Introduction
Pancreatic tumors are one of the deadliest
malignancies. In 2024, they were the third most deadly tumor
entity, despite ranking 10th in cancer incidence (1). A further increase in pancreatic
tumor-related deaths is expected in the coming years, so that it is
predicted to rank second within a few years (2). Of all pancreatic tumors, pancreatic
ductal adenocarcinoma (PDAC) is the most common and also the most
fatal (3). While life expectancy
for numerous other tumor diseases has increased significantly
thanks to improvements in therapy options, hardly any progress has
been made in this regard for PDAC, with an average 5-year survival
rate of 13% despite current standard therapy (4).
The use of high-dose vitamin C (ascorbic acid or
ascorbate), so-called pharmacological ascorbate, in tumor therapy
has been the subject of medical research for several decades,
starting with Linus Pauling's and Ewan Cameron's findings on
increased survival when treating tumor patients with high-dose
ascorbate (5,6). However, it was only numerous years
later that it became clear that the plasma ascorbate concentrations
required for antitumor efficacy can only be achieved by intravenous
administration (up to ~25 mM) (7),
but not by oral intake (up to ~0.2 mM) (8).
High-dose ascorbate has been investigated as a
monotherapy or combination therapy for various tumor entities in a
large number of studies. The majority of preclinical studies have
focused on the use of ascorbate in leukemia, colorectal carcinoma,
melanoma, and PDAC and have consistently shown that high-dose
ascorbate reduces the viability of tumor cells and decreases tumor
size in animals, as nicely summarized by Böttger et al
(9). With regard to pancreatic
cancer, an in vitro study by Kim et al (10) reported that high-dose ascorbate as
monotherapy reduces the proliferation and survival of PDAC cells by
inhibiting glucose metabolism. In addition, high-dose ascorbate
suppressed the invasion and metastasis of PDAC cells (10). The effects of high-dose ascorbate on
tumor cells are also reflected in studies carried out in
vivo. In mice with PDAC tumors, a reduction in tumor growth
(42%), metastasis, and ascites formation and also an improvement in
the survival rate were achieved with high-dose ascorbate (11). A recent randomized clinical trial
also provides evidence that supplementing standard therapy with
high-dose ascorbate prolongs the survival time of PDAC patients
(12).
Overall, the use of high-dose ascorbate is
considered safe and well tolerated, as has been observed in several
human studies (13–16). Chen et al (14) investigated the pharmacokinetics and
safety of intravenously administered vitamin C with increasing
doses in healthy individuals and cancer patients. The plasma
ascorbate concentration of >5 mM (equivalent to 88 mg/dl), which
is toxic for various cancer cell lines, could already be achieved
with infusions of 25 g vitamin C. No side effects were observed in
the participants in the present study, in which cardiac and liver
function as well as metabolic and coagulation parameters were
examined (14). In other studies,
however, mild side effects such as nausea, vomiting, headache, and
discomfort at the injection site were described. Severe side
effects can occur in patients with glucose-6-phosphate
dehydrogenase deficiency. This condition therefore represents a
contraindication for therapy with high-dose ascorbate (17).
Ascorbate has both antioxidant and pro-oxidant
properties. Which of these properties predominates depends, among
other things, on the local ascorbate concentration and the presence
of transition metals. At plasma concentrations in the micromolar
range, as achieved through diet or oral supplementation, ascorbate
has predominantly antioxidant functions, for example, as a radical
scavenger (18). However, if
pharmacological concentrations in the millimolar range are present,
as can be achieved by intravenous administration, ascorbate can
serve as a pro-drug for the formation of hydrogen peroxide
(H2O2), which is necessary for the
pro-oxidant effect (19,20). Catalytic metal ions play a decisive
role in ascorbate-induced H2O2 formation,
particularly free iron (so-called labile iron), which increases
H2O2 formation as part of the Fenton
reaction. Compared with healthy cells, cancer cells have larger
amounts of redox-active labile iron, so that high ascorbate
concentrations increase its pro-oxidative properties, which
explains the selective cytotoxicity of high-dose ascorbate towards
tumor cells (17,21). For this reason, an involvement of
the transport protein transferrin receptor 1 (TfR1), which mediates
cellular iron uptake in its trivalent form, in the antitumor
ascorbate effect is also suspected (22).
In recent years, there has been growing evidence of
increased cytotoxicity of high-dose ascorbate against tumor cells
in combination treatment with various iron compounds based on this
mechanism of action. While simultaneous treatment with iron and
ascorbate appears to be counterproductive in this context and
neutralizes the ascorbate effect, several studies show a successful
increase in treatment efficacy when high-dose ascorbate is used
after prior iron treatment (23–25).
Although in all three of these publications iron was used as a
preincubation prior to ascorbate treatment, different iron
compounds were used: While Piotrowsky et al (23) used 100 µM ferric chloride, Zhou
et al (24) used ferrous
ammonium sulphate (100 µM) as well as ammonium ferric citrate (100
µM), and Schoenfeld et al (25) used ferrous ammonium sulphate (100 µM
or 250 µM). There are also initial findings in PDAC cells, where
iron in the form of iron-hydroxyquinoline was used (26). However, the question of the success
of such a form of combination treatment with other iron compounds
in pancreatic carcinoma remains unanswered in order to investigate
the general validity of the recommendation for combination
treatment.
For this reason, the aim of the present study was to
investigate whether a combination treatment of iron and high-dose
ascorbate also leads to increased cytotoxicity against various
human pancreatic cancer cell lines. In addition, it should be
investigated whether the TfR1 expression in the tumor cells allows
conclusions to be drawn about the susceptibility to high-dose
ascorbate treatment.
Materials and methods
Cell culture and reagents
The human pancreatic carcinoma cell lines BxPC-3,
MIA PaCa-2, and PANC-1 were obtained from the German Collection of
Microorganisms and Cell Cultures (DSMZ). All cell lines were
cultured in RPMI-1640 medium (MilliporeSigma-Aldrich, cat. no.
R0883) with 10% fetal calf serum (FCS, MilliporeSigma-Aldrich, cat.
no. F7524) and 1% penicillin/streptomycin (MilliporeSigma-Aldrich,
cat. no. P4333) at 37°C and 5% CO2. HPDE6c7 cells were
obtained from MilliporeSigma-Aldrich (cat. no. SCC442) and cultured
in EpiVita Growth Medium for adult keratinocytes
(MilliporeSigma-Aldrich, cat. no. 141-500A) with 1%
penicillin/streptomycin, 50 µg/ml endothelial cell growth
supplement from bovine pituitary (MilliporeSigma-Aldrich, cat. no.
E0760) and 5 ng/ml epidermal growth factor (MilliporeSigma-Aldrich,
cat. no. SRP3027) at 37°C and 5% CO2. All cell lines
tested negative for mycoplasma infection.
Ascorbate (Pascorbin®) was purchased from
Pascoe pharmazeutische Praeparate GmbH (CAS: 50-81-7, cat. no.
00150343). Staurosporine (STS; CAS: 62996-74-1, cat. no. HY-15141)
and RAS-selective lethal 3 (RSL3; CAS: 1219810-16-8, cat. no.
HY-100218A) were obtained from MedChemExpress. Chloroquine (CQ) and
rapamycin (RAPA) were obtained from Enzo Biochem, Inc. (ENZ-51031).
Triton™ X-100, tert-butyl hydroperoxide (TBH), and
4-methylumbeliferyl heptanoate (MUH) were purchased from
MilliporeSigma-Aldrich (Triton™ X-100: CAS 9002-93-1, cat. no.
T8787; TBH: CAS 75-91-2, cat. no. 416665; MUH: CAS 18319-92-1, cat.
no. M2514). Deferoxamine mesylate (DFO) was obtained from Selleck
Chemicals LLC (cat. no. S5742).
Cell viability measurement
To determine cell viability, the MUH assay was
performed as previously described (23). The cells were seeded in 24-well
plates, according to their proliferation behavior, at a cell
density of 2×104 (PANC-1), 3×104 (MIA
PaCa-2), or 4×104 (BxPC-3) cells per well. Depending on
the planned experiments, FeCl3 was added to the medium
at a final concentration of 100 µM during seeding. 24 h after
seeding, the medium was discarded, the cells were washed twice with
PBS and treated with the indicated ascorbate concentrations for a
further 24 h. Triton™ X 100 served as a positive control.
To perform the MUH assay, the medium was discarded,
the cells were washed once with PBS and the MUH reagent was added
in phenol red-free, FCS-free RPMI-1640 medium (100 µg/ml). After an
1-h incubation period at 37°C, the fluorescence intensity at 460 nm
was measured which is proportional to the number of living cells
using the Synergy™ H1 microplate reader (BioTek Instruments, Inc.).
From this, the relative percentage of living cells in relation to
the untreated control was calculated.
