The analysis of microbiota composition has become
routine in our time, mostly due to the explosion and availability
of new technologies [metagenomic sequencing technologies that
incorporate next-generation DNA sequencing methods with the
computational approach of targeted (16S rRNA hypervariable regions)
or whole-genome shotgun sequence reads], that allow for the
identification and classification of a great variety of microbial
species (1,2). The genome of the microbiota is
100-fold more extensive than the human genome (3). The distribution of microbial cells
surpasses the number of all human cells, including somatic and germ
cells throughout the human body (4-8).
Microbiota is a term that has been established as the sum of
bacteria, fungi, parasites and viruses occupying the host (7). The microbiota genome is known as the
‘microbiome’, which acts as 'an indirect genome' with significant
functions that are several fold more numerous than those of the
human genome (6,7,9,10).
The microbiota and host form a complex ‘super-organism’ in which
their symbiotic association confers benefits to the host regarding
key aspects of life. However, defects in the host regulatory
circuits that control bacterial sensing and homeostasis, or
alterations in the microbiome, through environmental changes
(infection, diet or lifestyle), may disrupt the symbiotic
relationship and promote disease. Increasing evidence indicates a
key role for bacterial microbiota in carcinogenesis (6,7,9-11).
The International Agency for Research on Cancer (IARC) has
classified only 10 microorganisms from the total number of 3.7x1030
microorganisms as pathogenic (12).
The microbiota colonizes any surface in the human
body, with significant functional roles in homeostasis. Each
surface in different organs (such as the lungs, skin, vagina and
oral cavity) is exposed to the external environment and has its
distinct microbiome. The highest percentage of microbial mass is
located in the gastrointestinal tract (99%), which is the reason
why the intestinal microbiota is the most extensively investigated
type of microbiota thus far (13,14). In particular, 100 trillion
micro-organisms of distinct structures (bacteria, parasites, fungi
and viruses) are colonized in the human intestine, thus termed the
intestinal microbiome (15,16). The absolute number of
micro-organisms fluctuates between the mouth and rectum (17).
In order to establish the composition of the
microbiota, it is only reasonable to consider a human being from
the early stages of life, i.e. birth. The microbial composition is
acquired during the first three years of childhood; following the
initial stages, microbiota development continues with environmental
exposure (17-19,29).
As a general note, the gut microbiota displays a consistent
composition and differs among individuals. During late stages of
life, in the elderly, the composition of the microbiota changes
again, but retains a stable function (21-24).
With regards to the microbiota of the human
gastrointestinal tract, 50 distinct phyla and five hundred
bacterial species have been reported as dominant (25). High throughput sequencing
techniques have provided deep insight into the composition of the
microbiota (26). A high
proportion of gut microbiota is divided into three categories: The
Firmicutes (30-50%), the Bacteroidetes (20-40%) and Actinobacteria
(1-10%) (27). The majority of
the microbiota is strictly anaerobic, such as Bacteroides,
Eubacterium, Bifidobacterium, Fusobacterium, Peptostreptococcus and
Atopobium, whereas the minority is facultative anaerobic and
comprises Lactobacilli, Enterococci, Streptococci and
Enterobacteriaceae (28). The gut
microbiota varies in mass and composition along the gut, that is,
from the luminal to the mucosal regions (17,25). For example, microbial composition
is denser in the large intestine as compared to the small
intestine, potentially explaining the higher susceptibility of the
large intestine to cancer (30,31). The close association between the
gut microbiota and colon cancer was initially revealed in 1975,
when researchers observed that the potential of developing colon
cancer in a chemically-dependent manner was reduced in germ-free
rats (32). Furthermore, the
colon is dominated by Bacteroidetes and Actinobacteria (90%),
whereas the small intestine is colonized by Bacteroidetes and
Actinobacteria (50%). The composition of the microbiota in the
small intestine also includes other bacterial phyla, such as
Firmicutes species (40%) (33).
Despite the fluctuations of microbiota in composition and mass
along the gut, the main population of micro-organisms is most
certainly composed of bacteria, residing within the
gastrointestinal lumen (34),
either competing or cooperating with other micro-organisms
(35).
The symbiotic association of micro-organisms with
the human gut (host) is accomplished after several years of
co-evolution and co-adaptation, contributing to a balanced
homeostasis in the gut (4,36-38).
The microbiota plays a key role in homeostasis, as confirmed by its
participation in a wide range of processes, such as wound healing,
the maintenance of barrier function, and the modulation of cellular
growth and immune system regulation (39). Apart from participating in these
processes, the microbiota also expands to the digestion of complex
carbohydrates and the establishment of ecological niches that might
otherwise be occupied by non-innocuous microorganisms (11). The gut microbiota is characterized
by a high self-restitution capacity following perturbation
(40).
Other beneficial functions of the microbiota include
a significant contribution to the maturation of the immune system
and to the protection against infectious agents (41-45).
It is important to highlight that the murine gut microbiota
displays high homology to the human gut microbiota, paving the way
for translational research based on experimental mouse models
(46,47). For example, the microbiota has
been reported to actively participate in the developmental stages
of a healthy immune system, as illustrated by severe immune defects
in mice bred under germ-free conditions (18,48). As a general note, the immune
system of the gut is responsible for eradicating pathogenic
microbes, whereas at the same time it has developed
immuno-tolerance mechanisms against classical gut microbes. The
mucosal immune system is under the control of the adaptive immune
system and it functions in a cell-autonomous manner (39). The disruption of the microbiota
has been shown to confer a significant impact on the immune system
in many aspects. Importantly, the microbiota has been demonstrated
to affect immune structures, in addition to immune cell
populations. This was initially illustrated in experimental mice
bred under germ-free conditions, which displayed hallmark changes
in immunoglobulin A (IgA) secretion and functionality defects in
Peyer's patches and draining mesenteric lymph nodes (mLNs)
(49,50). Notably, in a previous study,
gut-associated lymphoid tissues (GALTs) were efficiently matured
with the concomitant activation of T cells and IgA-secreting plasma
cells in the presence of the microbiota, which mediated the
necessary signals for both epithelial and dendritic cell (DC)
activation (51). Furthermore,
the gut microbiota appears to be essential for the function of
basic immune populations, such as the secretion of interleukin
(IL)-22 by type 3 innate lymphoid cells (ILC3 cells). ILC3 cells
have been shown to be essential for the growth of T cells in a
microbiota status-dependent manner, independently of IL-22, IL-23
or IL-17 synthesis (52).
Subsequent scientific evidence has suggested that some bacterial
strains are particularly associated with the functions of the
immune system. For example, certain microbiota components seem to
initiate inflammation and to regulate immune cells within the
lamina propria of the intestine. The absence of segmented
filamentous bacteria (SFB) causes low enrichments of IgA titers,
the reduction of T helper 1 (Th1) and T helper 17 (Th17) cells, and
the alleviation of immune responses to classical intestinal
pathogens (Citrobacter rodentium and Salmonella spp.) (48,53,54). Yang et al highlighted the
significance of SFB in the function of T helper 17 (Th17) cells,
through the expression of a T cell receptor (TCR) directed to a
specific SFB antigen (55).
Despite the beneficial effects mediated by SFB, it was noted that
SFB also increases susceptibility to autoimmune disorders (56). Specifically, it has been
demonstrated that the induction of SFB to germ-free mice renders
them susceptible to the development of collagen-induced arthritis
(57). In addition, the
expression of the innate-like cytokine IL-17C seems to be under the
control of microbiota during intestinal tumorigenesis, as the
latter was shown to mediate Toll-like receptor (TLR)/MyD88
signaling, which in turn lead a to IL-17C upregulation during colon
cancer progression and ultimately to the uncontrolled proliferation
of intestinal epithelial cells (IECs) (58).
On the other hand, the gut microbiota also has the
capacity to induce an anti-inflammatory environment, by producing
certain metabolic by-products that maintain barrier integrity. For
example, the differentiation of IL-10-secreting Tregs has been
shown to be absolutely dependent on the signal transduction pathway
triggered by Bacteroides fragilis (51). Specifically, polysaccharide A
secreted by Bacteroides fragilis induces Treg cell expansion via
the TLR2 signaling pathway (51).
Similarly, it has been reported that Helicobacter hepaticus (Hh)
stimulates T cells to differentiate into T regulatory cells
(59). The induction of Tregs was
also observed following the incubation of clostridial strains,
conferring significant benefits to experimentally-induced colitis
(35,60). For example, Faecalibacterium
prausnitzii is a clostridial organism that has been shown to
protect patients from the onset of inflammatory bowel disease (IBD)
(61). The gut microbiota
therefore appears to be indispensable for the immune system as a
whole (62).
Apart from the gut, the skin also harbors a
significant number of microbial niches that sustain the recruitment
of Th1/Th17 cells and provide protection against pathogens, such as
Leishmania major (63).
Indicatively, the functionality and persistent response of CD8+ T
cells has been attributed to the skin microbe, Staphylococcus
epidermidis (64). The oral
cavity also contains microbial communities with key roles in
modulating persistent immune responses to various infections
(65,66). For example, it has been
demonstrated that in the absence of microbiota in the oral cavity,
immune cells cannot combat mucosal influenza virus (67). Even though the microbiota exerts
its effect on the immune system locally on each surface barrier,
the gut microbiome appears to be the most efficient in controlling
systemic immune homeostasis (67-69).
The insuperable effect of the gut microbiome on the immune system
has been attributed to its great variability, the high number of
associated micro-organisms and the relatively large surface area
that is available to expand on (13). In line with this, it has been
demonstrated that the incubation of certain bacterial strains in
the skin of germ-free mice does not seem to display systemic
effects and to reconstitute Th cell populations in the intestine
(63). The significance of the
presence of the microbiota in various anatomical sites was
established when experimental animal models lacking microbial
communities exhibited signs of autoimmune disorders (multiple
sclerosis or arthritis) (57,70,71).
However, as described above, the microbiota does not
only include populations of bacteria, but also consists of other
micro-organisms, such as archaea, fungi, viruses, etc. For example,
fungi such as Candida appear to be overexpressed in animal models
following treatment with antibiotics (72). Nonetheless, the role of microbiota
subtypes other than bacteria is still in its infancy and additional
studies are required in order to assess their impact on the immune
system (72,73).
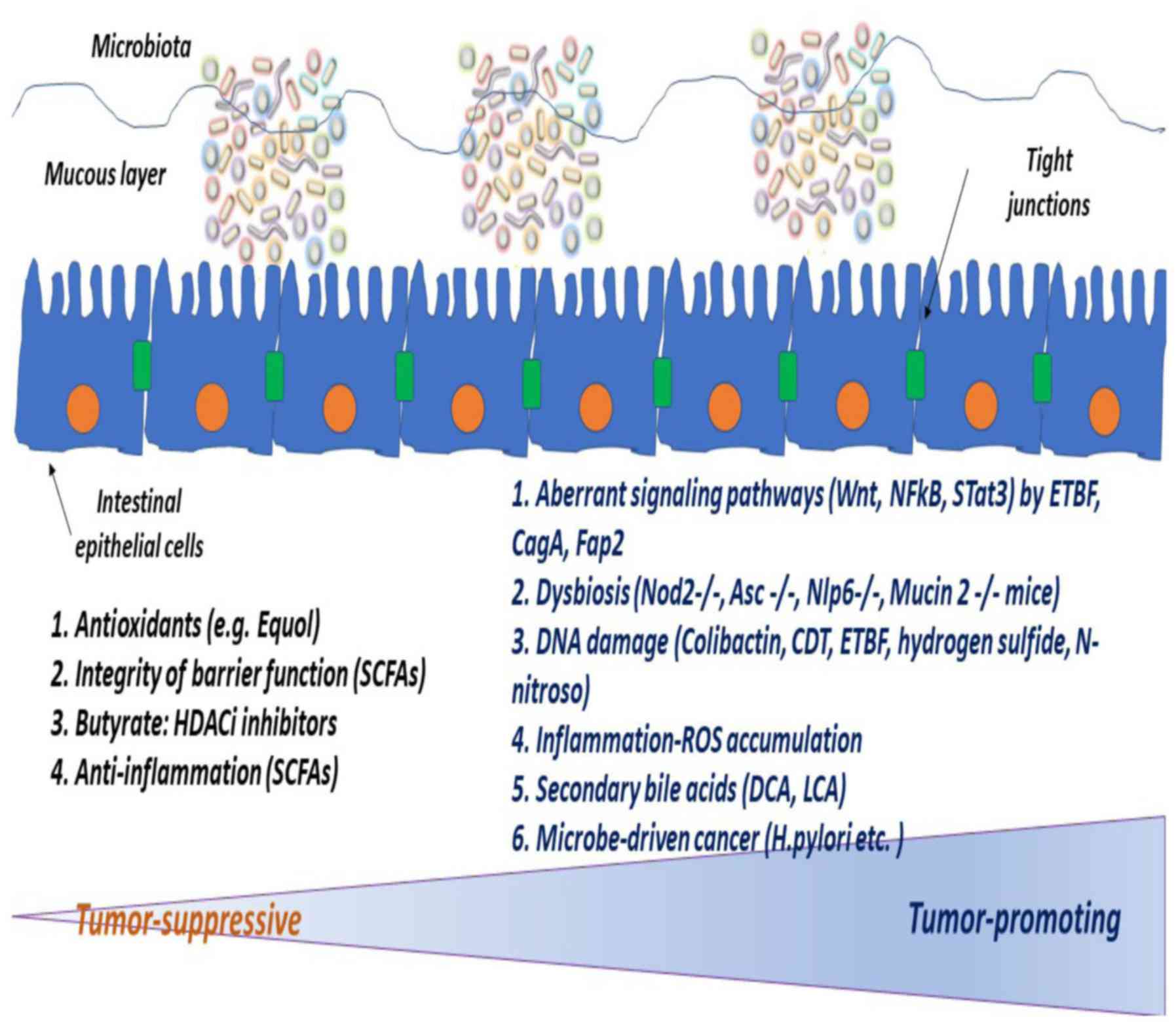
The protection of the gut against exogenous
pathogens is provided by the presence of the epithelial barrier.
The integrity of the epithelial barrier is mediated through tight
junctions between epithelial cells, the mucous layer, soluble
antibacterial factors and distinct cells of innate and adaptive
immunity (74-76).
Under normal gut conditions, one hundred trillion organisms
(particularly microbiota) thrive in the intestine, creating a
protected, warm and nutrient-rich microenvironment, which in turn
helps the microbiota to be in equilibrium with the host organism
(eubiosis). When eubiosis is disrupted, the composition of the
microbiota is altered and as a consequence, it is mostly
represented by facultative anaerobic bacteria instead of aerobic
bacteria (present in healthy conditions) (11). The interaction of the microbiota
with epithelial cells is indirect, through the mucous layer, which
separates the compartment of commensal bacteria from that of the
host (49). If one considers that
the gut microbiota is in close proximity to the IECs that line the
mucous layer, it is only to be expected that dysbiosis can rupture
the mucous layer, thereby leading to inflammatory conditions or
even cancer (7,11,62). The mucous layer can be ruptured by
microbial translocation, due to specific molecular alterations or
abnormal regulatory signals (77).
