Introduction
In recent years, a large number of studies have
focused on disorders of lipid and carbohydrate metabolism as the
etiologic factors of type 2 diabetes (T2D) and atherosclerosis, and
to be associated with their progression. Millions of people are
suffering from metabolic syndrome and cardiovascular failure
worldwide, which are considered the most common causes of human
mortality (1,2). The concept of ‘metabolic syndrome’ (MS)
(2), which indicates a close
association between certain types of hyperlipidemia and impaired
glucose tolerance, has been widely accepted. One of the most
important tasks for modern medicine is to identify novel, effective
and easily obtainable remedies with low toxicity for preventive and
adjunctive therapy of T2D and hyperlipidemia among natural sources
of biologically active substances (BAS), which possess various
mechanisms of protective action, aimed at restoring the disturbed
biochemical status of the body (3–7).
Marine organisms are a rich source of natural
products. To date, >20 million low-molecular-weight natural
marine BASs, which often differ from secondary metabolites of
terrestrial organisms in terms of chemical structure as well as in
their biological effects, have been isolated and identified. The
variety of chemical structures, the high biological activity and
the prospects for their effective therapeutic application are the
factors that attract increasing attention to substances of marine
origin (4). It should be noted that
certain major active components, i.e. rosmarinic acid and luteolin,
have been originally found in terrestrial plants, such as rosemary
and artichokes, while the disulfated derivative of luteolin may be
of specific marine origin, but is a metabolic product of luteolin
in the body (5–7).
The present review performed a brief analysis of
published experimental data by our and other groups and postulated
the possible mechanisms of the antidiabetic effects of the
polyphenol complex (PPC) isolated from seagrasses of the genus
Zostera (Z.; Z. marina and Z.
asiatica), and its main components, namely rosmarinic acid
(RA), luteolin (LT) and its disulfated derivative, 7,3′-disulfate
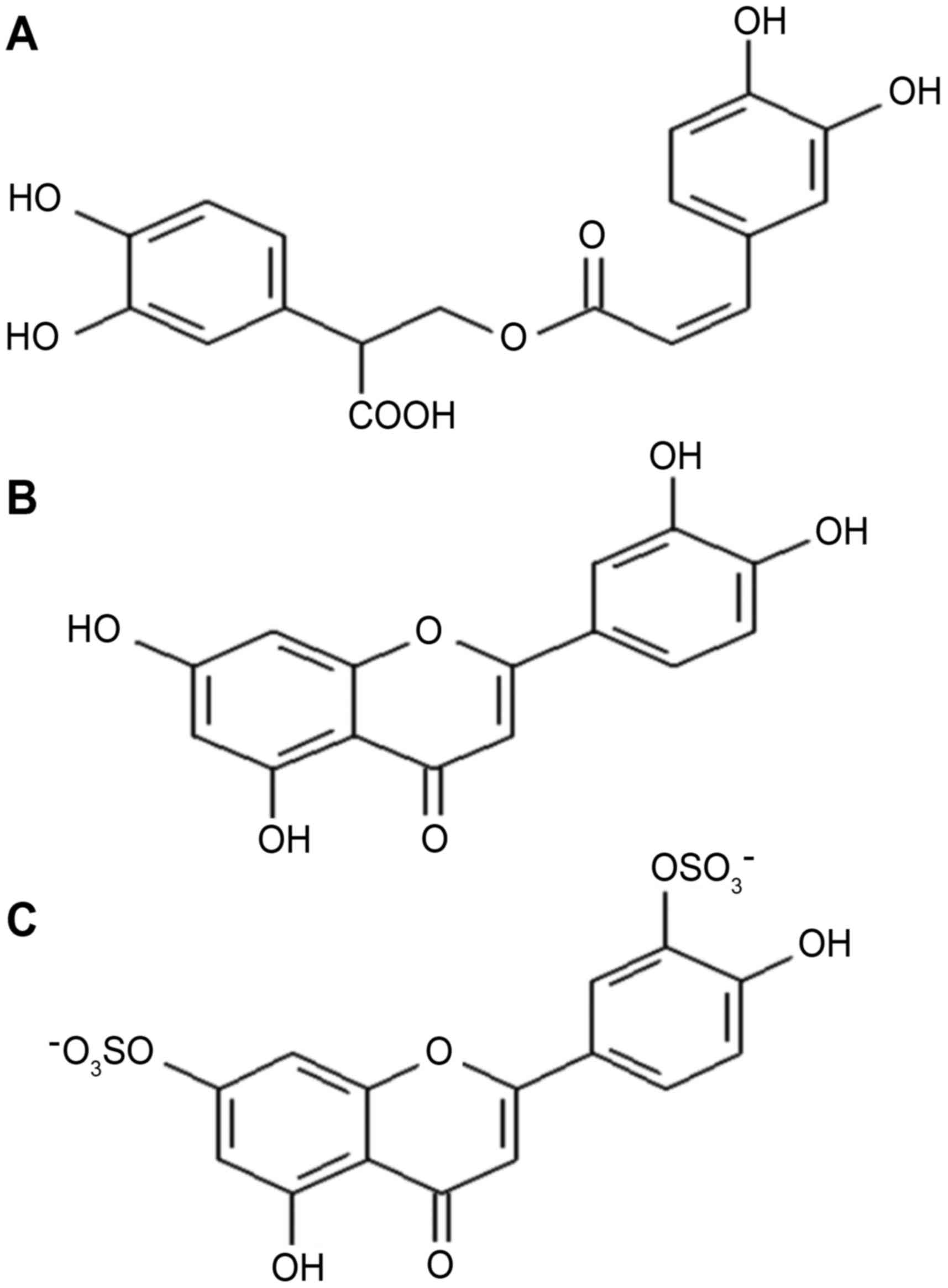
luteolin (DSL), whose chemical structures are presented in Fig. 1.
