western blotting indicated a
significant upregulation of p21 protein in the OXA group compared
with the Ctrl group, suggesting the induction of cell cycle arrest
(Fig. 1C). Since γ-H2AX is a
classic marker of DNA damage and replication stress and is a
central driver of senescence induction (20), its expression was assessed.
Immunofluorescence showed a stronger enhancement of γ-H2AX foci in
OXA group than in Ctrl group (Fig. 1D
and E).
SASP comprises the secretion of diverse inflammatory
cytokines, chemokines and proteases, which affects the tumor
microenvironment (TME) (21). Next,
the mRNA expression levels of major SASP components were evaluated
using RT-qPCR. RT-qPCR analysis indicated a significant
upregulation of various SASP factors, such as interleukin (IL)-6,
IL-8 and IL-1β in the OXA group relative to the Ctrl group in AGS
and MKN45 cells (Fig. 1F).
Finally, to clarify the cell cycle regulatory
network associated with senescence, the expression of key
regulatory proteins was examined (22). Compared with the Ctrl group, CDK4
and cyclin D1 were significantly downregulated in OXA group;
however, total RB protein levels remained unchanged, and p-RB
levels decreased. These results suggested that after OXA induction,
cell cycle progression was inhibited, and RB was maintained in a
hypo-phosphorylated active state (Fig.
1G).
In summary, it was established that OXA induces
senescence in AGS and MKN45 cells, as evidenced by elevated
SA-β-gal activity, p21 upregulation, cell cycle regulator
inhibition, DNA damage accumulation and enhanced SASP
secretion.
Integrated transcriptomic and
proteomic analyses identified the NR4A1-PI3K/AKT axis in
chemotherapy-induced senescence
To systematically elucidate the molecular mechanisms
of OXA-induced senescence in GC cells, integrative transcriptomic
and proteomic analyses of AGS cells were performed by comparing
Ctrl and OXA groups.
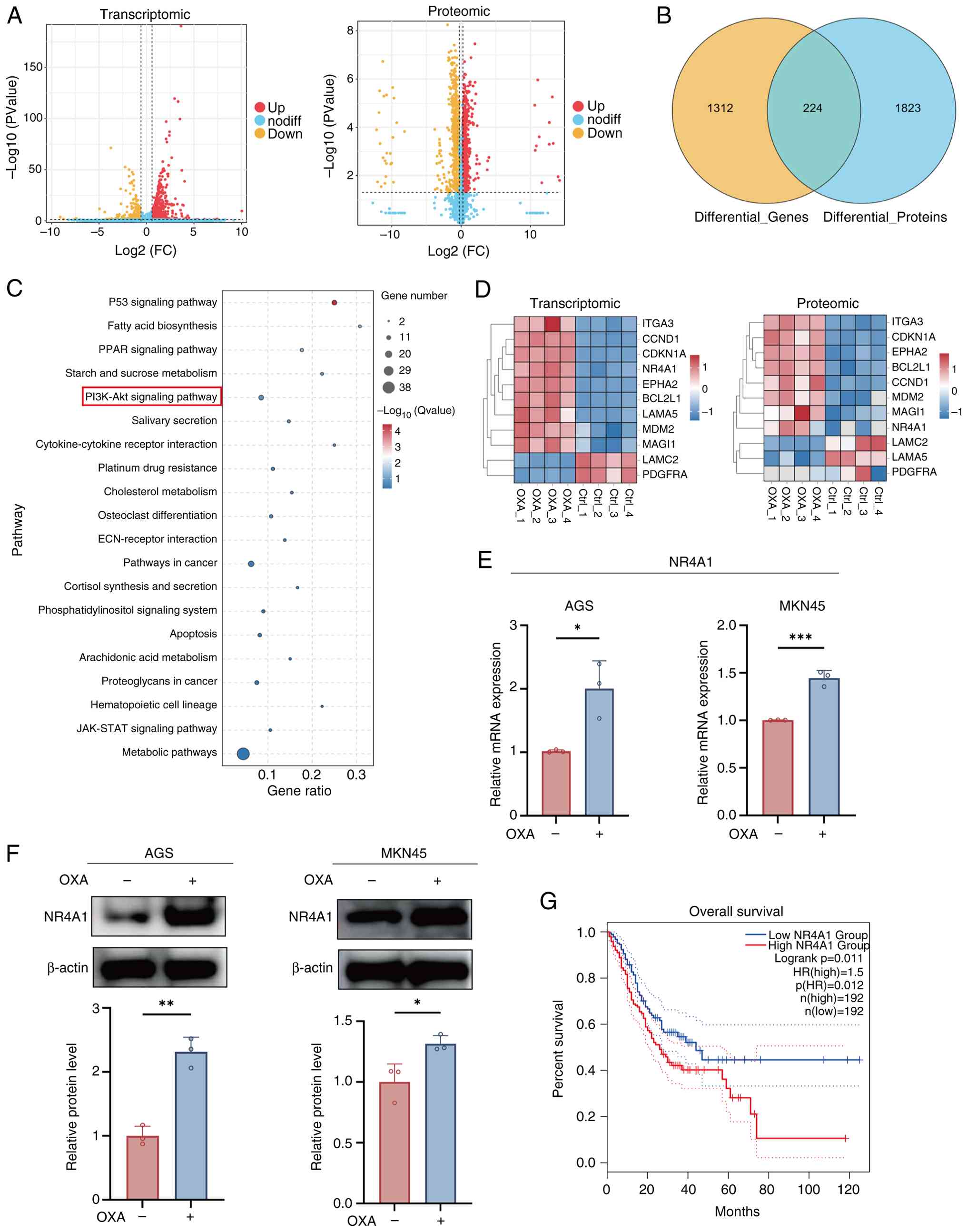
As demonstrated in Fig.
