Introduction
Growth hormone (GH), a 191-amino acid single-chain
peptide, is a peptide hormone with anabolic and
performance-enhancing effects (1).
Direct effects of GH include increasing muscle mass (2) Most of the indirect effects are mediated
by induction of insulin-like growth factor 1 (IGF-1) expression in
the liver and in peripheral tissues (3). An organ most affected by GH imbalance
(deficiency or excess) is the heart (4). Bodybuilders and athletes misuse
recombinant human GH (r-hGH) preparations to increase muscle mass
and strength; as such, r-hGH has been on the prohibited substances
list of the International Federations and the International Olympic
Committee since 1989. It is also on the 2010 World Anti-Doping
Agency list of prohibited substances (5,6).
Left ventricular hypertrophy resulting from swimming
exercise is an important physiological mechanism that increases
sarcomeres and cardiac cell growth to compensate for the chronic
increase in hemodynamic load (7).
Conversely, pathological left ventricular hypertrophy caused by
cardiovascular disease is associated with increased fibrosis and
decreased aerobic capacity leading to incompatible remodeling in
cardiac cells (7). Both in
physiological and pathological hypertrophy, intermediate signal
transduction pathways in the cytoplasm are triggered via
membrane-bound receptors. Signal transduction pathways critical for
myosin growth include phosphoinositide-3-kinase
(PI3K)/serine/threonine protein kinase (AKT)/mechanistic target of
rapamycin (mTOR), mitogen-activated protein kinase
(MAPK)/extracellular signal-regulated kinase (ERK), protein kinase
C and nuclear factor of activated T cells pathways, among others
(8). In particular, the PI3K/AKT/mTOR
signaling pathway has been reported to serve a regulatory role in
cell growth, metabolism, survival and angiogenesis, and to govern
the development and transformation of left ventricular hypertrophy
(7,9).
The ERK signaling pathway, the members of which belong to the MAPK
family, comprises a cascade consisting of a series of successively
acting kinases that finally results in the phosphorylation and
activation of terminal kinases including p38, c-Jun N-terminal
kinases and ERK. Following activation, each ERK phosphorylates
intracellular targets including transcription factors, resulting in
reprogramming of cardiac gene expression (10,11).
MicroRNAs (miRNA/miRs) are small RNA molecules of
18-25 nucleotides which do not encode proteins, and negatively
regulate gene expression (12,13). They
are involved in the regulation of several physiological responses
including cell apoptosis (14),
proliferation and differentiation (15). In addition, recent studies have
demonstrated that certain miRNAs are involved in cardiovascular
diseases and pathological left ventricular hypertrophy (16-18).
In particular, miR-21 and miR-133 have been demonstrated to
regulate cardiac hypertrophy (17,19).
miR-21 has also been implicated in the regulation of cardiac
fibrosis and cardiac muscle contractility (20), and has been shown to negatively
regulate phosphatase and tensin homolog (PTEN) gene expression
(7).
It is established that r-hGH may cause cardiac
hypertrophy (6). However, to the best
of our knowledge, there is no study demonstrating the mechanism
through which r-hGH, together with swimming exercise, affects
signaling pathways (PI3K/AKT/mTOR and ERK) that regulate cardiac
signaling genes and the miRNAs (miR-21 and miR-133) targeting the
genes in these signaling pathways. Thus, the present study aimed to
investigate i) the effect of r-hGH and exercise on cardiac
PI3K/AKT/mTOR and ERK signaling pathways, and ii) the role of
miR-21 and miR-133 expression in ventricular muscle.
Materials and methods
Animals and exercise protocols
A total of 36 male Sprague-Dawley rats (14 weeks
old) weighing 200-250 g were obtained from the Experimental Animals
Center of Trakya University, Edirne, Turkey. All animals were fed
daily with tap water and a standard pellet diet for rats under
optimal laboratory conditions (temperature: 22±2˚C; humidity:
50-55%; light/dark cycle: 12/12 h). The animals were randomly
stratified into sedentary control (SC, n=9), swimming exercise (SE,
n=8), r-hGH (GH, n=10) and swimming exercise plus r-hGH (SE-GH,
n=9) groups. For 8 weeks, from Monday to Friday, the rats in the SE
and SE-GH groups completed a swimming exercise for 1 h per day. A
low-intensity, long-period swimming exercise protocol was applied
(7,21). The exercise protocol was selected as
it has been tested in previous studies and been determined to be
effective in demonstrating cardiovascular adaptations and muscular
oxidative capacity (21,22). Animals in the swimming exercise groups
were allowed to swim for 1 h wearing a weight of 5% of their body
weight attached to the tail. Prior to the exercise program, the
rats in the swimming exercise groups were subjected to an
adaptation period of 5 days during which the duration and load were
gradually increased according to increase in animal weight. Animals
that could not adapt to the experiment (refused to swim or prone to
plunging) were excluded from the study. In addition, animals in the
non-exercise groups (SC and GH) were exposed to pool water by means
of an apparatus placed inside the pool for 8 weeks, and from Monday
to Friday, the rats in the SE group completed a 1-h swimming
exercise (7). The swimming pool
(120x80x75 cm) was filled with water to a depth of 65 cm. The pool
was divided by plastic barriers into twelve lanes. To avoid
interaction, each rat was placed into an individual lane. Water
temperature was maintained at 31±1˚C.
r-hGH was administered subcutaneously 5 days per
week for 8 weeks to the animals in the GH and SE-GH groups at a
dose of 0.3 (1 IU/day) mg/kg (Humatrope®, Eli Lilly and Company,
Indianapolis, IN, USA). The dose was selected based on previous
studies (18-21).
Animals in the SE-GH group were injected subcutaneously immediately
prior to exercise. Saline at the same dose (0.3 mg/kg) was injected
subcutaneously to the animals in the SE group.
The study was conducted in the Trakya University
Local Experimental Animals Research Center after obtaining approval
from the institutional Experimental Animals Ethics Committee
(approval no. TUHADYEK:2015/10).
Preparation of samples
After fasting for 24 h following the last exercise,
the rats received intraperitoneal thiopental (100 mg/kg) for
anesthesia. All rats were sacrificed by cervical dislocation and
blood (5 ml) was obtained from the heart for metabolic
investigations. Serum was separated from the blood samples via
centrifugation at 2,600 x g for 10 min at room temperature, and
stored at -20˚C to measure IGF-1 and insulin levels at a later time
point. Measurement of IGF1 and insulin was performed by means of
ELISAs. Isolated hearts were weighed and portions of the left
ventricular tissue were stored at -80˚C for mRNA and miRNA
measurements, and the remaining left ventricular tissue was
utilized to create paraffin blocks for immunohistochemical
evaluation.
Real-time reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analyses
Total RNA isolation and real-time PCR for
mRNA. Total RNA was isolated from the stored left ventricular
tissues using a Total RNA PureLink® RNA Mini kit as per the
manufacturer's instructions (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). The quantity of extracted RNA was
determined with a Qubit® Fluorometer (Invitrogen; Thermo Fisher
Scientific, Inc.).