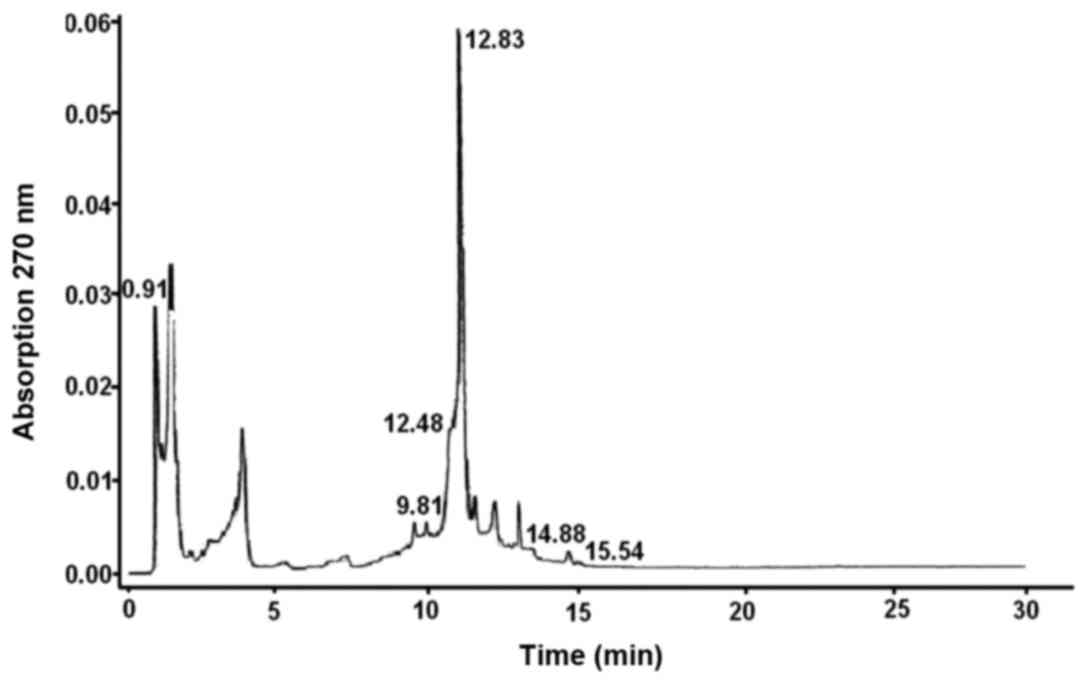
The chemical composition of PPC from seagrasses of
the genus Z. was analyzed by using high-performance liquid
chromatography/mass spectrometry (Fig.
2).
As shown in Fig. 2, a
major portion of the PPC is comprised of RA (45%) and mono- and
disulfated derivatives of LT (~45%); bioflavonoids including LT and
apigenin constitute the remaining portion. The Laboratory of
Biotechnology at the G.B. Elyakov Pacific Institute of Bioorganic
Chemistry, Far Eastern Branch of the Russian Academy of Sciences
(Vladivostok, Russia) has developed a method of isolating DSL
(Fig. 2) from seagrasses of the
genus Z. and performed extensive pharmacological trials in
experimental animal models to compare its activities to those of
the flavone LT and other commercial pharmaceuticals (8–10).
As recent studies on the antidiabetic properties of
the main constituents of the PPC, such as RA and LT, have made
significant progress, the present review focused on analyzing their
results to postulate possible mechanisms of action. In addition,
the present study attempted to elucidate why DSL has higher
activity than LT with regard to a number of pharmacological
properties. This information will be useful for the development of
more effective DSL-based remedies for adjunctive therapy of MS and
T2D.
Proposed mechanisms of the pharmacologic
activity of RA and LT as well as its analogues
Effect exerted on peroxisome
proliferator-activated receptor (PPAR) signaling pathways
According to studies by our group using experimental
animal models of hyperglycemia (model of alloxan-induced diabetes)
and hyperlipidemia (tyloxapol model), the PPC exerted potent
antidiabetic and antihyperlipidemic effects leading to substantial
normalization of clinical blood plasma parameters compared to those
in untreated animals. In most cases, the therapeutic effect of the
entire complex and its individual components proved to be stronger
than that of known commercial pharmaceutical drugs applied for
treating MS-associated pathologies (glibenclamide, metformin and
nicotinic acid), which were used as positive controls (5–7). The
high therapeutic activity of the PPC is undoubtedly determined by
the pharmacological properties of its main components RA, LT and
DSL, the biological activity and the possible mechanisms of action
of which are discussed below.
It has been established that the phenylpropanoid RA,
the flavone LT and its analogues exert pronounced antioxidant and
anti-inflammatory effects, and have modulating effects on PPARs,
which are among the main regulators of lipid and carbohydrate
metabolism (5,6,11,12).
PPAR has several known isoforms: PPARα, PPARγ
(subtypes 1 and 2) as well as PPARβ and PPARδ, which are all cell
receptors with a key role in preventing the pathologies associated
with the occurrence and progression of T2D and MS. PPARs are
nuclear receptors that regulate transcriptional activity; they can
be modulated by binding of a large variety of small ligands,
including certain phenolic compounds. PPARs have crucial roles as
molecular sensors of environmental factors, including nutritional
constituents to ‘fine-tune’ lipid and glucose homeostasis, and are
also important for other physiological processes. Subsequent to
activation, PPARs may form heterodimers with retinoid X receptors,
which bind to the PPAR response elements in the promoter regions of
expressed genes and either increase or reduce their transcription,
the main effect of which being an increase in the storage of fatty
acids in adipocytes, while the amount of fatty acids present in the
circulation is decreased (4–7,13,14).
PPARγ is predominantly expressed in adipose tissue.
The endogenous ligands for these receptors are free fatty acids
(FFAs) and a few of their exogenous ligands are used as therapeutic
agents for the treatment of T2D. Antagonists of PPARγ prevent
obesity and exert antidiabetic activity. Furthermore, they reduce
adhesion of monocytes to endothelial cells and inhibit the
inflammatory action of macrophages. Activation of PPARγ alters the
expression of genes involved in metabolic processes such as
adipogenesis, insulin signal transmission and glucose transport,
resulting in the reduction of the respective tissue's resistance to
insulin. RA and LT are PPARγ ligands with antioxidant and
anti-inflammatory properties (4–7,12,13,15).