2A, the transcriptomic volcano plot revealed 1,536 DEGs in the
OXA group (|log2FC| >0.585, P<0.05) with 1,193
upregulated and 343 downregulated genes. Proteomic analysis further
identified 2,047 proteins that were significantly altered
(|log2FC| >0.26, P<0.05), including 1,040
upregulated and 1,007 downregulated proteins. Venn diagram analysis
exhibited that 224 molecules were consistently differentially
expressed at both transcriptomic and proteomic levels (Fig. 2B), implicating a central role in
OXA-induced senescence.
KEGG enrichment of the overlapping differentially
expressed molecules revealed marked enrichment in pathways closely
associated with senescence, tumor progression and metabolism,
including ‘p53 signaling pathway’, ‘PI3K-AKT signaling pathway’,
‘platinum drug resistance’ and ‘metabolic pathways’ (Fig. 2C). Notably, the PI3K-AKT signaling
pathway was significantly enriched at both omics’ levels,
highlighting its critical role in chemotherapy-induced senescence.
The expression of PI3K-AKT pathway associated genes and their
corresponding proteins, was then visualized using heatmaps
(Fig. 2D).
Among these candidates, NR4A1 was explored, a
stress-responsive nuclear receptor that is regarded as a key
molecule linking signal transduction with metabolic fate. Emerging
evidence indicates that NR4A1 regulates cellular metabolism and
fate by suppressing PI3K/AKT signaling (23,24).
It was discovered that in GC, NR4A1 expression was elevated at the
transcriptional and protein levels in the OXA group. Consistent
with this, qPCR (Fig. 2E) and
western blotting (Fig. 2F)
confirmed elevated NR4A1 expression in OXA group compared with the
Ctrl group. Moreover, to enhance the clinical value of NR4A1, its
prognostic significance was further analyzed using the TCGA-STAD
dataset via GEPIA2. Notably, Kaplan-Meier survival analysis showed
a shorter OS in patients with high relative to low NR4A1 expression
(Fig. 2G).
Collectively, integrative multi-omics analyses
indicate that OXA induces cellular senescence in GC cells by
upregulating NR4A1 and suppressing the PI3K/AKT pathway. Notably,
NR4A1 expression was induced during OXA-induced senescence in
vitro, whereas intrinsically high NR4A1 expression was
correlated with poor prognosis, indicating a context-dependent role