RNA concentrations of the samples were adjusted to
200 ng/µl and cDNA synthesis was performed using a High Capacity
cDNA Reverse Transcription kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.). Gene expression was analyzed by means of
RT-qPCR. The PCR cycling conditions were an initial denaturation at
95˚C for 5 min followed by 40 cycles at 95˚C for 15 sec and 60˚C
for 45 sec and 72˚C for 15 sec. The primers, sequences of which are
provided in Table I, were used with
SYBR® Select Master Mix (Applied Biosystems; Thermo Fisher
Scientific, Inc.) to determine the changes in gene expression. Gene
expressions were determined as the relative fold change compared
with the control group, and normalized to β-actin mRNA
expression.
 | Table I.Quantitative real-time polymerase
chain reaction primers. |
Table I.
Quantitative real-time polymerase
chain reaction primers.
| Gene | Sequence,
5'-3' |
|---|
| PI3K | F:
CATGGATGCTTTGCAGGGTTT |
| | R:
CCAGATGTTCTCCATGATTCGGA |
| PTEN | F:
AGACCATAACCCACCACAGC |
| | R:
TACACCAGTCCGTCCTTTCC |
| AKT1 | F:
ACTGACATTGGACGGCTGAG |
| | R:
CAGGTGGGACTGTGATACGG |
| ERK1/2 | F:
GAGCCCAGGGGAACTGCT |
| | R:
TGGAAGCGGGCTGTCTC |
| ANP | F:
CTTCGGGGGTAGGATTGAC |
| | R:
CTTGGGATCTTTTGCGATCT |
| β-MHC | F:
CATCCCCAATGAGACGAAG |
| | R:
AGGCTCTTTCTGCTGGACA |
| β-actin | F:
AGAGGGAAATCGTGCGTGAC |
| | R:
AGGAAGGAAGGCTGGAAGAGA |
TaqMan miRNA assay and real-time PCR for
miRNA. miRNA isolation from left ventricular tissues was
performed with a mirVana miRNA Isolation kit (Ambion) as per the
manufacturer's instructions. cDNA for miRNA analysis was
synthesized from total RNA utilizing gene-specific primers
according to the TaqMan MicroRNA Reverse Transcription Assay
protocol (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
15 µl reactions obtained by the TaqMan MicroRNA Reverse
Transcription kit protocol were incubated in a thermal cycler (10
min at 95˚C followed by 40 cycles of 15 sec at 95˚C, 1 min at 60˚C
and 15 sec at 72˚C). PCR quantification was performed by means of a
TaqMan MicroRNA Assay (Thermo Fisher Scientific, Inc.) protocol.
The probe mix from the TaqMan MicroRNA Assay protocol contained
sequences for miR-21 and -133 (Taqman MicroRNA Assay 002393 and
002246; Thermo Fisher Scientific, Inc.). Samples were normalized by
evaluating against U6 gene expression (TaqMan MicroRNA Assay
001973).
Quantification
PCR product generation was monitored by measuring
the increase in fluorescence caused by probe presence in the TaqMan
MicroRNA Assay at each annealing phase or by the binding of
SYBR-Green to double-stranded DNA. A melting curve in the
SYBR-Green analysis was generated at the end of the reaction to
determine that only one product was amplified. Each assay was
performed in duplicate. The relative quantities of target gene
expression between sedentary rats and rats subjected to swimming
exercise with and without r-hGH treatment were compared following
normalization to the values of internal control (∆Cq). Fold change
in mRNA and miRNA expressions were calculated using the differences
in ∆Cq values between two samples (∆∆Cq) and the equation, 2-∆∆Cq
(23). The results were expressed as
a percentage of the sedentary control group.
ELISA measurements
The stored serum samples were evaluated using ELISA
kits for IGF-1 (Rat IGF-1, cat no. EK0377; Wuhan Boster Biological
Technology, Ltd., Wuhan, China) and insulin (Rat/Mouse insulin, cat
no. EZRMI-13K; EMD Millipore, Billerica, MA, USA) as per the
manufacturer's instructions.
In the experimental procedure for IGF-1 measurement,
a standard sandwich technique involving enzyme-linked immunosorbent
technology was used. Absorbance was measured at 450 nm with an
ELISA reader (FL 800; BioTek Instruments, Inc., Winooski, VT, USA).
The IGF-1 assay sensitivity was <5 pg/ml and the range of the
assay was 62.5-4.000 pg/ml.
In the experimental procedure for insulin
measurement, a double-antibody sandwich enzyme-linked immunosorbent
assay was used to determine insulin level in the samples.
Absorbance was measured at 450 nm with the ELISA reader. The
insulin assay sensitivity was 0.2 ng/ml (17.5 pM for a sample
volume of 10 µl) and the appropriate range of the assay was 0.2-10
ng/ml.
Two repeats were performed.
Immunohistochemical method and
evaluation
Left ventricular tissue samples were fixed for 24 h
at room temperature in 10% formalin solution. The samples were
subsequently passed through an ascending ethyl alcohol series
(60.0-70.0-80.0-90.0-99.9%). The tissues were exposed to two
changes of xylene for transparency and then embedded in paraffin.
Following deparaffinization of 5-µm sections from the blocks, one
section of each block was stained with hematoxylin and eosin
(H&E) stain for general histological examination of the
tissues. Counterstaining was performed with Hematoxylin for 3-5 min
at room temperature and eosin for different time periods under
visual control. For the other sections, a Ventana BenchMark XT
Automated IHC/ISH slide staining system (Ventana Medical Systems,
Inc., Tucson, AZ, USA) was used to apply immunohistological stains
against the following: PI3K catalytic subunit α (polypeptide p110;
PIK3CA; cat no. 09-482), PTEN (cat. no. 09-035), AKT1
[phosphorylated (p)-AKT Ser473], cat no. 07-1398), p-mTOR (p-mTOR
Ser2448 catalytic domain, cat no. 07-1415; all from EMD Millipore)
and ERK1/2 (p-ERK1/2 Thr202/Thr204; cat no. 4370S; Cell Signaling
Technology, Inc., Danvers, MA, USA).
Immunohistochemical scoring was conducted in a
manner entirely blinded to all clinical and biological variables
(24). PIK3CA staining was scored on
the following scale: 0, no staining; 1+, staining <50%; 2+,
staining ≥50% with weak intensity; 3+, ≥50% staining with strong
intensity (25).
PTEN expression was scored independently based on
the intensity and extent of staining. The intensity of positive
staining was scored from 0 to 2 as follows: 0 (none), 1 (weak;
intensity < positive control), 2 (strong; intensity ≥ positive
control). Positive staining was assessed using a semi-quantitative
grading system consisting of five categories: 0, <5% positive
cells; 1, 6-25% positive cells; 2, 26-50% positive cells; 3, 51-75%
positive cells; 4, 76-100% positive cells. The sum of the two
values provided the total score, and a score <4 was considered
PTEN-negative (26).
p-AKT, p-mTOR and ERK levels were stratified into
three categories based on staining intensity as well as positive
frequency according to a previously described scoring method
(9) with slight modification. Tissues
where <10% of the cells exhibited weak staining were scored as
0, tissues where >10% of the cells exhibited weak staining or
<20% exhibited strong staining were scored as 1, and tissues
where >20% of the cells exhibited strong staining were scored as
2(26).