Various studies have suggested that RA can function
as a PPARγ antagonist by inhibiting adipogenesis and
PPARγ-dependent gene expression (12), while LT can either function as a
PPARγ agonist by enhancing the expression of PPARγ-dependent genes
in adipocytes (16) or a PPARγ
antagonist (12,17). LT binds PPARγ, but unlike
thiazolidinediones (TZDs), which are a class of modern antidiabetic
and antihyperlipidemic medicinal drugs, it does not promote
adipocyte differentiation. LT exhibits mild partial
agonist/antagonist activity, inhibits several PPARγ target genes
and PPARγ-dependent adipogenesis, but activates the glucose
transporter type 4 (GLUT4) to a similar degree to that of RA,
implying gene-specific partial agonism. LT is thought to be useful
as a safe selective PPARγ modulator (5,6,18).
Orally ingested, LT glycosides are hydrolyzed by
β-glucosidase to LT and then absorbed into the blood as conjugated
metabolites, such as LT glucuronides and sulphates, which have
sulfate groups in an arrangement similar to the mono- and
disulfates of luteolin seagrass Zostera and are similar
species to DSL. Indeed, the free aglycone concentration in
circulating blood is undetectably low in humans and animals
(19,20). Therefore, luteolin metabolites are
presumably responsible for luteolin anti-inflammatory effects.
Luteolin glucuronides, particularly luteolin-7-O-glucuronide, have
been demonstrated to reduce the expression of inflammatory genes
the lipopolysaccharide-treated cells (20).
Should be emphasized that flavanone-class PPARγ
partial agonists, luteolin and hesperetin glucuronides, showed
similar activation profiles of the PPARγ ligand-binding domain
mutants, even though they have different side chain functionalities
(21). These glucuronides showed an
additive PPARγ-transactivating effect with the antidiabetic drug
troglitazone, a thiazolidine-2,4-dione which acts as a full PPARγ
agonist. The additive effect may indicate that flavanone
glucuronides activate PPARγ in a different manner to that of
thiazolidine-2,4-dione derivatives. These glucuronides and
troglitazone have distinct mechanisms of PPARγ activation, probably
due binding to different sites of PPARγ. This additive effect may
indicate that flavone glucuronides activate PPARγ in a different
manner from thiazolidine-2,4-dione derivatives, possibly by binding
to the PPARγ ligand-binding domain at a different site (21).
Thus, PPARs are among the main molecular targets of
RA, LT and several of its structural analogues and conjugates. TZDs
(pioglitazone, rosiglitazone) and fibrates have a selective effect
on PPARγ, which is a target for the treatment of T2D. PPARγ full
agonist TZDs are effective insulin sensitizers and
anti-inflammatory agents; however, their use is limited by adverse
effects. The major side effects of all TZDs are water retention
and, as a consequence, edema. Caution should therefore be taken
when TZDs are prescribed and patients should be made aware of the
potential water retention/weight gain (22,23).
Thus, the search for other effective and safe therapeutic agents
remains an urgent issue.
Antiinflammatory and antioxidant activity of
RA, LT and its analogues
Inflammation is considered a potential mechanism in
the progression of numerous pathologies, including insulin
resistance, cardiovascular diseases and T2D. RA and LT, as main PPC
components, modulate inflammatory responses in animal organisms via
various mechanisms, a few of which depend on PPARs, whereas others
are PPARs-independent but are regulated by numerous transcriptional
factors; therefore, the mechanisms of action of these phenolic
compounds have a pleiotropic pattern (19,24,25).
In the mechanism of damage and defense associated
with oxidative stress, the process of lipid peroxidation (LP) has
an important role. In the case of LP, the formation of pathological
changes in MS and T2D are predominantly the result of functional
impairment of the antioxidant system that seeks to protect the
organism. LP syndrome develops upon excessive accumulation of the
products of LP in the organism, which includes pathological
constituents such as damage to membrane lipids, lipoproteins and
proteins, inactivation of enzymes, and impairment of cell division
and phagocytosis. These pathological constituents subsequently lead
to changes in the structural and functional organization of the
membranes and the formation of various pathological processes.
Therefore, due to their anti-inflammatory and antioxidant actions,
the major components of PPC, RA, LT and its sulfated derivatives,
may be utilized for the treatment of MS and T2D (26,27). PPC
and its main components showed a high degree of safety and lack of
side effects (8,26).
LT is known to inhibit damage-inducing reactive
oxygen species (ROS) and is considered an antioxidant via various
mechanisms of action. First, LT functions as a scavenger of ROS
through being oxidized by it (24).
LT has the structural elements required for antioxidant activity of
flavonoids: Hydroxyl moieties at the 3′ and 4′ positions, a double
bond between carbon atoms in positions 2 and 3, and a carbonyl
group in position 4 (25). Hydrogen
atoms from the hydroxyl groups situated on the aromatic rings can
be donated to free radicals. As an aromatic compound, LT can
support unpaired electrons around the M-electron system. Direct
evidence that LT is a ROS scavenger was obtained from acellular
systems (27). Second, LT inhibits
ROS-generating oxidases. For instance, LT suppresses the formation
of superoxide anions (O2•−) by inhibiting the
activity of xanthine oxidase (28).
