Introduction
Neuropathic pain is an intractable affliction caused
as a result of various forms of damage to the nervous system. The
mechanisms underlying the development of neuropathic pain are yet
to be elucidated. Several previous studies using animal models have
suggested candidates that may be therapeutic targets for the
reduction of neuropathic pain. However, there are currently no good
pharmacotherapies for neuropathic pain (1). Substantial evidence has demonstrated
that central neuroimmune activation and neuroinflammation have
crucial roles in the generation and maintenance of chronic pain
(2). Subsequent to nerve injury,
microglia in the spinal cord become activated and pro-inflammatory
cytokines are released, including tumor necrosis factor (TNF)-α and
interleukin (IL)-1β. In addition, the activation of glia is
accompanied by the release of chemical mediators, including
substance P, inflammatory cytokines and chemokines, which
subsequently enhance the activation of spinal glia (3–7). The
suppression of microglia activity is has been explored as a
potential therapy for a number of central nervous system (CNS)
inflammatory diseases and neuropathic pain (8).
Toll like receptors (TLRs) are transmembrane
signaling molecules that participate in innate immune recognition
(9). Previous evidence has suggested
that TLR-4 is expressed by microglial cells in the CNS, and that
the activation of these cells is associated with the release of
pro-inflammatory cytokines (10).
Thus, the inhibition of TLR-4 may reduce inflammatory responses and
ameliorate the pain experienced after nerve-injury.
Peroxisome proliferator-activated receptor gamma
(PPAR-γ) is transcription factor that belongs to the nuclear
hormone receptor family. PPAR-γ agonists are currently employed
clinically for the management of type II diabetes mellitus.
Furthermore, previous studies have shown that PPAR-γ has the
potential to ameliorate the symptoms of nervous system-associated
disorders, such as spinal cord injury, amyotrophic lateral
sclerosis, allergic encephalomyelitis, Parkinson's disease,
cerebral hemorrhage and Alzheimer's disease (11–16).
However, studies concerning the underlying mechanism of PPAR-γ
agonists on neuroimmune activation associated with neuropathic
pain, are limited. Pioglitazone (Pio) is a promising PPAR-γ
antagonist, which may penetrate the blood-brain barrier to exert
its actions on the CNS (17).
Therefore, using a rat model of chronic constriction injury, the
current study was conducted to evaluate the role of orally
administered Pio on pain behaviors, inflammatory cytokines, glial
activation and the expression of TLR-4. The current study also
aimed to determine whether the analgesic effect of Pio was
associated with its impact on the expression of cytokines and
TLR-4, as well as glial activation markers involved in neuroimmune
activation.
Materials and methods
Experimental animals and
procedures
Male Sprague-Dawley rats (age, 45–50 days) weighing
between 200 and 250 g were housed in an approved facility and
maintained at 20–25°C, 50% humidity and under a 12-h dark:light
cycle, with food and water available ad libitum. All animal
experiments were performed in accordance with the National
Institutes of Health Guide regarding the care and use of animals
for experimental procedures. Efforts were made to limit distress
and to use the minimum number of rats necessary to achieve
statistical significance. The experimental protocol was reviewed
and approved by the Institutional Animal Care Committee of Jinling
Hospital (Nanjing, China).
Surgery to produce chronic constriction injury (CCI)
was first described by Bennett and Xie (18). Subsequent to the administration of
urethane anesthesia (1 g/kg, ip) the right sciatic nerve was
exposed, and proximal to the trifurcation, ~7 mm of the common
sciatic nerve was freed from the adherent tissue. Four 4-0 chromic
gut sutures, were tied around the nerve at intervals of ~1 mm, and
ligatures were secured loosely to ensure that, on visual
inspection, blood flow was not obstructed. The left sciatic nerve
was used as a control, and was exposed yet not ligated, whereas the
surgical incision was sutured and postsurgical recuperation was
monitored daily, and the rats recovered uneventfully. Particular
attention was applied to prevent infection, thus minimizing the
influence of inflammation. Rats were randomized into four equal
groups (n=8 per group) as follows: i) Sham-operation with vehicle
(0.5% carboxymethyl cellulose or Pio; sham group); ii) chronic
constriction injury with 2 ml/kg vehicle (vehicle group) or iii) 10
mg/kg Pio (Pio group); and iv) chronic constriction injury with
coadministration of 10 mg/kg Pio and 2 mg/kg GW9662 (a PPAR-γ
antagonist) administered by intraperitoneal (i.p.) injection (Pio +
GW group). The vehicle solution [0.5% CMC or Pio (Medical and
Pharmacological Company of Hengrui, China) was orally administered
1 h prior to surgery and continued daily to day 21
post-surgery.
Behavioral assessments
Mechanical hyperalgesia
Paw withdrawal mechanical threshold (PWMT) was
determined by applying a von Frey hair filament (Stoelting Co.,
Chicago, IL, USA) to the hind paw until a positive indicator of
pain behavior was elicited (19).
Evaluation thresholds were as follows: The mid-plantar paw was
assessed in the area of the sciatic nerve, avoiding the footpads.
The von Frey filaments with logarithmically incremental stiffness
(0.4–15.1 g) were applied serially to the paw via the up-down
method. The hairs were presented in ascending order of strength,
perpendicular to the plantar surface with sufficient force to cause
slight bending against the paw, and were held for 6–8 sec. A
positive response was recorded if the paw was sharply withdrawn.
Flinching immediately upon removal of the hair was also considered
a positive response. A 15.1 g hair was selected as the upper limit
for testing. If there was no response at 15.1 g pressure, rats were
assigned the upper limit value. A bending force able to evoke 50%
of the paw withdrawal occurrence was set as the PWMT.
Cold allodynia
Behavioral indicators of cold allodynia were
identified by immersing the tails of rats in cold water (4°C). As
previously described (20), each
animal was restrained in a plastic holder and its tail was allowed
to hang over the edge to facilitate the application of cold water
stimuli. Following the immersion of the tail, the latency of an
abrupt tail movement was measured, with a cut-off time of 15 sec.
Tail immersion assessment was repeated five times at 5-min
intervals. When calculating the average latency, the cut-off time
was assigned to rats with normal responses (an absence of tail
movement for 15 sec). Average latency was employed as a measure for
the severity of cold allodynia; a shorter latency was interpreted
as more severe cold allodynia.
Harvesting of tissue samples
Subsequent to the completion of the behavioral
assessment on day 21 post-operation, rats were anesthetized with an
overdose of sodium pentobarbital (70 mg/kg), the rat hearts were
exposed by thoracotomy and intracardially perfused with 250 ml cold
saline. Rats were then sacrificed by decapitation. The lumbar
spinal cord tissue proximal to the injury site was harvested and
kept on ice. The dorsal aspect of the spinal cord was dissected
from the ventral portion of the spinal cord. Following this, the
samples were removed quickly and stored in liquid nitrogen for
subsequent assays.