of NR4A1 in GC.
NR4A1-PI3K/AKT axis promotes
OXA-induced senescence in GC cells
To explore the role of NR4A1 in chemotherapy-induced
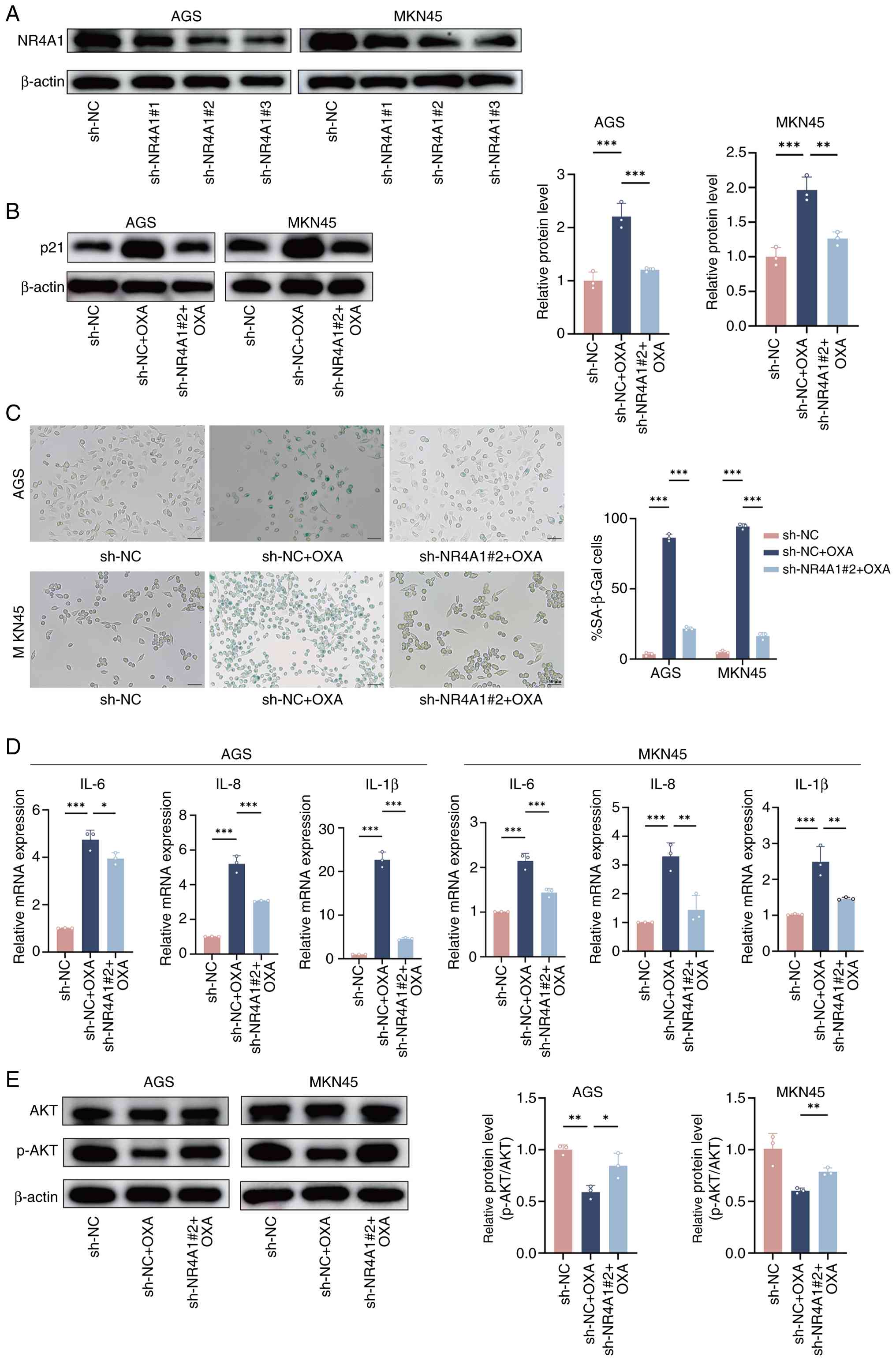
senescence, NR4A1-knockdown AGS and MKN45 cell lines were generated
using shRNA technology. Western blotting confirmed that NR4A1
protein levels were decreased in the sh-NR4A1 groups compared with
the sh-NC group, with sh-NR4A1#2 exhibiting the most efficient
knockdown (Fig. 3A). Consequently,
sh-NR4A1#2 was selected for subsequent experiments.
The senescence marker p21 expression was first
detected. As expected, p21 expression was significantly upregulated
in the sh-NC + OXA group relative to the sh-NC group. However, this
induction was significantly weakened in the sh-NR4A1#2 + OXA group
(Fig. 3B). These results indicated
that NR4A1 knockdown partially reversed the p21-mediated senescence
signal triggered by OXA. Consistently, SA-β-Gal staining indicated
that senescent cells increased substantially in the sh-NC + OXA
group compared with sh-NC, while this increase was significantly
reduced in the sh-NR4A1#2 + OXA group (Fig. 3C). Next, the SASP phenotype was
assessed using RT-qPCR. High expression of typical SASP factors was
found in the sh-NC + OXA group, but this upregulation was
significantly reduced in the sh-NR4A1#2 + OXA group (Fig. 3D).
Based on the aforementioned findings, the PI3K-AKT
pathway was highly enriched in transcriptomic and proteomic
analyses, and it was further explored whether NR4A1 was involved in
regulating this pathway. For this purpose, levels of phosphorylated
AKT at Ser473 (p-AKT) and total AKT, core indicators of PI3K/AKT
pathway activity, were analyzed by western blotting. It was found
that the sh-NC + OXA group significantly suppressed AKT
phosphorylation (decreased p-AKT/AKT ratio). This suppression was
effectively reversed in the sh-NR4A1#2 + OXA group, in which the
p-AKT/AKT ratio was restored (Fig.
3E).
In conclusion, NR4A1 is a critical regulator of
OXA-induced senescence in GC cells. NR4A1 knockdown weakened p21
upregulation, SA-β-Gal positivity and SASP secretion, and restored
PI3K/AKT-associated pathway activity. Collectively, these results
highlight a central role for the NR4A1-PI3K/AKT axis in
chemotherapy-induced cellular senescence.