Statistical analysis
All data are presented as the mean ± standard error
of the mean. All statistical analyses were conducted using SPSS for
Windows v21.0 (IBM Corp., Armonk, NY, USA). GraphPad Prism for
Windows v6.03 (GraphPad Software, Inc., La Jolla, CA, USA) was used
to construct the graphs. The one-sample Kolmogorov Smirnov test was
used to evaluate the normal distribution of data. The effects of GH
administration and swimming exercise were analyzed by two-way
analysis of variance. The Bonferroni post-hoc test was applied for
post-hoc comparisons between groups, and P<0.05 was considered
to indicate a statistically significant difference.
Results
Clinical characteristics
Results of all physiological and metabolic
parameters of the study evaluating the effects of r-hGH
administration and swimming exercise are presented in Table II. Comparison of heart weights
revealed an increase in the SE-GH group compared with in the SC
group (P<0.05). This significant difference was deemed to result
from the swimming exercise and r-hGH administration. In terms of
left ventricular weight, this was significantly increased in the
SE-GH group (P<0.05) compared with in the SC group (data not
shown). Cardiac hypertrophy grade was determined by heart
weight/body weight ratio, referred to as the heart weight index
(HWI; heart weight /body weight x1,000 g/g) (27,28).
Significant increase in this ratio was observed in the SE
(P<0.05) and SE-GH (P<0.001) groups compared with in the SC
group. The difference in left ventricular wall thickness, as
another parameter used to determine cardiac hypertrophy was not
statistically significant between the groups (28). Further, no significant differences
were observed between the groups when r-hGH administration plus
exercise was evaluated for the effect on the two metabolic
parameters, insulin and IGF-1.
| Table II.Effects of r-hGH and exercise on
physiological properties and metabolic parameters in rats. |
Table II.
Effects of r-hGH and exercise on
physiological properties and metabolic parameters in rats.
| |
P-valuea |
|---|
| |
------------------------------------------------------------------------------ |
|---|
| Parameters | SC (n=10) | SE (n=8) | GH (n=10) | SE-GH (n=9) | SE effect | GH effect | GH+SE effect |
|---|
| BW, g | 307.80±6.34 | 269.38±7.09 | 281.50±6.34 | 275.11±6.69 | <0.05 | 0.13 | <0.05 |
| HWI | 3.64±0.03 | 4.21±0.07 | 4.02±0.06 | 4.40±0.07 | <0.05 | 0.08 | <0.001 |
| LVWT, mm | 3.09±0.06 | 3.13±0.06 | 3.14±0.05 | 3.23±0.08 | 0.23 | 0.67 | 0.29 |
| Insulin, mIU/l | 0.68±0.13 | 0.44±0.08 | 0.58±0.12 | 0.57±0.11 | 0.89 | 0.29 | 0.34 |
| IGF-1, pg/ml |
5,736.05±306.62 |
6,410.46±226.46 |
6,260.12±202.55 |
5,922.10±142.87 | 0.69 | 0.97 | 0.23 |
Molecular markers of left ventricular
hypertrophy
Atrial natriuretic peptide (ANP), the two forms of
myosin heavy chain (α-MHC and β-MHC) and α-actin, which are
considered as molecular markers of cardiac hypertrophy (29), were evaluated. There was a significant
increase in ANP expression in the SE-GH group compared with in the
SC group (3.20-fold, P<0.05). β-MHC was increased in the SE-GH
group (6.76-fold, P<0.05) and decreased in the SE group
(0.34-fold, P<0.05) compared with in the control group (Fig. 1;
α-MHC and α-actin not shown).
Expression analysis of genes in the
PI3K/AKT/mTOR and ERK signaling pathways
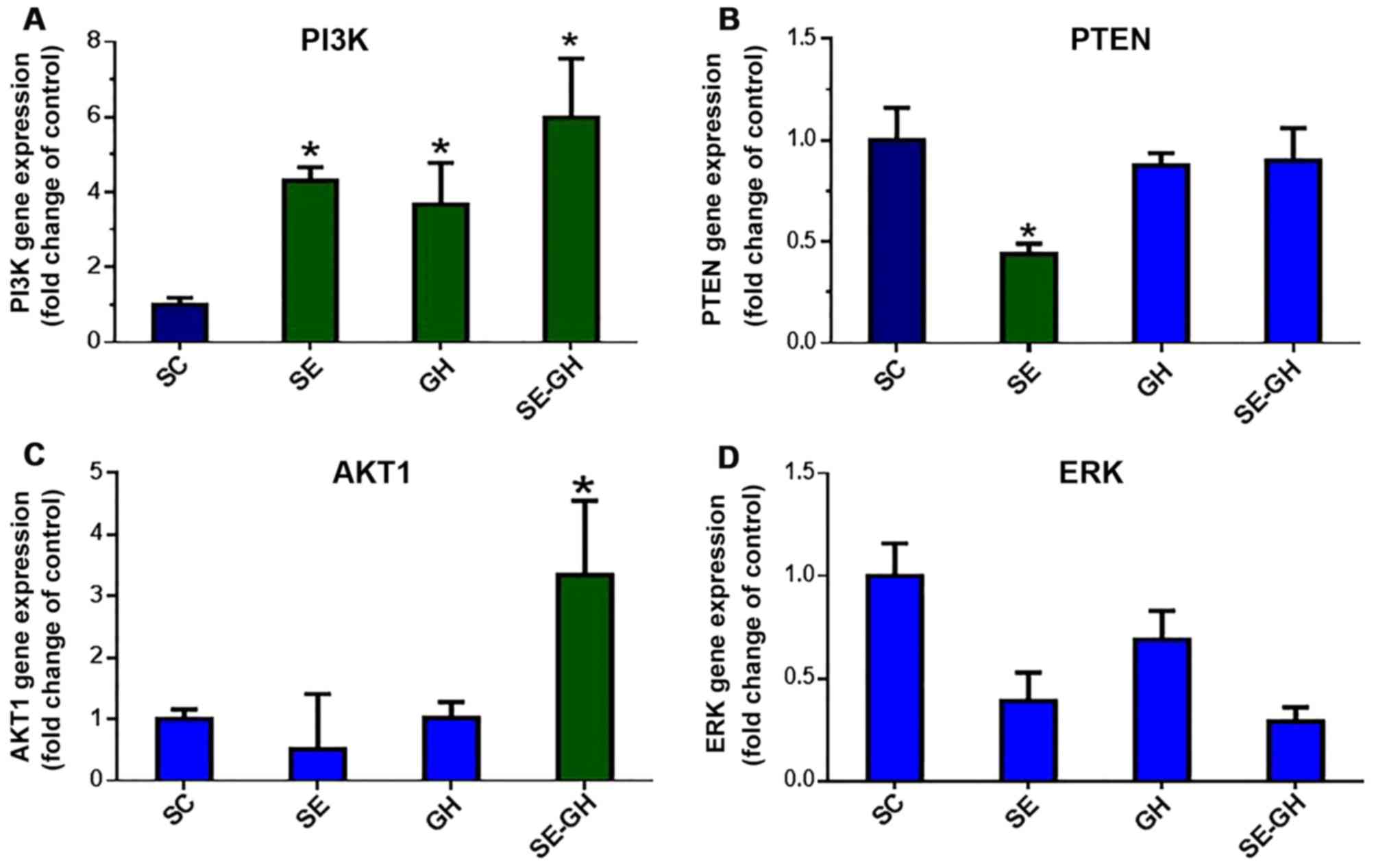
PCR results. PI3K gene expression, which is
established to serve an essential role in regulating cardiac growth
(7) was significantly higher in the
SE-GH group (5.98-fold, P<0.05) compared with in the SC group.