However, it remains elusive whether LT influences ROS formation in
the mitochondria of mammalian cells, which are the main site for
ROS formation, although it has an effect on the mitochondrial
electron transport chain in parasitic (leishmanial) cells (29). Third, LT may exert its antioxidative
action by protecting or enhancing endogenous antioxidants such as
superoxide dismutase, catalase, glutathione-S-transferase, and
glutathione reductase (30). As has
been recently shown, LT increased the endogenous mRNA and protein
levels of nuclear factor erythroid-2 p45-related factor 2 (Nrf2),
which is a critical intracellular regulator in the adaptive
response via regulation of a wide array of cytoprotective enzymes,
as well as Nrf2 target genes. LT significantly and dose-dependently
decreased the production of cytosolic phospholipase A2 (cPLA2),
nitric oxide (NO) and inducible NO synthase (iNOS), which were
induced by treatment of cells with 1 µg/ml lipopolysaccharide (LPS)
for 24 h. Thus, LT significantly activated the phosphoinositide
3-kinase/Nrf2/antioxidant response elements system, and this
activation may be responsible for its anti-inflammatory effects, as
demonstrated by the suppression of LPS-induced NO, iNOS and cPLA2
(31). Fourth, LT directly inhibits
the enzymes that catalyze oxidation of cellular components. For
instance, LT suppresses the formation of malondialdehyde in liver
lipids, which is stimulated by lipoxygenase (LOX), cyclooxygenase
(COX) and ascorbic acid (32).
Finally, LT chelates transition metal ions responsible for
production of ROS, and therefore inhibits reactions of LOX and
suppresses oxidation depending on non-transition metals (30). It should be noted that the LT may
also exert its antioxidant effects in vitro. For instance,
LT inhibited LPS-induced production of hydroxyl anions
(•OH) in macrophages by trapping
O2•− or by inhibiting the activity of
xanthine oxidase, or by a combination of the two mechanisms
(33).
It is important to emphasize that RA markedly
surpasses other known antioxidants, including LT, in tests for
antioxidant activity in the systems
(hemoglobin-H2O2-luminol) and
[2,2′-azobis(2-methylpropionamidine)dihydrochloride-luminol] and
with regard to the inhibition of peroxidation of linoleic acid.
According to the level of their antioxidant activity, the studied
substances can be arranged in the following order: RA >
dihydroquercetin > LT > trolox > ascorbic acid. RA
molecules mainly localized in lipid bilayer in the region of polar
groups, while not significantly affecting its structural and
functional properties, as the permeability of the planar bilayer
membranes was not changed by RA at 0.5–10 µg/ml. To fully prevent
LP, 1 mol% RA spontaneously incorporated in the lipid bilayer is
sufficient. The antioxidant effect of RA is based on its ability to
inhibit the initial stage of free radical reactions of LP, much
determined by the formation of ROS (34).
The pronounced antioxidant activity of RA has a
positive significance, particularly in the progression of oxidative
stress in various biological systems, which is considered to be the
process of excessive ROS production. The LP process has an
important role in the mechanisms of damage and protection during
oxidative stress (34). Oxidative
stress as well as inflammatory processes are implicated in diabetic
vascular complications. By exerting its antioxidant as well as
anti-inflammatory effects, RA protects aortic endothelial function
and ultrastructure against diabetes-induced damage (35).
LT is also known as an anti-inflammatory and
cytoprotective agent. It is successfully used for treating diseases
such as T2D, MS and obesity, which are associated with the
expansion of chronic inflammatory reactions, which in turn are
characterized by an increase in macrophage infiltration, changes in
cytokine production and activation of the inflammatory signaling
pathway in adipose tissue. In obesity, fatty acid levels,
particularly those of saturated free fatty acids, are elevated and
directly induce inflammatory responses in macrophages via toll-like
receptor 4 (TLR4) and LPS receptor. In numerous cell types,
including macrophages, nuclear factor κB (NF-κB) and c-Jun
N-terminal kinase (JNK) pathways are important modulators of
inflammatory gene expression downstream of TLR4. In this way,
adipocytes and macrophages interact in a paracrine manner and
create a self-amplifying cascade of inflammation that augments the
inflammatory changes and insulin resistance in obese adipose tissue
(34–39). It is therefore plausible that
pharmacological agents capable of stopping the cascade of
inflammatory reactions at the molecular level possess antidiabetic
properties. In macrophages, LT inhibited the release of the tumor
necrosis factor-α (TNF-α) and activation of NF-κB induced by LPS,
and reduced the mRNA levels TNF-α, interleukin-6 (IL-6) and
monocyte chemotactic factor-1. Upon stimulation with insulin, LT
increased glucose uptake by adipocytes and enhanced the expression
of adiponectin and liptin genes; however, the transcriptional
activity of PPARγ had the most pronounced effect on the presence of
LT (40,41).
Thus, LT reduces insulin resistance in obese adipose
tissue; in other words, it acts as an important corrective agent of
T2D-associated disorders at the cellular and molecular level. It
may therefore be assumed that the main components of PPC, RA and
LT, by interfering with the TLR4/NF-κB and TLR4/JNK axes, may be
useful in preventing the onset of insulin resistance in T2D
patients.
It is known that persistent hyperglycemia and
increased FFA levels contribute to oxidative stress, which is
thought to be the immediate cause of the onset and progression of
T2D and its complications (42).
Per os administration of RA (100 mg/kg) in a rat model of
T2D (high-fat diet and streptozotocin) greatly increased the
sensitivity to insulin; at the same time, levels of glucose,
glycated hemoglobin, glycation end products, TNF-α, IL-1β, IL-6,
NO, phosphorylated-JNK, P38 mitogen-activated protein kinases and
NF-κB in blood plasma significantly declined with a simultaneous
rise of the insulin levels in diabetic rats. Furthermore, treatment
with RA significantly decreased the levels of triglycerides, FFA,
cholesterol and LP in blood plasma and pancreas of rats. The
reduced activity of superoxide dismutase, catalase, glutathione
peroxidase and glutathione-S-transferase, and also the lower levels
of ceruloplasmin, vitamins C and E, and reduced glutathione in
blood plasma of diabetic rats were largely normalized as a result
of RA treatment. This indicated a high therapeutic potential of RA,
which was confirmed by the increased levels of Nrf2, which is a
critical intracellular regulator in adaptation processes via the
regulation of a wide array of cytoprotective enzymes (42).