NR4A1-PI3K/AKT axis-driven metabolic
reprogramming
To assess metabolic alterations during
chemotherapy-induced senescence, untargeted metabolomic profiling
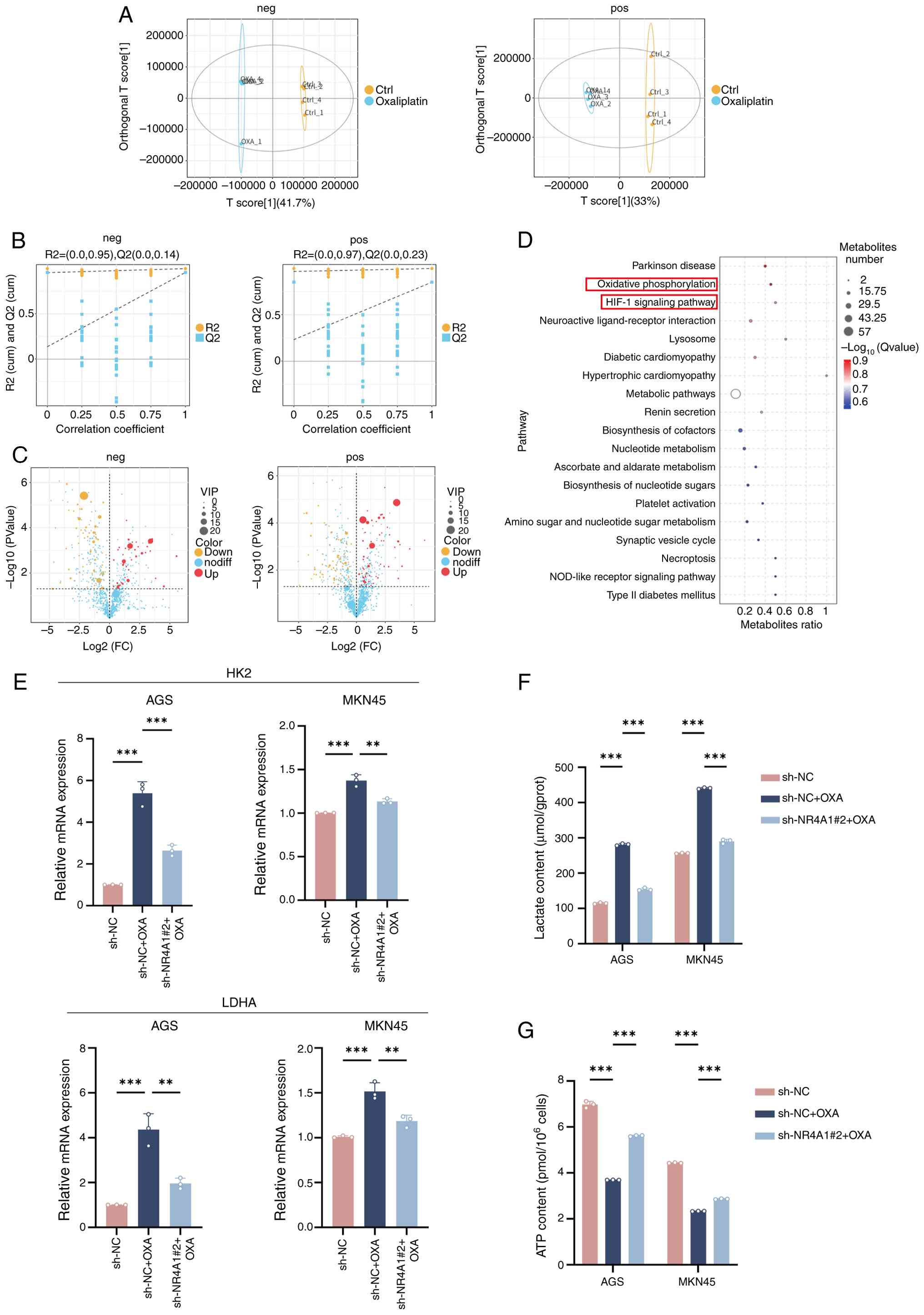
of AGS cells was performed in Ctrl and OXA groups. Score plots
demonstrated an obvious separation between Ctrl and OXA groups in
neg and pos ion detection modes, indicating favorable
reproducibility (Fig. 4A).
Permutation tests further confirmed that the model fits well in
both modes (Fig. 4B), supporting
the metabolomic analysis stability. The aforementioned results
prove the stability of the experiment and the reliability of the
observed metabolic differences. Differential metabolite analysis
revealed pronounced metabolic alterations in the OXA group under
both neg and pos ion modes. Specifically, 41 metabolites were
upregulated and 66 downregulated in the neg mode, while 59 were
upregulated and 76 downregulated in the pos mode (VIP >1,
P<0.05) (Fig. 4C). Following
integration of the differential metabolites from both neg and pos
modes, KEGG pathway enrichment analysis was performed (Fig. 4D). The analysis indicated that these
metabolites were substantially enriched in key metabolic pathways,
including ‘OXPHOS’, ‘HIF-1 signaling pathway’, ‘metabolic pathways’
and ‘nucleotide metabolism’ (Fig.
4D). Notably, the significant enrichment of OXPHOS and HIF-1
signaling, two central energy metabolism pathways, was
significantly enriched. This result suggests that OXA-induced
senescence in GC cells may drive metabolic reprogramming by
inhibiting OXPHOS and promoting glycolysis.