Significant increases were also observed in the SE and GH groups
(4.30- and 3.67-fold, P<0.05). PTEN is a major negative
regulator of PI3K gene expression (7). The present data demonstrated that PTEN
was significantly decreased in the SE group (0.44-fold, P<0.05)
compared with in the SC group; while no significant difference was
indicated when the two GH-treated groups were compared with the SC
group (Fig. 2).
AKT-1 gene expression, which is involved in the
regulation of protein synthesis and is a major suppressor of PI3K
(7), was significantly increased in
the SE-GH group (3.35-fold, P<0.05) compared with in the SC
group. The SE and GH groups were similar to the SC group in terms
of AKT-1 expression (Fig. 2).
Gene expression of ERK1/2 was unchanged with
exercise and/or r-hGH administration and remained comparable across
all groups (Fig. 2).
Immunohistochemical results
PI3Kα staining grades were significantly different
across the groups. The difference was a result of significantly
increased PI3K staining in left ventricular tissue of the SE, GH
and SE-GH groups compared with that in the SC group (Fig. 3). PTEN staining grades were similar
across all groups (not shown).
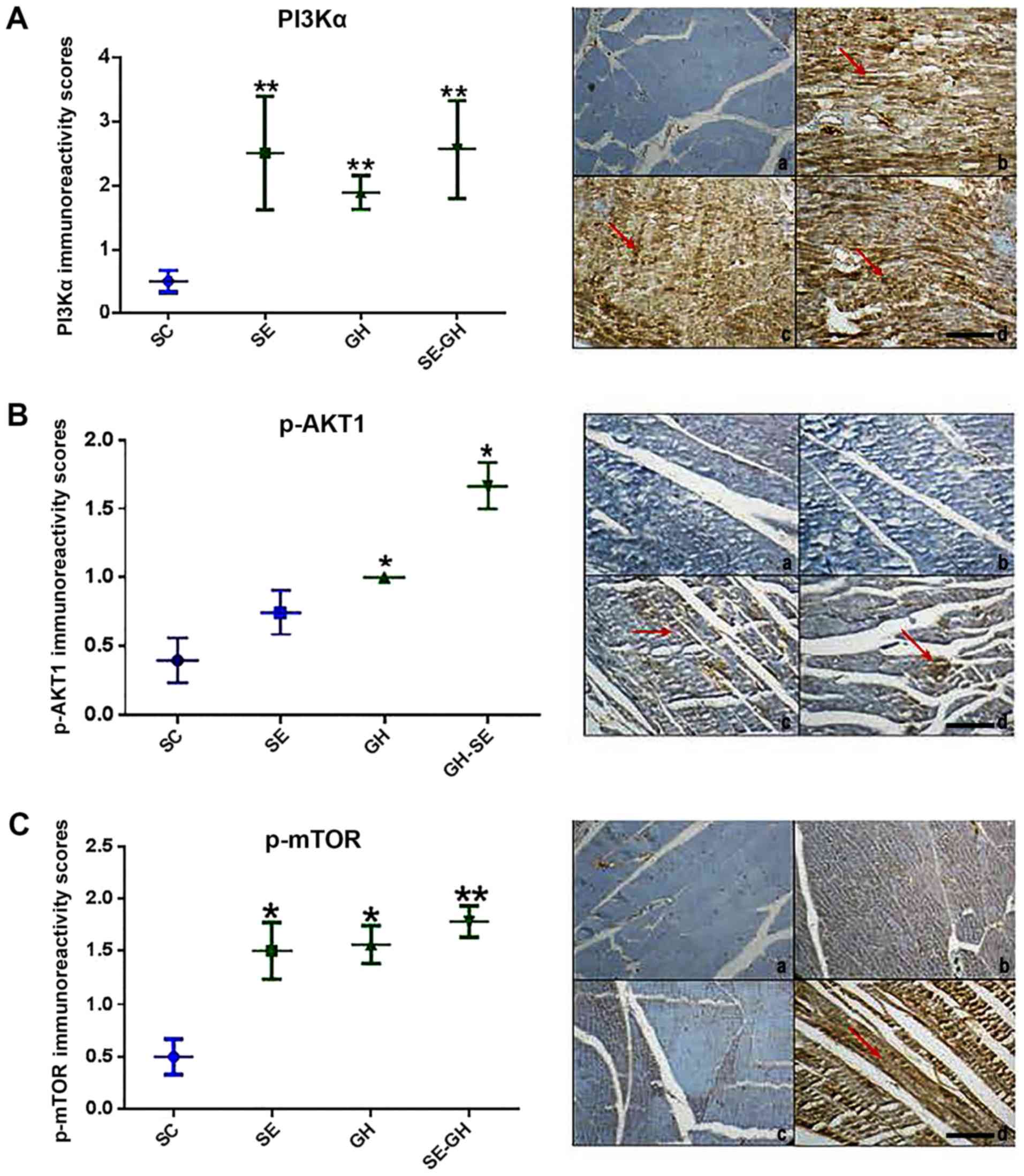 | Figure 3.Immunoperoxidase assay of the
PI3K/AKT/mTOR signaling pathway. (A) Cardiac P13Kα antibody. (A-a)
No staining was detected in the SC group. (A-b) Diffuse strong
positive staining was detected in the SE, (A-c) GH and (A-d) SE-GH
groups. (B) p-AKT antibody. (B-a) No staining was detected in the
SC and (B-b) SE groups. (B-c) Weak staining was detected in the
(B-d) SE-GH group. (C) p-mTOR antibody. (C-a) No staining was
detected in the SC group. (C-b) Diffuse-moderate staining was
detected in the SE group. (C-c) Focally moderate staining was
detected in the GH group and (C-d) diffuse-strong staining was
detected in the SE-GH group. Red arrows indicate areas where
staining was apparent. Original magnification, x200, scale bar, 100
µm. *P<0.05, **P<0.001. PI3Kα, phosphoinositide-3-kinase
catalytic α (polypeptide p-110); p-AKT, phospho-AKT (Ser473);
p-mTOR, phospho-mechanistic target of rapamycin (Ser2448-catalytic
domain); SC, sedentary control; SE, swimming exercise; GH, r-hGH
exposure; SE-GH, swimming exercise-r-hGH exposure; r-hGH,
recombinant human growth hormone. |
In terms of AKT, the groups revealed statistically
significant differences, with increased p-AKT-1 in the GH and SE-GH
groups compared with in the SC group (P<0.05). Regarding p-mTOR,
significant increases were observed in all test groups [SE
(P<0.05), GH (P<0.05) and SE-GH (P<0.01)] compared with SC
(Fig. 3).
No significant differences were noted in ERK1/2
protein levels across the groups (not shown).