Immunoblotting
A total of 1×105 (PANC-1),
1.5×105 (MIA PaCa-2), or 2×105 (BxPC-3) cells
were seeded in 6-well format and incubated for 24 h. To determine
basal protein expression, cell lysis was performed afterwards as
described below. For the experiments with iron, FeCl3
was added at different concentrations during seeding and
subsequently cell lysates were generated. Afterwards, the cells
were either lysed as described later (to investigate the influence
of iron on protein expression) or, to investigate the influence of
ascorbate treatment on protein expression, the medium was changed
24 h after seeding and the cells were treated with different
ascorbate concentrations for 6 h. A total of 10 µM STS served as a
positive control for apoptosis, 5 µM RSL3 as a positive control for
ferroptosis, and 60 µM CQ with 500 nM RAPA as a positive control
for autophagy. For the untreated control, the medium was also
changed, but without any added treatment. After treatment, the
cells were washed twice with PBS and lysed on ice by adding 50 µl
of lysis buffer [150 mM NaCl, 50 mM Tris-HCl, pH 7.4, 1% Triton™
X-100, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS),
10 µl/ml aprotinin, 10 µl/ml leupeptin, 10 µl/ml pepstatin A, 1.5
µl/ml PMSF, and 1 µl/ml Na3VO4]. After
incubation on ice for 30 min, cell debris was removed from the
lysates by centrifugation at 16,200 × g at 4°C for 20 min and
transferring the supernatant to a new reaction tube. The lysates
were stored at −20°C until further use. The protein concentration
of the lysates was determined using the Lowry method. From each
lysate, 35 µg protein was mixed with Laemmli buffer (250 nM
Tris-HCl pH 6.8, 40% glycerol, 8% SDS, 20% 2-mercaptoethanol, crumb
bromophenol blue) and boiled at 95°C for 5 min. The proteins were
then separated by electrophoresis in a 10 or 15% SDS polyacrylamide
gel. The proteins were subsequently transferred to a PVDF membrane,
blocked for 1 h at RT in 5% milk powder in Tris-buffered saline
containing 0.1% Tween-20 (TBST) and incubated overnight at 4°C with
the following primary antibodies [diluted at 1:1,000 in 5% bovine
serum albumin (BSA, cat. no. 8076.1, Carl Roth GmbH & Co.)]:
anti-Cleaved Caspase-3 (cat. no. ab2302; Abcam), anti-Glutathione
peroxidase 4 (GPX4; cat. no. 52455), anti-TfR1 (cat. no. 13113)
anti-GAPDH (cat. no. 2118) or anti-LC3B (cat. no. 2775; all from
Cell Signaling Technology, Inc.). The next day, incubation with the
secondary antibody [goat anti-rabbit IgG, horseradish peroxidase
(HRP)-coupled: cat. no. 7074s; 1:10,000 in 5% milk powder in TBST;
Cell Signaling Technology Inc.] in 5% milk powder solution in TBST
was carried out at RT for 45 min. After washing three times with
TBST, the proteins were analyzed by adding the WesternBright
chemiluminescence substrate Sirius (Biozym Scientific GmbH) in the
Fusion FX chemiluminescence detector (Vilber). The densitometric
evaluation was performed using ImageJ version 1.54r. (National
Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
The cell lines were seeded in the previously
mentioned cell numbers in 6-well format and incubated for 24 h at
37°C and 5% CO2. In the case of iron treatment, 100 µM
FeCl3 was added to the medium at seeding. This was
followed by RNA isolation using the Quick-RNA™ Miniprep Kit (Zymo
Research Corp.) according to the manufacturer's protocol.
Subsequently, 1 µg of RNA per sample was used for cDNA synthesis
using the iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Inc.)
according to the manufacturer's instructions. The SYBR®
Green RT-PCR Reaction Mix (Bio-Rad Laboratories, Inc.) and the
following primer pairs were used for qPCR: TfR1 forward,
5′-ATTGAACCTGGACTATGAGAG-3′ and reverse, 5′-TGGAAGTAGCACGGAAGA-3′;
GAPDH forward, 5′-GGTCGGAGTCAACGGATTTG-3′ and reverse,
5′-GGAAGATGGTGATGGGATTTC-3′. The qPCR program in the CFX96 Touch
Real-Time PCR Detection System (Bio-Rad Laboratories, Inc.) was
composed as follows: 2 min at 95°C and 40 cycles with 10 sec at
95°C, 30 sec at 58°C, and 30 sec at 72°C. The 2−ΔΔCq
method was used to calculate the relative gene expression of TfR1
as described by Livak and Schmittgen (27).
Fluorescence microscopy
The cells were seeded in the aforementioned cell
numbers in 24-well format on cover slips. After 24 h of treatment,
the medium was changed and the cells were treated with different
ascorbate concentrations for 6 h. Treatment with 5 µM STS
(apoptosis) or 1 µM RSL3 (ferroptosis) served as a positive
control. After treatment, the cells were washed once with PBS and
fixed by adding 4% PFA in PBS for 20 min. They were washed again
with PBS and quenched with 150 mM glycine in PBS for 10 min. After
a further washing step, phalloidin staining was performed. For
this, the cells were stained with phalloidin tetramethylrhodamine B
isothiocyanate (Phalloidin-TRITC, cat. no. P1951, 10 µg/ml in PBS;
MilliporeSigma-Aldrich) for 1.5 h at RT. The cells were washed once
with PBS and stained with diamidino-2-phenylindole (DAPI, cat. no.
6335.1, Carl Roth GmbH & Co.) solution (1:1,000 in PBS) for 20
min in the dark. After washing again with PBS and double-distilled
water, the cells were mounted on slides with mounting medium
(Fluoromount-G®, cat. no. 0100-01, SouthernBiotech).
Fluorescence images were captured with a Lionheart FX fluorescence
microscope (BioTek Instruments, Inc.).
Cell cycle analysis
Cell cycle analysis was performed to detect
apoptosis as previously described (23). The cells were seeded in the
aforementioned cell numbers per cell line in 6-well format and
incubated for 24 h. Subsequently, the medium was changed, and the
cells were treated with different ascorbate concentrations for 24
h. Treatment with 1 µM or 20 µM STS for 20 h served as a positive
control. After treatment, the cells were harvested by
trypsinization, washed once with PBS, and fixed by adding 70%
ice-cold ethanol at 4°C overnight. The cells were then washed twice
with PBS and incubated in PBS with 50 µg/ml propidium iodide (PI,
cat. no. P4170, Sigma-Aldrich; Merck KGaA) and 100 µg/ml RNAse
(cat. no. 10109142001, Roche Diagnostics) at 4°C for 20 min. This
was followed by flow cytometric analysis of the cell cycle phases
(NovoCyte® 2060R flow cytometer, Agilent Technologies,).
A total of 10,000 events per sample were recorded and three
independent experiments were performed.
Intracellular reactive oxygen species
(ROS) measurement
To analyze intracellular (ROS) levels, the
dichlorodihydrofluorescein diacetat (DCFH-DA) assay was performed
as previously described (28).
Cells were seeded in the aforementioned cell numbers in 24-well
format with or without the addition of 100 µM FeCl3. A
total of 24 h after seeding, cells were washed twice with PBS and
treated with different ascorbate concentrations or 500 µM TBH as a
positive control for 6 h. After treatment, the cells were harvested
by trypsinization and the cell pellet was washed twice with PBS.
This was followed by incubation with 5 µM DCFH-DA (cat. no. D6883,
MilliporeSigma-Aldrich) in phenol red-free, FCS-free RPMI-1640
medium at 37°C in the dark for 30 min. The cells were then pelleted
again and washed once with PBS, resuspended in phenol red-free,
FCS-free RPMI, and analyzed by flow cytometry (NovoCyte®
2060R flow cytometer; Agilent Technologies, Inc.). With absorbance
at 488 nm and detection of emission at 530 nm, 10,000 events per
sample were recorded and three independent experiments were
performed.
Quantification of the labile iron
pool
To measure the intracellular labile iron pool (LIP),
the calcein acetoxymethyl ester (AM) assay was performed as
described by Schönfeld et al (25). The cells were seeded in the
aforementioned cell numbers in 24-well format. During seeding,
FeCl3 was added to the cells at different
concentrations. A total of 24 h after seeding, the cells were
harvested by trypsinization, washed twice with PBS, and incubated
after addition of PBS with 500 nM calcein AM (cat. no. C1359,
MedChemExpress) for 15 min at 37°C in the dark. The cells were
pelleted, washed once more with PBS, and resuspended in PBS. The
cell suspension was divided into two flow cytometry tubes and
2′,2′-bipyridyl (BIP) was added to one of these tubes per treatment
at a final concentration of 100 µM. After a 15 min incubation at RT
in the dark, the cells were analyzed by flow cytometry
(NovoCyte® 2060R flow cytometer; Agilent Technologies,
Inc.) with absorbance at 488 nm and emission measurement at 515 nm.
The relative LIP was determined by subtracting the mean
fluorescence intensity without BIP addition from the mean
fluorescence intensity with BIP and normalizing the result to the
untreated control. For each sample, 10,000 events were recorded and
three independent experiments were performed.
Statistical analysis
The statistical data analysis and the calculation of
IC50 values were performed with GraphPad Prism version
10.4.1 (GraphPad Software, Inc.; Dotmatics). For multiple group
comparisons, one-way ANOVA with subsequent Dunnett's multiple
comparisons test was used for P-value calculation and significance
determination. P≤0.05 was considered to indicate a statistically
significant difference.
Results
Effect of high-dose ascorbate alone
and in combination with iron on the viability of pancreatic cancer
cells
The MUH assay was performed to investigate the
cytotoxic effect of high-dose ascorbate and its combination
treatment with ferric chloride on human pancreatic cancer cells.