One of the hallmarks of cancer is the uncontrolled
growth of malignant cells, which ultimately constitutes one of the
main causes of mortality. Colorectal cancer is the most predominant
type of cancer in the USA and the third most common cause of
mortality, therefore presenting a considerable tumor burden
(78). Colorectal cancer, as the
name implies, is characterized by carcinogenic alterations that
occur in the colon and rectal epithelial cells. Different forms of
colon cancer arise due to variations in genetic profiles,
histological patterns and sensitivity to potential therapeutic
drugs (79,80). Indeed, colorectal cancer can be
divided into three subtypes, the first of which (35%) is ultimately
linked to genetic alterations, the second (65%) is associated with
exogenous factors and the third, which accounts for 1% of all
colorectal cancer subtypes, is accompanied by chronic IBD (81). Consequently, the majority of
patients with colorectal cancer (95%) are not directly genetically
vulnerable to cancer, thereby supporting the notion that the gut
microbiota is actively implicated in cancer development (82).
The uncontrolled growth of intestinal malignant
cells begins with the conversion of normal epithelial cells into
hyperplastic cells. In this manner, epithelial cells lose their
morphological characteristics and become dysplastic. This is
followed by the invasion of hyperproliferative epithelial cells
into the gut lumen, where they form adenomas, and the subsequent
protrusion of epithelial cells into the gut, which ultimately leads
to cancer. From a genetic aspect, IECs are transformed into
hyperplastic intestinal cells after the following sequence of
genetic events: First the loss of tumor suppressor genes, such as
adenomatous polyposis coli (Apc) and subsequently mutations in
genes that encode the machinery for DNA repair, such as hMSH2.
Even though significant efforts have been made in
order to elucidate the driver mechanisms that cause colorectal
carcinogenesis, the landscape remains obscure. The effect of the
gut microbiota on carcinogenesis has become a burgeoning issue of
research in recent times, considering that the gut microbiota is
vital in sustaining the homeostasis and regulation of the immune
system. Since colorectal carcinogenesis is a multifactorial cancer
type with a genetic basis, it has been proposed that inflammatory
processes or perturbations of the intestinal microbiota can lead to
the cancer development through genetic alterations. The impact of
the gut microbiota appears to be more prominent in colon cancer,
independent of the type and cause, where it seems to systemically
influence cancer progression either as individual species or as a
microbial community (12).
If one considers that microbes create a dynamic
symbiotic interplay with the host, it is logical to assume that the
gut microbiota can function either as a blocker or as a potentiator
of colon cancer (Fig. 1). The
first efforts made in the treatment of cancer through the natural
effects of the gut microbiota were made in the late nineteenth
century, when it was realized that bacterial infections or
injections of heat-killed bacteria, such as Coley's toxin prevented
the development of sarcoma in patients (83,84). Nowadays, Coley's toxin, which
includes a mixture of killed bacteria (e.g., Streptococcus pyogenes
and Serratia marcescens) is regarded a precursor of current
immunotherapeutic agents. For example, the injection of
Mycobacterium bovis Bacillus Calmette-Guérin (BCG) is currently
used as the classical therapeutic approach in non-muscle invasive
bladder cancer (85). Consistent
with the above, experiments have demonstrated that animals bred
under germ-free conditions exhibit a stronger likelihood for
developing colon cancer than those bred under microbiota-rich
environments (86).
Additional in vivo experiments have indicated that
the microbiota has the capacity to induce cancer in a wider range
of organs than previously considered, including the skin, colon,
liver, breast and lungs (86-93).
In a similar manner, it was shown that the elimination of
intestinal microbiota (through antibiotics) ameliorates colorectal
cancer or hepatic carcinogenesis (88,94-97).
Consequently, the majority of studies have
concentrated on investigating the role of the gut microbiota in
colitis-associated colorectal cancers, using either germ-free
conditions or antibiotics (90,98). Using genetically engineered
experimental models specifically predisposed to cancer, such as
TCR-deficient (TCRb-/-) and double p53 KO (X p53-/-) mice, it was
previously demonstrated that the gut microbiota can indeed induce
colon cancer. The animals did not develop adenocarcinomas under
germ-free conditions (99);
however, TGFb1-/- mice gut-colonized with Helicobacter hepaticus
demonstrated a greater potential for colon cancer development
(100). Similarly, the gut
colonization of Rag2-/- mice with Helicobacter hepaticus, and that
of Tbet-/- Rag2-/- mice with microbiota, has been shown to induce
colon carcinogenesis (101,102). The resulting phenotype of these
experimental animal models (TGFb1-/- and Tbet-/- Rag2-/-) under
germ-free conditions was very similar to the phenotype following
antibiotic treatment (98,103).
Additional studies reported that IL10-/- mice developed tumors
following treatment with the chemical carcinogen azoxymethane (AOM)
and incubation with Enterococcus faecalis, as compared to mice bred
under germ-free conditions, which remained healthy (90,104).
However, since AOM, in combination with dextran
sulfate sodium (DSS), has been shown to induce colon cancer
formation, the role of the gut microbiota in colorectal
carcinogenesis in experimental mouse models may be obscured by the
effects of chemically induced carcinogenesis (92,105-110).
In other words, it is difficult to assess which animal models of
colorectal cancer are the most suitable for elucidating the role of
the microbiome in cancer development. Despite the seemingly
unsurpassed selective pressures displayed by malignant cells, the
concept of microbe-driven cancer formation does not cease to exist.
A significant number of research studies have explored whether
micro-organisms are directly involved in tumor progression or
whether they participate indirectly through their metabolites.
Recent metagenomic analyses have revealed essential differences
between healthy and cancer states and assessed the tumor-promoting
effects of microbiota in certain types of cancer. The mechanisms
through which specific microbes target certain cancers,
contributing to the acceleration of tumor progression remain
elusive. For example, human papillomavirus (HPV), hepatitis B
virus, hepatitis C virus, human herpesvirus 8 and human
T-lymphotropic virus 1 have been shown to trigger carcinogenesis
via well-defined processes (5,11).
Specifically, HPV can cause anogenital or oropharyngeal carcinomas,
hepatic B or C virus can induce hepatocellular carcinoma and human
immunodeficiency virus (HIV) or Epstein-Barr virus (EBV) or human
T-cell lymphotropic virus type 1 (HTLV-1) can lead to lymphoma
(7). As regards individual
bacterial species, it has been revealed that specific bacterial
strains drive carcinogenesis (111). As highlighted by epidemiological
data, the most highly-associated microbe to cancer development,
especially gastric cancer, is Helicobacter pylori (H. pylori)
(112). Helicobacter pylori
releases toxins (CagA or VacA) that cause cytoskeletal
rearrangements which cannot be surpassed by the host repair
mechanisms (113,114). Notably, the close association of
Helicobacter pylori with gastric cancer has been regarded a
landmark discovery and was awarded a Nobel Prize (115). Furthermore, Streptococcus bovis,
Helicobacter pylori, Bacteroides fragilis, Enterococcus faecalis,
Clostridium septicum, Fusobacterium spp. and Escherichia coli have
also been identified as bacterial species that can lead to the
development of intestinal neoplasms. Other bacterial strains,
including Bacteroides fragilis, Escherichia coli, Enterococcus
faecalis and Fusobacterium nucleatum, have been identified in
experimental animal models chemically predisposed to colon cancer
as microbes with an ability to modulate normal immune responses
(Table I) (116-121).
An important breakthrough was achieved in the case
of Fusobacterium nucleatum. In general, Fusobacterium nucleatum is
localized in the oral cavity. As a resident member of the oral
microbiota, Fusobacterium nucleatum has been extensively studied
and has been found highly associated with periodontitis and
appendicitis (122).
Fusobacterium nucleatum is an anaerobic Gram-negative rod bacterium
and one of the leading micro-organisms in intrauterine infections
causing premature death. Compared to the gut microbiota of healthy
individuals, Fusobacteria are often found in patients with colon
adenocarcinoma and IBD, thus suggesting an association between
Fusobacteria and the colon inflammatory environment (120,123-126).
In the human body, Fusobacterium nucleatum can be introduced
through the oral cavity and transferred into the gastrointestinal
tract, thereby affecting human colon adenocarcinoma. From an immune
point of view, Fusobacterium nucleatum triggers inflammatory
signaling pathways and functions as a shield to tumor cells against
an immune attack (12,127). It has been proposed that certain
bacterial strains possess many adhesins, mediating their binding to
TLR4 and RIG-I, as well as their direct interaction with natural
killer (NK) cells via binding to the NKp46 receptor (128-130).
In this context, the Fap2 protein of Fusobacterium nucleatum has
been shown to bind to the human (not mouse) TIGIT [T cell
immunoreceptor with Ig and ITIM (immunoreceptor tyrosine-based
inhibitory motif)] receptor present on NK cells and T cells in a
hemagglutination-dependent manner (Table I) (127). The hemagglutination potency of
Fap2 has been tightly linked to TIGIT suppression. In this manner,
Fusobacterium nucleatum manages to abolish NK cell-mediated
destruction of human cancer cells. Notably, Fusobacterium nucleatum
has been shown to bind to many tumor cells through interactions of
its Fap2 protein with the Gal-GalNac protein of cancer cells.
Consistent with the above, the exposure of experimental ApcMin/+
animal models to Fusobacterium nucleatum has been shown to cause
enriched myeloid cell infiltration (predominantly DCs, macrophages)
and the activation of the nuclear factor (NF)-κB signal
transduction pathway. In addition, Fusobacterium nucleatum has been
shown to augment the numbers of two types of myeloid-derived
suppressor cells (MDSCs), thus inhibiting activated T cell
responses and exacerbating small intestinal adenocarcinoma
development (Table I) (120). Substantial experimental data
have highlighted that Fusobacterium nucleatum-fed mice are enriched
for tumor-associated macrophages (TAMs), tumor-associated
neutrophils (TANs), CD103+ regulatory DCs, thus exhibiting a
promotion of neoplastic progression (Table I) (120).
Fusobacterium nucleatum seems to recapitulate tumor
progression through its effects on the tumor microenvironment. For
example, Fusobacterium nucleatum has been shown to synthesize
hydrogen sulfide following red meat consumption, thus promoting DNA
damage responses and genomic instability in colon epithelial cells,
which can in turn lead to tumor development (131,132). The onset of colorectal carcinoma
and the extent of tumor progression appear to be more prominent in
individuals with mutations or perturbations in the DNA-damage
response (e.g., ATR and ATM). In support of this notion, both in
vitro and in vivo xenograft experiments have highlighted the
potential of Fusobacterium nucleatum to trigger the Wnt/β-catenin
pathway, via FadA binding, which is highly associated with the
proliferation of neoplastic cells (126).
Furthermore, it has been illustrated that
enterotoxigenic Bacteroides fragilis (ETBF) can potentiate tumor
formation by activating the signal transducer and activator of
transcription 3 (STAT3) signaling transduction pathway and
recruiting T helper 17 cells (Th17) in ApcMin/+ mice (Table I) (132). The oncogenicity of Bacteroides
fragilis became evident from its capacity to trigger the
Wnt/β-catenin pathway, thus aiding in distant site colonization by
tumor cells. Overall, Bacteroides, Escherichia coli and
Enterococcus faecalis are the bacterial strains found to be DNA
damage inducers in animal models (Table II) (119,133-136).
Enterococcus faecalis has been shown to stimulate the DNA damage
response in epithelial cells by secreting high levels of reactive
oxygen species (ROS) (119,137), Bacteroides fragilis has been
shown to trigger the Wnt signaling pathway and Escherichia coli is
able to initiate double-stranded DNA breaks, thus resulting in
increased genomic instability (133-135).
Escherichia coli, in particular, has been found to contain
polyketide synthase pathogenicity islands (pks), which contain the
gene responsible for the toxin (colibactin) that triggers DNA
damage response in IECs (Table
II) (134,138,139). The significant oncogenicity of
Escherichia coli was highlighted when IL10-/- mice developed
intestinal tumorigenesis following treatment with AOM and
Escherichia coli strains, without any indication of inflammatory
sites (134). Therefore, these
microbes (Enterococcus faecalis, Bacteroides fragilis, Escherichia
coli) are considered to be key mediators of the DNA damage response
and tumor progression, without the need for a pro-inflammatory
environment (Table II). Last but
not least, other bacterial strains have been shown to use other,
multifaceted mechanisms in order to induce carcinogenesis in animal
models (Table III) (140-143).
Even though recent data have suggested that
individual micro-organisms specifically influence the formation of
cancer, carcinogenesis may also be the result of altered microbiota
composition (dysbiosis) (144).
The defects in the symbiotic interplay between the host and
intestinal microbiota can result in alterations in the composition
of the microbiota or in defects in the regulatory signals that
orchestrate the normal association of microbiota with the host. In
this manner, the balance in the commensal community changes (termed
dysbiosis) and colon cancer can become the following event
(6,8). Possible causes of dysbiosis can be
either pathogenic microorganisms or environmental cues, such as
antibiotics, xenobiotics or obesity (11). Other causes may be genetic defects
in epithelial, myeloid, or lymphoid cells of the gut, which can
stimulate dysbiosis and consequently lead to inflammatory states,
such as Crohn's disease, which may in turn confer some
predisposition to carcinogenesis (35).
Dysbiotic bacteria appear to be indispensable to the
creation of an inflammatory environment in the gut. Nevertheless,
additional genetic changes are required for the initiation of
colorectal carcinogenesis (11,145,146).
A significant number of studies have investigated
the precise mechanisms through which the microbiota can be
implicated in tumor development. In general, experimental mouse
models chemically predisposed to colon cancer exhibit a lower
incidence of tumor formation when treated with antibiotics or when
bred under germ-free conditions, as compared to animals bred under
conventional conditions (86,92,93,95-97,147,148).
It remains to be clarified whether inflammation precedes or follows
dysbiosis, before ultimately leading to cancer. On one hand, it has
been suggested that a dysbiotic microbial community can lead to
carcinogenesis by inducing chronic inflammation (149). For example, IL-18-, IL-18R- and
MyD88-deficient mice are unable to mount adequate immune responses
due to intestinal dysbiosis that occurs through the expansion of
bacterial phyla Bacteroidetes (Prevotellaceae) and TM7, which
ultimately leads to colon cancer (108,150,151). Other immune-deficient mice
[Nod2-/-, Asc-/- (also known as Pycard-/-) and Nlrp6-/-] also
display dysbiotic microbiota and exhibit carcinogenesis (152,153). The functional significance of
dysbiotic microbiota has been highlighted by the fact that the
transfer of dysbiotic intestinal microbiota in healthy mice renders
them susceptible to colon cancer (151). Similarly, genetically-edited
mice (Tlr5 or IL1- or Tbx1 or Rag2 immunologically ablated) display
prominent signs of colon cancer due to dysbiotic microbiota, as
compared to immunologically wild-type mice (111). Last but not least, mice
deficient for mucin (total Mucin 2 KO) present a defective
intestinal barrier and for this they have been classified as models
of intestinal neoplasia (154).
On the other hand, it has been proposed that
inflammation causes barrier deterioration (dysbiosis), thus
facilitating bacterial translocation, which in turn facilitates the
creation of amplification feedback loops between intestinal barrier
loss and carcinogenesis (145).