Reverse-transcription polymerase chain reaction
(RT-PCR)for the determination of IL-1β and TNF-α mRNA expression
levels
Total RNA was extracted with Tri Pure Isolation
Reagent (Roche Diagnostics, Basel, Switzerland) and quantified by
absorption at 260 nm. RNA samples all had OD260/280 ratios of
1.8–2.0, indicating good quality. RT was performed using Reverse
Transcription System (Promega Corp., Madison, WI, USA) according to
the manufacturer's protocol. β-actin as normalization control.
Primer sequences were as follows: IL-1β sense
5′-TGATGTTCCCATTAGACAGC-3′ and antisense
5′-GAGGTGCTGATGTACCAGTT-3′; TNF-α, sense 5′-CTCAGCCTCTTCTCCTTCCT-3′
and antisense 5′-ATGGGCTCATACCAGGGCTT-3′; and β-actin, sense
5′-CTGGCCGGGACCTGACTGACTACCTC-3′ and antisense
5′-AAACAAATAAAGCCATGCCAATCTCA-3′. The 100 µl reaction volume
consisted of 2 µl cDNA template (Promega Corporation, Madison, WI,
USA), 1.5 mmol/l MgCl2, 2.5 U Taq DNA polymerase, 100
µmol/l dNTP, 0.1 µmol/l primer and 1X Taq DNA polymerase
magnesium-free buffer (Promega Corp.). Next, the reaction mixture
was supplemented with two drops of mineral oil (Sigma-Aldrich;
Merck Millipore, Darmstadt, Germany) and incubated in a
thermocycler (Mini-Cycler PTC 150, MJ Research Inc., Hercules, CA,
USA) programmed to initially denature at 95°C for 2 min, then
denature at 95°C for 1 min, anneal at 60°C for 1 min and extend at
72°C for 2 min for a total of 30 cycles. The final cycle was
followed by incubation at 72°C for 5 min, and cooling to 4°C. The
PCR products were 206 bp (IL-1β), 191 bp (TNF-α) and 650 bp
(β-actin), respectively. Subsequently, PCR products were
electrophoresed on a 15 g/l agarose gel stained with ethidium
bromide. Digital images of the gel were captured and analyzed using
Scion Image v. 4.03 software (Scion Corporation; Informer
Technologies, Inc. Frederick, MD, USA). Values in each sample were
normalized with β-actin.
RT-PCR for the determination of TLR4 mRNA
expression levels
Total RNA was extracted with TriPure Isolation
Reagent and quantified by absorption at 260 nm. RT was performed
using a Promega Reverse Transcription System according to the
manufacturer's protocol. β-actin was used as an internal control
for normalization. Primer sequences were as follows: TLR4, sense
5′-AGCTTTGGTCAGTTGGCTCT-3′ and antisense
5′-CAGGATGACACCATTGAAGC-3′; and β-actin, sense
5′-CACCCGCGAGTACAACCTTC-3′ and antisense
5′-CCCATACCCACCATCACACC-3′. Cycling conditions were as follows:
95°C for 2 min, then 95°C for 1 min, 60°C for 1 min and 72°C for 2
min for 30 cycles. PCR amplification products were separated on an
ethidium bromide-stained 1.5% agarose gel, and visualized by
ultraviolet light. mRNA expression levels of TLR4 were calculated
as a percentage of the mRNA expression levels of β-actin in the
same tissue sample. Amplified bands showed the predicted sizes to
be as follows: TLR4 111 bp and β-actin 149 bp.
Immunohistochemistry
Immunoreactivity analysis of the microglial
activation marker Mac-1, or CR3/CD11b, was performed using
immunohistochemistry. To suppress endogenous peroxidase activity,
4-µm sections were treated with 0.3% H2O2 in
Tris-buffered saline (TBS) for 20 min at room temperature.
Following this, sections were incubated overnight at 4°C with
monoclonal mouse anti-Mac-1 (cat. no. 554980; 1:1,000; BD
Pharmingen, San Diego, CA, USA) in TBS with 3% normal goat serum
and 0.5% Triton-X-100. Subsequently, sections were incubated with
biotin-conjugated goat anti-mouse (cat. no. 115-065-003; 1:400;
Jackson ImmunoResearch, West Grove, PA) for 2 h at room
temperature, incubated with an avidin-biotin complex solution
(1:400; Vector Laboratories, Burlingame, CA, USA) for 1 h at room
temperature, followed by staining with 0.5 mg/ml
3,3′-diaminobenzidine tetrahydrochloride (Sigma-Aldrich). Finally,
sections were mounted on gelatin-coated slides, dried, dehydrated,
and coverslipped with Permount. Staining was evaluated by light
microscopy.
Statistical analysis
Values are expressed as means ± standard error of
the mean. For behavioral data, comparisons between groups were
performed using repeated measures of analysis of variance (ANOVA).
The effects of surgery and the administration of therapies on the
cytokines and TLR-4 were determined by one-way ANOVA followed by
Bonferroni or Tamhane's T2 test based on equal variances assumed or
otherwise. Data were analyzed using SPSS statistical software for
Windows (version 13.0; SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Behavioral assessments
Mechanical hyperalgesia and cold allodynia were
evidenced by PWMT (Fig. 1) and
response latency (RL) (Fig. 2)
findings. All rats demonstrated similar responsiveness to
mechanical and cold stimuli prior to surgery (Figs. 1 and 2). It was observed that CCI produced
significant mechanical hyperalgesia and cold allodynia in the
ipsilateral hind paw of rats on postoperative day 3, and these
effects persisted to postoperative day 21. Pio (10 mg/kg)
administration was able to attenuate the development of mechanical
hyperalgesia and cold allodynia in rats with CCI compared with
those who received the vehicle treatment on days 3, 7, 14 and 21
post-operation (P<0.01 for 10 mg/kg). When coadministered with
Pio, GW9662 (2 mg/kg, i.p.), an antagonist of PPAR-γ, was able to
reverse the analgesic effect of Pio (P<0.01; Figs. 1 and 2).