While no cytotoxicity was observed after 24 h of treatment at the
concentration of 0.2 mM (achievable in plasma by oral
supplementation), treatment with pharmacological ascorbate
concentrations (only achievable by intravenous administration) led
to a rapid decrease in relative cell viability in all cell lines
investigated (Fig. 1A). While the
BxPC-3 cells proved to be the most resistant cell line with a
significant reduction of the number of viable cells from 0.8 mM
ascorbate and an IC50 of 0.89 mM, a significant
cytotoxic effect on the MIA PaCa-2 and PANC-1 cells occurred from
0.4 mM onwards, with IC50 values of 0.43 mM (MIA PaCa-2)
and 0.45 mM (PANC-1), respectively. In all three cell lines
analyzed, the relative cell viability dropped to below 20% at 2 mM
ascorbate at the latest.
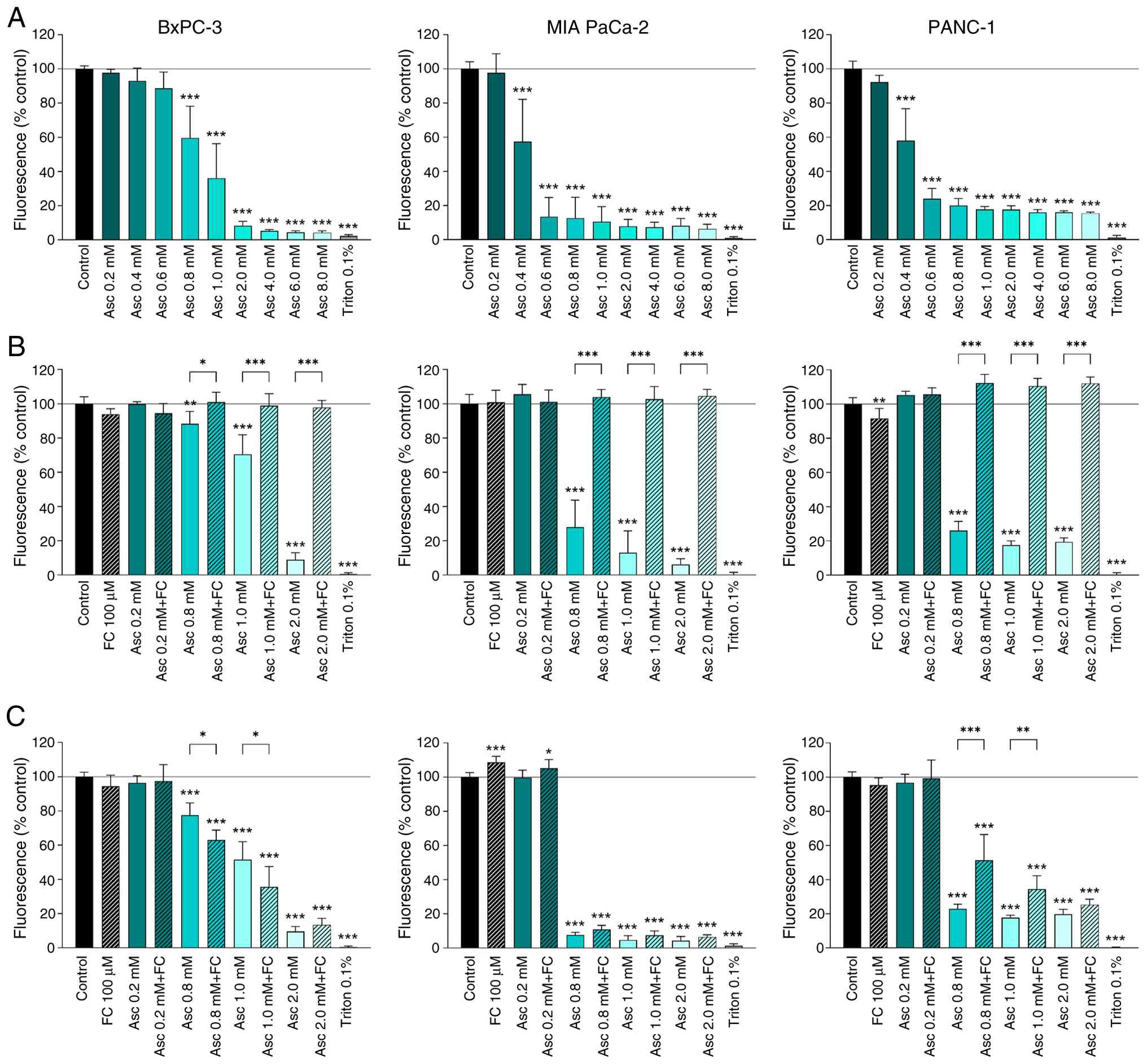 | Figure 1.Influence of high-dose ascorbate
alone or in combination with ferric iron on the viability of human
pancreatic cancer cell lines. The cell lines BxPC-3, MIA PaCa-2 and
PANC-1 were either treated for 24 h with the indicated ascorbate
concentrations alone (A), in the form of coincubation
simultaneously with ascorbate and 100 µM FC (B), or incubated with
100 µM FC for 24 h as a preincubation before ascorbate treatment
(C). Triton™ X-100 at 0.1% (v/v) served as positive control. Cell
viability was measured after treatment by MUH assay. The results
are presented as a percentage of fluorescence intensity relative to
the untreated control. Three independent experiments were performed
in duplicates. Error bars represent the mean ± SD, statistical
analysis with one-way ANOVA and subsequent Dunnett's multiple
comparisons test, confidence interval 95%. *P≤0.05, **P≤0.01, and
***P≤0.001. Asc, ascorbate; FC, ferric chloride; MUH,
4-methylumbelliferyl heptanoate. |
Two different approaches were investigated for the
combination treatment of ascorbate and iron. In the co-incubation
with FeCl3, the treatment was carried out simultaneously
with both substances (Fig. 1B). In
this case, the combination treatment led to a complete inhibition
of ascorbate-induced cytotoxicity. This was true for all cell lines
analyzed as well as for all concentrations used.
In the preincubation with iron, the 24 h ascorbate
treatment followed a previous treatment for 24 h with 100 µM
FeCl3 (Fig. 1C). In this
case, the influence of the combination treatment varied depending
on the cell line. In BxPC-3 cells, the combination treatment with
0.8 and 1 mM ascorbate led to a significantly greater decrease in
cell viability than ascorbate treatment alone (0.8 mM: 78% vs. 63%,
1 mM: 52% vs. 36%). In the MIA PaCa-2 cells, none of the analyzed
concentrations differed significantly between combination treatment
and ascorbate treatment alone. On the other hand, the PANC-1 cells
showed an inhibition of the ascorbate effect in the concentrations
0.8 and 1 mM by combination treatment with preincubated
FeCl3 (0.8 mM: 23% vs. 51%, 1 mM: 18% vs. 35%).
To increase the transferability of the results, the
experiments were performed in the form of preincubation with holo
transferrin instead of FeCl3 for all three cell lines
(Fig. S1). Consistent with the
data on preincubation with FeCl3, treatment with holo
transferrin only led to a slight increase in efficacy when BxPC-3
cells were treated with 2 mM ascorbate, while no effect on
treatment efficacy was observed in MIA PaCa-2 and PANC-1 cells.
Investigation of apoptosis as a
possible ascorbate-induced cell death mechanism
To detect possible apoptosis as an ascorbate-induced
cell death mechanism, caspase-3 cleavage after treatment was
analyzed by flow cytometry (Fig.
2A) and western blotting (Fig.
2B). Neither method was able to detect any activation of
caspase 3 after 6 h of treatment for any of the ascorbate
concentrations or cell lines analyzed. This also applied to
ascorbate concentrations that led to a decrease in cell viability
to below 20% in the MUH assay.