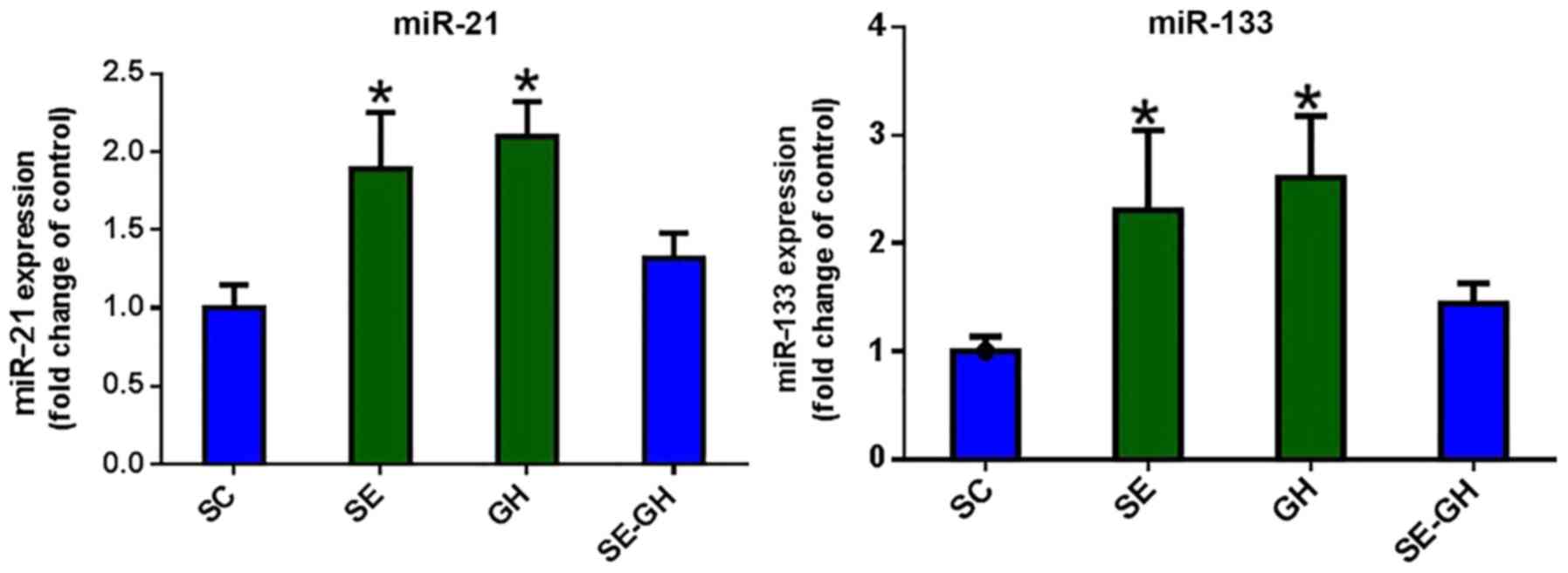
miRNA analysis
miR-21 gene expression was determined to be
increased in the SE and GH groups, compared with the SC group
(P<0.05). When miR-133 levels were compared, significant
increases were observed in the SE and GH groups relative to levels
in the SC group (P<0.05). Expression levels of miR-21/133 in the
SE-GH group did not exhibit any significant change compared with
those in the SC group (Fig. 4).
Discussion
It is known to use r-hGH to improve performance and
muscle mass by some athletes (30).
However, no study to our knowledge has demonstrated the mechanism
through which r-hGH, together with swimming exercise, may affect
signaling pathways (PI3K/AKT/mTOR and ERK) that regulate cardiac
signaling genes and associated miRNAs (miR-21 and miR-133)
(31). A key finding of the present
study was the altered expression of genes in the PI3K/AKT/mTOR
signaling pathway with r-hGH administration and swimming exercise.
With this, the present study demonstrated that r-hGH administration
may trigger the similar signal transduction pathways in ventricular
muscle. However, no change was detected between the groups in terms
of ERK1/2 gene expression, which is involved in the ERK signaling
pathway that is active during cardiac pathological hypertrophy
(Fig. 5).
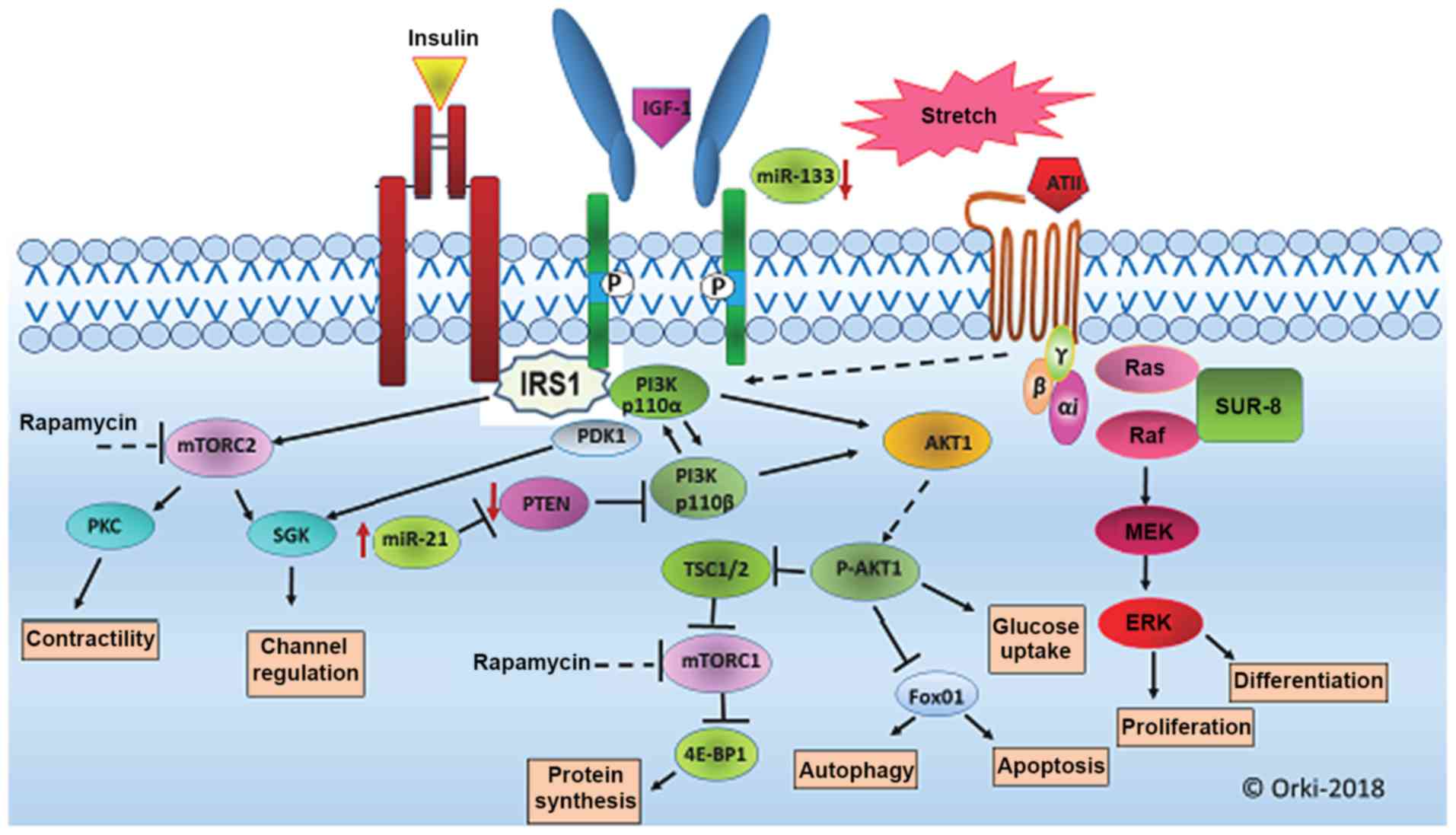 | Figure 5.PI3K/AKT/mTOR and ERK signalling
pathways associated with cardiac hypertrophy. Signal transduction
pathways critical for myosin growth include PI3K/AKT/mTOR,
MAPK/ERK. The PI3K/AKT/mTOR signaling pathway has been reported to
serve a regulatory role in cell growth, metabolism, survival and
angiogenesis, and to govern the development and transformation of
left ventricular hypertrophy. The ERK signaling pathway, the
members of which belong to the MAPK family, comprises a cascade of
a series of successively acting kinases that finally lead to
phosphorylation and activation of terminal kinases including p38,
c-Jun N-terminal kinases and ERK. miR-133 has a critical role in
determining cardiomyocyte hypertrophy and miR-21 has also been
implicated in the regulation of cardiac fibrosis and cardiac muscle
contractility, and has been shown to negatively regulate
phosphatase and tensin homolog (PTEN) gene expression. PI3K,
phosphoinositide-3-kinase; AKT, serine/threonine protein kinase;
mTOR, mechanistic target of rapamycin; MAPK, mitogen-activated
protein kinase; ERK, extracellular signal-regulated kinase; Ang II,
angiotensin II; DAG, diacylglycerol; Et, endothelin; FGF,
fibroblast growth factor; G, G protein; IGF, insulin-like growth
factor; IP3, inositol 1,4,5-triphosphate; MEK, MAP kinase kinase;
NFAT, nuclear factor of activated T cells; PKC, protein kinase C;
PLC, phospholipase C; RAF, mitogen-activated protein (MAP) kinase
kinase kinase; RAS, monomeric GTPase. |
Dissimilar to the present study, previous studies
planned GH administration based on the therapeutic approach in
children or adults with GH deficiency (4). Various studies have been performed on
the use of r-hGH as a doping hormone in sports (5,6). However,
the present study is seemingly the first to investigate the effects
of r-hGH administration in conjunction with swimming exercise on
PI3K/AKT/ mTOR and ERK signaling pathways, known to regulate
cardiac hypertrophy, and on miRNAs associated with cardiac
hypertrophy (miR-21 and miR-133) in rats. In the present study,
r-hGH was administered at 0.3 mg/kg (1 IU/day) concurrently with
swimming exercise 5 times a week for 8 weeks.
A limitation of the current study is the evaluation
of one dose level, when different doses may demonstrate the
potential harmful effects of r-hGH. We plan to assess the effects
of higher dose r-hGH treatment in the future.
Heart weight/ body weight, myocyte diameter and
sedentary bradycardia are among the determinants of physiological
left ventricular hypertrophy (7,22,28). The current study data demonstrated
left ventricular hypertrophy formation based on increase in to
heart weight/ body weight ratio (HWI) in SE and SE-GH rats.
ANP, β-MHC and α-actin levels have been reported as
molecular markers of pathological hypertrophy (32,33).
Increased expression of ANP and β-MHC was found in the SE-GH group
in the current study; while relatively small changes were
determined in ANP gene expression. No changes in these factors were
identified in the SE and GH groups. It is noteworthy that markedly
greater increases in ANP gene expression have been reported under