To verify the role of NR4A1 in metabolic
reprogramming, the mRNA expression levels of key glycolytic enzymes
were measured. RT-qPCR analysis demonstrated significant
upregulation of lactate dehydrogenase A (LDHA) and hexokinase 2
(HK2) in the sh-NC + OXA group relative to the sh-NC group, whereas
this effect was significantly reversed in the sh-NR4A1#2 + OXA
group (Fig. 4E). The functional
experiments supported these findings. Lactate levels were
substantially increased in the sh-NC + OXA group and were
significantly reduced in the sh-NR4A1#2 + OXA group (Fig. 4F). Concurrently, ATP detection
showed a notable decrease in cellular ATP content in the sh-NC +
OXA group relative to sh-NC, reflecting impaired OXPHOS function.
Notably, ATP production was restored in the sh-NR4A1#2 + OXA group,
exceeding that in the sh-NC + OXA group (Fig. 4G).
In conclusion, OXA-induced GC cells triggered
extensive metabolic reprogramming characterized by OXPHOS
inhibition and glycolysis enhancement (‘Warburg effect’). NR4A1 is
a central mediator of OXA-induced metabolic reprogramming. NR4A1
knockdown effectively reversed the upregulation of glycolytic genes
(LDHA and HK2) by OXA, alleviated ATP generation suppression, and
reduced lactate accumulation. Building on previous findings, these
data demonstrate that NR4A1-mediated inhibition of the PI3K/AKT
pathway serves as a novel bridge connecting chemotherapy-induced
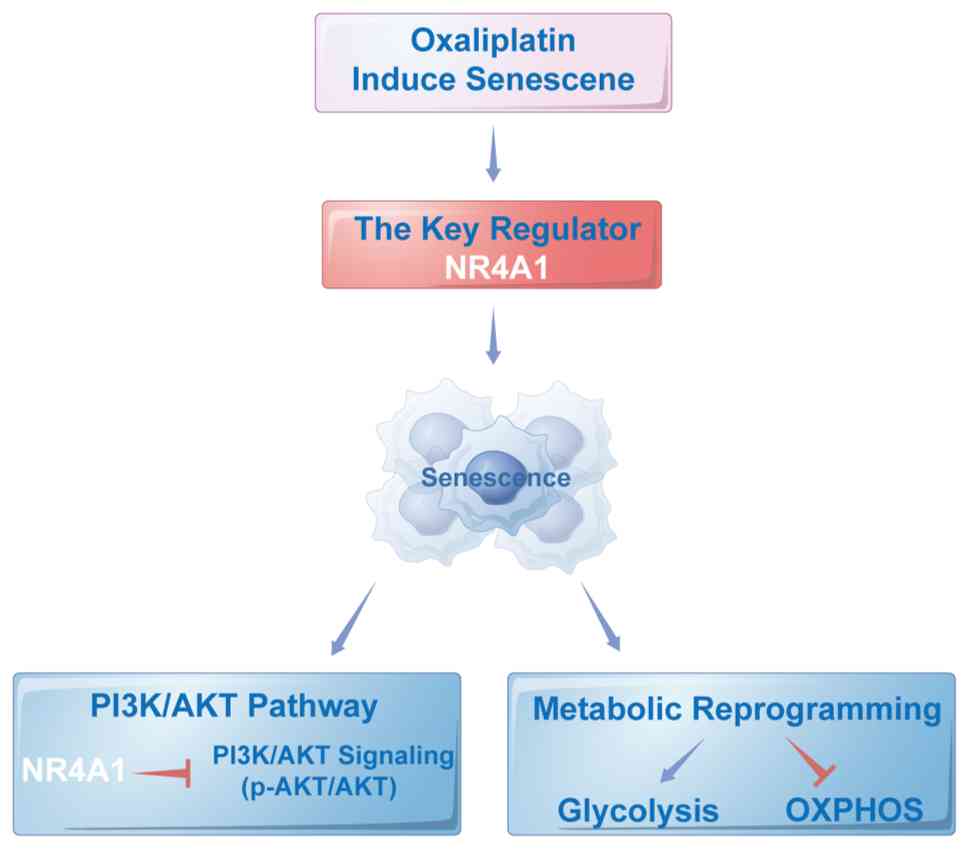
stress to metabolic reprogramming (Fig.
5).
Discussion
GC remains a major global health challenge and is a
prominent cause of cancer deaths. Increasing evidence indicates
that chemotherapy, particularly with platinum-based agents, can
induce TIS. TIS has emerged as a unique cellular response with dual
effects on tumors. While it limits tumor proliferation through
stable cell cycle arrest, the accompanying SASP remodels the TME,
which can promote immune evasion and contribute to recurrence.
Accordingly, understanding the molecular mechanisms of
chemotherapy-induced senescence in GC is essential for identifying
actionable targets and optimizing therapeutic strategies. The
present findings support previous studies that chemotherapy can
induce TIS (25,26). It was first confirmed that OXA
effectively induces a senescence phenotype in GC cells,
characterized by p21 upregulation, elevated SA-β-Gal positivity,
accumulation of γ-H2AX foci, and enhanced SASP secretion.