For example, if mice display defects in pattern recognition
receptors which specifically bind to microbial-associated molecular
patterns (MAMPs), then these mice are characterized by bacterial
translocation, dysbiosis and ultimately exhibit carcinogenesis
(134,152,153).
In addition to the local gut microbiota effect on
cancer described above, other changes can cause long-distance
effects, determining the outcome of neoplasms other than colorectal
cancer (e.g., pancreatic, liver and breast cancer) (88,94,155-157).
A characteristic example of cancer caused by the distant effect of
dysbiotic microbiota is represented by hepatocellular carcinoma.
The long-distance effects to host organs are exerted by dysbiotic
intestinal microbiota either via the activation of pro-inflammatory
MAMPs or via the secretion of bacterial metabolites. For example,
the gut microbiota is capable of stimulating hepatocellular
carcinoma following entry into the liver through the portal vein
(88,94,158,159). Similarly, it has been
demonstrated that antibiotics ameliorate the progression of
hepatocellular carcinoma (88,159,160). Notably, dysbiotic microbiota
have been shown to influence estrogen metabolism, thereby affecting
tumors in distant sites (7). In
the lungs, Candida overgrowth has been observed following
antibiotic-mediated gut dysbiosis, with subsequent increases in
plasma prostaglandin E2 levels and macrophage differentiation
towards the M2 lineage (72).
These findings are in agreement with the results of epidemiological
studies supporting a strong link between dysbiosis and the
development of extracolonic neoplasms, including breast carcinoma
(161,162). It was concluded that bacteria
and their products are systemically distributed throughout the
body, compromising the integrity of the intestinal barrier
(94).
If one considers that cancer is tightly associated
with genetic diseases, it makes sense to assume that the microbiota
exerts a tumor-promoting role via genotoxic stress. The gut
microbiota is highly implicated in colorectal carcinogenesis
through the secretion of toxic metabolites as by-products of
fermentation. Toxic metabolites bind to specific surface receptors
on intestinal cells, thereby affecting key signaling pathways. As a
general note, microbiota-secreted toxins trigger DNA damage
response, causing cell cycle arrest, which is often followed by
apoptosis (116,134,139,163-167).
A particular toxin, termed CagA, secreted by
Helicobacter pylori, has been shown to induce both inflammation and
cancer (168,169). ETBF, another well-described
toxin, is secreted by Bacteroides fragilis and is also implicated
in colorectal carcinogenesis. Specifically, ETBF binds to the
epithelial receptor, stimulating the Wnt and NF-κB signal
transduction pathways, thus leading to enhanced cell proliferation
and DNA damage response (133,171-172). ETBF
stimulates IL-17 synthesis in the ApcMin/+ mouse model, thereby
predisposing it to intestinal neoplasia (116,172,173). The underlying molecular
mechanisms of ETBF have been shown to include the epithelial damage
through E-cadherin cleavage, which in turn activates the
β-catenin/Wnt pathway and the STAT3 signaling pathway (critical for
the growth of malignant cells) (116,174).
Since DNA damage is tightly associated with genomic
instability, proteins that are responsible for all the changes
caused by double-strand DNA breaks, such as cytolethal distending
toxin (CDT) and colibactin, can be regarded as true genotoxins
(139,165). Colibactin, however, stands out
among other toxins, as it is capable of inducing oxidative burst,
in addition to causing genome instability (134,139).
Bacteria-secreted toxins may also have a profound
impact on the oxidation status of cancer cells (175). Enterococcus faecalis seems to be
the main bacterial strain producing reactive oxygen intermediates
(superoxide and hydrogen peroxide) and inducing harmful changes in
epithelial cells and malignant transformation. Notably, these
effects are exacerbated in IL-10-deficient mice, suggesting that
the microbiota leads to colorectal carcinogenesis via reactive
toxins in an established inflammatory environment (104,176). It has also been indicated that
IECs are toxically hampered by sulfate-reducing bacteria through
production of hydrogen sulfide (H2S) (177,178). Finally, it has been argued that
bacteria can obtain virulence factors and convert them to
pathogens. The capacity of bacteria to bind to IECs seems to be
facilitated via the acquisition of virulence factors (11,179-181).
For example, FadA has been identified as the virulence factor
secreted by Fusobacterium nucleatum in order to activate colorectal
cancer (126,182). Similarly, the afa and eae
adhesins have been identified as the virulence factors released by
Escherichia coli strains in order to drive intestinal malignant
transformation (107,183).
Recent experimental data have confirmed that the gut
microbiota can synthesize an enormous quantity of metabolic
by-products that affect tumor progression either positively or
negatively, upon interaction with the host. In general, the
microbiota is responsible for metabolizing dietary factors into
bioactive food components. Commencal bacteria are known to exert
their fermentation capacity in the gut, metabolizing non-digestible
carbohydrates such as polysaccharides (e.g., resistant starch,
cellulose, hemicellulose, pectins and gums), oligosaccharides, and
lignins into short-chain fatty acids (SCFAs). The SCFAs are
composed of acetate, propionate and butyrate, which are regarded as
tumor suppressors with great anti-inflammatory and chemo-preventive
properties (184,185). SCFAs are final fermentation
products of dietary fiber in gut bacteria and provide the
appropriate energy to sustain the health of gut epithelial cells.
The type of diet directly influences bacterial abundance and
composition, as indicated by technologies, such as metagenomics
(effect of diet on microbiota), metaproteomics (microbial gene
expression) and metabolomics (microbial metabolites). On the other
hand, the microbiota can exert beneficial effects on the host
organism, as it is responsible for vitamin synthesis, such as
vitamin K and most B vitamins (186). In addition, carbohydrates,
branched chain amino acids, ammonia, amines, phenols, indoles and
phenylacetic acid are also generated through the actions of gut
microbiota (187,188). Several Bacteroides spp. and some
Firmicutes have been classified as the bacteria responsible for the
synthesis of phenylacetic acid, phenols, indoles and p-cresol.
These metabolites are known to be quite toxic as they cause the
nitrogen alkylation of DNA (189,190). For example, N-nitroso compounds
(NOCs) are exogenously supplied or endogenously synthesized through
the nitrosation of amines by gut microbiota. The abundance of NOCs
has been positively linked to an increased incidence of colorectal
cancer in European populations (94). Some products, such as ammonium are
carcinogenic despite being produced at low concentrations (191).
SCFAs, such as acetate, propionate and butyrate are
efficiently absorbed by the gut lumen, despite differences in their
distribution and their effects on host cell metabolism. Each SCFA
has specific characteristics that distinguish it from the other
SFCAs. Despite low concentration levels of butyrate in the systemic
circulation, IECs predominantly use butyrate to fuel their energy
stores (60-70%). Normal colonocytes exploit butyrate as their
primary energy source, as butyrate follows the procedure of
mitochondrial β-oxidation every 7 days (192-194).
With respect to the distribution of other SCFAs, the liver has the
greatest metabolizing capacity of propionate, while most of the
peripheral blood is occupied by high concentrations of acetate
(0.10-0.15 mM) (195).
Butyrate and niacin also constitute main
representative metabolites of microbiota-secreted SCFAs that have
been shown to influence the immune system through two opposing
mechanisms. From one perspective, microbial metabolites may mediate
their action through binding to the GPR109A receptor, thereby
triggering IL-18 synthesis in IECs and affecting DCs, macrophages
and T cells (211). Form another
perspective, gut microbial metabolites may exert anti-inflammatory
properties, supporting Treg differentiation and expansion, thus
establishing an immunosuppressive micro-environment (201).
Importantly, the gut microbiota is significantly
implicated in metabolizing certain food supplements and nutrients.
For example, berries and nuts involve ellagic acid, which is
converted to urolithins by gut microbiota. Urolithins diminish Cox2
levels, thus exhibiting a certain anti-cancer effect (212,213). Daidzen is another nutrient
metabolized by gut sulfate-reducing microbiota into equols
(214,215). Epidemiological data from Asian
populations have reported an association between high urinary or
plasma equol concentrations and a decreased breast and prostate
cancer risk (216). Another
characteristic example is the elimination by certain gut microbiota
(Lactobacilli and Bifidobacteria) of linoleic acid levels, which
are regarded very toxic as they convert omega-6 to omega-3 and
produce prostaglandins (217).
Finally, resveratrol constitutes another example of the
metabolizing effect mediated by gut microbiota (218).
In contrast to the above, the gut microbiota may
promote carcinogenesis through the synthesis of secondary bile
acids. Characteristically, a minor portion of primary bile acids
(5%) escapes the classical enterohepatic circulation and reaches
the colon. The following procedure deconjugates and transforms
primary bile acids into secondary bile acids (such as DCA and LCA)
though the action of specific bacteria (219). The presence of mutations that
are insensitive to apoptosis enables secondary bile salts to act as
promoters of tumorigenesis (220). DCA is such a toxic metabolite,
provoking epithelial DNA damage and apoptosis in a p53-independent
but PKC-ERK1/2-dependent manner, with direct associations to the
formation of colon cancer or hepatocellular carcinoma or esophageal
cancer (94,221-225).
Recent data illustrate that bacteria in Clostridium cluster IX are
responsible for enrichment of DCA levels in obese mice, rendering
them highly susceptible to cancer formation (144). Similarly, certain types of
microbiota that convert ethanol into acetaldehyde have been
regarded as major stimulators of carcinogenesis (191).
Last but not least, the cumulative exposure of
humans to xenobiotics or pharmaceuticals has helped in the
understanding that gut microbiota may have direct or indirect
implications in the breakdown of such substances (226).
Significant efforts are being made in order to
manipulate gut microbiota for preventive, diagnostic and
therapeutic purposes. For the diagnostic purposes, identification
of specific bacterial strains can offer enormous benefit in the
context of new, reliable, non-invasive biomarkers for cancer. For
cancer prevention purposes, Fusobacterium nucleatum has been proven
to be a valuable prognostic biomarker, if one considers the
abundance of Fusobacterium nucleatum in patients with high-grade
colon cancer and adenomas (227,228). This has been supported by
elevated fecal levels of Fusobacterium nucleatum in patients with
colorectal cancer (120,227,229,230). Notably, recent evidence has
suggested that the percentage of Fusobacterium nucleatum present in
fecal samples is inversely associated with the survival of patients
with colon cancer (231).
A subset of microbes have also been shown to reduce
chronic inflammation or to mitigate malignant transformation
(132,232,233). Below, we discuss some potential
avenues through which microbiota can be therapeutically
exploited.
First of all, the gut microbiota appears to be
essential to the effectiveness of classical chemotherapeutic drugs
such as oxiplatin, cisplatin, and cyclophosphamide (CTX). In
general, chemotherapeutic compounds elicit toxic effects on tumor
cells, including ROS activation by myeloid cells, intrinsic
mitochondrial apoptosis, and the stimulation of inflammatory genes
(132,234,235). The beneficial contribution of
the microbiota to chemotherapy has been determined by the
composition of the microbiota on myeloid cells (24,236). Similarly, microbiota located on
myeloid cells has been found to exert a positive effect on cancer
immunotherapy or total body irradiation (TBI). Therefore, the
importance of the gut microbiota in cancer therapy emerges from its
interaction with anti-neoplastic agents in a bidirectional
manner.
On the one hand, many current anti-cancer
therapeutic strategies (chemotherapy and radiation therapy)
negatively affect microbial composition, by fostering dysbiosis
(24,236). Radiation therapy, allogeneic
stem cell transplantation and several chemotherapeutic agents,
including irinotecan and 5-fluorouracil, appear to negatively
affect the composition of the gut microbiota (237-239).
On the other hand, a considerable body of evidence has demonstrated
that the gut microbiota is of the utmost importance to the efficacy
of therapeutic drugs, by eliminating side-effects and by
interfering in a pharmacodynamic or immunological manner (132,234,240,241).
Currently, therapeutic interventions based on the
gut microbiota are categorized as follows: i) Antibiotics; ii)
probiotics; ii) prebiotics; and iv) postbiotics. Each therapeutic
perspective of the gut microbiota is distinct. Antibiotics are
usually used for the eradication of specific bacterial strains.
Probiotics are living bacteria and prebiotics are non-digestible
compounds, both of which provide strong support to the host.
Postbiotics are non-viable products of microbiota, recapitulating a
wide range of functions in the human body. All of these therapeutic
categories have been shown to confer significant benefits to the
host.
Probiotics and prebiotics are known for their
capacity to sustain a balanced microbial community, obviating
pro-inflammatory or signaling pathways that lead to carcinogenesis
(5-8,11,242,243).
Probiotics are usually administered as a curative strategy for
antibiotic-mediated dysbiosis and side-effects in studies with mice
and humans (244). Probiotics
are innocuous microbes, critical for homeostasis, preventing entry
of pathogens by stimulating AMPS, IgA and contributing to
intestinal barrier integrity (243,245,246). A number of studies have proposed
probiotics as a preventive intervention for inflammatory bowel
disease or ulcerative colitis (247-251).
Notably, the chemo-preventive efficacy of
probiotics and prebiotics seems to be higher than that elicited by
antibiotics, through alleviation of inflammation. Antibiotics are
not only insufficient as chemopreventive agents, but they can also
eliminate commensal homeostatic bacteria and make certain bacterial
strains resistant (252).
However, human microbiome reconstitution following antibiotic
treatment is defective due to impairment of commensal microbial
community (253-255),
and a time-consuming process (256). For this reason, efforts have
focused on devising therapeutic strategies which sustain the
microbial composition and population, thereby conferring benefit to
the host. Based on data derived from 20 studies, 36% of patients
with IBD who were transplanted fecal-derived microbiota from
healthy donors exhibited an alleviation of symptoms (257). In another case, fecal microbiota
transplantation has been shown to alleviate diarrhea symptoms in
individuals with severe Clostridium difficile infections, following
the use of antibiotics (258).
The most impressive results were derived from the study by Suez et
al, who demonstrated that autologous fecal microbiome
transplantation was able to reconstitute the microbial community in
its initial configuration in both murine and human samples
following treatment with antibiotics. The rapid and complete
recovery of the microbiome niche in aFMT-samples following the use
of antibiotics, as compared to incomplete niche following treatment
with probiotics, was compelling (259).
Mounting evidence suggests that the microbiota can
be a determinant factor in modulating the host immune response.
Several studies have demonstrated the crucial role of the
microbiota in the response of distinct cancer types to classical
immunotherapy (immune checkpoint inhibitors) (260-264).
For example, the gut microbiota can positively influence the
effectiveness of recently developed immunotherapeutic molecules
[cytotoxic T lymphocyte associated protein 4 (CTLA4) or programmed
death protein 1 (PD-1) antibodies]. The effects of germ-free state
or the effects of colonization with specific bacterial strains on
therapies using immune checkpoint inhibitors have been
investigated. Bacteroides spp. appears to be necessary in the
anti-CTLA treatment against sarcomas (265) and Bifidobacterium seems to be
essential in anti-PDL1 therapy against melanoma (266). Furthermore, the microbiota seem
to elicit an efficient response to immunotherapy
[CpG-oligodeoxynucleotides (ODN) with neutralization antibody
against IL-10], as indicated by experiments using mice (132). Similarly, mice grown under
conventional conditions have exhibited stronger responses to
CpG-ODN than TLR4-deficient mice (132).