conditions of acute pressure or volume overload (10- to 20-fold
increase), which may represent a different pathogenetic response
(29).
Previous studies have shown that activation of the
PI3K/AKT signaling cascade results in myocardial hypertrophy
(33,34). In a cardiac hypertrophy study
utilizing aortic narrowing surgery, PI3K-negative transgenic mice
(dnPI3K) were determined to have smaller hearts (appearance and
weight) compared with controls. Additionally, a significant
increase in heart weight was observed among dNPI3K mice compared
with the control group in response to placement of an aortic band
and swimming exercise (8). However,
it has been suggested that PI3K signaling is not essential for
pathological hypertrophy development induced by increased pressure,
but is important for physiological cardiac hypertrophy development
induced by swimming exercise (34).
In the present study, PI3K gene and PI3Kα protein expression levels
were increased to a greater extent in the SE-GH group compared with
levels in the other groups. Furthermore, HWI values were
significantly higher in the SE-GH group. This increase appeared to
be a result of both the swimming exercise and r-hGH administration.
This suggests that r-hGH administration for 8 weeks does not cause
as much damage as pathological cardiac hypertrophy induced by an
aortic band, but differs from the physiological hypertrophy induced
by swimming exercise. Greater understanding of the mechanism
involved requires further studies performed with higher doses of
r-hGH over longer durations.
PTEN is established as a major negative regulator of
the PI3K/AKT signaling pathway (35).
Interestingly, heart weight has been reported to be increased (by
50%) in PTEN (null) rats (36).
Certain studies have demonstrated negative regulation of PTEN gene
expression by miR-21(31). miR-21 has
also been shown to serve a role in cardiomyocyte hypertrophy
(37). In a study by Ma et al
(7), PTEN expression was
significantly decreased at the gene and protein levels in the SE
group compared with in the control group, and the expression of
PTEN-targeting miR-21 was also observed to be significantly
increased. Similarly, in the present study, a significant decrease
in PTEN was observed in the SE group compared with in the control
group, concomitant with a significant increase in miR-21 gene
expression. However, in the GH and SE-GH groups, no difference was
observed in PTEN gene expression compared with the control, while
miR-21 was significantly increased in the GH group; the miR-21
expression was similar to that of controls in the SE-GH group.
These findings suggests that r-hGH administration may impair the
regulation between miR-21 and its target PTEN.
AKT-1 has been demonstrated to be necessary for the
development of physiological hypertrophy induced by exercise
(38) and for the regulation of
normal cardiac growth (39). Previous
reports have concluded that cardiac AKT-1 may regulate cardiac
hypertrophy when studying models of cardiac hypertrophy or ischemia
(32,38). In the study of Ma et al (7), it was concluded that AKT1 upregulated
PI3Kα through phosphorylation in left ventricular hypertrophy
induced by swimming exercise. Left ventricular hypertrophy (LVH)
induced by swimming exercise training is an important physiological
mechanism that compensates for chronic increases in hemodynamics
(7). Its comparison to the previously
reported Akt2 deficiency phenotype reveals the non-redundant
functions of Akt1 and Akt2 genes with respect to organismal growth
and insulin-regulated glucose metabolism (39). To assess the role of AKT1, the present
study tested AKT1 gene and p-AKT1 protein expression. A significant
increase in AKT1 gene expression was observed in the SE-GH group
compared with in the controls, and p-AKT1 protein levels were
significantly higher in the GH and SE-GH groups than in the control
group. This demonstrates that AKT1 may increase the upregulation of
PI3Kα and result in the relatively highest elevation in PI3K gene
expression observed in the SE-GH group. Expression of the mTOR gene
has been associated with decreased cardiac hypertrophy through
inhibition of mTOR with rapamycin in humans and animal models
(34,40). In the current study,
phosphoser2448mTOR protein expression was determined to be
increased in SE, GH and SE-GH rats compared with in controls. These
results may aid in clarifying the cause of hypertrophy following
SE-GH exposure.