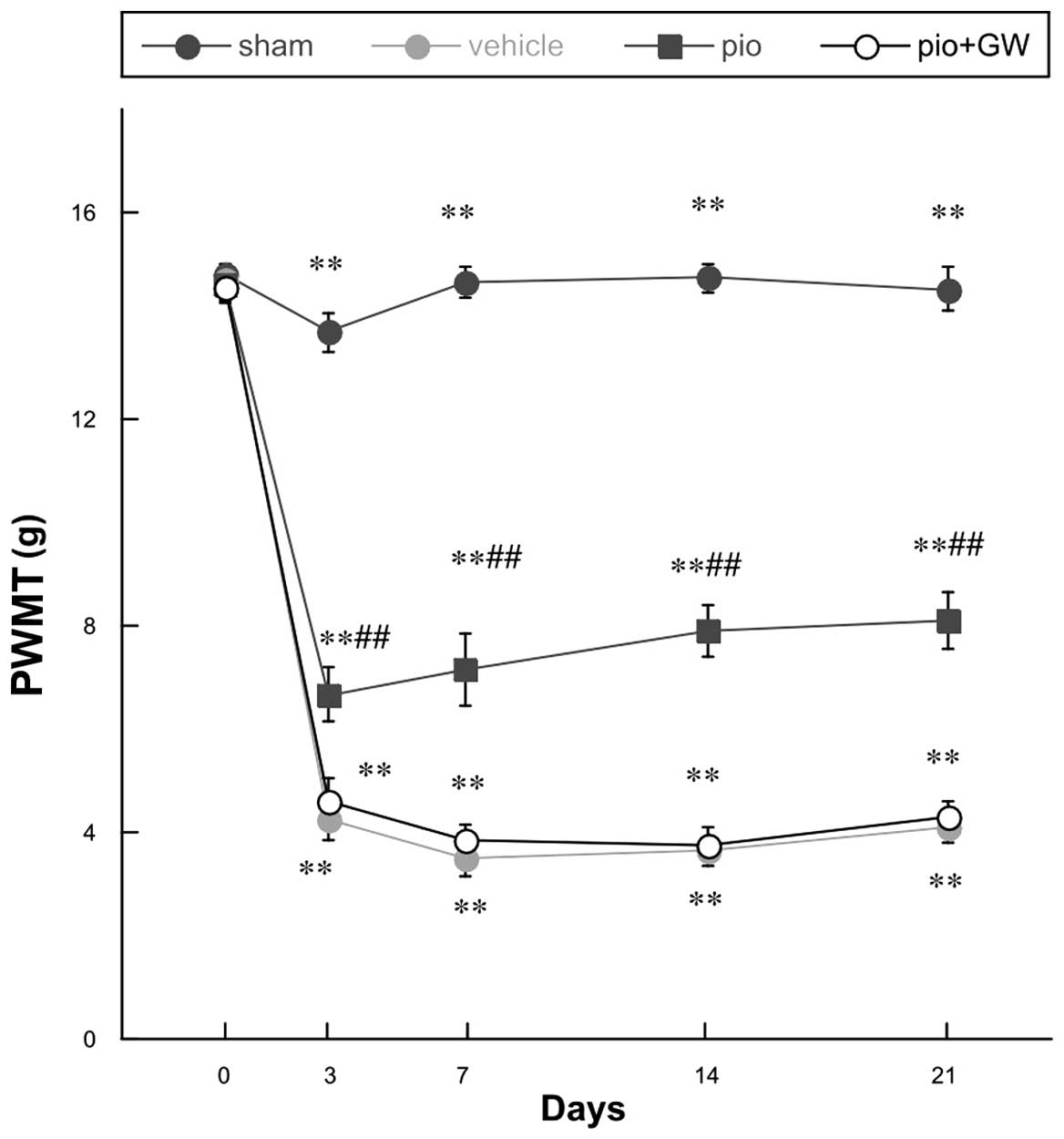 | Figure 1.Changes in the PWMT prior to surgery
and on days 0, 3, 7, 14 and 21 post-surgery. Compared with the sham
group, there was a significant decrease in PWMT following chronic
constriction injury (**P<0.01) in each of the remaining groups.
Pio administration attenuated the reduction in PWMT on
postoperative days 3, 7, 14 and 21 post-surgery compared with the
vehicle group (##P<0.01), and this effect was
reversed by co-administration with GW. PWMT, paw withdrawal
mechanical threshold; Pio, pioglitazone; GW, GW9662. |
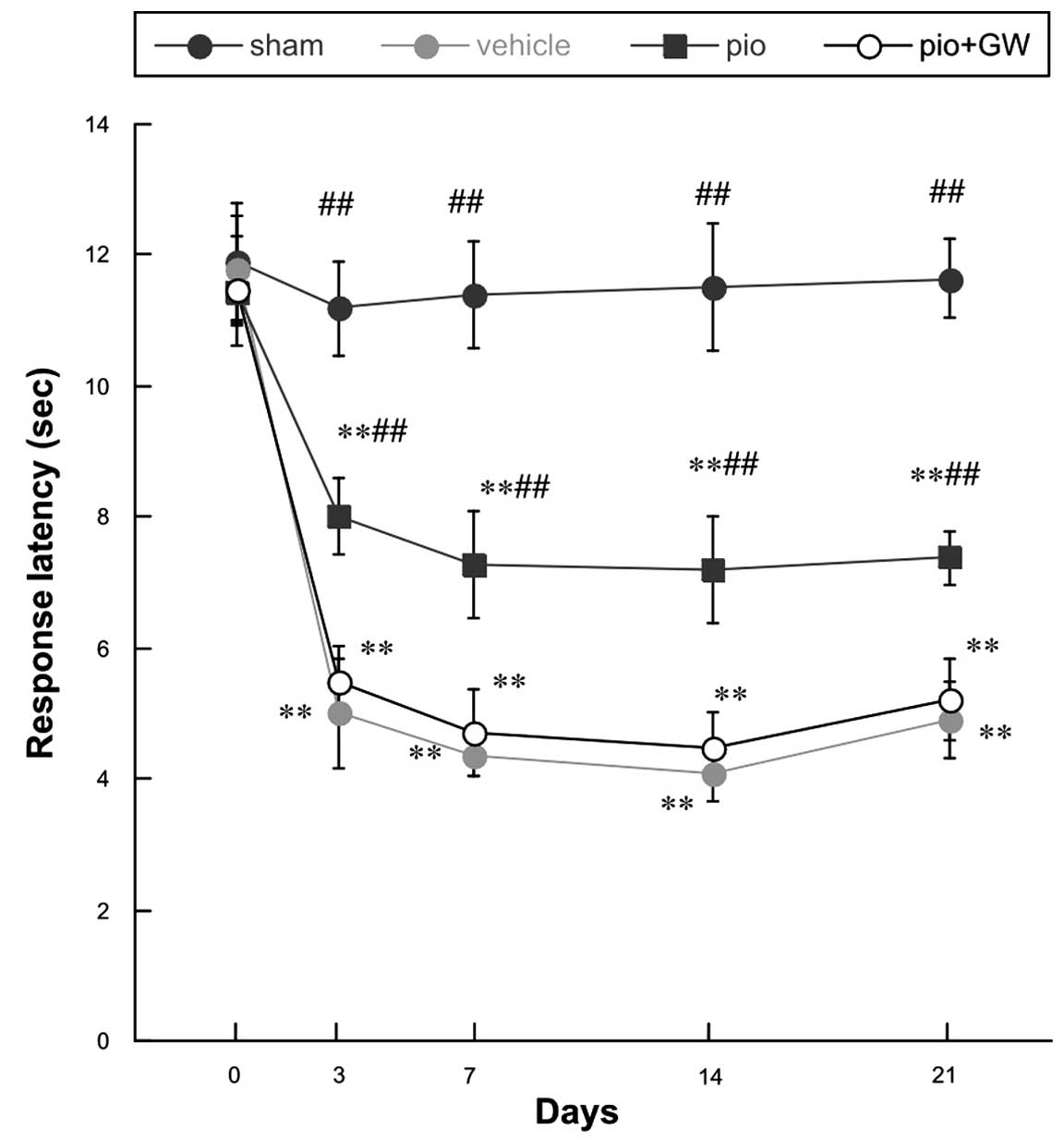 | Figure 2.Changes in the response latency prior
to surgery and on days 0, 3, 7, 14 and 21 post-surgery. Compared