Additional research efforts are required in order
to elucidate the mechanisms through which the gut microbiota
modulates the clinical effectiveness of various drugs, thus
facilitating the design of appropriate personalized therapies based
on the microbiota profile of an individual patient. The binding of
the Fap2 protein of Fusobacterium nucleatum to the Ig and ITIM
domains (TIGIT) of the human inhibitory receptor that is present on
NK cells protects tumors from an immune attack by NK cells
(127). Therefore, the presence
of Fusobacterium nucleatum in patients may be a direct determinant
and/or predictor of resistance to immunotherapy, and special
considerations will have to be taken into account when designing
personalized therapies for this particular patient group.
Certain issues will also have to be addressed
before we move forward in the design of more personalized
therapies. For example, the gut microbiota can be directed towards
a specific immune population so as to serve as a tool for enriching
the specific immune population against cancer. A better
understanding of the microbiome effect on anti-PD1 therapy,
currently applied to various types of cancer, may help to address
the question (262-264).
Another issue may be whether host T cells can be equipped with a
TCR that is specific for a bacterial epitope and thereby to
orchestrate an appropriate immune response. The ultimate goal will
be to use microbiota or microbiota-derived molecules as novel
immunotherapeutic approaches that will spare patients from the
side-effects associated with systemic immunotherapies. For example,
an organized and enriched CD8+ T cell response already looks
promising in effectively enhancing the therapeutic action of immune
checkpoint inhibitors in melanoma without adverse effects (262).
A considerable body of evidence exists nowadays
that supports how essential the microbiota is in deciding the fate
of neoplastic formations, their progression and their sensitivity
to classical therapeutic drugs. The effect of the microbiota on
cancer is usually elicited locally but it can also be developed
systemically, through alterations in the whole immunological
milieu. The knowledge pertaining to the microbiome expands rapidly,
however therapeutic interventions of intestinal carcinogenesis are
still limited. Further experiments will be critical in
understanding the underlying molecular mechanisms of microbiota,
using animal models or epidemiological data derived from clinical
trials towards inventing new treatments. Nevertheless, the
available methodologies need to incorporate new technologies in
order to facilitate the growth of microbes in conditions that are a
direct replica to those within the gastrointestinal tract of the
human body. The combination of metagenomics (effect of diet on
microbiota), metaproteomics (microbial gene expression), and
metabolomics (microbial metabolites) seems to play an important
role in developing strategies for disease prevention.
Not applicable.
No funding was received.
Not applicable.
All authors (SB, MA, DAS, AMK, IC and VZ) were
involved in the design and conception of the study and have revised
and approved the final manuscript. SB performed the literature
search, has written the manuscript, has critically analyzed the
existing knowledge and has designed the picture and the tables. MA
contributed to editing the manuscript concerning the role of
microbiota in metabolism and role of dysbiotic microbiota in
inflammation and colorectal cancer, AMK contributed to editing of
manuscript concerning the role of microbiota in genotoxic stress,
IC contributed to editing the manuscript concerning the role of
microbiota in homeostasis, DA contributed to editing of manuscript
concerning association of cancer therapy with microbiota. SB, MA,
DAS, AMK, IC and VZ were significantly involved in the drafting of
the manuscript. All authors have taken the responsibility for
publishing this review paper and all authors have read and approved
the final manuscript.
Not applicable.
Not applicable.
DAS is the Editor-in-Chief for the journal, but had
no personal involvement in the reviewing process, or any influence
in terms of adjudicating on the final decision, for this article.
All the other authors do not have any competing interests to
declare.
|
1
|
Iida N, Dzutsev A, Stewart CA, Smith L,
Bouladoux N, Weingarten RA, Molina DA, Salcedo R, Back T, Cramer S,
et al: Commensal bacteria control cancer response to therapy by
modulating the tumor microenvironment. Science. 342:967–970.
2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Boleij A, Hechenbleikner EM, Goodwin AC,
Badani R, Stein EM, Lazarev MG, Ellis B, Carroll KC, Albesiano E,
Wick EC, et al: The Bacteroides fragilis toxin gene is prevalent in
the colon mucosa of colorectal cancer patients. Clin Infect Dis.
60:208–215. 2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Qin J, Li R, Raes J, Arumugam M, Burgdorf
KS, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T, et al:
MetaHIT Consortium: A human gut microbial gene catalogue
established by metagenomic sequencing. Nature. 464:59–65.
2010.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Blaser MJ: Who are we? Indigenous microbes
and the ecology of human diseases. EMBO Rep. 7:956–960.
2006.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Bultman SJ: Emerging roles of the
microbiome in cancer. Carcinogenesis. 35:249–255. 2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Clemente JC, Ursell LK and Parfrey LW and
Knight R: The impact of the gut microbiota on human health: An
integrative view. Cell. 148:1258–1270. 2012.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Plottel CS and Blaser MJ: Microbiome and
malignancy. Cell Host Microbe. 10:324–335. 2011.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Petersen C and Round JL: Defining
dysbiosis and its influence on host immunity and disease. Cell
Microbiol. 16:1024–1033. 2014.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Dietert RR and Dietert JM: The microbiome
and sustainable healthcare. Healthcare (Basel). 3:100–129.
2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Arslan N: Obesity, fatty liver disease and
intestinal microbiota. World J Gastroenterol. 20:16452–16463.
2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Schwabe RF and Jobin C: The microbiome and
cancer. Nat Rev Cancer. 13:800–812. 2013.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Garrett WS: Cancer and the microbiota.
Science. 348:80–86. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Human Microbiome Project Consortium:
Structure, function and diversity of the healthy human microbiome.
Nature. 486:207–214. 2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Grice EA and Segre JA: The skin
microbiome. Nat Rev Microbiol. 9:244–253. 2011.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Suau A, Bonnet R, Sutren M, Godon JJ,
Gibson GR, Collins MD and Doré J: Direct analysis of genes encoding
16S rRNA from complex communities reveals many novel molecular
species within the human gut. Appl Environ Microbiol. 65:4799–4807.
1999.PubMed/NCBI
|
|
16
|
Savage DC: Microbial ecology of the
gastrointestinal tract. Annu Rev Microbiol. 31:107–133.
1977.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Neish AS: Microbes in gastrointestinal
health and disease. Gastroenterology. 136:65–80. 2009.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Goncharova GI, Dorofeĭchuk VG,
Smolianskaia AZ and Sokolova KIa: Microbial ecology of the
intestines in health and in pathology. Antibiot Khimioter.
34:462–466. 1989.(In Russian). PubMed/NCBI
|
|
19
|
Dominguez-Bello MG, Blaser MJ, Ley RE and
Knight R: Development of the human gastrointestinal microbiota and
insights from high-throughput sequencing. Gastroenterology.
140:1713–1719. 2011.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Mulder IE, Schmidt B, Lewis M, Delday M,
Stokes CR, Bailey M, Aminov RI, Gill BP, Pluske JR, Mayer CD, et
al: Restricting microbial exposure in early life negates the immune
benefits associated with gut colonization in environments of high
microbial diversity. PLoS One. 6(e28279)2011.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Claesson MJ, Cusack S, O'Sullivan O,
Greene-Diniz R, de Weerd H, Flannery E, Marchesi JR, Falush D,
Dinan T, Fitzgerald G, et al: Composition, variability, and
temporal stability of the intestinal microbiota of the elderly.
Proc Natl Acad Sci USA. 108((Suppl 1)): 4586–4591. 2011.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Rajilić-Stojanović M, Heilig HGHJ,
Molenaar D, Kajander K, Surakka A, Smidt H and de Vos WM:
Development and application of the human intestinal tract chip, a
phylogenetic microarray: Analysis of universally conserved
phylotypes in the abundant microbiota of young and elderly adults.
Environ Microbiol. 11:1736–1751. 2009.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Turnbaugh PJ, Bäckhed F, Fulton L and
Gordon JI: Diet-induced obesity is linked to marked but reversible
alterations in the mouse distal gut microbiome. Cell Host Microbe.
3:213–223. 2008.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zwielehner J, Liszt K, Handschur M, Lassl
C, Lapin A and Haslberger AG: Combined PCR-DGGE fingerprinting and
quantitative-PCR indicates shifts in fecal population sizes and
diversity of Bacteroides, bifidobacteria and Clostridium cluster IV
in institutionalized elderly. Exp Gerontol. 44:440–446.
2009.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Eckburg PB, Bik EM, Bernstein CN, Purdom
E, Dethlefsen L, Sargent M, Gill SR, Nelson KE and Relman DA:
Diversity of the human intestinal microbial flora. Science.
308:1635–1638. 2005.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Sommer F and Bäckhed F: The gut microbiota
- masters of host development and physiology. Nat Rev Microbiol.
11:227–238. 2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Bäckhed F, Ley RE, Sonnenburg JL, Peterson
DA and Gordon JI: Host-bacterial mutualism in the human intestine.
Science. 307:1915–1920. 2005.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Tlaskalová-Hogenová H, Stepánková R,
Hudcovic T, Tucková L, Cukrowska B, Lodinová-Zádníková R, Kozáková
H, Rossmann P, Bártová J, Sokol D, et al: Commensal bacteria
(normal microflora), mucosal immunity and chronic inflammatory and
autoimmune diseases. Immunol Lett. 93:97–108. 2004.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wang L, Zhao N, Zhang F, Yue W and Liang
M: Effect of taurine on leucocyte function. Eur J Pharmacol.
616:275–280. 2009.PubMed/NCBI View Article : Google Scholar
|
|
30
|
O'Hara AM and Shanahan F: The gut flora as
a forgotten organ. EMBO Rep. 7:688–693. 2006.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Proctor LM: The human microbiome project
in 2011 and beyond. Cell Host Microbe. 10:287–291. 2011.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Weisburger JH, Reddy BS, Narisawa T and
Wynder EL: Germ-free status and colon tumor induction by
N-methyl-N'-nitro-N-nitrosoguanidine. Proc Soc Exp Biol Med.
148:1119–1121. 1975.PubMed/NCBI
|
|
33
|
Peterson DA, Frank DN, Pace NR and Gordon
JI: Metagenomic approaches for defining the pathogenesis of
inflammatory bowel diseases. Cell Host Microbe. 3:417–427.
2008.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Walsh CJ, Guinane CM, O'Toole PW and
Cotter PD: Beneficial modulation of the gut microbiota. FEBS Lett.
588:4120–4130. 2014.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Kamada N and Núñez G: Role of the gut
microbiota in the development and function of lymphoid cells. J
Immunol. 190:1389–1395. 2013.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Blaser MJ and Falkow S: What are the
consequences of the disappearing human microbiota? Nat Rev
Microbiol. 7:887–894. 2009.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Hooper LV and Macpherson AJ: Immune
adaptations that maintain homeostasis with the intestinal
microbiota. Nat Rev Immunol. 10:159–169. 2010.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Palmer C, Bik EM, DiGiulio DB, Relman DA
and Brown PO: Development of the human infant intestinal
microbiota. PLoS Biol. 5(e177)2007.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Neish AS: Mucosal immunity the microbiome.
Ann Am Thorac Soc. 11((Suppl 1)): S28–S32. 2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Lozupone CA, Stombaugh JI, Gordon JI,
Jansson JK and Knight R: Diversity, stability and resilience of the
human gut microbiota. Nature. 489:220–230. 2012.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Morgan XC and Huttenhower C: Chapter 12:
Human microbiome analysis. PLOS Comput Biol.
8(e1002808)2012.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Hooper LV, Littman DR and Macpherson AJ:
Interactions between the microbiota and the immune system. Science.
336:1268–1273. 2012.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Khoruts A, Dicksved J, Jansson JK and
Sadowsky MJ: Changes in the composition of the human fecal
microbiome after bacteriotherapy for recurrent Clostridium
difficile-associated diarrhea. J Clin Gastroenterol. 44:354–360.
2010.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Reid G, Younes JA, Van der Mei HC, Gloor
GB, Knight R and Busscher HJ: Microbiota restoration: Natural and
supplemented recovery of human microbial communities. Nat Rev
Microbiol. 9:27–38. 2011.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Swiatczak B, Rescigno M and Cohen IR:
Systemic features of immune recognition in the gut. Microbes
Infect. 13:983–991. 2011.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Arthur JC and Jobin C: The struggle within
Microbial influences on colorectal cancer. Inflamm Bowel Dis.
17:396–409. 2011.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Ley RE, Bäckhed F, Turnbaugh P, Lozupone
CA, Knight RD and Gordon JI: Obesity alters gut microbial ecology.
Proc Natl Acad Sci USA. 102:11070–11075. 2005.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Ivanov II, Frutos R de L, Manel N,
Yoshinaga K, Rifkin DB, Sartor RB, Finlay BB and Littman DR:
Specific microbiota direct the differentiation of IL-17-producing
T-helper cells in the mucosa of the small intestine. Cell Host
Microbe. 4:337–349. 2008.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Johansson MEV, Jakobsson HE,
Holmén-Larsson J, Schütte A, Ermund A, Rodríguez-Piñeiro AM, Arike
L, Wising C, Svensson F, Bäckhed F, et al: Normalization of host
intestinal mucus layers requires long-term microbial colonization.
Cell Host Microbe. 18:582–592. 2015.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Spiljar M, Merkler D and Trajkovski M: The
immune system bridges the gut microbiota with systemic energy
homeostasis: focus on TLRs, mucosal barrier, and SCFAs. Front
Immunol. 8(1353)2017.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Round JL and Mazmanian SK: Inducible
Foxp3+ regulatory T-cell development by a commensal bacterium of
the intestinal microbiota. Proc Natl Acad Sci USA. 107:12204–12209.
2010.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Hepworth MR, Monticelli LA, Fung TC,
Ziegler CG, Grunberg S, Sinha R, Mantegazza AR, Ma HL, Crawford A,
Angelosanto JM, et al: Innate lymphoid cells regulate CD4+ T-cell
responses to intestinal commensal bacteria. Nature. 498:113–117.
2013.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Gaboriau-Routhiau V, Rakotobe S, Lécuyer
E, Mulder I, Lan A, Bridonneau C, Rochet V, Pisi A, De Paepe M,
Brandi G, et al: The key role of segmented filamentous bacteria in
the coordinated maturation of gut helper T cell responses.
Immunity. 31:677–689. 2009.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Garland CD, Lee A and Dickson MR:
Segmented filamentous bacteria in the rodent small intestine: Their
colonization of growing animals and possible role in host
resistance to Salmonella. Microb Ecol. 8:181–190. 1982.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Yang Y, Torchinsky MB, Gobert M, Xiong H,
Xu M, Linehan JL, Alonzo F, Ng C, Chen A, Lin X, et al: Focused
specificity of intestinal TH17 cells towards commensal bacterial
antigens. Nature. 510:152–156. 2014.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Schnupf P, Gaboriau-Routhiau V and
Cerf-Bensussan N: Host interactions with segmented filamentous
bacteria: An unusual trade-off that drives the post-natal
maturation of the gut immune system. Semin Immunol. 25:342–351.
2013.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Wu HJ, Ivanov II, Darce J, Hattori K,
Shima T, Umesaki Y, Littman DR, Benoist C and Mathis D:
Gut-residing segmented filamentous bacteria drive autoimmune
arthritis via T helper 17 cells. Immunity. 32:815–827.