It has been reported that the activation of cardiac
myocytes in response to any stress stimulation occurs by means of
the ERK1/2 signaling pathway (8).
Exercise and pressure overload may activate the ERK1/2 signaling
pathway (10,39). Another study has suggested that the
hypertrophic response to exercise-induced load increase is not
reduced, although phosphorylation of ERK1/2 is essentially
eliminated, following the stimulation of increased pressure
(40,41). Additionally, it has been reported that
MAPK kinase-ERK1/2 inhibition does interfere with hypertrophic
morphology (19). In the present
study, it was observed that post-exercise development of left
ventricular hypertrophy did not differ between the groups in terms
of ERK1/2 signaling: ERK1/2 gene and protein expression was similar
across all groups. With this, the data indicates that the ERK1/2
signaling pathway may not be activated during physiological left
ventricular hypertrophy caused by swimming exercise and r-hGH
administration over 8 weeks.
miR-133, which is established to be highly expressed
in adult heart and skeletal muscles, serves a role in cell
differentiation and proliferation (42,43).
miR-133 has a critical role in determining cardiomyocyte
hypertrophy; its overexpression may inhibit hypertrophy whereas its
suppression may induces hypertrophy in vitro and in vivo (20). Previous study has shown that miR-133
may controls cardiac hypertrophy and decrease significantly in the
presence of cardiac hypertrophy and heart failure (44). Thus, the reduction of miR-133
expression may be involved in the development and progression of
chronic heart failure (45). The
present study identified increased miR-133 expression in the
ventricular tissue of SE and GH rats compared with controls.
However, the SE-GH group was similar to the controls with regard to
miR-133 expression.
In conclusion, the present study demonstrated that
r-hGH administration with swimming exercise over 8 weeks altered
PI3K, PTEN, AKT-1 and mTOR expression in rat heart. As these
factors are involved in the PI3K/AKT/mTOR signaling pathway, as a
major regulator of cell proliferation, metabolism, survival and
angiogenesis, these changes may have contributed to cardiac
hypertrophy. Additionally, r-hGH administration was observed to
impair the negative regulation between miR-21 and its target, PTEN.
Greater understanding of this mechanism requires further studies on
more genes and miRNAs involved in the PI3K/AKT/mTOR and ERK
signaling pathways.
Acknowledgments
Not applicable.
Funding
The current study was funded by Trakya University
Scientific Research Projects Unit (Project no. 2015-36; Edirne,
Turkey).
Availability of data and materials
The materials and data described in the manuscript
are available on reasonable request for non-commercial
purposes.
Authors' contributions
OP designed the experiments, and coordinated the
project and interpreted results. SAV interpreted the results. ZBD
performed molecular studies and interpreted results. ET was
responsible for morphological evaluations and interpretation of
results. PT and AD were involved in the animal protocols and
determination of blood parameters.
Ethics approval and consent to
participate
The study was conducted in the Trakya University
Local Experimental Animals Research Center after obtaining approval
from the Local Experimental Animals Ethics Committee. (approval no.
TUHADYEK:2015/10).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Graham MR, Baker JS, Evans P, Kicman A,
Cowan D, Hullin D, Thomas N and Davies B: Physical effects of
short-term recombinant human growth hormone administration in
abstinent steroid dependency. Horm Res. 69:343–354. 2008.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Jenkins PJ: Growth hormone and exercise.
Clin Endocrinol (Oxf). 50:683–689. 1999.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Castellano G, Affuso F, Conza PD and Fazio
S: The GH/IGF-1 Axis and Heart Failure. Curr Cardiol Rev.
5:203–215. 2009.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Colao A, Marzullo P, Di Somma C and
Lombardi G: Growth hormone and the heart. Clin Endocrinol (Oxf).
54:137–154. 2001.PubMed/NCBI View Article : Google Scholar
|
|
5
|
World Anti-Doping Agency: The 2010
Prohibited List. International Standard. https://www.wada-ama.org/sites/default/files/resources/files/WADA_Prohibited_List_2010_EN.pdf.
Accessed September 19. 2009.
|
|
6
|
Voss SC, Giraud S, Alsayrafi M, Bourdon
PC, Schumacher YO, Saugy M and Robinson N: The effect of a period
of intensive exercise on the isoform test to detect growth hormone
doping in sports. Growth Horm IGF Res. 23:105–108. 2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ma Z, Qi J, Meng S, Wen B and Zhang J:
Swimming exercise training-induced left ventricular hypertrophy
involves microRNAs and synergistic regulation of the PI3K/AKT/mTOR
signaling pathway. Eur J Appl Physiol. 113:2473–2486.
2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Heineke J and Molkentin JD: Regulation of
cardiac hypertrophy by intracellular signalling pathways. Nat Rev
Mol Cell Biol. 7:589–600. 2006.PubMed/NCBI View
Article : Google Scholar
|
|
9
|
Guertin DA and Sabatini DM: Defining the
role of mTOR in cancer. Cancer Cell. 12:9–22. 2007.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Sugden PH and Clerk A: ‘Stress-responsive’
mitogen-activated protein kinases (c-Jun N-terminal kinases and p38
mitogen-activated protein kinases) in the myocardium. Circ Res.
83:345–352. 1998.PubMed/NCBI
|
|
11
|
Purcell NH, Wilkins BJ, York A,
Saba-El-Leil MK, Meloche S, Robbins J and Molkentin JD: Genetic
inhibition of cardiac ERK1/2 promotes stress-induced apoptosis and
heart failure but has no effect on hypertrophy in vivo. Proc Natl
Acad Sci USA. 104:14074–14079. 2007.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Hutvágner G and Zamore PD: A microRNA in a
multiple-turnover RNAi enzyme complex. Science. 297:2056–2060.
2002.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Bao H, Hu S, Zhang C, Shi S, Qin W, Zeng
C, Zen K and Liu Z: Inhibition of miRNA-21 prevents fibrogenic
activation in podocytes and tubular cells in IgA nephropathy.
Biochem Biophys Res Commun. 444:455–460. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Da Costa Martins PA and De Windt LJ:
Targeting microRNA targets. Circ Res. 111:506–508. 2012.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Cheng YS, Tang YQ, Dai DZ and Dai Y: AQP4
knockout mice manifest abnormal expressions of calcium handling
proteins possibly due to exacerbating pro-inflammatory factors in
the heart. Biochem Pharmacol. 83:97–105. 2012.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Da Costa Martins PA and De Windt LJ:
MicroRNAs in control of cardiac hypertrophy. Cardiovasc Res.
93:563–572. 2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wang L, Li X, Zhou Y, Shi H, Xu C, He H,
Wang S, Xiong X, Zhang Y, Du Z and et al: Downregulation of miR-133
via MAPK/ERK signaling pathway involved in nicotine-induced
cardiomyocyte apoptosis. Naunyn Schmiedebergs Arch Pharmacol.
387:197–206. 2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Dong DL, Chen C, Huo R, Wang N, Li Z, Tu
YJ, Hu JT, Chu X, Huang W and Yang BF: Reciprocal repression
between microRNA-133 and calcineurin regulates cardiac hypertrophy:
A novel mechanism for progressive cardiac hypertrophy.