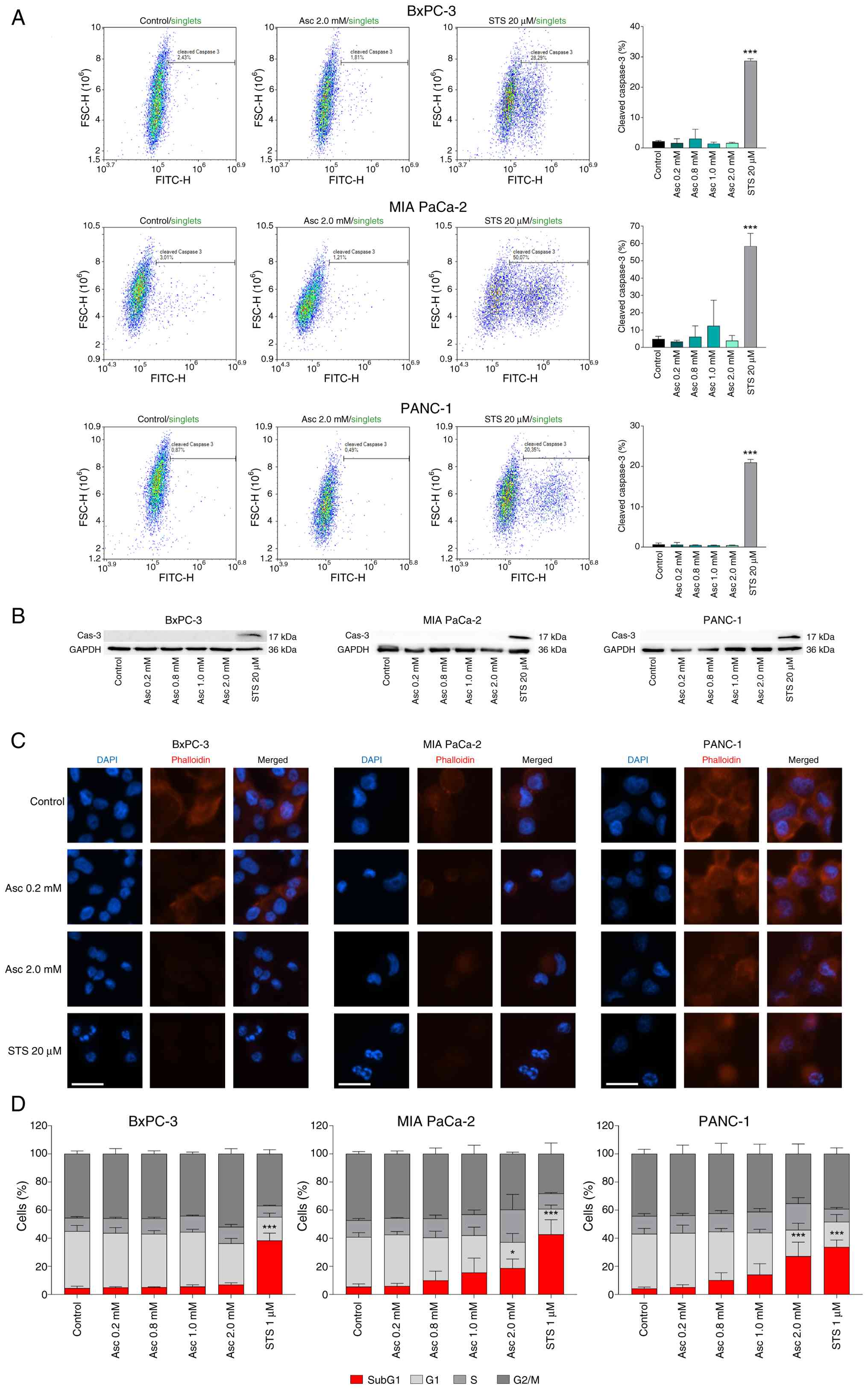 | Figure 2.Investigation of the effect of
high-dose ascorbate treatment on different apoptosis markers in
human pancreatic cancer cell lines. A possible induction of
apoptosis in the human pancreatic cancer cell lines BxPC-3, MIA
PaCa-2, and PANC-1 was investigated by testing caspase-3 cleavage
by flow cytometry (A) and western blotting (B). Cells were treated
for 6 h with the indicated ascorbate concentrations. 20 µM STS
served as positive control. Three independent experiments were
performed. Morphological nuclear changes were detected by
fluorescence microscopy. The white scale bars represent 25 µm. (C).
Cells were treated with the indicated ascorbate concentrations for
6 h and then fixed. 10 µM STS served as positive control. The
nuclei were stained with DAPI (blue), the cytoskeleton with
phalloidin (red). A representative experiment is shown. Cell cycle
analysis was performed by flow cytometry (D). Cells were treated
with the indicated ascorbate concentrations for 24 h, fixed,
stained with PI, and subsequently detected. Treatment with 1 µM STS
for 20 h served as positive control. Three independent experiments
were performed. Error bars represent the mean ± SD, statistical
analysis with one-way ANOVA and subsequent Dunnett's multiple
comparisons test, confidence interval 95%. *P≤0.05 and ***P≤0.001.
Asc, ascorbate; DAPI, diamidino-2-phenylindole; PI, propidium
iodide; STS, staurosporine. |
As a further indication of possible apoptotic
ascorbate-mediated cell death, apoptosis-typical nuclear
fragmentation was examined using fluorescence staining (Fig. 2C). While ascorbate treatment visibly
reduced the phalloidin staining of the filamentous actin, no
apoptotic cell nuclear fragmentation could be detected in any of
the cell lines examined due to the highly cytotoxic ascorbate
concentration of 2.0 mM and thus no evidence of apoptotic cell
death was found here either.
To confirm these results, a flow cytometric cell
cycle analysis was performed by PI staining after ascorbate
treatment (Fig. 2D). In agreement
with the previous results, no increase in the SubG1 fraction as an
indication of apoptosis-typical DNA fragmentation was observed in
the BxPC-3 cell line. In the MIA PaCa-2 and PANC-1 cells, a
significant increase in the SubG1 fraction was evident at the
highest ascorbate concentration (2 mM) analyzed. However, the lower
ascorbate concentrations of up to 1 mM, which also caused a
decrease in cell viability of at least 80%, similarly provided no
evidence of apoptotic cell death.
Evaluation of ferroptosis as an
ascorbate-induced cell death mechanism
Based on the hypothesized main mechanism of action
of high-dose ascorbate in cancer cells, ferroptotic cell death is
often suspected in this context (23,29).
For this reason, ferroptotic cell death following ascorbate
treatment was evaluated by investigating the effect of the
ferroptosis inhibitor DFO (Fig.
3A). In BxPC-3 and PANC-1 cells, the addition of DFO to the
treatment at all effective ascorbate concentrations led to a
significant, in some cases complete suppression of the ascorbate
effect, which was reflected in a lower decrease in cell viability
in the MUH assay. In the MIA PaCa-2 cells, DFO also had a strong
inhibitory effect at ascorbate concentrations of 0.8 and 1 mM. At 2
mM, however, ascorbate cytotoxicity predominated and DFO only led
to a slight, non-significant inhibition of the treatment
effect.
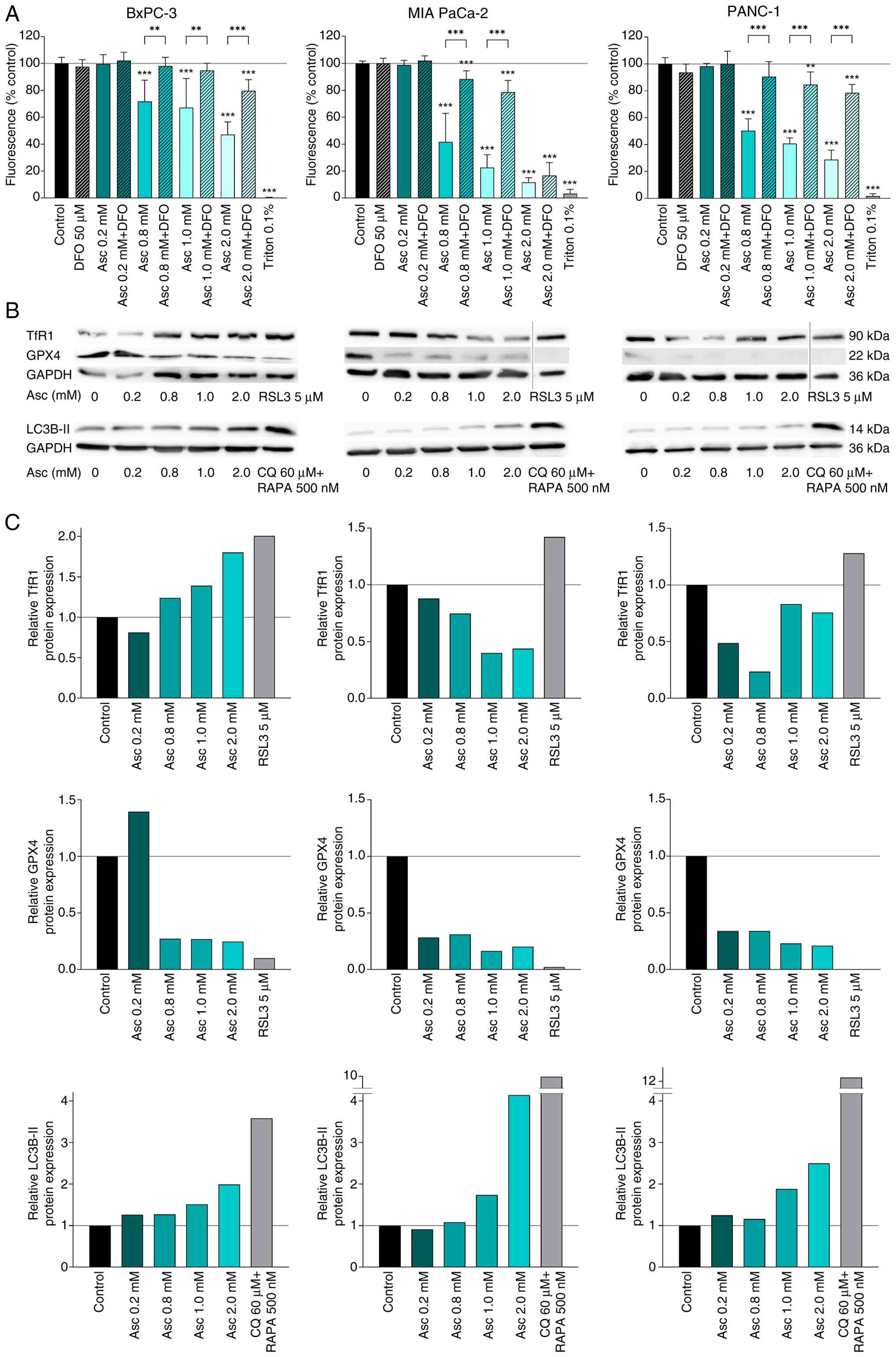 | Figure 3.Investigation of the effect of
high-dose ascorbate treatment on different ferroptosis markers in
human pancreatic cancer cell lines. The human pancreatic cancer
cell lines BxPC-3, MIA PaCa-2, and PANC-1 were treated with or
without the ferroptosis inhibitor DFO for 24 h, after which cell
viability was measured using the MUH assay (A). Triton™ X-100 at
0.1% (v/v) served as positive control. The results are presented as
a percentage of the fluorescence intensity of the untreated
control. Three independent experiments were performed in
duplicates. As further signs of ferroptosis, protein expression of
TfR1, GPX4, and LC3B-II was detected by western blotting (B) and
quantified densitometrically (C). Cells were treated with the
indicated ascorbate concentrations for 6 h, after which a western
blot was performed. 5 µM RSL3 or 60 µM CQ together with 500 nM RAPA
were used as positive controls. Signal intensity was analyzed
densitometrically and normalized to GAPDH. One representative
experiment out of two is shown. Error bars represent the mean ± SD,
statistical analysis with one-way ANOVA and subsequent Dunnett's
multiple comparisons test, confidence interval 95%. **P≤0.01 and
***P≤0.001. Asc, ascorbate; CQ, chloroquine; DFO, deferoxamine
mesylate; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; GPX4,
glutathione peroxidase 4; MUH, 4-methylumbelliferyl heptanoate;
RAPA, rapamycin; RSL3, RAS-selective lethal 3; TfR1, transferrin
receptor 1. |
Further indications of ferroptosis are increased
TfR1 expression and reduced expression of the antioxidant active
enzyme GPX4 (30). Therefore, both
parameters were also analyzed by western blotting (Fig. 3B). In addition, the expression of
both proteins was analyzed densitometrically (Fig. 3C). In order to detect autophagy in
the tumor cells, which is another feature of ferroptotic cell death
(30), LC3B-II expression was also
examined after ascorbate treatment.