with the sham group, there was a significant decrease in RL
following chronic constriction injury (**P<0.01). Pio
administration attenuated the reduction in RL on postoperative days
3, 7, 14 and 21 post-surgery, compared with the vehicle group
(##P<0.01), and this effect was reversed by
co-administration with GW. Pio, pioglitazone; GW, GW9662. |
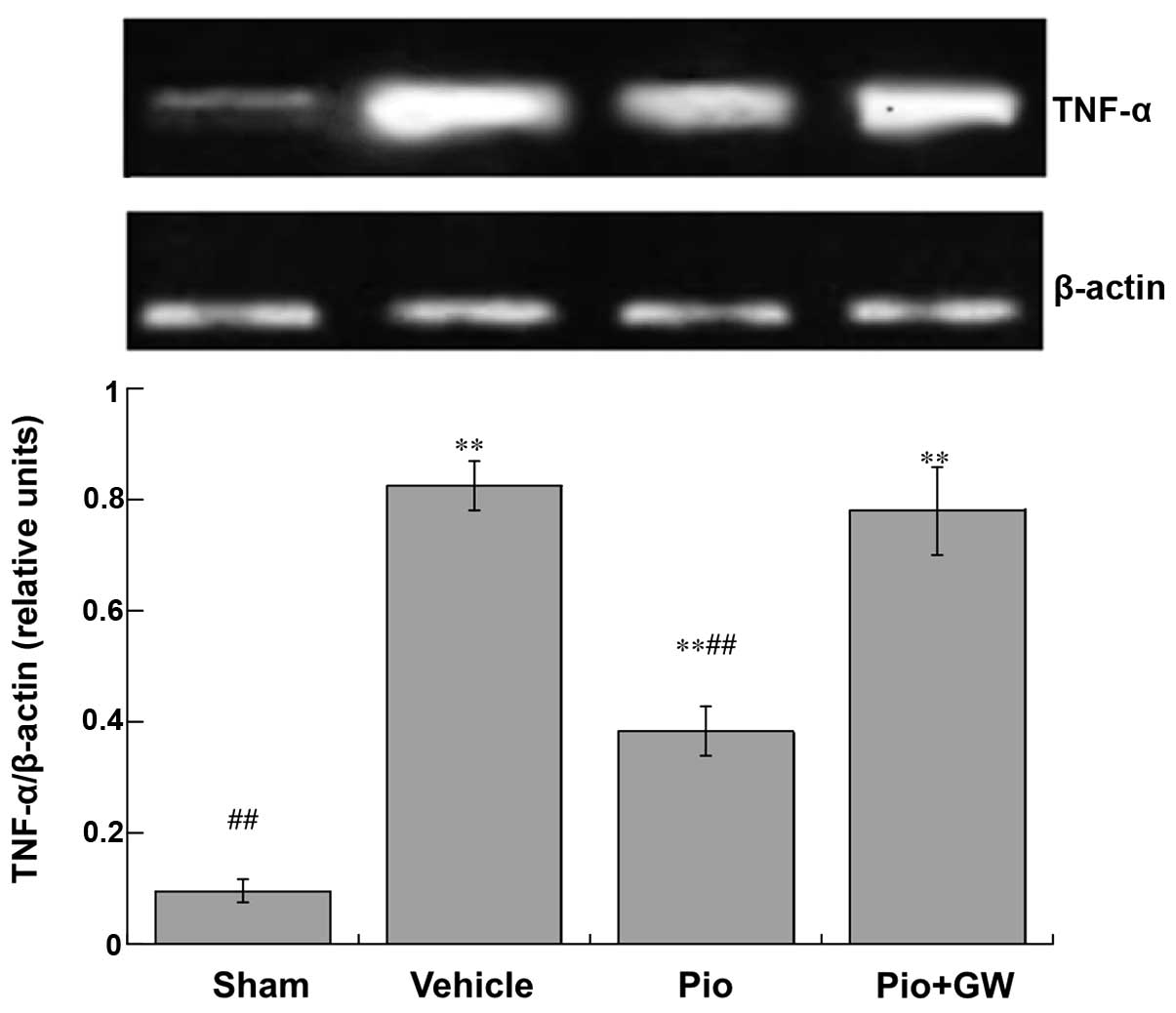
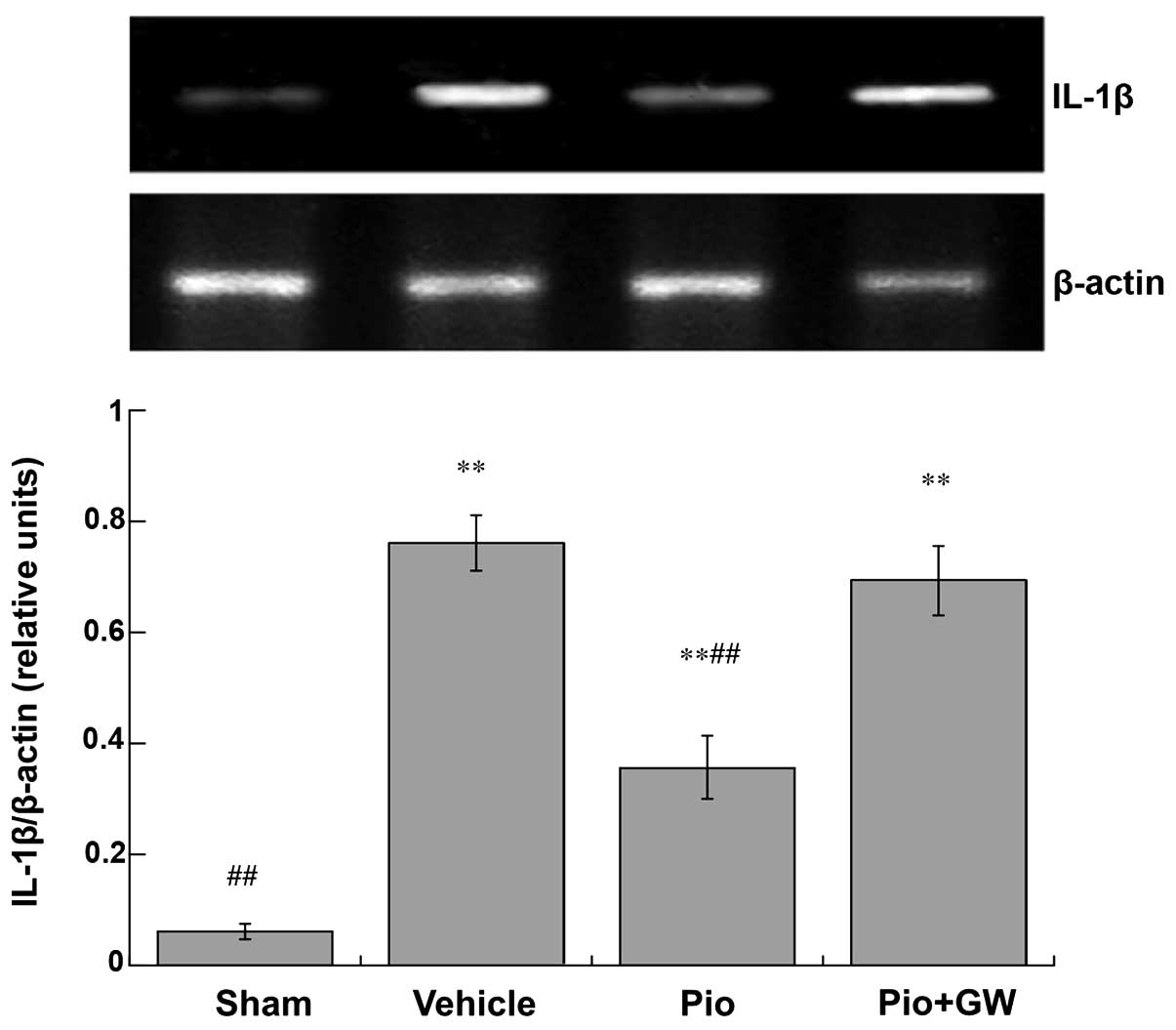
Production of cytokines
Expression levels of TNF-α (Fig. 3) and IL-1β (Fig. 4) in the spinal cord were
significantly increased in CCI rats compared with saline-treated
rats. CCI-induced neuropathic pain was associated with a
significant increase in the protein expression levels of IL-1β and