Histological, ultrastructural and
immunohistochemical results have demonstrated that oral
administration of RA protects pancreatic β-cells from oxidative
stress caused by high-fat diet and streptozotocin in an
experimental model of diabetes. It can be assumed that oral
administration of RA relieves the symptoms of diabetes mostly due
to its antioxidant properties (34,43).
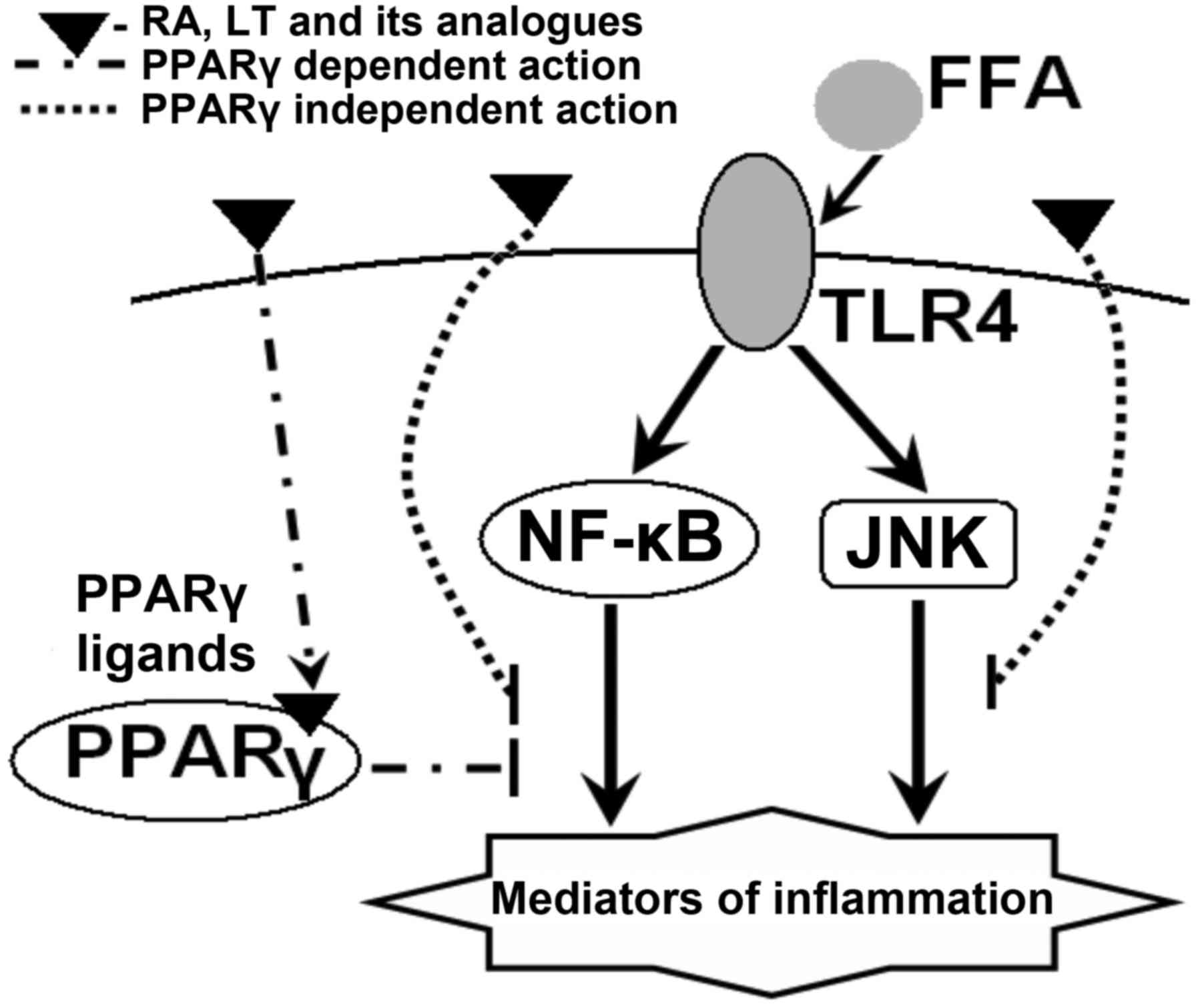
Fig. 3 depicts a
probable pattern of changes in functions and expression of genes
participating in inflammatory signaling pathways in diabetes under
the effect of RA, LT and its analogues.
Metformin, a pharmaceutical drug most widely used in
the treatment of T2D, has a mechanism of action similar to that of
LT and RA. Metformin blocks the synthesis of the transcription
factor NF-κB, which controls the downstream synthesis of certain
molecules involved in inflammatory responses. The fact that
metformin also activates the synthesis of Nrf2, which controls the
expression of numerous enzymes involved in the antioxidant
protection of the body, is equally important. These enzymes
scavenge ROS, thus stopping the activity of macrophages, while
their deregulation or loss of function causes mortality of
pancreatic β-cells, resulting in diabetes (44).
It should be noted that besides the action through
PPAR-dependent and -independent signaling pathways, RA, LT and its
analogues exhibit a pharmacological activity via other
intracellular signaling and biochemical pathways: i) They inhibit
the formation of enzymes (e.g. PLA2, COX and LOX) involved in the
synthesis of eicosanoids, and as a result, the content of
proinflammatory molecules (prostaglandins and leukotrienes)
declines; ii) they also inhibit factors of transcriptional
activation that modulate expression of proinflammatory genes
(COX-2, iNOS, TNF-α as well as IL-1β and −6) (4–9,13), as presented in Fig. 4.