While increased TfR1 expression was observed in the
BxPC-3 cells starting at 0.8 mM 6 h ascorbate treatment, the MIA
PaCa-2 and PANC-1 cells showed reduced expression as a result of
treatment. GPX4 expression was significantly reduced in all three
cell lines after ascorbate treatment, in the MIA PaCa-2 cells and
PANC-1 cells even at the lowest ascorbate concentration of 0.2 mM,
strongly indicating ferroptotic cell death. In agreement with these
results, a clear induction of autophagy was observed in all three
cell lines, as indicated by increased LC3B-II expression. Ascorbate
treatment in the MIA PaCa-2 cells led to the strongest increase in
LC3B-II expression, followed by the PANC-1 and finally the BxPC-3
cells, reflecting the sensitivity of the different cell lines to
high-dose ascorbate.
Influence of iron on ROS induction of
high-dose ascorbate and TfR1 expression
Based on the observed signs of ferroptotic cell
death after ascorbate treatment with simultaneous reduction of the
ascorbate effect by combination with iron, the influence of 24 h
iron treatment on ascorbate-induced ROS formation (DCFH-DA assay)
as well as intracellular LIP (calcein AM assay) and TfR1 expression
(western blotting and qPCR) was investigated (Fig. 4). While ascorbate treatment alone
led to a concentration-dependent increase in intracellular ROS
levels in all cell lines, co-incubation with 100 µM ferric chloride
simultaneously with ascorbate treatment resulted in a significant
and almost complete inhibition of intracellular ROS formation in
the pancreatic cancer cell lines (Fig.
4A).
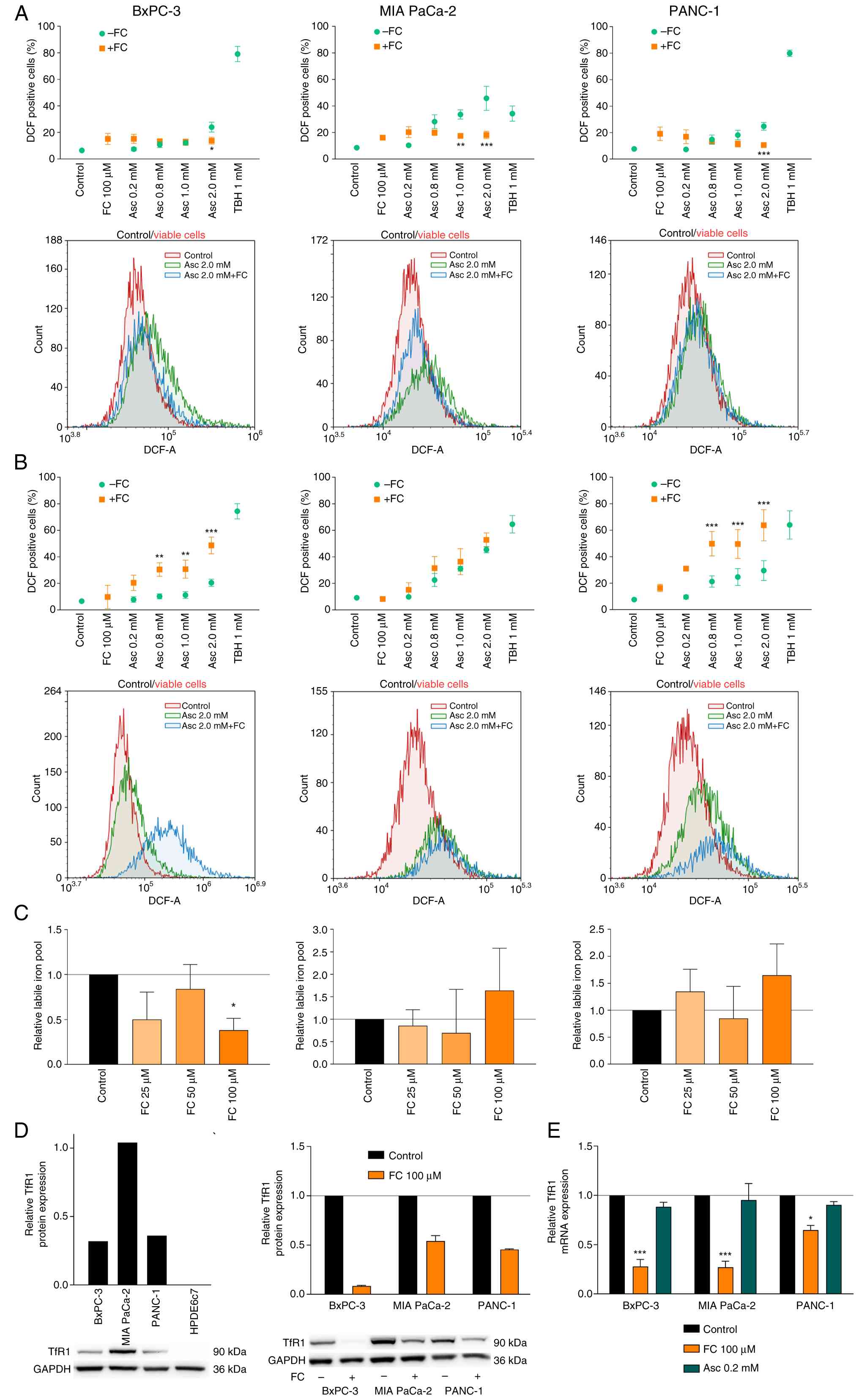 | Figure 4.The influence of iron and TfR1 on the
ascorbate effect in human pancreatic cancer cell lines. To
investigate the effect of iron on ascorbate action, the three human
pancreatic cancer cell lines BxPC-3, MIA PaCa-2, and PANC-1 were
either incubated simultaneously for 6 h with ascorbate at different
concentrations and 100 µM ferric chloride (FC) (A) or treated with
ascorbate after 24 h preincubation with ferric chloride (B). The
intracellular ROS levels were determined after treatment by flow
cytometry using the DCFH-DA assay. The percentage of DCF-positive
cells is shown as a measure of intracellular ROS accumulation.
Statistically significant differences between the combination
treatment with iron and ascorbate treatment alone are marked by
asterisks. Three independent experiments were performed. The
intracellular LIP after 24-h incubation with ferric chloride was
determined by flow cytometry using the calcein AM assay (C). The
relative labile iron pool is shown in relation to the untreated
control. Three independent experiments were performed. The basal
protein expression of TfR1 in the pancreatic cancer cell lines and
the non-malignant pancreatic ductal epithelial cell line HPDE6c7
was determined by western blotting, as well as the influence of
ferric chloride on TfR1 expression in the three pancreatic cancer
cell lines (D). The western blot results were also analyzed
densitometrically. A representative experiment is shown for basal
expression. For TfR1 expression after iron treatment, two
independent experiments were performed, a representative western
blot is shown. The influence of the 24-h ferric chloride treatment
as well as ascorbate treatment was additionally determined at the
mRNA level by qPCR (E). Error bars represent the mean ± SD,
statistical analysis with one-way ANOVA and subsequent Dunnett's
multiple comparisons test, confidence interval 95%. *P≤0.05,
**P≤0.01, and ***P≤0.001. Asc, ascorbate; DCFH-DA,
dichlorodihydrofluorescein diacetate; DCF, dichlorofluorescein; FC,
ferric chloride; LIP, labile iron pool; ROS, reactive oxygen
species; TfR1, transferrin receptor 1. |
By contrast, preincubation with 100 µM ferric
chloride for 24 h and subsequent ascorbate treatment showed a
significant increase in intracellular ROS levels in BxPC-3 and
PANC-1 cells from 0.8 mM. However, in MIA PaCa-2 cells, no
difference in ROS induction was observed between ascorbate
treatment alone and ascorbate treatment after prior iron
preincubation (Fig. 4B).