TNF-α (P<0.01). Pio was able to significantly suppress the
elevation of IL-1β and TNF-α (both P<0.01). However, GW9662 (2
mg/kg, i.p.) was able to reduce the decrease in IL-1β and TNF-α
levels induced by Pio in the spinal cord.
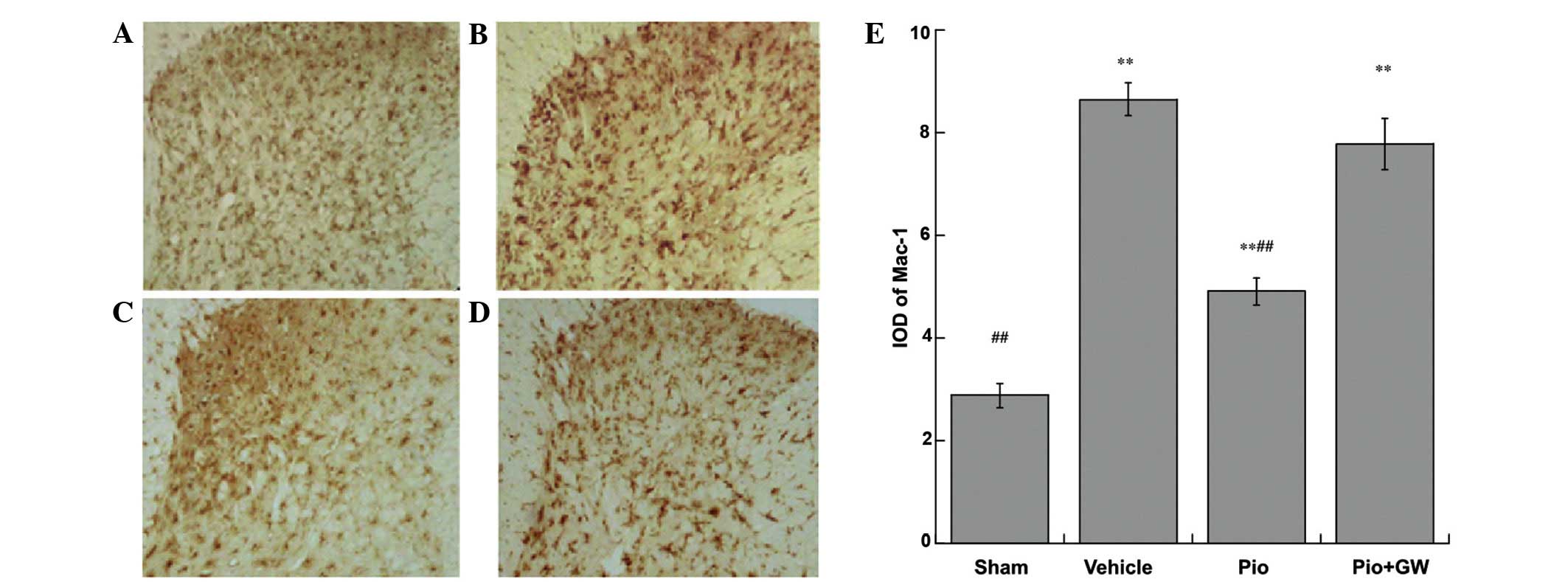
Expression of glial activation
markers
The expression of Mac-1 were investigated in the
sciatic nerves of rats by immunohistochemical analysis (Fig. 5). Pio-treated rats exhibited a
significant attenuation in Mac-1 immunoreactivity. The integral
optical density of Mac-1 immunoreactive cells increased
significantly (P<0.01) in the Pio group compared with the sham
group and vehicle groups, and this effect was reversed by GW9662
coadministration (P<0.01).