2010.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Song X, Gao H, Lin Y, Yao Y, Zhu S, Wang
J, Liu Y, Yao X, Meng G, Shen N, et al: Alterations in the
microbiota drive interleukin-17C production from intestinal
epithelial cells to promote tumorigenesis. Immunity. 40:140–152.
2014.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Xu M, Pokrovskii M, Ding Y, Yi R, Au C,
Harrison OJ, Galan C, Belkaid Y, Bonneau R and Littman DR:
c-MAF-dependent regulatory T cells mediate immunological tolerance
to a gut pathobiont. Nature. 554:373–377. 2018.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Kamada N and Núñez G: Regulation of the
immune system by the resident intestinal bacteria.
Gastroenterology. 146:1477–1488. 2014.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Martín R, Miquel S, Chain F, Natividad JM,
Jury J, Lu J, Sokol H, Theodorou V, Bercik P, Verdu EF, et al:
Faecalibacterium prausnitzii prevents physiological damages in a
chronic low-grade inflammation murine model. BMC Microbiol.
15(67)2015.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Honda K and Littman DR: The microbiota in
adaptive immune homeostasis and disease. Nature. 535:75–84.
2016.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Naik S, Bouladoux N, Wilhelm C, Molloy MJ,
Salcedo R, Kastenmuller W, Deming C, Quinones M, Koo L, Conlan S,
et al: Compartmentalized control of skin immunity by resident
commensals. Science. 337:1115–1119. 2012.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Naik S, Bouladoux N, Linehan JL, Han SJ,
Harrison OJ, Wilhelm C, Conlan S, Himmelfarb S, Byrd AL, Deming C,
et al: Commensal-dendritic-cell interaction specifies a unique
protective skin immune signature. Nature. 520:104–108.
2015.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Hajishengallis G, Liang S, Payne MA,
Hashim A, Jotwani R, Eskan MA, McIntosh ML, Alsam A, Kirkwood KL,
Lambris JD, et al: Low-abundance biofilm species orchestrates
inflammatory periodontal disease through the commensal microbiota
and complement. Cell Host Microbe. 10:497–506. 2011.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Hajishengallis G and Lamont RJ: Breaking
bad: Manipulation of the host response by Porphyromonas gingivalis.
Eur J Immunol. 44:328–338. 2014.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Abt MC, Osborne LC, Monticelli LA, Doering
TA, Alenghat T, Sonnenberg GF, Paley MA, Antenus M, Williams KL,
Erikson J, et al: Commensal bacteria calibrate the activation
threshold of innate antiviral immunity. Immunity. 37:158–170.
2012.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Belkaid Y and Naik S: Compartmentalized
and systemic control of tissue immunity by commensals. Nat Immunol.
14:646–653. 2013.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Ichinohe T, Pang IK, Kumamoto Y, Peaper
DR, Ho JH, Murray TS and Iwasaki A: Microbiota regulates immune
defense against respiratory tract influenza A virus infection. Proc
Natl Acad Sci USA. 108:5354–5359. 2011.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Chervonsky AV: Microbiota autoimmunity.
Cold Spring Harb Perspect Biol. 5(a007294)2013.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Lee YK, Menezes JS, Umesaki Y and
Mazmanian SK: Proinflammatory T-cell responses to gut microbiota
promote experimental autoimmune encephalomyelitis. Proc Natl Acad
Sci USA. 108((Suppl 1)): 4615–4622. 2011.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Kim YG, Udayanga KGS, Totsuka N, Weinberg
JB, Núñez G and Shibuya A: Gut dysbiosis promotes M2 macrophage
polarization and allergic airway inflammation via fungi-induced
PGE2. Cell Host Microbe. 15:95–102. 2014.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Mangeney M, Pothlichet J, Renard M, Ducos
B and Heidmann T: Endogenous retrovirus expression is required for
murine melanoma tumor growth in vivo. Cancer Res. 65:2588–2591.
2005.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Vaishnava S, Behrendt CL, Ismail AS,
Eckmann L and Hooper LV: Paneth cells directly sense gut commensals
and maintain homeostasis at the intestinal host-microbial
interface. Proc Natl Acad Sci USA. 105:20858–20863. 2008.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Ménard S, Cerf-Bensussan N and Heyman M:
Multiple facets of intestinal permeability and epithelial handling
of dietary antigens. Mucosal Immunol. 3:247–259. 2010.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Mortha A, Chudnovskiy A, Hashimoto D,
Bogunovic M, Spencer SP, Belkaid Y and Merad M:
Microbiota-dependent crosstalk between macrophages and ILC3
promotes intestinal homeostasis. Science.
343(1249288)2014.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Rescigno M: Intestinal microbiota and its
effects on the immune system. Cell Microbiol. 16:1004–1013.
2014.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics 2014. CA Cancer J Clin. 64:104–117.
2014.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Shen H, Yang J, Huang Q, Jiang MJ, Tan YN,
Fu JF, Zhu LZ, Fang XF and Yuan Y: Different treatment strategies
and molecular features between right-sided and left-sided colon
cancers. World J Gastroenterol. 21:6470–6478. 2015.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Tamas K, Walenkamp AM, de Vries EG, van
Vugt MA, Beets-Tan RG, van Etten B, de Groot DJ and Hospers GA:
Rectal and colon cancer: Not just a different anatomic site. Cancer
Treat Rev. 41:671–679. 2015.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Carethers JM and Jung BH: Genetics and
genetic biomarkers in sporadic colorectal Cancer. Gastroenterology.
149:1177–1190.e3. 2015.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Watson AJ and Collins PD: Colon cancer A
civilization disorder. Dig Dis. 29:222–228. 2011.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Starnes CO: Coley's toxins in perspective.
Nature. 357:11–12. 1992.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Hoption Cann SA, van Netten JP and van
Netten C: Dr William Coley and tumour regression: A place in
history or in the future. Postgrad Med J. 79:672–680.
2003.PubMed/NCBI
|
|
85
|
Grange JM, Bottasso O, Stanford CA and
Stanford JL: The use of mycobacterial adjuvant-based agents for
immunotherapy of cancer. Vaccine. 26:4984–4990. 2008.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Reddy BS, Mastromarino A and Wynder EL:
Further leads on metabolic epidemiology of large bowel cancer.
Cancer Res. 35:3403–3406. 1975.PubMed/NCBI
|
|
87
|
Schreiber H, Nettesheim P, Lijinsky W,
Richter CB and Walburg HE Jr: Induction of lung cancer in germfree,
specific-pathogen-free, and infected rats by
N-nitrosoheptamethyleneimine: Enhancement by respiratory infection.
J Natl Cancer Inst. 49:1107–1114. 1972.PubMed/NCBI
|
|
88
|
Dapito DH, Mencin A, Gwak GY, Pradere JP,
Jang MK, Mederacke I, Caviglia JM, Khiabanian H, Adeyemi A,
Bataller R, et al: Promotion of hepatocellular carcinoma by the
intestinal microbiota and TLR4. Cancer Cell. 21:504–516.
2012.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Reddy BS and Watanabe K: Effect of
intestinal microflora on 2,2'-dimethyl-4-aminobiphenyl-induced
carcinogenesis in F344 rats. J Natl Cancer Inst. 61:1269–1271.
1978.PubMed/NCBI
|
|
90
|
Uronis JM, Mühlbauer M, Herfarth HH,
Rubinas TC, Jones GS and Jobin C: Modulation of the intestinal
microbiota alters colitis-associated colorectal cancer
susceptibility. PLoS One. 4(e6026)2009.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Lofgren JL, Whary MT, Ge Z, Muthupalani S,
Taylor NS, Mobley M, Potter A, Varro A, Eibach D, Suerbaum S, et
al: Lack of commensal flora in Helicobacter pylori-infected INS-GAS
mice reduces gastritis and delays intraepithelial neoplasia.
Gastroenterology. 140:210–220. 2011.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Vannucci L, Stepankova R, Kozakova H,
Fiserova A, Rossmann P and Tlaskalova-Hogenova H: Colorectal
carcinogenesis in germ-free and conventionally reared rats:
Different intestinal environments affect the systemic immunity. Int
J Oncol. 32:609–617. 2008.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Dove WF, Clipson L, Gould KA, Luongo C,
Marshall DJ, Moser AR, Newton MA and Jacoby RF: Intestinal
neoplasia in the ApcMin mouse: Independence from the microbial and
natural killer (beige locus) status. Cancer Res. 57:812–814.
1997.PubMed/NCBI
|
|
94
|
Yoshimoto S, Loo TM, Atarashi K, Kanda H,
Sato S, Oyadomari S, Iwakura Y, Oshima K, Morita H, Hattori M, et
al: Obesity-induced gut microbial metabolite promotes liver cancer
through senescence secretome. Nature. 499:97–101. 2013.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Chen GY, Shaw MH, Redondo G and Núñez G:
The innate immune receptor Nod1 protects the intestine from
inflammation-induced tumorigenesis. Cancer Res. 68:10060–10067.
2008.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Grivennikov SI, Wang K, Mucida D, Stewart
CA, Schnabl B, Jauch D, Taniguchi K, Yu GY, Osterreicher CH, Hung
KE, et al: Adenoma-linked barrier defects and microbial products
drive IL-23/IL-17-mediated tumour growth. Nature. 491:254–258.
2012.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Klimesova K, Kverka M, Zakostelska Z,
Hudcovic T, Hrncir T, Stepankova R, Rossmann P, Ridl J, Kostovcik
M, Mrazek J, et al: Altered gut microbiota promotes
colitis-associated cancer in IL-1 receptor-associated kinase
M-deficient mice. Inflamm Bowel Dis. 19:1266–1277. 2013.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Garrett WS, Punit S, Gallini CA, Michaud
M, Zhang D, Sigrist KS, Lord GM, Glickman JN and Glimcher LH:
Colitis-associated colorectal cancer driven by T-bet deficiency in
dendritic cells. Cancer Cell. 16:208–219. 2009.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Kado S, Uchida K, Funabashi H, Iwata S,
Nagata Y, Ando M, Onoue M, Matsuoka Y, Ohwaki M and Morotomi M:
Intestinal microflora are necessary for development of spontaneous
adenocarcinoma of the large intestine in T-cell receptor beta chain
and p53 double-knockout mice. Cancer Res. 61:2395–2398.
2001.PubMed/NCBI
|
|
100
|
Engle SJ, Ormsby I, Pawlowski S, Boivin
GP, Croft J, Balish E and Doetschman T: Elimination of colon cancer
in germ-free transforming growth factor beta 1-deficient mice.
Cancer Res. 62:6362–6366. 2002.PubMed/NCBI
|
|
101
|
Erdman SE, Poutahidis T, Tomczak M, Rogers
AB, Cormier K, Plank B, Horwitz BH and Fox JG: CD4+ CD25+
regulatory T lymphocytes inhibit microbially induced colon cancer
in Rag2-deficient mice. Am J Pathol. 162:691–702. 2003.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Garrett WS, Lord GM, Punit S,
Lugo-Villarino G, Mazmanian SK, Ito S, Glickman JN and Glimcher LH:
Communicable ulcerative colitis induced by T-bet deficiency in the
innate immune system. Cell. 131:33–45. 2007.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Garrett WS, Gallini CA, Yatsunenko T,
Michaud M, DuBois A, Delaney ML, Punit S, Karlsson M, Bry L,
Glickman JN, et al: Enterobacteriaceae act in concert with the gut
microbiota to induce spontaneous and maternally transmitted
colitis. Cell Host Microbe. 8:292–300. 2010.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Balish E and Warner T: Enterococcus
faecalis induces inflammatory bowel disease in interleukin-10
knockout mice. Am J Pathol. 160:2253–2257. 2002.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Zhan Y, Chen PJ, Sadler WD, Wang F, Poe S,
Núñez G, Eaton KA and Chen GY: Gut microbiota protects against
gastrointestinal tumorigenesis caused by epithelial injury. Cancer
Res. 73:7199–7210. 2013.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Hussain SP, Hofseth LJ and Harris CC:
Radical causes of cancer. Nat Rev Cancer. 3:276–285.
2003.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Maddocks ODK, Short AJ, Donnenberg MS,
Bader S and Harrison DJ: Attaching and effacing Escherichia coli
downregulate DNA mismatch repair protein in vitro and are
associated with colorectal adenocarcinomas in humans. PLoS One.
4(e5517)2009.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Salcedo R, Worschech A, Cardone M, Jones
Y, Gyulai Z, Dai RM, Wang E, Ma W, Haines D, O'hUigin C, et al:
MyD88-mediated signaling prevents development of adenocarcinomas of
the colon: Role of interleukin 18. J Exp Med. 207:1625–1636.
2010.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Saleh M and Trinchieri G: Innate immune
mechanisms of colitis and colitis-associated colorectal cancer. Nat
Rev Immunol. 11:9–20. 2011.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Rakoff-Nahoum S, Paglino J,
Eslami-Varzaneh F, Edberg S and Medzhitov R: Recognition of
commensal microflora by toll-like receptors is required for
intestinal homeostasis. Cell. 118:229–241. 2004.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Sears CL and Garrett WS: Microbes,
microbiota, and colon cancer. Cell Host Microbe. 15:317–328.
2014.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Eslick GD: Helicobacter pylori infection
causes gastric cancer? A review of the epidemiological,
meta-analytic, and experimental evidence. World J Gastroenterol.
12:2991–2999. 2006.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Buti L, Spooner E, van der Veen AG,
Rappuoli R, Covacci A and Ploegh HL: Helicobacter pylori
cytotoxin-associated gene A (CagA) subverts the
apoptosis-stimulating protein of p53 (ASPP2) tumor suppressor
pathway of the host. Proc Natl Acad Sci USA. 108:9238–9243.
2011.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Wroblewski LE, Peek RM Jr and Wilson KT:
Helicobacter pylori and gastric cancer: Factors that modulate
disease risk. Clin Microbiol Rev. 23:713–739. 2010.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Marshall BJ and Warren JR: Unidentified
curved bacilli in the stomach of patients with gastritis and peptic
ulceration. Lancet. 1:1311–1315. 1984.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Wu S, Rhee KJ, Albesiano E, Rabizadeh S,
Wu X, Yen HR, Huso DL, Brancati FL, Wick E, McAllister F, et al: A
human colonic commensal promotes colon tumorigenesis via activation
of T helper type 17 T cell responses. Nat Med. 15:1016–1022.
2009.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Geis AL, Fan H, Wu X, Wu S, Huso DL, Wolfe
JL, Sears CL, Pardoll DM and Housseau F: Regulatory T-cell response
to enterotoxigenic Bacteroides fragilis colonization triggers
IL17-dependent colon carcinogenesis. Cancer Discov. 5:1098–1109.
2015.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Kim SC, Tonkonogy SL, Albright CA, Tsang
J, Balish EJ, Braun J, Huycke MM and Sartor RB: Variable phenotypes
of enterocolitis in interleukin 10-deficient mice monoassociated
with two different commensal bacteria. Gastroenterology.
128:891–906. 2005.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Wang X, Yang Y, Moore DR, Nimmo SL,
Lightfoot SA and Huycke MM: 4-Hydroxy-2-nonenal mediates
genotoxicity and bystander effects caused by Enterococcus
faecalis-infected macrophages. Gastroenterology. 142:543–551, e7.