Hypertension. 55:946–952. 2010.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Dong S, Ma W, Hao B, Hu F, Yan L, Yan X,
Wang Y, Chen Z and Wang Z: microRNA-21 promotes cardiac fibrosis
and development of heart failure with preserved left ventricular
ejection fraction by up-regulating Bcl-2. Int J Clin Exp Pathol.
7:565–574. 2014.PubMed/NCBI
|
|
21
|
DA Silva ND Jr, Fernandes T, Soci UP,
Monteiro AW and Phillips MI and DE Oliveira EM: Swimming training
in rats increases cardiac MicroRNA-126 expression and angiogenesis.
Med Sci Sports Exerc. 44:1453–1462. 2012.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Oliveira EM, Sasaki MS, Cerêncio M,
Baraúna VG and Krieger JE: Local renin-angiotensin system regulates
left ventricular hypertrophy induced by swimming training
independent of circulating renin: A pharmacological study. J Renin
Angiotensin Aldosterone Syst. 10:15–23. 2009.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Hirata Y, Murai N, Yanaihara N, Saito M,
Saito M, Urashima M, Murakami Y, Matsufuji S and Okamoto A:
MicroRNA-21 is a candidate driver gene for 17q23-25 amplification
in ovarian clear cell carcinoma. BMC Cancer. 14(799)2014.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Iijima Y, Seike M, Noro R, Ibi T, Takeuchi
S, Mikami I, Koizumi K, Usuda J and Gemma A: Prognostic
significance of PIK3CA and SOX2 in Asian patients with lung
squamous cell carcinoma. Int J Oncol. 46:505–512. 2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhou X, Tan M, Stone Hawthorne V, Klos KS,
Lan KH, Yang Y, Yang W, Smith TL, Shi D and Yu D: Activation of the
Akt/mammalian target of rapamycin/4E-BP1 pathway by ErbB2
overexpression predicts tumor progression in breast cancers. Clin
Cancer Res. 10:6779–6788. 2004.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Domenighetti AA, Danes VR, Curl CL,
Favaloro JM, Proietto J and Delbridge LM: Targeted GLUT-4
deficiency in the heart induces cardiomyocyte hypertrophy and
impaired contractility linked with Ca(2+) and proton flux
dysregulation. J Mol Cell Cardiol. 48:663–672. 2010.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chung E and Diffee GM: Effect of aging on
power output properties in rat skinned cardiac myocytes. J Gerontol
A Biol Sci Med Sci. 66:1267–1273. 2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Vikstrom KL, Bohlmeyer T, Factor SM and
Leinwand LA: Hypertrophy, pathology, and molecular markers of
cardiac pathogenesis. Circ Res. 82:773–778. 1998.PubMed/NCBI
|
|
30
|
Saugy M, Robinson N, Saudan C, Baume N,
Avois L and Mangin P: Human growth hormone doping in sport. Br J
Sports Med. 40((Suppl 1)): i35–i39. 2006.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Cheng Y, Ji R, Yue J, Yang J, Liu X, Chen
H, Dean DB and Zhang C: MicroRNAs are aberrantly expressed in
hypertrophic heart: Do they play a role in cardiac hypertrophy? Am
J Pathol. 170:1831–1840. 2007.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Soci UP, Fernandes T, Hashimoto NY, Mota
GF, Amadeu MA, Rosa KT, Irigoyen MC, Phillips MI and Oliveira EM:
MicroRNAs 29 are involved in the improvement of ventricular
compliance promoted by aerobic exercise training in rats. Physiol
Genomics. 43:665–673. 2011.PubMed/NCBI View Article : Google Scholar
|
|
33
|
McMullen JR and Jennings GL: Differences
between pathological and physiological cardiac hypertrophy: Novel
therapeutic strategies to treat heart failure. Clin Exp Pharmacol
Physiol. 34:255–262. 2007.PubMed/NCBI View Article : Google Scholar
|
|
34
|
McMullen JR, Shioi T, Zhang L, Tarnavski
O, Sherwood MC, Kang PM and Izumo S: Phosphoinositide
3-kinase(p110alpha) plays a critical role for the induction of
physiological, but not pathological, cardiac hypertrophy. Proc Natl
Acad Sci USA. 100:12355–12360. 2003.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Maehama T and Dixon JE: PTEN: A tumour
suppressor that functions as a phospholipid phosphatase. Trends
Cell Biol. 9:125–128. 1999.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Crackower MA, Oudit GY, Kozieradzki I,
Sarao R, Sun H, Sasaki T, Hirsch E, Suzuki A, Shioi T, Irie-Sasaki
J and et al: Regulation of myocardial contractility and cell size
by distinct PI3K-PTEN signaling pathways. Cell. 110:737–749.
2002.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Bai L, Liang R, Yang Y, Hou X, Wang Z, Zhu
S, Wang C, Tang Z and Li K: MicroRNA-21 Regulates PI3K/Akt/mTOR
Signaling by Targeting TGFβI during Skeletal Muscle Development in
Pigs. PLoS One. 10(e0119396)2015.PubMed/NCBI View Article : Google Scholar
|
|
38
|
DeBosch B, Treskov I, Lupu TS, Weinheimer
C, Kovacs A, Courtois M and Muslin AJ: Akt1 is required for
physiological cardiac growth. Circulation. 113:2097–2104.
2006.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Cho H, Thorvaldsen JL, Chu Q, Feng F and
Birnbaum MJ: Akt1/PKBalpha is required for normal growth but
dispensable for maintenance of glucose homeostasis in mice. J Biol
Chem. 276:38349–38352. 2001.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Shioi T, McMullen JR, Tarnavski O,
Converso K, Sherwood MC, Manning WJ and Izumo S: Rapamycin
attenuates load-induced cardiac hypertrophy in mice. Circulation.
107:1664–1670. 2003.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Leontieva OV, Paszkiewicz GM and
Blagosklonny MV: Mechanistic or mammalian target of rapamycin
(mTOR) may determine robustness in young male mice at the cost of
accelerated aging. Aging (Albany NY). 4:899–916. 2012.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Chen JF, Mandel EM, Thomson JM, Wu Q,
Callis TE, Hammond SM, Conlon FL and Wang DZ: The role of
microRNA-1 and microRNA-133 in skeletal muscle proliferation and
differentiation. Nat Genet. 38:228–233. 2006.PubMed/NCBI View
Article : Google Scholar
|
|
43
|
Horie T, Ono K, Nishi H, Iwanaga Y, Nagao
K, Kinoshita M, Kuwabara Y, Takanabe R, Hasegawa K, Kita T and et
al: MicroRNA-133 regulates the expression of GLUT4 by targeting
KLF15 and is involved in metabolic control in cardiac myocytes.
Biochem Biophys Res Commun. 389:315–320. 2009.PubMed/NCBI View Article : Google Scholar
|
|
44
|
van Rooij E, Sutherland LB, Liu N,
Williams AH, McAnally J, Gerard RD, Richardson JA and Olson EN: A
signature pattern of stress-responsive microRNAs that can evoke
cardiac hypertrophy and heart failure. Proc Natl Acad Sci USA.
103:18255–18260. 2006.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Abdellatif M: The role of microRNA-133 in
cardiac hypertrophy uncovered. Circ Res. 106:16–18. 2010.PubMed/NCBI View Article : Google Scholar
|