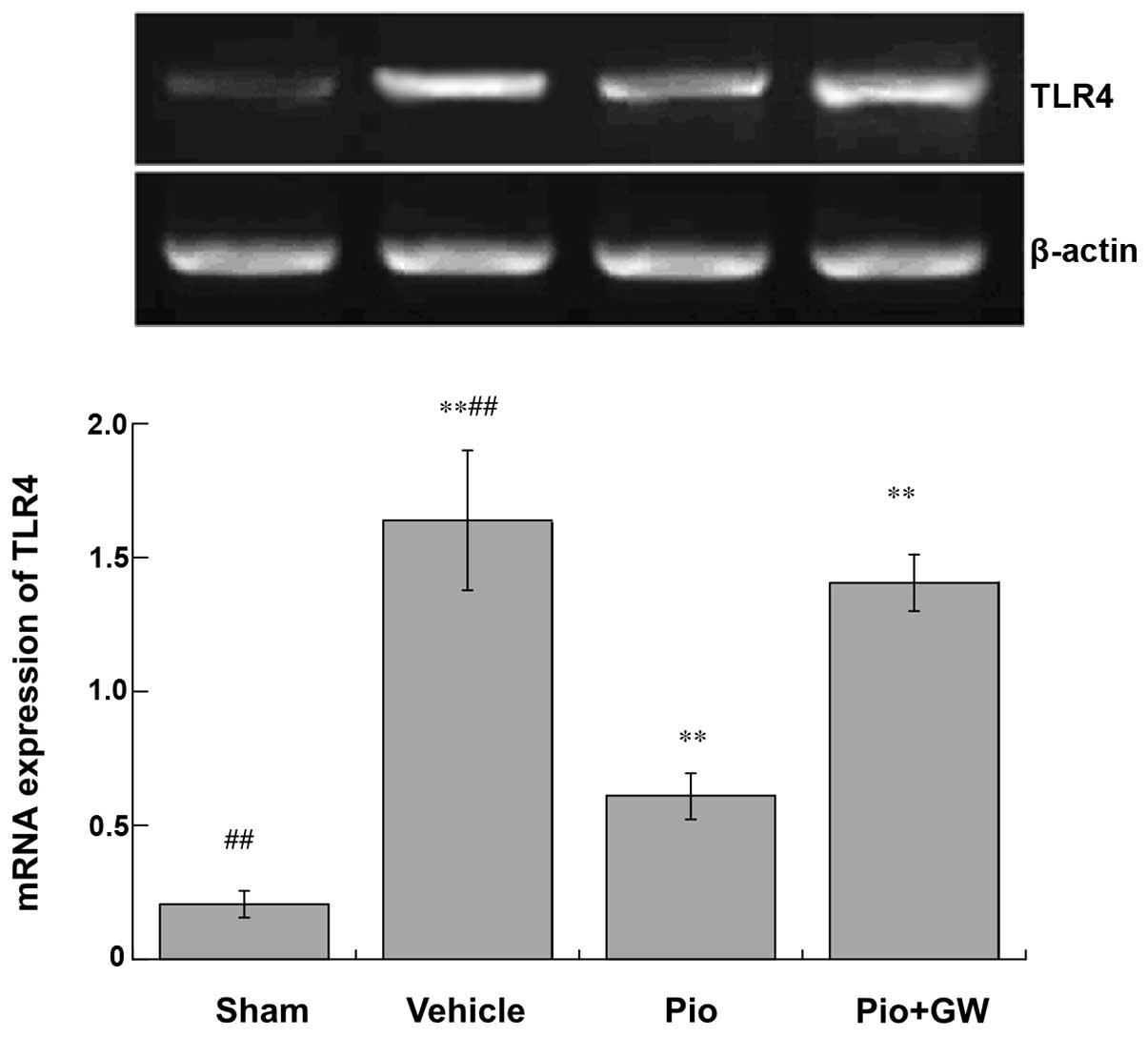
mRNA expression of TLR-4
mRNA expression levels of TLR-4 were significantly
increased in rats on postoperative day 21 in the vehicle group
compared with the sham group (P<0.01). Pio administration
significantly attenuated the increase in TLR-4 expression levels
compared with vehicle administration (P<0.01), which was
significantly reversed by GW9662 coadministration (P<0.01;
Fig. 6).
Discussion
The present study investigated the potential
therapeutic value of Pio, which is a PPAR-γ agonist, in the
treatment of neuropathic pain using a rat model of CCI. Pio was
effective in reducing the incidence of the development of
mechanical hyperalgesia and the upregulation of Mac-1 and
cytokines, and suppressing the activity of TLR-4 in rats after CCI.
GW9662 reversed the aforementioned changes produced by Pio in rats
with CCI.
Neuropathic pain is characterized by hyperalgesia,
allodynia and spontaneous pain. At present, available treatments
for neuropathic pain are neither adequate nor effective, partially
due to the fact that the underlying mechanisms are insufficiently
understood. Current therapies for the treatment of peripheral
neuropathic pain involve numerous drug classes, including opioids,
gabapentinoids, antidepressants, antiepileptic drugs, local
anesthetics and capsaicin. However, the majority of these
treatments have limited efficacy or produce undesirable adverse
effects (1). Thus, increasing
research effort has been applied to the development of novel
therapeutic targets and drugs for the treatment of neuropathic
pain.
Rosiglitazone and Pio are synthetic PPAR-γ agonists
of the thiazolidinedione (TZD) class. As insulin sensitizers, TZDs
remain an important pharmacotherapeutic class in the treatment of
type II diabetes. Several studies have suggested that systemically
administered TZD class drugs facilitate recovery from behavioral
changes during neuropathic pain associated with tibial and sural
nerve transection, in addition to carrageenan-induced inflammation,
spared nerve injury and L5 nerve transection-induced neuropathy
(21–25). The findings of the current study were
consistent with the aforementioned observations that Pio attenuated
the development of mechanical hyperalgesia and cold allodynia. Of
greater note was the observation that the effects of Pio were
successfully blocked by the coadministration of GW9662. Thus, the
present study concluded that PPAR-γ substantially contributes to
the antinoceptive role of Pio, with respect to the amelioration of
neuropathic pain. However, it should be noted that the
administration of Pio for 3 weeks did not produce a significant
effect on the glucose levels in rats in the current study, and that
the glucose levels were within the normal range.