2012.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Kostic AD, Chun E, Robertson L, Glickman
JN, Gallini CA, Michaud M, Clancy TE, Chung DC, Lochhead P, Hold
GL, et al: Fusobacterium nucleatum potentiates intestinal
tumorigenesis and modulates the tumor-immune microenvironment. Cell
Host Microbe. 14:207–215. 2013.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Abed J, Emgård JEM, Zamir G, Faroja M,
Almogy G, Grenov A, Sol A, Naor R, Pikarsky E, Atlan KA, et al:
Fap2 mediates fusobacterium nucleatum colorectal adenocarcinoma
enrichment by binding to tumor-expressed Gal-GalNAc. Cell Host
Microbe. 20:215–225. 2016.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Signat B, Roques C, Poulet P and Duffaut
D: Fusobacterium nucleatum in periodontal health and disease. Curr
Issues Mol Biol. 13:25–36. 2011.PubMed/NCBI
|
|
123
|
Strauss J, Kaplan GG, Beck PL, Rioux K,
Panaccione R, Devinney R, Lynch T and Allen-Vercoe E: Invasive
potential of gut mucosa-derived Fusobacterium nucleatum positively
correlates with IBD status of the host. Inflamm Bowel Dis.
17:1971–1978. 2011.PubMed/NCBI View Article : Google Scholar
|
|
124
|
Sobhani I, Tap J, Roudot-Thoraval F,
Roperch JP, Letulle S, Langella P, Corthier G, Tran Van Nhieu J and
Furet JP: Microbial dysbiosis in colorectal cancer (CRC) patients.
PLoS One. 6(e16393)2011.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Castellarin M, Warren RL, Freeman JD,
Dreolini L, Krzywinski M, Strauss J, Barnes R, Watson P,
Allen-Vercoe E, Moore RA, et al: Fusobacterium nucleatum infection
is prevalent in human colorectal carcinoma. Genome Res. 22:299–306.
2012.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Rubinstein MR, Wang X, Liu W, Hao Y, Cai G
and Han YW: Fusobacterium nucleatum promotes colorectal
carcinogenesis by modulating E-cadherin/β-catenin signaling via its
FadA adhesin. Cell Host Microbe. 14:195–206. 2013.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Gur C, Ibrahim Y, Isaacson B, Yamin R,
Abed J, Gamliel M, Enk J, Bar-On Y, Stanietsky-Kaynan N,
Coppenhagen-Glazer S, et al: Binding of the Fap2 protein of
Fusobacterium nucleatum to human inhibitory receptor TIGIT protects
tumors from immune cell attack. Immunity. 42:344–355.
2015.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Liu H, Redline RW and Han YW:
Fusobacterium nucleatum induces fetal death in mice via stimulation
of TLR4-mediated placental inflammatory response. J Immunol.
179:2501–2508. 2007.PubMed/NCBI View Article : Google Scholar
|
|
129
|
Lee P and Tan KS: Fusobacterium nucleatum
activates the immune response through retinoic acid-inducible gene
I. J Dent Res. 93:162–168. 2014.PubMed/NCBI View Article : Google Scholar
|
|
130
|
Chaushu S, Wilensky A, Gur C, Shapira L,
Elboim M, Halftek G, Polak D, Achdout H, Bachrach G and Mandelboim
O: Direct recognition of Fusobacterium nucleatum by the NK cell
natural cytotoxicity receptor NKp46 aggravates periodontal disease.
PLoS Pathog. 8(e1002601)2012.PubMed/NCBI View Article : Google Scholar
|
|
131
|
Huycke MM and Gaskins HR: Commensal
bacteria, redox stress, and colorectal cancer Mechanisms and
models. Exp Biol Med (Maywood). 229:586–597. 2004.PubMed/NCBI
|
|
132
|
Rooks MG and Garrett WS: Bacteria, food,
and cancer. F1000 Biol Rep 3. 12:2011.PubMed/NCBI View
Article : Google Scholar
|
|
133
|
Goodwin AC, Destefano Shields CE, Wu S,
Huso DL, Wu X, Murray-Stewart TR, Hacker-Prietz A, Rabizadeh S,
Woster PM, Sears CL, et al: Polyamine catabolism contributes to
enterotoxigenic Bacteroides fragilis-induced colon tumorigenesis.
Proc Natl Acad Sci USA. 108:15354–15359. 2011.PubMed/NCBI View Article : Google Scholar
|
|
134
|
Arthur JC, Perez-Chanona E, Mühlbauer M,
Tomkovich S, Uronis JM, Fan TJ, Campbell BJ, Abujamel T, Dogan B,
Rogers AB, et al: Intestinal inflammation targets cancer-inducing
activity of the microbiota. Science. 338:120–123. 2012.PubMed/NCBI View Article : Google Scholar
|
|
135
|
Cougnoux A, Dalmasso G, Martinez R, Buc E,
Delmas J, Gibold L, Sauvanet P, Darcha C, Déchelotte P, Bonnet M,
et al: Bacterial genotoxin colibactin promotes colon tumour growth
by inducing a senescence-associated secretory phenotype. Gut.
63:1932–1942. 2014.PubMed/NCBI View Article : Google Scholar
|
|
136
|
Mangerich A, Knutson CG, Parry NM,
Muthupalani S, Ye W, Prestwich E, Cui L, McFaline JL, Mobley M, Ge
Z, et al: Infection-induced colitis in mice causes dynamic and
tissue-specific changes in stress response and DNA damage leading
to colon cancer. Proc Natl Acad Sci USA. 109:E1820–E1829.
2012.PubMed/NCBI View Article : Google Scholar
|
|
137
|
Wang X, Allen TD, May RJ, Lightfoot S,
Houchen CW and Huycke MM: Enterococcus faecalis induces aneuploidy
and tetraploidy in colonic epithelial cells through a bystander
effect. Cancer Res. 68:9909–9917. 2008.PubMed/NCBI View Article : Google Scholar
|
|
138
|
Buc E, Dubois D, Sauvanet P, Raisch J,
Delmas J, Darfeuille-Michaud A, Pezet D and Bonnet R: High
prevalence of mucosa-associated Ecoli producing cyclomodulin and
genotoxin in colon cancer. PLoS One. 8(e56964)2013.PubMed/NCBI View Article : Google Scholar
|
|
139
|
Cuevas-Ramos G, Petit CR, Marcq I, Boury
M, Oswald E and Nougayrède JP: Escherichia coli induces DNA damage
in vivo and triggers genomic instability in mammalian cells. Proc
Natl Acad Sci USA. 107:11537–11542. 2010.PubMed/NCBI View Article : Google Scholar
|
|
140
|
Devkota S, Wang Y, Musch MW, Leone V,
Fehlner-Peach H, Nadimpalli A, Antonopoulos DA, Jabri B and Chang
EB: Dietary-fat-induced taurocholic acid promotes pathobiont
expansion and colitis in IL10-/- mice. Nature. 487:104–108.
2012.PubMed/NCBI View Article : Google Scholar
|
|
141
|
Maggio-Price L, Treuting P, Zeng W, Tsang
M, Bielefeldt-Ohmann H and Iritani BM: Helicobacter infection is
required for inflammation and colon cancer in SMAD3-deficient mice.
Cancer Res. 66:828–838. 2006.PubMed/NCBI View Article : Google Scholar
|
|
142
|
Chu F-F, Esworthy RS, Chu PG, Longmate JA,
Huycke MM, Wilczynski S and Doroshow JH: Bacteria-induced
intestinal cancer in mice with disrupted Gpx1 and Gpx2 genes.
Cancer Res. 64:962–968. 2004.PubMed/NCBI View Article : Google Scholar
|
|
143
|
Ellmerich S, Schöller M, Duranton B, Gossé
F, Galluser M, Klein JP and Raul F: Promotion of intestinal
carcinogenesis by Streptococcus bovis. Carcinogenesis. 21:753–756.
2000.PubMed/NCBI
|
|
144
|
Sheflin AM, Whitney AK and Weir TL:
Cancer-promoting effects of microbial dysbiosis. Curr Oncol Rep.
16(406)2014.PubMed/NCBI View Article : Google Scholar
|
|
145
|
Jobin C: Colorectal cancer: CRC - all
about microbial products and barrier function? Nat Rev
Gastroenterol Hepatol. 9:694–696. 2012.PubMed/NCBI View Article : Google Scholar
|
|
146
|
Rao VP, Poutahidis T, Ge Z, Nambiar PR,
Boussahmain C, Wang YY, Horwitz BH, Fox JG and Erdman SE: Innate
immune inflammatory response against enteric bacteria Helicobacter
hepaticus induces mammary adenocarcinoma in mice. Cancer Res.
66:7395–7400. 2006.PubMed/NCBI View Article : Google Scholar
|
|
147
|
Reddy BS, Weisburger JH, Narisawa T and
Wynder EL: Colon carcinogenesis in germ-free rats with
1,2-dimethylhydrazine and N-methyl-n'-nitro-N-nitrosoguanidine.
Cancer Res. 34:2368–2372. 1974.PubMed/NCBI
|
|
148
|
Reddy BS, Narisawa T, Wright P, Vukusich
D, Weisburger JH and Wynder EL: Colon carcinogenesis with
azoxymethane and dimethylhydrazine in germ-free rats. Cancer Res.
35:287–290. 1975.PubMed/NCBI
|
|
149
|
Winter SE, Lopez CA and Bäumler AJ: The
dynamics of gut-associated microbial communities during
inflammation. EMBO Rep. 14:319–327. 2013.PubMed/NCBI View Article : Google Scholar
|
|
150
|
Allen IC, TeKippe EM, Woodford RM, Uronis
JM, Holl EK, Rogers AB, Herfarth HH, Jobin C and Ting JP: The NLRP3
inflammasome functions as a negative regulator of tumorigenesis
during colitis-associated cancer. J Exp Med. 207:1045–1056.
2010.PubMed/NCBI View Article : Google Scholar
|
|
151
|
Elinav E, Strowig T, Kau AL, Henao-Mejia
J, Thaiss CA, Booth CJ, Peaper DR, Bertin J, Eisenbarth SC, Gordon
JI, et al: NLRP6 inflammasome regulates colonic microbial ecology
and risk for colitis. Cell. 145:745–757. 2011.PubMed/NCBI View Article : Google Scholar
|
|
152
|
Couturier-Maillard A, Secher T, Rehman A,
Normand S, De Arcangelis A, Haesler R, Huot L, Grandjean T,
Bressenot A, Delanoye-Crespin A, et al: NOD2-mediated dysbiosis
predisposes mice to transmissible colitis and colorectal cancer. J
Clin Invest. 123:700–711. 2013.PubMed/NCBI View Article : Google Scholar
|
|
153
|
Hu B, Elinav E, Huber S, Strowig T, Hao L,
Hafemann A, Jin C, Wunderlich C, Wunderlich T, Eisenbarth SC, et
al: Microbiota-induced activation of epithelial IL-6 signaling
links inflammasome-driven inflammation with transmissible cancer.
Proc Natl Acad Sci USA. 110:9862–9867. 2013.PubMed/NCBI View Article : Google Scholar
|
|
154
|
Velcich A, Yang W, Heyer J, Fragale A,
Nicholas C, Viani S, Kucherlapati R, Lipkin M, Yang K and
Augenlicht L: Colorectal cancer in mice genetically deficient in
the mucin Muc2. Science. 295:1726–1729. 2002.PubMed/NCBI View Article : Google Scholar
|
|
155
|
Ochi A, Nguyen AH, Bedrosian AS, Mushlin
HM, Zarbakhsh S, Barilla R, Zambirinis CP, Fallon NC, Rehman A,
Pylayeva-Gupta Y, et al: MyD88 inhibition amplifies dendritic cell
capacity to promote pancreatic carcinogenesis via Th2 cells. J Exp
Med. 209:1671–1687. 2012.PubMed/NCBI View Article : Google Scholar
|
|
156
|
Michaud DS, Joshipura K, Giovannucci E and
Fuchs CS: A prospective study of periodontal disease and pancreatic
cancer in US male health professionals. J Natl Cancer Inst.
99:171–175. 2007.PubMed/NCBI View Article : Google Scholar
|
|
157
|
Farrell JJ, Zhang L, Zhou H, Chia D,
Elashoff D, Akin D, Paster BJ, Joshipura K and Wong DT: Variations
of oral microbiota are associated with pancreatic diseases
including pancreatic cancer. Gut. 61:582–588. 2012.PubMed/NCBI View Article : Google Scholar
|
|
158
|
Wiest R and Garcia-Tsao G: Bacterial
translocation (BT) in cirrhosis. Hepatology. 41:422–433.
2005.PubMed/NCBI View Article : Google Scholar
|
|
159
|
Seki E, De Minicis S, Osterreicher CH,
Kluwe J, Osawa Y, Brenner DA and Schwabe RF: TLR4 enhances TGF-beta
signaling and hepatic fibrosis. Nat Med. 13:1324–1332.
2007.PubMed/NCBI View
Article : Google Scholar
|
|
160
|
Yu LX, Yan HX, Liu Q, Yang W, Wu HP, Dong
W, Tang L, Lin Y, He YQ, Zou SS, et al: Endotoxin accumulation
prevents carcinogen-induced apoptosis and promotes liver
tumorigenesis in rodents. Hepatology. 52:1322–1333. 2010.PubMed/NCBI View Article : Google Scholar
|
|
161
|
Xuan C, Shamonki JM, Chung A, Dinome ML,
Chung M, Sieling PA and Lee DJ: Microbial dysbiosis is associated
with human breast cancer. PLoS One. 9(e83744)2014.PubMed/NCBI View Article : Google Scholar
|
|
162
|
Velicer CM, Heckbert SR, Lampe JW, Potter
JD, Robertson CA and Taplin SH: Antibiotic use in relation to the
risk of breast cancer. JAMA. 291:827–835. 2004.PubMed/NCBI View Article : Google Scholar
|
|
163
|
Wu S, Powell J, Mathioudakis N, Kane S,
Fernandez E and Sears CL: Bacteroides fragilis enterotoxin induces
intestinal epithelial cell secretion of interleukin-8 through
mitogen-activated protein kinases and a tyrosine kinase-regulated
nuclear factor-kappaB pathway. Infect Immun. 72:5832–5839.
2004.PubMed/NCBI View Article : Google Scholar
|
|
164
|
Nougayrède JP, Taieb F, De Rycke J and
Oswald E: Cyclomodulins: Bacterial effectors that modulate the
eukaryotic cell cycle. Trends Microbiol. 13:103–110.
2005.PubMed/NCBI View Article : Google Scholar
|
|
165
|
Nesić D, Hsu Y and Stebbins CE: Assembly
and function of a bacterial genotoxin. Nature. 429:429–433.
2004.PubMed/NCBI View Article : Google Scholar
|
|
166
|
Oswald E, Nougayrède JP, Taieb F and Sugai
M: Bacterial toxins that modulate host cell-cycle progression. Curr
Opin Microbiol. 8:83–91. 2005.PubMed/NCBI View Article : Google Scholar
|
|
167
|
Travaglione S, Fabbri A and Fiorentini C:
The Rho-activating CNF1 toxin from pathogenic E. coli: A risk
factor for human cancer development? Infect Agent Cancer.