The influence of a 24 h incubation with ferric
chloride on the LIP of pancreatic cancer cells was investigated
using the calcein AM assay (Fig.
4C). In BxPC-3 cells, all concentrations analyzed resulted in a
reduction of the LIP that reached significance at 100 µM. The MIA
PaCa-2 and PANC-1 cells, on the other hand, showed no significant
change in the LIP due to iron treatment.
The basal expression of TfR1 in the three pancreatic
cancer cell lines and in the non-malignant human pancreatic ductal
epithelial cell line HPDE6c7 was determined by western blotting and
quantified densitometrically (Fig.
4D). Interestingly, HPDE6c7 cells showed almost no detectable
TfR1 protein expression, whereas the most ascorbate-sensitive cell
line MIA PaCa-2 showed the highest expression, followed by BxPC-3
and PANC-1 cells. Furthermore, the influence of 24 h iron treatment
on TfR1 expression was analyzed by western blotting and qPCR to
find a possible explanation for the observations relating the LIP
(Fig. 4E). Ferric chloride
treatment led to a significant reduction in TfR1 expression, with
an almost complete inhibition of protein expression being observed
in the BxPC-3 cells both at the mRNA and protein level. By
contrast, treatment with the highest orally achievable ascorbate
concentration had no effect on TfR1 mRNA expression in BxPC-3 and
PANC-1 cells. Gene expression increased slightly in MIA PaCa-2
cells, but this increase was also not significant.
Discussion
To the best of our knowledge, the present study is
the first to investigate the antitumor efficacy of a combination
treatment of high-dose ascorbate and iron in pancreatic cancer
cells. Despite the general assumption that the effect of ascorbate
against tumor cells is mainly based on ROS induction involving
intracellular labile iron (9), the
combination treatment in the present model did not lead to an
increase in cytotoxicity, on the contrary even to a decrease in
efficacy in most cases. While simultaneous treatment with ascorbate
and iron also led to a reduction in cytotoxicity in other tumor
entities in previous studies, the treatment regimen of
preincubation often resulted in increased potency (23,31,24).
In glioblastoma, for example, increased ascorbate-induced
cytotoxicity was shown in vitro after prior incubation of
the cells with FeCl3 (23). The same has been shown in
vitro with colon carcinoma cells and in osteosarcoma (31,24).
However, in a recent study on prostate cancer cells,
Deme et al (32) also showed
a reduction in the effect of ascorbate after preincubation of the
cells with FeCl3 for 24 h prior to ascorbate treatment.
These partly contradictory data suggest that the benefit of iron
may be tumor entity-specific. The mutation of the KRAS gene could
play a role here. For example, the cell lines mentioned, in which
preincubation with iron led to an increase in the ascorbate effect,
predominantly carry the KRAS wild-type gene. In the present study,
too, the only cell line in which at least a slight increase in
ascorbate cytotoxicity could be achieved was a KRAS wild-type cell
line. By contrast, the cell lines in which preincubation with
FeCl3 had the opposite effect were KRAS-mutated cells.
On the other hand, the increased overall sensitivity of
KRAS-mutated cancer cells is described in the clinical phase III
study by Wang et al (33) on
the use of high-dose ascorbate in metastatic colorectal cancer.
While the addition of high-dose ascorbate to the treatment regimen
FOLFOX (leucovorin, 5-fluorouracil, and oxaliplatin) and
bevacizumab showed no benefit on progression-free survival,
objective response rate, and overall survival, a significant effect
of the additional high-dose ascorbate administration was found on
progression-free survival in the subpopulation of patients with
KRAS-mutated metastatic colorectal cancer. However, these effects
were observed without targeted iron supplementation.
It appears that iron supplementation is beneficial
in patients without KRAS mutation, but detrimental in patients with
KRAS mutation. If this is confirmed in further studies and/or in
other tumor entities, this would mean that before using high-dose
ascorbate in tumor patients, the KRAS status should be analyzed in
order to identify a possible contraindication for the additional
administration of iron or to increase the effectiveness of the
therapy through a rational combination treatment.
It is surprising, however, that increased
intracellular ROS formation was observed for the PANC-1 cells after
6 h of ascorbate treatment and prior preincubation with iron, with
simultaneously reduced cytotoxicity of this form of combination
treatment. However, since ascorbate treatment alone caused a
low-threshold ROS induction in this cell line, it can be assumed
that the cytotoxicity here is not solely due to ROS formation, but
that other mechanisms also play a role, as summarized by Böttger
et al (9).
The exact cell death mechanism by which high-dose
ascorbate causes the death of tumor cells has not yet been
conclusively clarified. Previous studies suggest that the form of
cell death may vary between different tumor entities and depending
on the concentration used (29).
However, as summarized by Szarka et al (29), apoptotic cell death can be ruled out
in most cases, which is presumably related to the fact that
numerous tumor cells lose or greatly reduce their ability to
undergo classical apoptosis during transformation (34). The present study also found minor
indications of apoptosis after ascorbate treatment of human
pancreatic cancer cells. In particular, caspase-3 activation, which
is crucial for apoptosis, could not be detected in any of the cell
lines analyzed using two different detection methods.
The current understanding of the antitumor mechanism
of action of high-dose ascorbate as well as the data of this study
suggest a key involvement of intracellular labile iron and the
promotion of intracellular oxidative stress (35). Both factors are also crucial
hallmarks of ferroptosis, which is why this form of cell death is
discussed in connection with ascorbate (29). Nevertheless, signs of ferroptosis
after ascorbate treatment have so far only been detected in
oropharyngeal cancer and anaplastic thyroid cancer (36,37).
In pancreatic carcinoma, there have been no studies on ferroptotic
cell death after vitamin C treatment, but only indications that
ascorbate can increase erastin-induced ferroptosis (38). In the present study, it could now be
shown for the first time, to the best of our knowledge, that
high-dose ascorbate triggers cell death in human pancreatic cancer
cells, causing major signs of ferroptosis. Interestingly,
ferroptosis has increasingly become the focus of cancer therapy
research in recent years, as various drugs that induce ferroptosis
appear to be a promising option for overcoming drug resistance,
especially in therapy-resistant tumors (39). High-dose ascorbate could therefore
represent an effective alternative here, especially for second-line
therapy.
Another important finding of the present study is
the role of TfR1 in the application of high-dose ascorbate alone
and in combination with iron. Interestingly, the cell line MIA
PaCa-2, which was the most sensitive to vitamin C, shows by far the
highest TfR1 expression, which could explain the high
susceptibility of these cells: Namely, increased intracellular LIP
would also increase intracellular ROS formation, as expected.
In line with this, treatment of the cells with
FeCl3 uniformly led to a significant, in some cases
almost complete, decrease in TfR1 expression. As a result, the
intracellular LIP of the cells could not be further increased or
even decreased, which explains the partially reduced or unchanged
ascorbate effect when combined with FeCl3. The current
understanding of the mechanism of action of high-dose ascorbate on
PDAC cells, including the findings of the present study, is
summarized in Fig. 5.