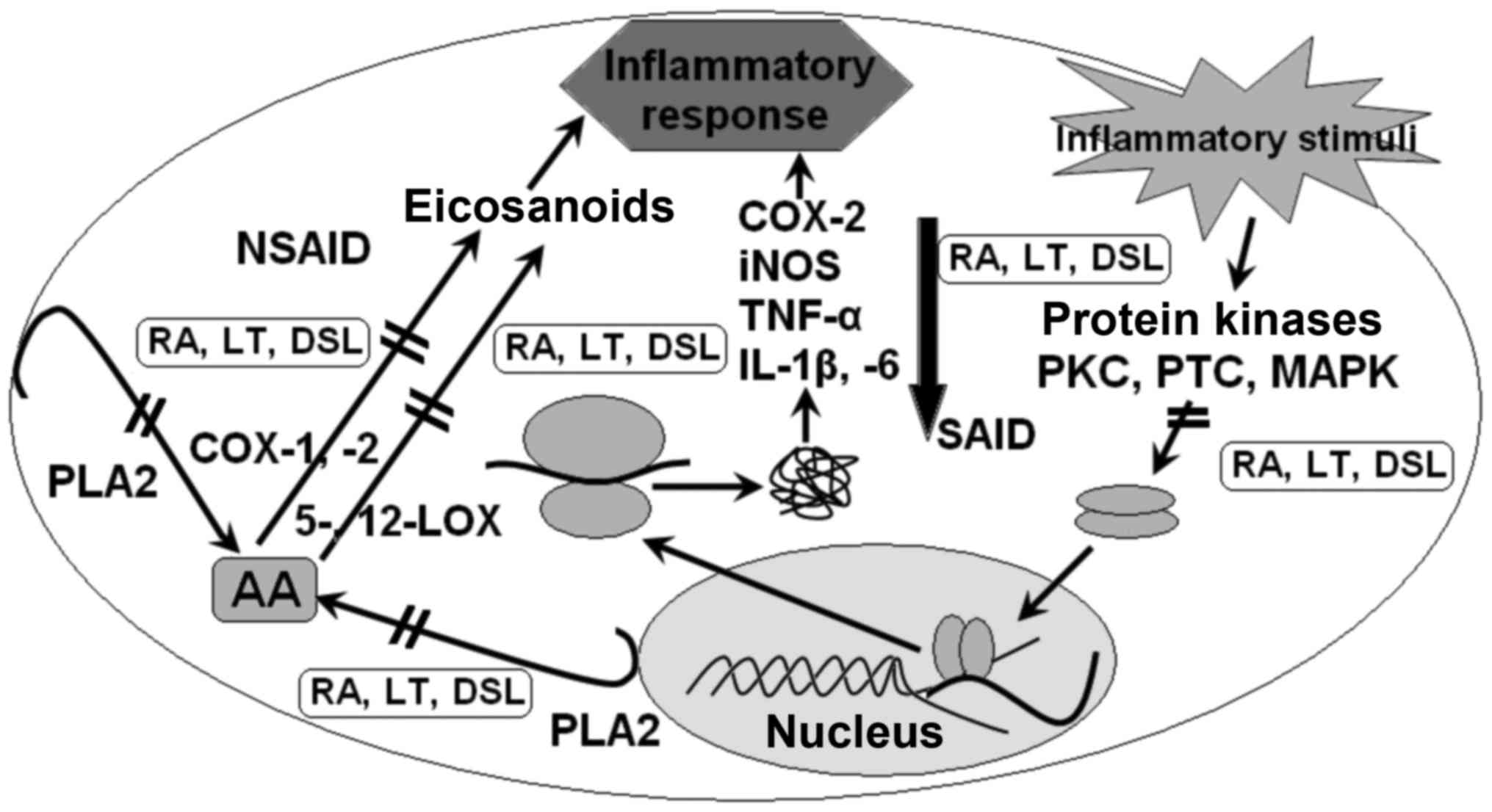 | Figure 4.Proposed mechanisms of the AA of RA,
LT and DSL. RA, rosmarinic acid; LT, luteolin; DSL, 7,3′-disulfate
luteolin; NSAID, non-steroidal anti-inflammatory agent; SAID,
steroidal anti-inflammatory agent; PKC, protein kinase C; PTC,
protein tyrosine kinase; MAPK, mitogen-activated protein kinase;
iNOS, inducible nitric oxide synthase; COX, cyclooxygenase; LOX,
lipoxygenase; TNF-α, tumor necrosis factor; IL, interleukin; PLA2,
phospholipase A2; AA, arachidonic acid. |
The basic mechanisms of these properties are not
fully understood, but can partly be explained by the redox
properties of these compounds. This means that in various cell
systems, they may engage in redox reactions, thus performing
functions as donors and acceptors of electrons and protons. The
redox properties of RA, LT, and most phenolic compounds are not
only confined to their antioxidant and anti-inflammatory activity.
Moreover, it was shown that LT exhibits oxidant properties when
acting on cells at low concentrations (19). Experimental studies by our group on
the combined use of PPC from seagrasses of the genus Z. and
its main components, RA and DSL, with metformin, based on an
alloxan-induced diabetes model, did not reveal any synergistic or
additive effects. It is important to emphasize that the protective
activity of PPC was higher than that of its main components, RA and
DSL (4–7).
A different pattern was observed when physical
exercise was combined with the therapeutic use of antioxidants
(vitamins C and E) and metformin. A combined therapy of this type
appeared to have opposing physiological consequences (45,46).
With this regard, it is important to be aware of the molecular
mechanisms of action of various substances with anti-diabetic
properties when used in combination for the purpose of enhancing
their protective effects against the progression of pathological
processes; it may also be important to be aware of their action
towards normal cells.
Obviously, the mechanisms of modulation of cellular
signaling pathways differ between normal and pathological cells.
For instance, LT inhibits JNK in macrophages but activates this
kinase in tumor cells (47).
Furthermore, LT inhibits the activity of NF-κB in epithelial cells
and macrophages during inflammation by suppressing the activation
of inhibitor of NF-κB, while in tumor cells, participation of NF-κB
in nuclear events is apparently not suppressed in the presence of
LT (48).
It may be assumed that various mechanisms of NF-κB
suppression exist in normal and pathological cells, which obviously
depends on the redox status of these cells and/or on the
redox-regulating properties of the active phenolic agent,
particularly LT.