3(4)2008.PubMed/NCBI View Article : Google Scholar
|
|
168
|
Fox JG and Wang TC: Inflammation, atrophy,
and gastric cancer. J Clin Invest. 117:60–69. 2007.PubMed/NCBI View Article : Google Scholar
|
|
169
|
Ohnishi N, Yuasa H, Tanaka S, Sawa H,
Miura M, Matsui A, Higashi H, Musashi M, Iwabuchi K, Suzuki M, et
al: Transgenic expression of Helicobacter pylori CagA induces
gastrointestinal and hematopoietic neoplasms in mouse. Proc Natl
Acad Sci USA. 105:1003–1008. 2008.PubMed/NCBI View Article : Google Scholar
|
|
170
|
Sears CL: Enterotoxigenic Bacteroides
fragilis: A rogue among symbiotes. Clin Microbiol Rev. 22:349–369.
2009.PubMed/NCBI View Article : Google Scholar
|
|
171
|
Wu S, Lim KC, Huang J, Saidi RF and Sears
CL: Bacteroides fragilis enterotoxin cleaves the zonula adherens
protein, E-cadherin. Proc Natl Acad Sci USA. 95:14979–14984.
1998.PubMed/NCBI
|
|
172
|
Wu S, Shin J, Zhang G, Cohen M, Franco A
and Sears CL: The Bacteroides fragilis toxin binds to a specific
intestinal epithelial cell receptor. Infect Immun. 74:5382–5390.
2006.PubMed/NCBI View Article : Google Scholar
|
|
173
|
Toprak NU, Yagci A, Gulluoglu BM, Akin ML,
Demirkalem P, Celenk T and Soyletir G: A possible role of
Bacteroides fragilis enterotoxin in the aetiology of colorectal
cancer. Clin Microbiol Infect. 12:782–786. 2006.PubMed/NCBI View Article : Google Scholar
|
|
174
|
Wu S, Morin PJ, Maouyo D and Sears CL:
Bacteroides fragilis enterotoxin induces c-Myc expression and
cellular proliferation. Gastroenterology. 124:392–400.
2003.PubMed/NCBI View Article : Google Scholar
|
|
175
|
Owen RW, Spiegelhalder B and Bartsch H:
Generation of reactive oxygen species by the faecal matrix. Gut.
46:225–232. 2000.PubMed/NCBI View Article : Google Scholar
|
|
176
|
Huycke MM, Abrams V and Moore DR:
Enterococcus faecalis produces extracellular superoxide and
hydrogen peroxide that damages colonic epithelial cell DNA.
Carcinogenesis. 23:529–536. 2002.PubMed/NCBI
|
|
177
|
Wallace JL: Hydrogen sulfide-releasing
anti-inflammatory drugs. Trends Pharmacol Sci. 28:501–505.
2007.PubMed/NCBI View Article : Google Scholar
|
|
178
|
Attene-Ramos MS, Wagner ED, Gaskins HR and
Plewa MJ: Hydrogen sulfide induces direct radical-associated DNA
damage. Mol Cancer Res. 5:455–459. 2007.PubMed/NCBI View Article : Google Scholar
|
|
179
|
Le Gall T, Clermont O, Gouriou S, Picard
B, Nassif X, Denamur E and Tenaillon O: Extraintestinal virulence
is a coincidental by-product of commensalism in B2 phylogenetic
group Escherichia coli strains. Mol Biol Evol. 24:2373–2384.
2007.PubMed/NCBI View Article : Google Scholar
|
|
180
|
Escobar-Páramo P, Grenet K, Le Menac'h A,
Rode L, Salgado E, Amorin C, Gouriou S, Picard B, Rahimy MC,
Andremont A, et al: Large-scale population structure of human
commensal Escherichia coli isolates. Appl Environ Microbiol.
70:5698–5700. 2004.PubMed/NCBI View Article : Google Scholar
|
|
181
|
Darfeuille-Michaud A, Boudeau J, Bulois P,
Neut C, Glasser AL, Barnich N, Bringer MA, Swidsinski A, Beaugerie
L and Colombel JF: High prevalence of adherent-invasive Escherichia
coli associated with ileal mucosa in Crohn's disease.
Gastroenterology. 127:412–421. 2004.PubMed/NCBI View Article : Google Scholar
|
|
182
|
Han YW, Ikegami A, Rajanna C, Kawsar HI,
Zhou Y, Li M, Sojar HT, Genco RJ, Kuramitsu HK and Deng CX:
Identification and characterization of a novel adhesin unique to
oral fusobacteria. J Bacteriol. 187:5330–5340. 2005.PubMed/NCBI View Article : Google Scholar
|
|
183
|
Prorok-Hamon M, Friswell MK, Alswied A,
Roberts CL, Song F, Flanagan PK, Knight P, Codling C, Marchesi JR,
Winstanley C, et al: Colonic mucosa-associated diffusely adherent
afaC+ Escherichia coli expressing lpfA and pks are increased in
inflammatory bowel disease and colon cancer. Gut. 63:761–770.
2014.PubMed/NCBI View Article : Google Scholar
|
|
184
|
Morrison DJ and Preston T: Formation of
short chain fatty acids by the gut microbiota and their impact on
human metabolism. Gut Microbes. 7:189–200. 2016.PubMed/NCBI View Article : Google Scholar
|
|
185
|
Belcheva A, Irrazabal T, Robertson SJ,
Streutker C, Maughan H, Rubino S, Moriyama EH, Copeland JK,
Surendra A, Kumar S, et al: Gut microbial metabolism drives
transformation of MSH2-deficient colon epithelial cells. Cell.
158:288–299. 2014.PubMed/NCBI View Article : Google Scholar
|
|
186
|
LeBlanc JG, Milani C, de Giori GS, Sesma
F, van Sinderen D and Ventura M: Bacteria as vitamin suppliers to
their host: A gut microbiota perspective. Curr Opin Biotechnol.
24:160–168. 2013.PubMed/NCBI View Article : Google Scholar
|
|
187
|
Russell WR, Gratz SW, Duncan SH, Holtrop
G, Ince J, Scobbie L, Duncan G, Johnstone AM, Lobley GE, Wallace
RJ, et al: High-protein, reduced-carbohydrate weight-loss diets
promote metabolite profiles likely to be detrimental to colonic
health. Am J Clin Nutr. 93:1062–1072. 2011.PubMed/NCBI View Article : Google Scholar
|
|
188
|
Windey K, De Preter V and Verbeke K:
Relevance of protein fermentation to gut health. Mol Nutr Food Res.
56:184–196. 2012.PubMed/NCBI View Article : Google Scholar
|
|
189
|
Neis EP, Dejong CH and Rensen SS: The role
of microbial amino acid metabolism in host metabolism. Nutrients.
7:2930–2946. 2015.PubMed/NCBI View Article : Google Scholar
|
|
190
|
Verbeke KA, Boobis AR, Chiodini A, Edwards
CA, Franck A, Kleerebezem M, Nauta A, Raes J, van Tol EA and Tuohy
KM: Towards microbial fermentation metabolites as markers for
health benefits of prebiotics. Nutr Res Rev. 28:42–66.
2015.PubMed/NCBI View Article : Google Scholar
|
|
191
|
Louis P, Hold GL and Flint HJ: The gut
microbiota, bacterial metabolites and colorectal cancer. Nat Rev
Microbiol. 12:661–672. 2014.PubMed/NCBI View Article : Google Scholar
|
|
192
|
Donohoe DR, Garge N, Zhang X, Sun W,
O'Connell TM, Bunger MK and Bultman SJ: The microbiome and butyrate
regulate energy metabolism and autophagy in the mammalian colon.
Cell Metab. 13:517–526. 2011.PubMed/NCBI View Article : Google Scholar
|
|
193
|
Roediger WE: Role of anaerobic bacteria in
the metabolic welfare of the colonic mucosa in man. Gut.
21:793–798. 1980.PubMed/NCBI View Article : Google Scholar
|
|
194
|
Roediger WE: Utilization of nutrients by
isolated epithelial cells of the rat colon. Gastroenterology.
83:424–429. 1982.PubMed/NCBI
|
|
195
|
Sleeth ML, Thompson EL, Ford HE,
Zac-Varghese SE and Frost G: Free fatty acid receptor 2 and
nutrient sensing: A proposed role for fibre, fermentable
carbohydrates and short-chain fatty acids in appetite regulation.
Nutr Res Rev. 23:135–145. 2010.PubMed/NCBI View Article : Google Scholar
|
|
196
|
Pessione E: Lactic acid bacteria
contribution to gut microbiota complexity: Lights shadows. Front
Cell Infect Microbiol. 2(86)2012.PubMed/NCBI View Article : Google Scholar
|
|
197
|
Hamer HM, Jonkers D, Venema K, Vanhoutvin
S, Troost FJ and Brummer RJ: Review article: The role of butyrate
on colonic function. Aliment Pharmacol Ther. 27:104–119.
2008.PubMed/NCBI View Article : Google Scholar
|
|
198
|
Fung KYC, Cosgrove L, Lockett T, Head R
and Topping DL: A review of the potential mechanisms for the
lowering of colorectal oncogenesis by butyrate. Br J Nutr.
108:820–831. 2012.PubMed/NCBI View Article : Google Scholar
|
|
199
|
Wilson AJ, Chueh AC, Tögel L, Corner GA,
Ahmed N, Goel S, Byun DS, Nasser S, Houston MA, Jhawer M, et al:
Apoptotic sensitivity of colon cancer cells to histone deacetylase
inhibitors is mediated by an Sp1/Sp3-activated transcriptional
program involving immediate-early gene induction. Cancer Res.
70:609–620. 2010.PubMed/NCBI View Article : Google Scholar
|
|
200
|
Chang PV, Hao L, Offermanns S and
Medzhitov R: The microbial metabolite butyrate regulates intestinal
macrophage function via histone deacetylase inhibition. Proc Natl
Acad Sci USA. 111:2247–2252. 2014.PubMed/NCBI View Article : Google Scholar
|
|
201
|
Smith PM, Howitt MR, Panikov N, Michaud M,
Gallini CA, Bohlooly YM, Glickman JN and Garrett WS: The microbial
metabolites, short-chain fatty acids, regulate colonic Treg cell
homeostasis. Science. 341:569–573. 2013.PubMed/NCBI View Article : Google Scholar
|
|
202
|
Furusawa Y, Obata Y, Fukuda S, Endo TA,
Nakato G, Takahashi D, Nakanishi Y, Uetake C, Kato K, Kato T, et
al: Commensal microbe-derived butyrate induces the differentiation
of colonic regulatory T cells. Nature. 504:446–450. 2013.PubMed/NCBI View Article : Google Scholar
|
|
203
|
Arpaia N, Campbell C, Fan X, Dikiy S, van
der Veeken J, deRoos P, Liu H, Cross JR, Pfeffer K, Coffer PJ, et
al: Metabolites produced by commensal bacteria promote peripheral
regulatory T-cell generation. Nature. 504:451–455. 2013.PubMed/NCBI View Article : Google Scholar
|
|
204
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the Warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009.PubMed/NCBI View Article : Google Scholar
|
|
205
|
Donohoe DR, Holley D, Collins LB,
Montgomery SA, Whitmore AC, Hillhouse A, Curry KP, Renner SW,
Greenwalt A, Ryan EP, et al: A gnotobiotic mouse model demonstrates
that dietary fiber protects against colorectal tumorigenesis in a
microbiota- and butyrate-dependent manner. Cancer Discov.
4:1387–1397. 2014.PubMed/NCBI View Article : Google Scholar
|
|
206
|
Donohoe DR, Collins LB, Wali A, Bigler R,
Sun W and Bultman SJ: The Warburg effect dictates the mechanism of
butyrate-mediated histone acetylation and cell proliferation. Mol
Cell. 48:612–626. 2012.PubMed/NCBI View Article : Google Scholar
|
|
207
|
Sebastián C and Mostoslavsky R: Untangling
the fiber yarn: Butyrate feeds Warburg to suppress colorectal
cancer. Cancer Discov. 4:1368–1370. 2014.PubMed/NCBI View Article : Google Scholar
|
|
208
|
Davie JR: Inhibition of histone
deacetylase activity by butyrate. J Nutr. 133((Suppl 7)):
2485S–2493S. 2003.PubMed/NCBI View Article : Google Scholar
|
|
209
|
Gonçalves P and Martel F: Butyrate and
colorectal cancer: The role of butyrate transport. Curr Drug Metab.
14:994–1008. 2013.PubMed/NCBI View Article : Google Scholar
|
|
210
|
Thangaraju M, Cresci GA, Liu K, Ananth S,
Gnanaprakasam JP, Browning DD, Mellinger JD, Smith SB, Digby GJ,
Lambert NA, et al: GPR109A is a G-protein-coupled receptor for the
bacterial fermentation product butyrate and functions as a tumor
suppressor in colon. Cancer Res. 69:2826–2832. 2009.PubMed/NCBI View Article : Google Scholar
|
|
211
|
Singh N, Gurav A, Sivaprakasam S, Brady E,
Padia R, Shi H, Thangaraju M, Prasad PD, Manicassamy S, Munn DH, et
al: Activation of Gpr109a, receptor for niacin and the commensal
metabolite butyrate, suppresses colonic inflammation and
carcinogenesis. Immunity. 40:128–139. 2014.PubMed/NCBI View Article : Google Scholar
|
|
212
|
Larrosa M, González-Sarrías A,
García-Conesa MT, Tomás-Barberán FA and Espín JC: Urolithins,
ellagic acid-derived metabolites produced by human colonic
microflora, exhibit estrogenic and antiestrogenic activities. J
Agric Food Chem. 54:1611–1620. 2006.PubMed/NCBI View Article : Google Scholar
|
|
213
|
González-Sarrías A, Larrosa M,
Tomás-Barberán FA, Dolara P and Espín JC: NF-kappaB-dependent
anti-inflammatory activity of urolithins, gut microbiota ellagic
acid-derived metabolites, in human colonic fibroblasts. Br J Nutr.
104:503–512. 2010.PubMed/NCBI View Article : Google Scholar
|
|
214
|
Atkinson C, Frankenfeld CL and Lampe JW:
Gut bacterial metabolism of the soy isoflavone daidzein: Exploring
the relevance to human health. Exp Biol Med (Maywood). 230:155–170.
2005.PubMed/NCBI
|
|
215
|
Bolca S, Possemiers S, Herregat A,
Huybrechts I, Heyerick A, De Vriese S, Verbruggen M, Depypere H, De
Keukeleire D, Bracke M, et al: Microbial and dietary factors are
associated with the equol producer phenotype in healthy
postmenopausal women. J Nutr. 137:2242–2246. 2007.PubMed/NCBI View Article : Google Scholar
|
|
216
|
Lampe JW: Emerging research on equol
cancer. J Nutr. 140:1369S–1372S. 2010.PubMed/NCBI View Article : Google Scholar
|
|
217
|
Davis CD and Milner JA: Gastrointestinal
microflora, food components and colon cancer prevention. J Nutr
Biochem. 20:743–752. 2009.PubMed/NCBI View Article : Google Scholar
|
|
218
|
Bode LM, Bunzel D, Huch M, Cho GS, Ruhland
D, Bunzel M, Bub A, Franz CM and Kulling SE: In vivo and in vitro
metabolism of trans-resveratrol by human gut microbiota. Am J Clin
Nutr. 97:295–309. 2013.PubMed/NCBI View Article : Google Scholar
|
|
219
|
Greer JB and O'Keefe SJ: Microbial
induction of immunity, inflammation, and cancer. Front Physiol.