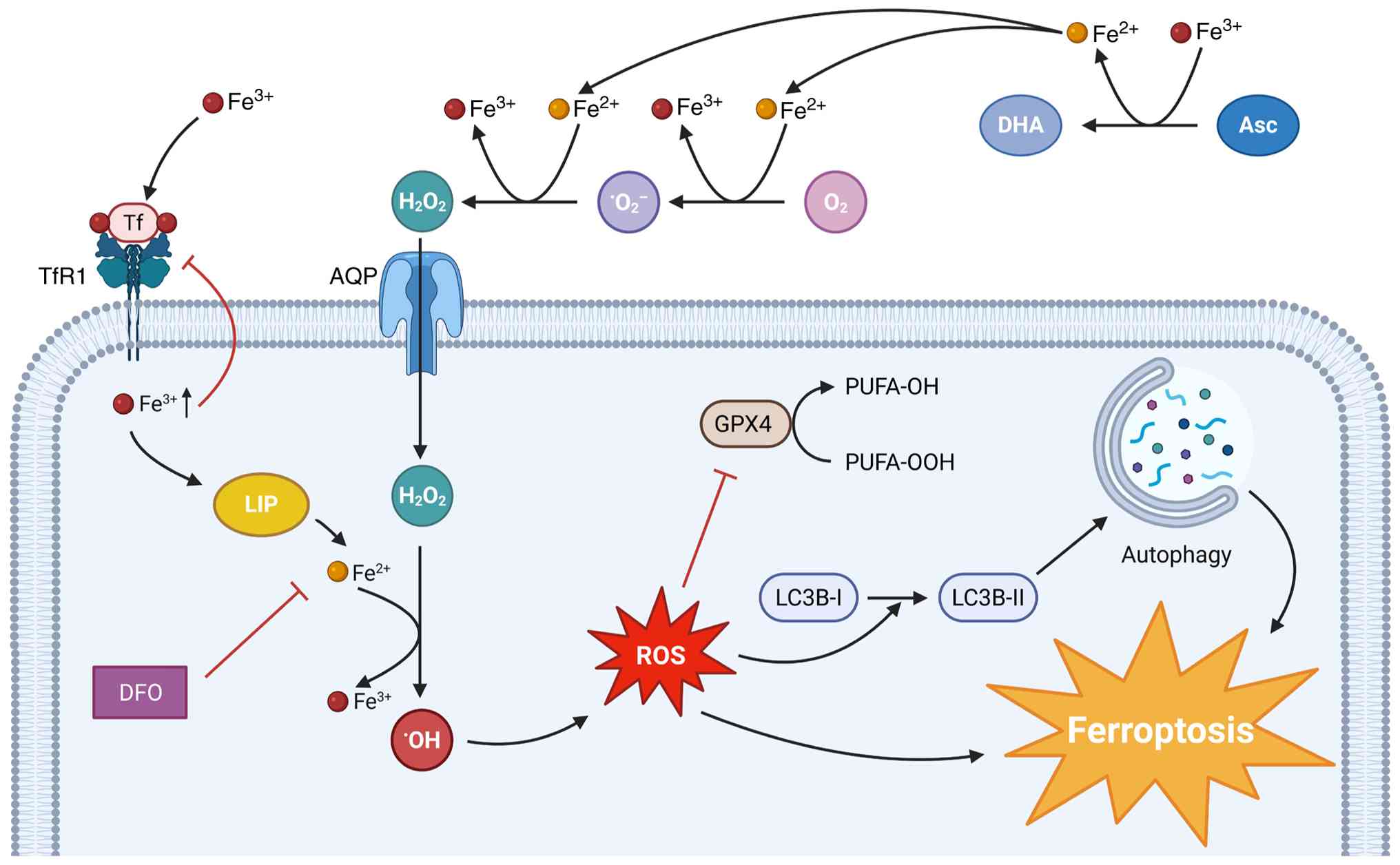 | Figure 5.Hypothesized mechanism of
ascorbate-induced cell death in pancreatic cancer cells. Asc is
being oxidized outside the cell to DHA by ferric iron. This results
in the accumulation of ferrous iron, which reacts with molecular
oxygen and then with the superoxide radical anion to form hydrogen
peroxide. Hydrogen peroxide enters the cancer cell via AQPs. Ferric
iron itself enters the cell via TfR1, a process that is inhibited
by intracellular iron accumulation and increased LIP. Once inside
the cancer cell, hydrogen peroxide reacts with ferrous iron to form
the hydroxyl radical, causing oxidative stress through ROS
accumulation. This inhibits the antioxidant activity of GPX4,
promotes autophagy, and finally induces ferroptotic cell death.
Ferroptosis is inhibited by DFO. Asc, ascorbate; AQP, aquaporin;
DFO, deferoxamine mesylate; DHA dehydroascorbic acid; GPX4,
glutathione peroxidase 4; LIP, labile iron pool; PUFA,
polyunsaturated fatty acids; ROS, reactive oxygen species; TfR1,
transferrin receptor 1. |
When trying to transfer this in vitro data
onto the much more complex environment of a living organism, there
are some difficulties that should be taken into consideration. One
relevant point is that, in a clinical context, intravenous iron is
usually administered in the form of iron-carbohydrate complexes,
such as iron carboxymaltose, rather than the iron compounds used in
most cell culture studies. These complexes are mainly taken up by
macrophages in the liver, spleen, and bone marrow, and are
continuously and in a controlled way delivered to transferrin,
which distributes them throughout the body and to potential tumor
tissue (40). The complexity of the
pharmacokinetics of systemic iron supplementation makes it
difficult to translate the in vitro data into a clinical
context. For this reason, in vivo experiments with clinical
iron compounds are needed to answer the remaining questions about
transferring the results of the present study into a therapeutic
approach.
Overall, it was shown for the first time that
FeCl3 does not synergize with high-dose ascorbate in the
treatment of pancreatic cancer cells and may even reduce the
antitumor effect of ascorbate. The explanation for this could be
the reduced TfR1 expression after iron treatment, which results in
reduced LIP. For this reason, when using high-dose ascorbate in
tumor therapy, simultaneous iron administration should be carried
out with caution and patients' iron levels should be monitored.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Mrs. Monika
Schumacher (Technical Assistant, Department of Nutritional
Biochemistry, Institute of Nutritional Sciences, University of
Hohenheim) for her excellent technical support in conducting the
experiments.
Funding
The present study was supported by the Dr. Hans Fritz Stiftung
(grant no. 3140080501), the Ministry of Rural Affairs and Consumer
Protection Baden-Wuerttemberg [grant no. AZ: 16 (34) 8402.43/0454
E], the PASCOE pharmazeutische Praeparate GmbH (grant nos.
D.31.15100 and D.31.22506) and the Orthomol Pharmazeutische
Vertriebs GmbH (grant no. 3140080701). Publishing fees supported by
Funding Programme Open Access Publishing of University of
Hohenheim.
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
AP conceptualized the study, curated data, conducted
investigation, developed methodology, and visualized data. CL
developed methodology and performed software analysis. HS, KD, KS,
JS, SH, and LM curated data, conducted investigation and developed
methodology. OR conceptualized the study and acquired funding. MB
conceptualized and supervised the study and acquired funding. SV
conceptualized and supervised the study, acquired funding, and
conducted project administration. All authors wrote, reviewed, and
edited the manuscript. All authors read and approved the final
version of the manuscript. MB and SV confirm the authenticity of
all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Use of artificial intelligence tools
During the preparation of this work, artificial
intelligence tools were used to improve the readability and
language of the manuscript or to generate images, and subsequently,
the authors revised and edited the content produced by the
artificial intelligence tools as necessary, taking full
responsibility for the ultimate content of the present
manuscript.
Glossary
Abbreviations
Abbreviations:
|
AM
|
acetoxymethyl ester
|
|
BIP
|
2′,2′-bipyridyl
|
|
BSA
|
bovine serum albumin
|
|
CQ
|
chloroquine
|
|
DAPI
|
diamidino-2-phenylindole
|
|
DCFH-DA
|
dichlorodihydrofluorescein
diacetate
|
|
DFO
|
deferoxamine mesylate
|
|
FCS
|
fetal calf serum
|
|
GPX4
|
glutathione peroxidase 4
|
|
HRP
|
horseradish peroxidase
|
|
LIP
|
labile iron pool
|
|
MUH
|
4-methylumbeliferyl heptanoate
|
|
PI
|
propidium iodide
|
|
RAPA
|
rapamycin
|
|
ROS
|
reactive oxygen species
|
|
RSL3
|
RAS-selective lethal 3
|
|
STS
|
staurosporine
|
|
TCA
|
trichloroacetic acid
|
|
TBH
|
tert-butyl hydroperoxide
|
|
TBST
|
Tris-buffered saline containing 0.1%
Tween-20
|
|
TfR1
|
transferrin receptor 1
|
References
|
1
|
Siegel RL, Giaquinto AN and Jemal A:
Cancer statistics, 2024. CA Cancer J Clin. 74:12–49.
2024.PubMed/NCBI
|
|
2
|
Rahib L, Wehner MR, Matrisian LM and Nead
KT: Estimated projection of US cancer incidence and death to 2040.
JAMA Netw Open. 4:e2147082021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
American Cancer Society, . Cancer facts
& figures. 2024.Available from:. https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/2024-cancer-facts-figures.html
|
|
4
|
National Cancer Institute, . Cancer Stat
Facts: Common Cancer Sites. 2025.Available from:. https://seer.cancer.gov/statfacts/html/common.html
|
|
5
|
Cameron E and Pauling L: Supplemental
ascorbate in the supportive treatment of cancer: Prolongation of
survival times in terminal human cancer. Proc Natl Acad Sci USA.
73:3685–3689. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cameron E and Pauling L: Supplemental
ascorbate in the supportive treatment of cancer: Reevaluation of
prolongation of survival times in terminal human cancer. Proc Natl
Acad Sci USA. 75:4538–4542. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ou J, Zhu X, Lu Y, Zhao C, Zhang H, Wang
X, Gui X, Wang J, Zhang X, Zhang T and Pang CLK: The safety and
pharmacokinetics of high dose intravenous ascorbic acid synergy
with modulated electrohyperthermia in Chinese patients with stage
III–IV non-small cell lung cancer. Eur J Pharm Sci. 109:412–418.