Features of the antidiabetic action of
DSL
Flavonoids are thought to exhibit a high bioactivity
in pure form, but their metabolites are far less active and are
mainly the ‘deposited’ form of these medicinal agents. However,
recent findings contradict this dogma. In numerous cases,
flavonoids do not lose their activity upon metabolization by
processes including sulfation or glycosylation, particularly with
regard to their anti-inflammatory and antioxidant potential.
Moreover, in certain cases, the effectiveness of the protective
action of conjugated forms of phenolic compounds on certain
molecular targets in cells and tissues was markedly increased as
compared to that of the native phenols, while their cytotoxicity
was declined (8,9,49).
Sulfation is an important metabolic pathway of
flavonoids in plants. Previous studies by our group found a
significant amount of DSL in seagrasses of the Z. family
(Z. marina and Z. asiatica) and developed a simple
method to isolate it. These studies showed that in numerous cases,
the pharmacological activity of DSL was higher than that of LT
(7–10). It may be assumed that DSL is a
natural water-soluble form of LT, which is absorbed and reaches the
blood plasma of animals and human through the intestine, skipping
the stages of modification in intestinal and liver cells. DSL is
likely to have a lower toxic potential than LT and thus, the
efficiency of its pharmacological action may be increased.
When flavonoids enter the human body, sulfation and
glycosylation represent the main ways of their metabolic
transformation. It is logical to assume that therapeutic and
preventive medications based on flavonoid derivatives, to be
developed in the future, are equally promising to non-modified
flavonoids that are already widely used in medical practice
(8,9).
A study by our group assessing the actions of the
sulfated conjugate LT-DSL in models of T2D and hyperlipidemia
revealed a higher therapeutic activity than LT (5–9). It may
be assumed that, similar to resveratrol (50), sulfated LT is absorbed by epithelial
cells more effectively than pure LT. In addition, it is possible
that, similar to estrone sulfate (51), sulfated conjugates of LT are an
inactive pool of this flavone and can reach target cells upon
hydrolysis by sulphatases.
As is known, organic anions of the type of sulfated
conjugates of phenolic compounds, with the assistance of designated
transporters (symporters) and a significant proton gradient,
possess the ability to permeate into intestinal epithelial cells,
followed by their rapid transport into blood plasma (8,52). These
specific transporters are members of a large family of organic
anion transporters and are located at the apical part of membranes
of epithelial cells in the small intestine and other organs. They
are responsible for transportation of large amounts of anionic
organic molecules, which differ structurally from each other.
Our research and analysis of literature data
indicates that DSL is able to reach the blood plasma of animals and
humans through the intestine, skipping the stages of modification
in intestinal and liver cells, which increases its bioabsorbability
and effectiveness of its pharmacological action. DSL and LT are
likely to interact with the same target receptors and exert a
pharmacological effect. DSL serves as an inactive pool of LT and
reaches target receptors inside these cells only when hydrolyzed by
sulfatases (enzymes of the esterase class that catalyze the
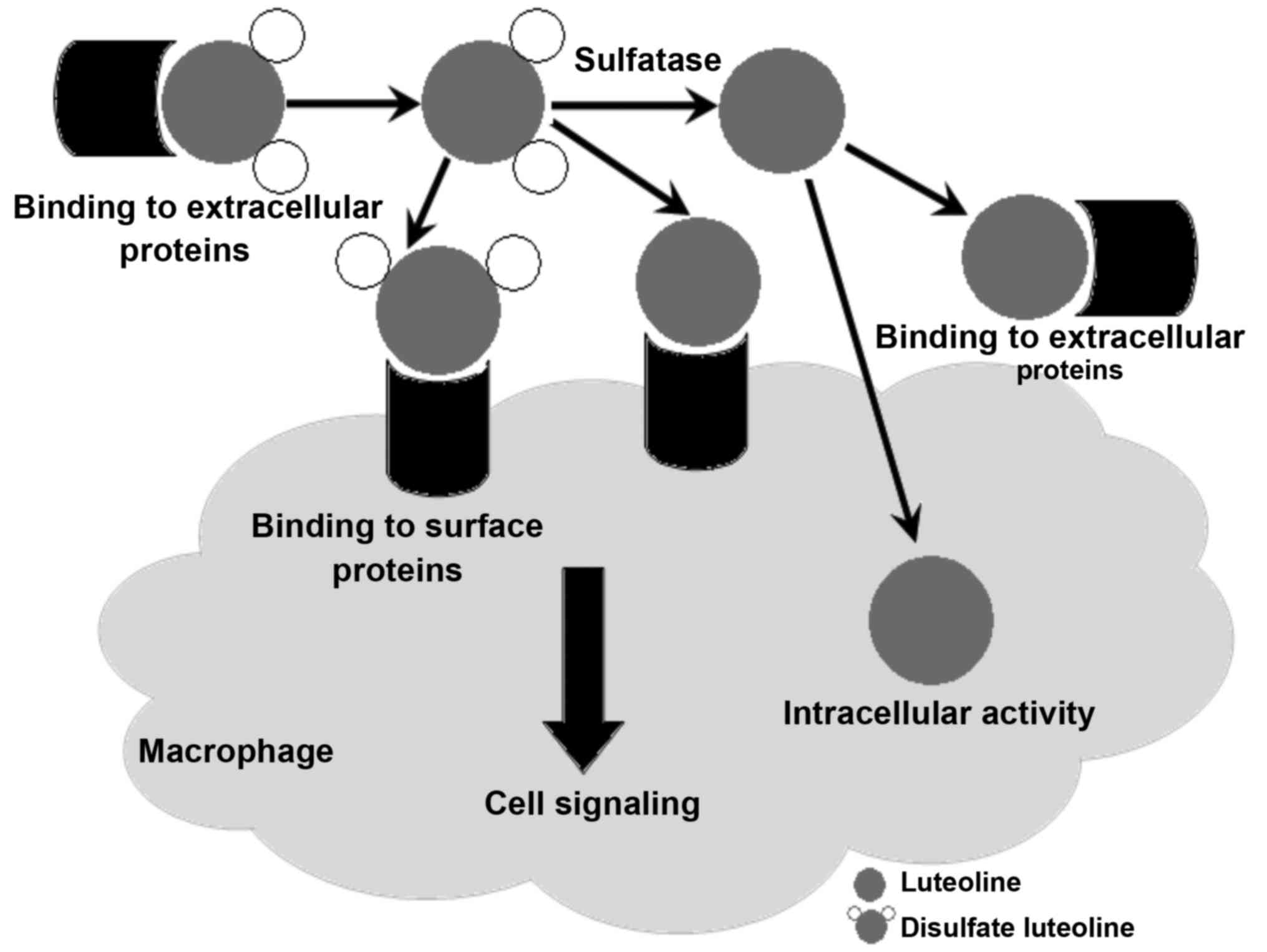
hydrolysis of sulfate esters). Fig.