An increasing number of studies have implicated
neuroimmune activation, which has been associated with glial
activation and inflammatory cytokine expression following nerve
injury, to be associated with the generation and maintenance of
behavioral changes during neuropathic pain (26–28). In
the spinal chord, activated microglia lead to hyperalgesia by
releasing algesic soluble factors, including TNF-α and IL-1β
(29). Proinflammatory cytokines
have been hypothesized to contribute to the sensitization of the
CNS, which results in lower thresholds and spontaneous ectopic
neuronal firing (30). As a
consequence, the deleterious escalation of a pathological glial
activation ensues, and induces the expression levels of numerous
factors, including cyclooxygenase-2, inducible nitric oxide (NO)
synthase and substance P (8).
Elevated levels of the aforementioned substances leads to increased
nociceptive activity and thus severe and persistent neuropathic
pain (31). Positive feedback loops
between inflammatory mediators and glial activation, and between
glia and neurons, lead to the enhancement and maintenance of
neuropathic pain. Previous studies have indicated that, in glial
cells, ligand-dependent activation of PPAR-γ has an
anti-inflammatory effect by suppressing the expression of various
inflammatory response genes, including those encoding inducible NO
synthase, TNF-α and IL-1β (32–34). As
demonstrated in the present study, microglial activation, indicated
by increased mRNA expression levels of the surface marker, Mac-1,
led to elevated expression levels of TNF-α and IL-1β inflammatory
mediators, which may, in part, account for the mechanical allodynia
observed in nerve injury models of CCI (35–37).
PPAR-γ receptors are expressed on glial cells in the CNS (38) and spinal cord (39), and the present results indicated that
Pio significantly decreased Mac-1 immunoreactivity and the
production of TNF-α and IL-1β in the lumbar dorsal horn. Thus, we
hypothesize that Pio inhibits the maintenance of neuropathic pain
by decreasing microglia activity, which is the primary source of
inflammatory cytokines in the dorsal horn.
It has previously been suggested that the TLR-4
signaling pathway has an important role in neuropathic pain
(40,41). Proinflammatory cytokines (IL-1β,
TNF-α, IL-6 and INF-γ) are released by the microglial TLR-4
receptor, which is one of several neuron-to-glia signalling
receptors. TLR-4 is a transmembrane receptor protein with
extracellular leucine-rich repeat domains and a cytoplasmic
signaling domain. In the CNS, TLR-4 is expressed on microglia, and
possibly astrocytes and endothelial cells; however, it is not
expressed on neurons (42). Previous
studies have suggested several potential mechanisms that may
mediate the neuroprotective actions of Pio, including diminished
inflammation, and activation of TLR, nuclear factor-κB and
antiapoptotic genes (11,21,43).
However, to the best of our knowledge, the current study is the
first to demonstrate that Pio is able to inhibit TLR-4 expression
during neuropathic pain. This was in agreement with the findings of
a previous study, which reported that Pio inhibits the activation
of TLR-4 in lipopolysaccharide-stimulated microglia cultures.
Further studies are required to elucidate the role of various
signaling pathways besides that of TLR-4 signaling in peripheral
nerve injury.
In conclusion, the present study demonstrated that
Pio administration had an antiallodynic effect in a rat model of
neuropathic pain by inhibiting glial activation and the expression
of TLR-4, which consequently inhibited the production of
proinflammatory cytokines. The results concerning the protective
effects of Pio on hyperalgesia, glial activation, and inflammatory
response in the present study may provide a novel therapeutic
strategy for the treatment of neuropathic pain.
References
|
1
|
Scholz J and Woolf CJ: The neuropathic
pain triad: Neurons, immune cells and glia. Nat Neurosci.
10:1361–1368. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Romero-Sandoval E Alfonso and Sweitzer S:
Nonneuronal central mechanisms of pain: Glia and immune response.
Prog Mol Biol Transl Sci. 131:325–358. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aloisi F: Immune function of microglia.
Glia. 36:165–179. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
DeLeo JA and Yezierski RP: The role of
neuroinflammation and neuroimmune activation in persistent pain.
Pain. 90:1–6. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dong Y and Benveniste EN: Immune function
of astrocytes. Glia. 36:180–190. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mantyh PW, Clohisy DR, Koltzenburg M and
Hunt SP: Molecular mechanisms of cancer pain. Nat Rev Cancer.
2:201–209. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ruohonen S, Jagodi M, Khademi M, Taskinen
HS, Ojala P, Olsson T and Röyttä M: Contralateral non-operated
nerve to transected rat sciatic nerve shows increased expression of
IL-1beta, TGF-beta1, TNF-alpha and IL-10. J Neuroimmunol.
132:11–17. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ji RR, Berta T and Nedergaard M: Glia and
pain: Is chronic pain a gliopathy? Pain. 154:S10–28. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wolska A, Lech-Maranda E and Robak T:
Toll-like receptors and their role in hematologic malignancies.
Curr Mol Med. 9:324–335. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Buchanan MM, Hutchinson M, Watkins LR and
Yin H: Toll-like receptor 4 in CNS pathologies. J Neurochem.
114:13–27. 2010.PubMed/NCBI
|
|
11
|
Park SW, Yi JH, Miranpuri G, Satriotomo I,
Bowen K, Resnick DK and Vemuganti R: Thiazolidinedione class of
peroxisome proliferator-activated receptor gamma agonists prevents
neuronal damage, motor dysfunction, myelin loss, neuropathic pain
and inflammation after spinal cord injury in adult rats. J
Pharmacol Exp Ther. 320:1002–1012. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao Y, Patzer A, Herdegen T, Gohlke P and
Culman J: Activation of cerebral peroxisome proliferator-activated
receptors gamma promotes neuroprotection by attenuation of neuronal
cyclooxygenase-2 overexpression after focal cerebral ischemia in
rats. FASEB J. 20:1162–1175. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao X and Li LY: PPAR-alpha agonist
fenofibrate induces renal CYP enzymes and reduces blood pressure
and glomerular hypertrophy in Zucker diabetic fatty rats. Am J
Nephrol. 28:598–606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Diab A, Hussain RZ, Lovett-Racke AE,
Chavis JA, Drew PD and Racke MK: Ligands for the peroxisome
proliferator-activated receptor-gamma and the retinoid X receptor
exert additive anti-inflammatory effects on experimental autoimmune
encephalomyelitis. J Neuroimmunol. 148:116–126. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Breidert T, Callebert J, Heneka MT,
Landreth G, Launay JM and Hirsch EC: Protective action of the
peroxisome proliferator-activated receptor-gamma agonist
pioglitazone in a mouse model of Parkinson's disease. J Neurochem.
82:615–624. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pedersen WA, McMillan PJ, Kulstad JJ,
Leverenz JB, Craft S and Haynatzki GR: Rosiglitazone attenuates
learning and memory deficits in Tg2576 Alzheimer mice. Exp Neurol.