Subsequently, through integrated transcriptomic and proteomic
analyses, it was observed that the PI3K/AKT pathway is critical for
senescence regulation and NR4A1 was further identified as a
potential regulator of TIS. Functional experiments demonstrated
that NR4A1 negatively regulates the PI3K/AKT pathway, consistent
with previous studies (27,28).
In parallel, metabolomic profiling revealed that
OXA-induced senescence was accompanied by typical metabolic
reprogramming, namely, enhanced glycolysis and suppressed OXPHOS.
These observations strongly support the emerging concept of
‘metabolism-senescence combination’ (29,30).
Based on these findings, it was demonstrated that NR4A1 knockdown
reversed OXA-induced metabolic reprogramming, which was reflected
by the downregulation of glycolytic enzymes (LDHA and HK2),
restored ATP production, and reduced lactate accumulation.
Collectively, these results indicate that NR4A1 improves the
establishment and stabilization of the senescent phenotype by
maintaining PI3K/AKT signaling in a low-activity state and driving
an imbalance between glycolysis and energy metabolism.
The present in vitro findings demonstrate
that NR4A1 is upregulated under OXA-induced stress and promotes a
stable senescent state by suppressing PI3K/AKT signaling and
metabolic reprogramming. However, clinical cohorts reveal a
paradoxical observation between high NR4A1 expression and poor
prognosis in patients with GC. This discrepancy can be explained by
the context-dependent nature of NR4A1 biology (31). In vitro, NR4A1 elevation
represents a short-term stress response that drives tumor cells
toward proliferation arrest and senescence. Conversely,
persistently elevated NR4A1 levels in tumors likely reflect a
distinct long-term program involving metabolic adaptation, chronic
inflammation, or an immunosuppressive microenvironment, all of
which are features commonly associated with adverse outcomes
(32).
Moreover, TIS can inhibit proliferation in the short
term; however, persistent SASP can promote immune evasion,
angiogenesis, EMT, and ultimately facilitate tumor recurrence or
drug resistance. Therefore, NR4A1-driven senescence may
simultaneously confer short-term growth inhibition and long-term
tumor progression risk, consistent with the observed survival
association (33). Notably, tumors
with high NR4A1 expression, despite being more aggressive, may be
more dependent on the NR4A1-mediated senescence pathway, and
therefore, more sensitive to chemotherapy-induced senescence. In
summary, the current mechanistic findings highlight the acute and
pro-senescence effects of NR4A1, whereas clinical data reflect its
complex and long-term association with tumor biology. Collectively,
these findings reveal that NR4A1 has dual and temporally distinct
roles in GC.
From a clinical standpoint, TIS represents a
‘double-edged sword’ in which it inhibits tumor progression and
facilitates immune evasion and tumor recurrence due to SASP
secretion. Therefore, the recently proposed ‘one-two punch’
strategy, which involves the induction of senescence followed by
targeted senolysis, represents a promising anti-cancer therapeutic
strategy (34–36). In the present study, findings
suggest that the intervention of the NR4A1-PI3K/AKT axis and the
NR4A1-metabolic axis may collectively amplify the tumor-suppressive
effect of therapy-induced senescence. However, they simultaneously
provide new opportunities for senolytic and SASP-modulating
interventions. This strategy could potentially improve
chemosensitivity and reduce drug resistance and recurrence
risk.
The present study provides two major innovations.
First, it was proposed that NR4A1 acts as an important regulator of
chemotherapy-induced senescence in GC. Second, the relationship
between the NR4A1-PI3K/AKT axis and metabolic reprogramming was
revealed, providing a new idea for enhancing the aging effect
through metabolic intervention and offering a theoretical basis for
metabolic-targeted anti-cancer therapy. Nevertheless, the present
study has certain limitations. Firstly, experiments were primarily
reliant on GC cell models, which necessitates future validation in
other types of cancer to determine whether the observed mechanisms
are widely applicable. Second, future studies should analyze the
NR4A1 regulatory network and explore its potential in combination
with immunotherapy or metabolic therapy.
In conclusion, the present study established NR4A1
as an important mediator of OXA-induced senescence in GC cells. It
acts by suppressing the PI3K/AKT pathway and driving metabolic
transitions characterized by impaired OXPHOS and enhanced
glycolysis. The present study provides innovative mechanistic
insights into the interaction between chemotherapy, signaling
pathways and metabolic plasticity, and highlights NR4A1 as a
promising therapeutic target for enhancing senescence-based
strategies. Interventions targeting the NR4A1-PI3K/AKT-metabolic
axis may amplify the tumor-suppressive effect of therapy-induced
senescence while reducing its tumorigenic consequences, offering a
reasonable framework for the integration of ‘one-two punch’
strategies or immunotherapy.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Precision Medicine Joint
Cultivation Fund Project of Natural Science Foundation of Hebei
(grant no. H2021402007), the Basic Research Special Project of
Natural Science Foundation of Hebei (grant no. H2022402009), the
Cooperative Project of the Affiliated Hospital of Hebei University
of Engineering (grant no. KFKT2024-05), the Scientific Research
Project of the Administration of Traditional Chinese Medicine of
Hebei (grant no. 2022149) and the Handan Clinical Medicine
Excellent Talent Project (grant no. ZF2023221).