2017. View Article : Google Scholar
|
|
8
|
Padayatty SJ, Sun H, Wang Y, Riordan HD,
Hewitt SM, Katz A, Wesley RA and Levine M: Vitamin C
pharmacokinetics: Implications for oral and intravenous use. Ann
Intern Med. 140:533–537. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Böttger F, Vallés-Martí A, Cahn L and
Jimenez CR: High-dose intravenous vitamin C, a promising
multi-targeting agent in the treatment of cancer. J Exp Clin Cancer
Res. 40:3432021. View Article : Google Scholar
|
|
10
|
Kim JH, Hwang S, Lee JH, Im SS and Son J:
Vitamin C suppresses pancreatic carcinogenesis through the
inhibition of both glucose metabolism and wnt signaling. Int J Mol
Sci. 23:122492022. View Article : Google Scholar
|
|
11
|
O'Leary BR, Alexander MS, Du J, Moose DL,
Henry MD and Cullen JJ: Pharmacological ascorbate inhibits
pancreatic cancer metastases via a peroxide-mediated mechanism. Sci
Rep. 10:176492020. View Article : Google Scholar
|
|
12
|
Bodeker KL, Smith BJ, Berg DJ,
Chandrasekharan C, Sharif S, Fei N, Vollstedt S, Brown H, Chandler
M, Lorack A, et al: A randomized trial of pharmacological
ascorbate, gemcitabine, and nab-paclitaxel for metastatic
pancreatic cancer. Redox Biol. 77:1033752024. View Article : Google Scholar
|
|
13
|
Alexander MS, Wilkes JG, Schroeder SR,
Buettner GR, Wagner BA, Du J, Gibson-Corley K, O'Leary BR, Spitz
DR, Buatti JM, et al: Pharmacologic ascorbate reduces
radiation-induced normal tissue toxicity and enhances tumor
radiosensitization in pancreatic cancer. Cancer Res. 78:6838–6851.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen P, Reed G, Jiang J, Wang Y, Sunega J,
Dong R, Ma Y, Esparham A, Ferrell R, Levine M, et al:
Pharmacokinetic evaluation of intravenous Vitamin C: A classic
pharmacokinetic study. Clin Pharmacokinet. 61:1237–1249. 2022.
View Article : Google Scholar
|
|
15
|
Polireddy K, Dong R, Reed G, Yu J, Chen P,
Williamson S, Violet PC, Pessetto Z, Godwin AK, Fan F, et al: High
dose parenteral ascorbate inhibited pancreatic cancer growth and
metastasis: Mechanisms and a phase I/IIa study. Sci Rep.
7:171882017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Welsh JL, Wagner BA, van't Erve TJ, Zehr
PS, Berg DJ, Halfdanarson TR, Yee NS, Bodeker KL, Du J, Roberts LJ
II, et al: Pharmacological ascorbate with gemcitabine for the
control of metastatic and node-positive pancreatic cancer (PACMAN):
Results from a phase I clinical trial. Cancer Chemother Pharmacol.
71:765–775. 2013. View Article : Google Scholar
|
|
17
|
Fan D, Liu X, Shen Z, Wu P, Zhong L and
Lin F: Cell signaling pathways based on vitamin C and their
application in cancer therapy. Biomed Pharmacother. 162:1146952023.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Piotrowsky A, Burkard M, Schmieder H,
Venturelli S, Renner O and Marongiu L: The therapeutic potential of
vitamins A, C, and D in pancreatic cancer. Heliyon. 11:e415982025.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Du J, Cullen JJ and Buettner GR: Ascorbic
acid: Chemistry, biology and the treatment of cancer. Biochim
Biophys Acta. 1826:443–457. 2012.
|
|
20
|
González-Montero J, Chichiarelli S, Eufemi
M, Altieri F, Saso L and Rodrigo R: Ascorbate as a bioactive
compound in cancer therapy: The old classic strikes back.
Molecules. 27:38182022. View Article : Google Scholar
|
|
21
|
Schoenfeld JD, Alexander MS, Waldron TJ,
Sibenaller ZA, Spitz DR, Buettner GR, Allen BG and Cullen JJ:
Pharmacological ascorbate as a means of sensitizing cancer cells to
Radio-chemotherapy while protecting normal tissue. Semin Radiat
Oncol. 29:25–32. 2019. View Article : Google Scholar
|
|
22
|
Leischner C, Marongiu L, Piotrowsky A,
Niessner H, Venturelli S, Burkard M and Renner O: Relevant membrane
transport proteins as possible gatekeepers for effective
pharmacological ascorbate treatment in cancer. Antioxidants
(Basel). 12:9162023. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Piotrowsky A, Burkard M, Hammerschmidt K,
Ruple HK, Nonnenmacher P, Schumacher M, Leischner C, Berchtold S,
Marongiu L, Kufer TA, et al: Analysis of High-dose
ascorbate-induced cytotoxicity in human glioblastoma cells and the
role of dehydroascorbic acid and iron. Antioxidants (Basel).
13:10952024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou L, Zhang L, Wang S, Zhao B, Lv H and
Shang P: Labile iron affects pharmacological ascorbate-induced
toxicity in osteosarcoma cell lines. Free Radic Res. 54:385–396.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schoenfeld JD, Sibenaller ZA, Mapuskar KA,
Wagner BA, Cramer-Morales KL, Furqan M, Sandhu S, Carlisle TL,
Smith MC, Abu Hejleh T, et al: O2·- and H2O2-mediated
disruption of fe metabolism causes the differential susceptibility
of NSCLC and GBM cancer cells to pharmacological ascorbate. Cancer
Cell. 31:487–500.e8. 2017. View Article : Google Scholar
|
|
26
|
Du J, Wagner BA, Buettner GR and Cullen
JJ: Role of labile iron in the toxicity of pharmacological
ascorbate. Free Radic Biol Med. 84:289–295. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Burkard M, Niessner H, Leischner C,
Piotrowsky A, Renner O, Marongiu L, Lauer UM, Busch C, Sinnberg T
and Venturelli S: High-dose ascorbate in combination with Anti-PD1
checkpoint inhibition as treatment option for malignant melanoma.
Cells. 12:2542023. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Szarka A, Kapuy O, Lőrincz T and Bánhegyi
G: Vitamin C and cell death. Antioxid Redox Signal. 34:831–844.
2021. View Article : Google Scholar
|
|
30
|
Chen X, Comish PB, Tang D and Kang R:
Characteristics and biomarkers of ferroptosis. Front Cell Dev Biol.
9:6371622021. View Article : Google Scholar
|
|
31
|
Brandt KE, Falls KC, Schoenfeld JD, Rodman
SN, Gu Z, Zhan F, Cullen JJ, Wagner BA, Buettner GR, Allen BG, et
al: Augmentation of intracellular iron using iron sucrose enhances
the toxicity of pharmacological ascorbate in colon cancer cells.
Redox Biol. 14:82–87. 2018. View Article : Google Scholar
|
|
32
|
Deme S, Ramezani I, Coulter J, Paller C
and Bressler J: Effects of hypoxia and iron on ascorbic
acid-mediated cytotoxicity in prostate cancer cell lines. Toxicol
Appl Pharmacol. 497:1172592025. View Article : Google Scholar
|
|
33
|
Wang F, He MM, Xiao J, Zhang YQ, Yuan XL,
Fang WJ, Zhang Y, Wang W, Hu XH, Ma ZG, et al: A randomized,
open-label, multicenter, phase 3 study of high-dose Vitamin C Plus
FOLFOX ± bevacizumab versus FOLFOX ± bevacizumab in unresectable
untreated metastatic colorectal cancer (VITALITY study). Clin
Cancer Res. 28:4232–4239. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mohammad RM, Muqbil I, Lowe L, Yedjou C,
Hsu HY, Lin LT, Siegelin MD, Fimognari C, Kumar NB, Dou QP, et al:
Broad targeting of resistance to apoptosis in cancer. Semin Cancer
Biol. 35 (Suppl(0)):S78–S103. 2015. View Article : Google Scholar
|
|
35
|
Renner O, Burkard M, Michels H, Vollbracht
C, Sinnberg T and Venturelli S: Parenteral high-dose ascorbate-A
possible approach for the treatment of glioblastoma (Review). Int J
Oncol. 58:352021. View Article : Google Scholar
|
|
36
|
Wu K, Liu L, Wu Z, Huang Q, Zhou L, Xie R
and Wang M: Ascorbic acid induces ferroptosis via STAT3/GPX4
signaling in oropharyngeal cancer. Free Radic Res. 58:117–129.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang X, Xu S, Zhang L, Cheng X, Yu H, Bao
J and Lu R: Vitamin C induces ferroptosis in anaplastic thyroid
cancer cells by ferritinophagy activation. Biochem Biophys Res
Commun. 551:46–53. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu Y, Huang P, Li Z, Xu C, Wang H, Jia B,
Gong A and Xu M: Vitamin C sensitizes pancreatic cancer cells to
erastin-induced ferroptosis by activating the AMPK/Nrf2/HMOX1
pathway. Oxid Med Cell Longev. 2022:53612412022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang C, Liu X, Jin S, Chen Y and Guo R:
Ferroptosis in cancer therapy: A novel approach to reversing drug
resistance. Mol Cancer. 21:472022. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Schaefer B, Meindl E, Wagner S, Tilg H and
Zoller H: Intravenous iron supplementation therapy. Mol Aspects
Med. 75:1008622020. View Article : Google Scholar : PubMed/NCBI
|