199:265–273. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Heneka MT, Sastre M, Dumitrescu-Ozimek L,
Hanke A, Dewachter I, Kuiperi C, O'Banion K, Klockgether T, Van
Leuven F and Landreth GE: Acute treatment with the PPAR-gamma
agonist pioglitazone and ibuprofen reduces glial inflammation and
Abeta1-42 levels in APPV717I transgenic mice. Brain. 128:1442–1453.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bennett GJ and Xie YK: A peripheral
mononeuropathy in rat that produces disorders of pain sensation
like those seen in man. Pain. 33:87–107. 1998. View Article : Google Scholar
|
|
19
|
Chaplan SR, Bach FW, Pogrel JW, Chung JM
and Yaksh TL: Quantitative assessment of tactile allodynia in the
rat paw. J Neurosci Methods. 53:55–63. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim YI, Na HS, Yoon YW, Han HC, Ko KH and
Hong SK: NMDA receptors are important for both mechanical and
thermal allodynia from peripheral nerve injury in rats.
Neuroreport. 8:2149–2153. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jain V, Jaggi AS and Singh N: Ameliorative
potential of rosiglitazone in tibial and sural nerve
transection-induced painful neuropathy in rats. Pharmacol Res.
59:385–392. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jia H, Zhu S, Ji Q, Hui K, Duan M, Xu J
and Li W: Repeated administration of pioglitazone attenuates
development of hyperalgesia in a rat model of neuropathic pain. Exp
Clin Psychopharmacol. 18:359–365. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Maeda T, Kiguchi N, Kobayashi Y, Ozaki M
and Kishioka S: Pioglitazone attenuates tactile allodynia and
thermal hyperalgesia in mice subjected to peripheral nerve injury.
J Pharmacol Sci. 108:341–347. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takahashi Y, Hasegawa-Moriyama M, Sakurai
T and Inada E: The macrophage-mediated effects of the peroxisome
proliferator-activated receptor-gamma agonist rosiglitazone
attenuate tactile allodynia in the early phase of neuropathic pain
development. Anesth Analg. 113:398–404. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Churi SB, Abdel-Aleem OS, Tumber KK,
Scuderi-Porter H and Taylor BK: Intrathecal rosiglitazone acts at
peroxisome proliferator-activated receptor-gamma to rapidly inhibit
neuropathic pain in rats. J Pain. 9:639–649. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang ZJ, Cao DL, Zhang X, Ji RR and Gao
YJ: Chemokine contribution to neuropathic pain: Respective
induction of CXCL1 and CXCR2 in spinal cord astrocytes and neurons.
Pain. 154:2185–2197. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Akimoto N, Ifuku M, Mori Y and Noda M:
Effects of chemokine (C-C motif) ligand 1 on microglial function.
Biochem Biophys Res Commun. 436:455–461. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Clark AK, Old EA and Malcangio M:
Neuropathic pain and cytokines: Current perspectives. J Pain Res.
6:803–814. 2013.PubMed/NCBI
|
|
29
|
Ji RR and Suter MR: P38 MAPK, microglial
signaling, and neuropathic pain. Mol Pain. 3:332007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Moalem G and Tracey DJ: Immune and
inflammatory mechanisms in neuropathic pain. Brain Res Rev.
51:240–264. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Inoue M, Yamada T and Ueda H: Low dose of
kyotorphin (tyrosine-arginine) induces nociceptive responses
through a substance P release from nociceptor endings. Brain Res
Mol Brain Res. 69:302–305. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zingarelli B, Sheehan M, Hake PW, O'Connor
M, Denenberg A and Cook JA: Peroxisome proliferator activator
receptor-gamma ligands, 15-deoxy-Delta(12,14)-prostaglandin J2 and
ciglitazone, reduce systemic inflammation in polymicrobial sepsis
by modulation of signal transduction pathways. J Immunol.
171:6827–6837. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fuenzalida K, Quintanilla R, Ramos P,
Piderit D, Fuentealba RA, Martinez G, Inestrosa NC and Bronfman M:
Peroxisome proliferator-activated receptor gamma up-regulates the
Bcl-2 anti-apoptotic protein in neurons and induces mitochondrial
stabilization and protection against oxidative stress and
apoptosis. J Biol Chem. 282:37006–37015. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bernardo A, Ajmone-Cat MA, Gasparini L,
Ongini E and Minghetti L: Nuclear receptor peroxisome
proliferator-activated receptor-gamma is activated in rat
microglial cells by the anti-inflammatory drug HCT1026, a
derivative of flurbiprofen. J Neurochem. 92:895–903. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Watkins LR, Milligan ED and Maier SF:
Glial activation: A driving force for pathological pain. Trends
Neurosci. 24:450–455. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Luna-Medina R, Cortes-Canteli M, Alonso M,
Santos A, Martínez A and Perez-Castillo A: Regulation of
inflammatory response in neural cells in vitro by
thiadiazolidinones derivatives through peroxisome
proliferator-activated receptor gamma activation. J Biol Chem.
280:21453–21462. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen Y, Willcockson HH and Valtschanoff
JG: Influence of the vanilloid receptor TRPV1 on the activation of
spinal cord glia in mouse models of pain. Exp Neurol. 220:383–390.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Moreno S, Farioli-Vecchioli S and Cerù MP:
Immunolocalization of peroxisome proliferator-activated receptors
and retinoid X receptors in the adult rat CNS. Neuroscience.
123:131–145. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Diab A, Deng C, Smith JD, Hussain RZ,
Phanavanh B, Lovett-Racke AE, Drew PD and Racke MK: Peroxisome
proliferator-activated receptor-gamma agonist 15-deoxy-Delta
(12,14)-prostaglandin J2 ameliorates experimental autoimmune
encephalomyelitis. J Immunol. 168:2508–2515. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hutchinson MR, Zhang Y, Brown K, Coats BD,
Shridhar M, Sholar PW, Patel SJ, Crysdale NY, Harrison JA, Maier
SF, et al: Non-stereoselective reversal of neuropathic pain by
naloxone and naltrexone: Involvement of toll-like receptor 4
(TLR4). Eur J Neurosci. 28:20–29. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cao L, Tanga FY and DeLeo JA: The
contributing role of CD14 in toll-like receptor 4 dependent
neuropathic pain. Neuroscience. 158:896–903. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Miyake K: Innate immune sensing of
pathogens and danger signals by cell surface Toll-like receptors.
Semin Immunol. 19:3–10. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gurley C, Nichols J, Liu S, Phulwani NK,
Esen N and Kielian T: Microglia and Astrocyte Activation by
Toll-Like Receptor Ligands: Modulation by PPAR-gamma Agonists. PPAR
Res. 2008:4531202008. View Article : Google Scholar : PubMed/NCBI
|