Availability of data and materials
The data generated in the present study may be found
in the Open Archive of the China National Center for Bioinformation
under accessions numbers HRA015931, OMIX014177 or OMIX014178, or at
the following URL: https://ngdc.cncb.ac.cn/.
Authors' contributions
TZ designed the research, conducted the experiments
and wrote the manuscript. YW assisted with the experiments and data
analysis. JZ and XY contributed to data analysis and participated
in data discussion. YS contributed to data interpretation,
participated in data discussion and critically revised the
manuscript. ZZ contributed to study conception, experimental
design, and critical revision of the manuscript. TZ and ZZ confirm
the authenticity of all the raw data. All authors read and approved
the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI
|
|
2
|
Mikula-Pietrasik J, Niklas A, Uruski P,
Tykarski A and Ksiazek K: Mechanisms and significance of
therapy-induced and spontaneous senescence of cancer cells. Cell
Mol Life Sci. 77:213–229. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Faheem MM, Seligson ND, Ahmad SM, Rasool
RU, Gandhi SG, Bhagat M and Goswami A: Convergence of
therapy-induced senescence (TIS) and EMT in multistep
carcinogenesis: Current opinions and emerging perspectives. Cell
Death Discov. 6:512020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Takasugi M, Yoshida Y and Ohtani N:
Cellular senescence and the tumour microenvironment. Mol Oncol.
16:3333–3351. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang L, Lankhorst L and Bernards R:
Exploiting senescence for the treatment of cancer. Nat Rev Cancer.
22:340–355. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
McHugh D, Duran I and Gil J: Senescence as
a therapeutic target in cancer and age-related diseases. Nat Rev
Drug Discov. 24:57–71. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pan W, Tan Y, Chen X, Zeng L, Lv Y and
Yang J: miRNA-204-5p acts as a tumor suppressor in gastric cancer
by inhibiting cell migration, invasion, and glycolysis via the
RAB22A/PI3K/AKT axis. Sci Rep. 15:295362025. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yan S, Hu X, Wu Y, Ye W, Zhu Y, He Y, Zhan
F, Wu W and Ma Z: ITGA4 contributes to 5-fluorouracil resistance by
Up-Regulating PI3K/AKT signaling: evidence from network
pharmacology, molecular docking and experimental verification. Drug
Des Devel Ther. 19:4105–4122. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fontana F, Giannitti G, Marchesi S and
Limonta P: The PI3K/Akt pathway and glucose metabolism: A dangerous
liaison in cancer. Int J Biol Sci. 20:3113–3125. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang CY, Tan XH, Yang HH, Jin L, Hong JR,
Zhou Y and Huang XT: COX-2/sEH dual inhibitor alleviates hepatocyte
senescence in NAFLD mice by restoring autophagy through
Sirt1/PI3K/AKT/mTOR. Int J Mol Sci. 23:82672022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tang Q, Markby GR, MacNair AJ, Tang K,
Tkacz M, Parys M, Phadwal K, MacRae VE and Corcoran BM:
TGF-β-induced PI3K/AKT/mTOR pathway controls myofibroblast
differentiation and secretory phenotype of valvular interstitial
cells through the modulation of cellular senescence in a naturally
occurring in vitro canine model of myxomatous mitral valve disease.
Cell Prolif. 56:e134352023. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Efimova EV, Appelbe OK, Ricco N, Lee SS,
Liu Y, Wolfgeher DJ, Collins TN, Flor AC, Ramamurthy A, Warrington
S, et al: O-GlcNAcylation enhances double-strand break repair,
promotes cancer cell proliferation, and prevents therapy-induced
senescence in irradiated tumors. Mol Cancer Res. 17:1338–1350.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gu J, Wang J, Liu X, Sai K, Mai J, Xing F,
Chen Z, Yang X, Lu W, Guo C, et al: IL-6 derived from
therapy-induced senescence facilitates the glycolytic phenotype in
glioblastoma cells. Am J Cancer Res. 11:458–478. 2021.PubMed/NCBI
|
|
14
|
Mikula-Pietrasik J, Witucka A, Pakula M,
Uruski P, Begier-Krasinska B, Niklas A, Tykarski A and Ksiazek K:
Comprehensive review on how platinum- and taxane-based chemotherapy
of ovarian cancer affects biology of normal cells. Cell Mol Life
Sci. 76:681–697. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Melones-Herrero J, Alcala S, Ruiz-Canas L,
Benitez-Buelga C, Batres-Ramos S, Cales C, Lorenzo O, Perona R,
Quiroga AG, Sainz B Jr and Sanchez-Perez I: Platinum iodido drugs
show potential anti-tumor activity, affecting cancer cell
metabolism and inducing ROS and senescence in gastrointestinal
cancer cells. Commun Biol. 7:3532024. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tang Z, Kang B, Li C, Chen T and Zhang Z:
GEPIA2: an enhanced web server for large-scale expression profiling
and interactive analysis. Nucleic Acids Res. 47((W1)): W556–W560.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee BY, Han JA, Im JS, Morrone A, Johung
K, Goodwin EC, Kleijer WJ, DiMaio D and Hwang ES:
Senescence-associated beta-galactosidase is lysosomal
beta-galactosidase. Aging Cell. 5:187–195. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yan J, Chen S, Yi Z, Zhao R, Zhu J, Ding S
and Wu J: The role of p21 in cellular senescence and aging-related
diseases. Mol Cells. 47:1001132024. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Biran A, Zada L, Abou Karam P, Vadai E,
Roitman L, Ovadya Y, Porat Z and Krizhanovsky V: Quantitative
identification of senescent cells in aging and disease. Aging Cell.