1(168)2011.PubMed/NCBI View Article : Google Scholar
|
|
220
|
Ridlon JM, Kang DJ and Hylemon PB: Bile
salt biotransformations by human intestinal bacteria. J Lipid Res.
47:241–259. 2006.PubMed/NCBI View Article : Google Scholar
|
|
221
|
Wells JE, Williams KB, Whitehead TR,
Heuman DM and Hylemon PB: Development and application of a
polymerase chain reaction assay for the detection and enumeration
of bile acid 7alpha-dehydroxylating bacteria in human feces. Clin
Chim Acta. 331:127–134. 2003.PubMed/NCBI View Article : Google Scholar
|
|
222
|
Rubin DC, Shaker A and Levin MS: Chronic
intestinal inflammation: Inflammatory bowel disease and
colitis-associated colon cancer. Front Immunol.
3(107)2012.PubMed/NCBI View Article : Google Scholar
|
|
223
|
Powolny A, Xu J and Loo G: Deoxycholate
induces DNA damage and apoptosis in human colon epithelial cells
expressing either mutant or wild-type p53. Int J Biochem Cell Biol.
33:193–203. 2001.PubMed/NCBI View Article : Google Scholar
|
|
224
|
Imray CH, Radley S, Davis A, Barker G,
Hendrickse CW, Donovan IA, Lawson AM, Baker PR and Neoptolemos JP:
Faecal unconjugated bile acids in patients with colorectal cancer
or polyps. Gut. 33:1239–1245. 1992.PubMed/NCBI View Article : Google Scholar
|
|
225
|
Nagengast FM, Grubben MJ and van Munster
IP: Role of bile acids in colorectal carcinogenesis. Eur J Cancer.
31A:1067–1070. 1995.PubMed/NCBI View Article : Google Scholar
|
|
226
|
Carmody RN and Turnbaugh PJ:
Host-microbial interactions in the metabolism of therapeutic and
diet-derived xenobiotics. J Clin Invest. 124:4173–4181.
2014.PubMed/NCBI View Article : Google Scholar
|
|
227
|
Flanagan L, Schmid J, Ebert M, Soucek P,
Kunicka T, Liska V, Bruha J, Neary P, Dezeeuw N, Tommasino M, et
al: Fusobacterium nucleatum associates with stages of colorectal
neoplasia development, colorectal cancer and disease outcome. Eur J
Clin Microbiol Infect Dis. 33:1381–1390. 2014.PubMed/NCBI View Article : Google Scholar
|
|
228
|
McCoy AN, Araújo-Pérez F, Azcárate-Peril
A, Yeh JJ, Sandler RS and Keku TO: Fusobacterium is associated with
colorectal adenomas. PLoS One. 8(e53653)2013.PubMed/NCBI View Article : Google Scholar
|
|
229
|
Fukugaiti MH, Ignacio A, Fernandes MR,
Ribeiro Júnior U, Nakano V and Avila-Campos MJ: High occurrence of
Fusobacterium nucleatum and Clostridium difficile in the intestinal
microbiota of colorectal carcinoma patients. Braz J Microbiol.
46:1135–1140. 2015.PubMed/NCBI View Article : Google Scholar
|
|
230
|
Zackular JP, Rogers MAM, Ruffin MT IV and
Schloss PD: The human gut microbiome as a screening tool for
colorectal cancer. Cancer Prev Res (Phila). 7:1112–1121.
2014.PubMed/NCBI View Article : Google Scholar
|
|
231
|
Mima K, Nishihara R, Qian ZR, Cao Y,
Sukawa Y, Nowak JA, Yang J, Dou R, Masugi Y, Song M, et al:
Fusobacterium nucleatum in colorectal carcinoma tissue and patient
prognosis. Gut. 65:1973–1980. 2016.PubMed/NCBI View Article : Google Scholar
|
|
232
|
Ballal SA, Veiga P, Fenn K, Michaud M, Kim
JH, Gallini CA, Glickman JN, Quéré G, Garault P, Béal C, et al:
Host lysozyme-mediated lysis of Lactococcus lactis facilitates
delivery of colitis-attenuating superoxide dismutase to inflamed
colons. Proc Natl Acad Sci USA. 112:7803–7808. 2015.PubMed/NCBI View Article : Google Scholar
|
|
233
|
Veiga P, Gallini CA, Beal C, Michaud M,
Delaney ML, DuBois A, Khlebnikov A, van Hylckama Vlieg JE, Punit S,
Glickman JN, et al: Bifidobacterium animalis subsplactis fermented
milk product reduces inflammation by altering a niche for
colitogenic microbes. Proc Natl Acad Sci USA. 107:18132–18137.
2010.PubMed/NCBI View Article : Google Scholar
|
|
234
|
Viaud S, Saccheri F, Mignot G, Yamazaki T,
Daillère R, Hannani D, Enot DP, Pfirschke C, Engblom C, Pittet MJ,
et al: The intestinal microbiota modulates the anticancer immune
effects of cyclophosphamide. Science. 342:971–976. 2013.PubMed/NCBI View Article : Google Scholar
|
|
235
|
Siddik ZH: Cisplatin: Mode of cytotoxic
action and molecular basis of resistance. Oncogene. 22:7265–7279.
2003.PubMed/NCBI View Article : Google Scholar
|
|
236
|
Yang J, Liu KX, Qu JM and Wang XD: The
changes induced by cyclophosphamide in intestinal barrier and
microflora in mice. Eur J Pharmacol. 714:120–124. 2013.PubMed/NCBI View Article : Google Scholar
|
|
237
|
Nam YD, Kim HJ, Seo JG, Kang SW and Bae
JW: Impact of pelvic radiotherapy on gut microbiota of
gynecological cancer patients revealed by massive pyrosequencing.
PLoS One. 8(e82659)2013.PubMed/NCBI View Article : Google Scholar
|
|
238
|
Jenq RR, Ubeda C, Taur Y, Menezes CC,
Khanin R, Dudakov JA, Liu C, West ML, Singer NV, Equinda MJ, et al:
Regulation of intestinal inflammation by microbiota following
allogeneic bone marrow transplantation. J Exp Med. 209:903–911.
2012.PubMed/NCBI View Article : Google Scholar
|
|
239
|
Von Bültzingslöwen I, Adlerberth I, Wold
AE, Dahlén G and Jontell M: Oral and intestinal microflora in
5-fluorouracil treated rats, translocation to cervical and
mesenteric lymph nodes and effects of probiotic bacteria. Oral
Microbiol Immunol. 18:278–284. 2003.PubMed/NCBI View Article : Google Scholar
|
|
240
|
Wallace BD, Wang H, Lane KT, Scott JE,
Orans J, Koo JS, Venkatesh M, Jobin C, Yeh LA, Mani S, et al:
Alleviating cancer drug toxicity by inhibiting a bacterial enzyme.
Science. 330:831–835. 2010.PubMed/NCBI View Article : Google Scholar
|
|
241
|
Lam W, Bussom S, Guan F, Jiang Z, Zhang W,
Gullen EA, Liu SH and Cheng YC: The four-herb Chinese medicine
PHY906 reduces chemotherapy-induced gastrointestinal toxicity. Sci
Transl Med. 2(45ra59)2010.PubMed/NCBI View Article : Google Scholar
|
|
242
|
Cesaro C, Tiso A, Del Prete A, Cariello R,
Tuccillo C, Cotticelli G, Del Vecchio Blanco C and Loguercio C: Gut
microbiota and probiotics in chronic liver diseases. Dig Liver Dis.
43:431–438. 2011.PubMed/NCBI View Article : Google Scholar
|
|
243
|
Hemarajata P and Versalovic J: Effects of
probiotics on gut microbiota: Mechanisms of intestinal
immunomodulation and neuromodulation. Therap Adv Gastroenterol.
6:39–51. 2013.PubMed/NCBI View Article : Google Scholar
|
|
244
|
Ekmekciu I, von Klitzing E, Fiebiger U,
Neumann C, Bacher P, Scheffold A, Bereswill S and Heimesaat MM: The
probiotic compound VSL#3 modulates mucosal, peripheral, and
systemic immunity following murine broad-spectrum antibiotic
treatment. Front Cell Infect Microbiol. 7(167)2017.PubMed/NCBI View Article : Google Scholar
|
|
245
|
Rao RK and Samak G: Protection and
restitution of gut barrier by probiotics Nutritional and clinical
implications. Curr Nutr Food Sci. 9:99–107. 2013.PubMed/NCBI
|
|
246
|
Kamada N, Chen GY, Inohara N and Núñez G:
Control of pathogens and pathobionts by the gut microbiota. Nat
Immunol. 14:685–690. 2013.PubMed/NCBI View Article : Google Scholar
|
|
247
|
Jonkers D and Stockbrügger R: Probiotics
and inflammatory bowel disease. J R Soc Med. 96:167–171.
2003.PubMed/NCBI
|
|
248
|
Celiberto LS, Bedani R, Rossi EA and
Cavallini DC: Probiotics: The scientific evidence in the context of
inflammatory bowel disease. Crit Rev Food Sci Nutr. 57:1759–1768.
2017.PubMed/NCBI View Article : Google Scholar
|
|
249
|
Sheil B, Shanahan F and O'Mahony L:
Probiotic effects on inflammatory bowel disease. J Nutr. 137:(Suppl
2):819S–824S. 2007.PubMed/NCBI View Article : Google Scholar
|
|
250
|
Tamboli CP, Caucheteux C, Cortot A,
Colombel JF and Desreumaux P: Probiotics in inflammatory bowel
disease: A critical review. Best Pract Res Clin Gastroenterol.
17:805–820. 2003.PubMed/NCBI View Article : Google Scholar
|
|
251
|
Bibiloni R, Fedorak RN, Tannock GW, Madsen
KL, Gionchetti P, Campieri M, De Simone C and Sartor RB: VSL#3
probiotic-mixture induces remission in patients with active
ulcerative colitis. Am J Gastroenterol. 100:1539–1546.
2005.PubMed/NCBI View Article : Google Scholar
|
|
252
|
Kim S, Covington A and Pamer EG: The
intestinal microbiota: Antibiotics, colonization resistance, and
enteric pathogens. Immunol Rev. 279:90–105. 2017.PubMed/NCBI View Article : Google Scholar
|
|
253
|
Dethlefsen L, Huse S, Sogin ML and Relman
DA: The pervasive effects of an antibiotic on the human gut
microbiota, as revealed by deep 16S rRNA sequencing. PLoS Biol.
6(e280)2008.PubMed/NCBI View Article : Google Scholar
|
|
254
|
Dethlefsen L and Relman DA: Incomplete
recovery and individualized responses of the human distal gut
microbiota to repeated antibiotic perturbation. Proc Natl Acad Sci
USA. 108((Suppl 1)): 4554–4561. 2011.PubMed/NCBI View Article : Google Scholar
|
|
255
|
Jernberg C, Löfmark S, Edlund C and
Jansson JK: Long-term ecological impacts of antibiotic
administration on the human intestinal microbiota. ISME J. 1:56–66.
2007.PubMed/NCBI View Article : Google Scholar
|
|
256
|
Lankelma JM, Cranendonk DR, Belzer C, de
Vos AF, de Vos WM, van der Poll T and Wiersinga WJ:
Antibiotic-induced gut microbiota disruption during human
endotoxemia: A randomised controlled study. Gut. 66:1623–1630.
2017.PubMed/NCBI View Article : Google Scholar
|
|
257
|
Lopez J and Grinspan A: Fecal microbiota
transplantation for inflammatory bowel disease. Gastroenterol
Hepatol (NY). 12:374–379. 2016.PubMed/NCBI
|
|
258
|
McFarland LV: Epidemiology, risk factors
and treatments for antibiotic-associated diarrhea. Dig Dis.
16:292–307. 1998.PubMed/NCBI View Article : Google Scholar
|
|
259
|
Suez J, Zmora N, Zilberman-Schapira G, Mor
U, Dori-Bachash M, Bashiardes S, Zur M, Regev-Lehavi D, Ben-Zeev
Brik R, Federici S, et al: Post-antibiotic gut mucosal microbiome
reconstitution is impaired by probiotics and improved by autologous
FMT. Cell. 174:1406–1423, e16. 2018.PubMed/NCBI View Article : Google Scholar
|
|
260
|
Chaput N, Lepage P, Coutzac C, Soularue E,
Le Roux K, Monot C, Boselli L, Routier E, Cassard L, Collins M, et
al: Baseline gut microbiota predicts clinical response and colitis
in metastatic melanoma patients treated with ipilimumab. Ann Oncol.
28:1368–1379. 2017.PubMed/NCBI View Article : Google Scholar
|
|
261
|
Frankel AE, Coughlin LA, Kim J, Froehlich
TW, Xie Y, Frenkel EP and Koh AY: Metagenomic shotgun sequencing
and unbiased metabolomic profiling identify specific human gut
microbiota and metabolites associated with immune checkpoint
therapy efficacy in melanoma patients. Neoplasia. 19:848–855.
2017.PubMed/NCBI View Article : Google Scholar
|
|
262
|
Gopalakrishnan V, Spencer CN, Nezi L,
Reuben A, Andrews MC, Karpinets TV, Prieto PA, Vicente D, Hoffman
K, Wei SC, et al: Gut microbiome modulates response to anti-PD-1
immunotherapy in melanoma patients. Science. 359:97–103.
2018.PubMed/NCBI View Article : Google Scholar
|
|
263
|
Matson V, Fessler J, Bao R, Chongsuwat T,
Zha Y, Alegre ML, Luke JJ and Gajewski TF: The commensal microbiome
is associated with anti-PD-1 efficacy in metastatic melanoma
patients. Science. 359:104–108. 2018.PubMed/NCBI View Article : Google Scholar
|
|
264
|
Routy B, Le Chatelier E, Derosa L, Duong
CPM, Alou MT, Daillère R, Fluckiger A, Messaoudene M, Rauber C,
Roberti MP, et al: Gut microbiome influences efficacy of PD-1-based
immunotherapy against epithelial tumors. Science. 359:91–97.
2018.PubMed/NCBI View Article : Google Scholar
|
|
265
|
Vétizou M, Pitt JM, Daillère R, Lepage P,
Waldschmitt N, Flament C, Rusakiewicz S, Routy B, Roberti MP, Duong
CP, et al: Anticancer immunotherapy by CTLA-4 blockade relies on
the gut microbiota. Science. 350:1079–1084. 2015.PubMed/NCBI View Article : Google Scholar
|
|
266
|
Sivan A, Corrales L, Hubert N, Williams
JB, Aquino-Michaels K, Earley ZM, Benyamin FW, Lei YM, Jabri B,
Alegre ML, et al: Commensal Bifidobacterium promotes antitumor
immunity and facilitates anti-PD-L1 efficacy. Science.
350:1084–1089. 2015.PubMed/NCBI View Article : Google Scholar
|