16:661–671. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dong Z, Luo Y, Yuan Z, Tian Y, Jin T and
Xu F: Cellular senescence and SASP in tumor progression and
therapeutic opportunities. Mol Cancer. 23:1812024. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gorgoulis V, Adams PD, Alimonti A, Bennett
DC, Bischof O, Bishop C, Campisi J, Collado M, Evangelou K,
Ferbeyre G, et al: Cellular senescence: Defining a path forward.
Cell. 179:813–827. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ma YL, Kong CY, Guo Z, Wang MY, Wang P,
Liu FY, Yang D, Yang Z and Tang QZ: Semaglutide ameliorates cardiac
remodeling in male mice by optimizing energy substrate utilization
through the Creb5/NR4a1 axis. Nat Commun. 15:47572024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li JM, Song ZH, Li Y, Chen HW, Li H, Yuan
L, Li J, Lv WY, Liu L and Wang N: NR4A1 silencing alleviates
high-glucose-stimulated HK-2 cells pyroptosis and fibrosis via
hindering NLRP3 activation and PI3K/AKT pathway. World J Diabetes.
16:975442025.PubMed/NCBI
|
|
25
|
Kallenbach J, Atri Roozbahani G, Heidari
Horestani M and Baniahmad A: Distinct mechanisms mediating
therapy-induced cellular senescence in prostate cancer. Cell
Biosci. 12:2002022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang D, Zhang JW, Xu H, Chen X, Gao Y,
Jiang HG, Wang Y, Wu H, Yang L, Wang WB, et al: Therapy-induced
senescent tumor cell-derived extracellular vesicles promote
colorectal cancer progression through SERPINE1-mediated NF-ĸB p65
nuclear translocation. Mol Cancer. 23:702024. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang H, Wei Z, Xu C, Fang F, Wang Z, Zhong
Y and Wang X: Nuclear receptor 4A1 ameliorates UUO-induced renal
fibrosis by inhibiting the PI3K/AKT pathway. Sci Rep. 14:247872024.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lei L, Guo Q, Cheng Y, Gui Y, Li W, Zhao
Y, Xu Z, Luo Y, Wu G, Wang JZ, et al: Age-dependent elevation of
Nr4a1 attenuates PI3K/AKT/GSK3beta pathway and mediates tau
hyperphosphorylation and cognitive impairments. J Adv Res.
S2090-1232(25)00442-4. 2025.(Epub ahead of print).
|
|
29
|
Kirkland JL and Tchkonia T: Cellular
senescence: A translational perspective. EBioMedicine. 21:21–28.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Herranz N and Gil J: Mechanisms and
functions of cellular senescence. J Clin Invest. 128:1238–1246.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Safe S and Karki K: The paradoxical roles
of orphan nuclear receptor 4A (NR4A) in cancer. Mol Cancer Res.
19:180–191. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ma QX, Yin M and Lei QY: A nucleotide
regulates NR4A1 status in gastric cancer. Mol Cell. 85:4299–4300.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Faget DV, Ren Q and Stewart SA: Unmasking
senescence: Context-dependent effects of SASP in cancer. Nat Rev
Cancer. 19:439–453. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang C, Vegna S, Jin H, Benedict B,
Lieftink C, Ramirez C, de Oliveira RL, Morris B, Gadiot J, Wang W,
et al: Inducing and exploiting vulnerabilities for the treatment of
liver cancer. Nature. 574:268–272. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qing Y, Li H, Zhao Y, Hu P, Wang X, Yu X,
Zhu M, Wang H, Wang Z, Guo Q and Hui H: One-Two punch therapy for
the treatment of T-Cell malignancies involving p53-dependent
cellular senescence. Oxid Med Cell Longev. 2021:55295182021.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang T, Liu W, Shen Q, Tao R, Li C, Shen
Q, Lin Y, Huang Y, Yang L, Xie G, et al: Combination of PARP
inhibitor and CDK4/6 inhibitor modulates cGAS/STING-dependent
therapy-induced senescence and provides ‘one-two punch’ opportunity
with anti-PD-L1 therapy in colorectal cancer. Cancer Sci.
114:4184–4201. 2023. View Article : Google Scholar : PubMed/NCBI
|