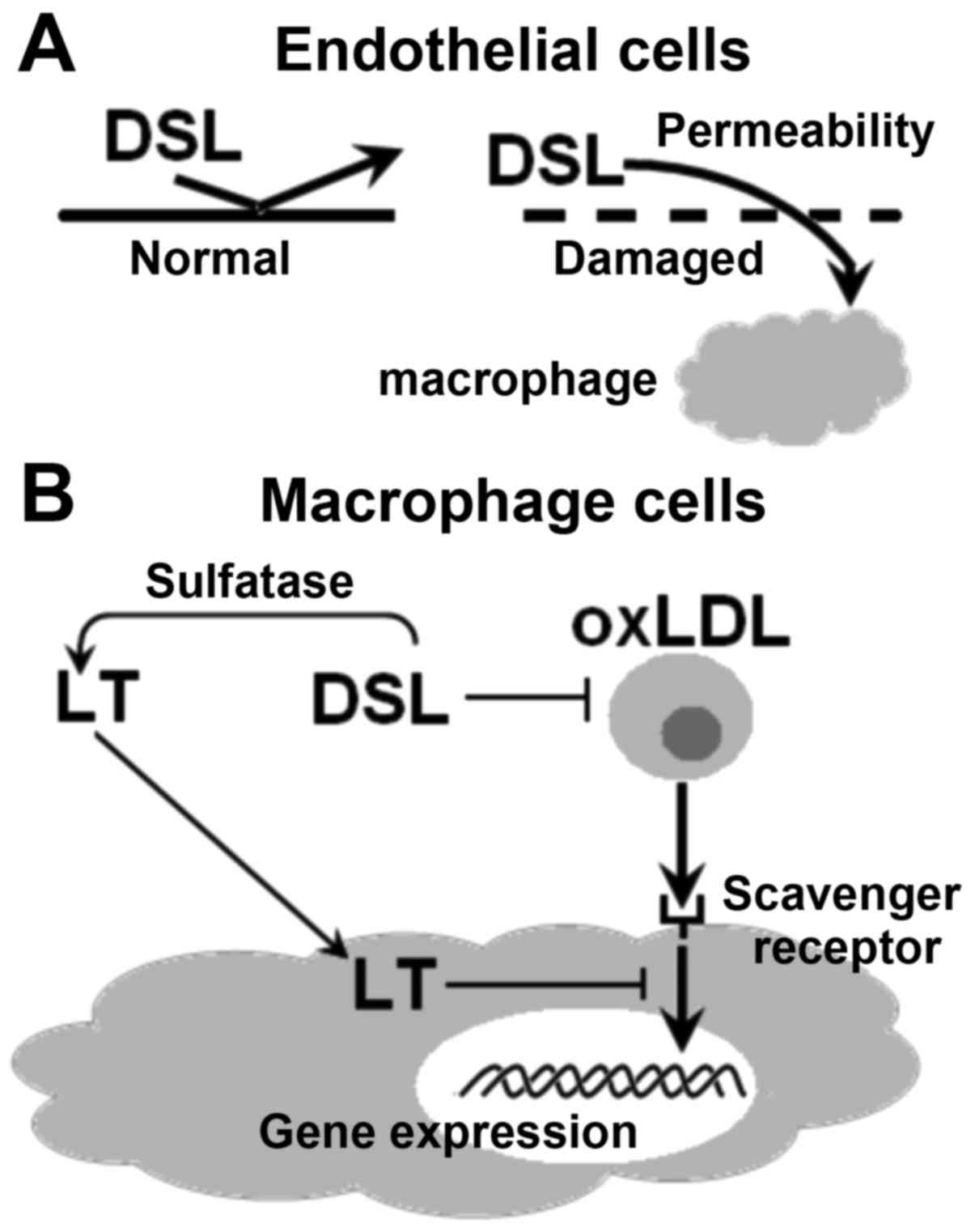
5 shows several variants of interaction of LT and DSL with
receptor protein targets on the plasma membrane of macrophage cells
and inside these cells.
After entering the bloodstream, DSL and LT begin to
interact with endothelial cells of blood vessels, which may be in
the normal or a pathological state (e.g. diabetes, hyperlipidemia
or atherosclerosis-associated cardiovascular diseases) (Fig. 6A). Fig.
6B depicts the proposed pathways of inhibition of endocytosis
of oxidized low-density lipoproteins (oxLDL), mediated by scavenger
receptors (SR), in macrophage cells in the presence of LT and DSL.
It may be assumed that the interaction of DSL with SR, localized on
the plasma membrane of macrophages, selectively blocks the binding
of oxLDL to these receptors and their delivery into the cell. At
the early stages of atherosclerosis, SR are considered the most
important structural components, which are responsible for the
absorption of oxLDL by macrophages, subsequently resulting in the
formation of foam cells (precursors of atherosclerotic plaques).
Thus, the inhibitory effect of LT on this process may also occur
inside a macrophage cell (8,9).
Based on the abovementioned information, the
protective action of LT and DSL in the normal state and in the case
of cardiovascular diseases and T2D may be divided into several
stages: i) Damaged walls of blood vessels allow for entry of
metabolites from circulating blood; ii) DSL is safely absorbed and
subsequently delivered to target cells; iii) DSL interacts with SR
located on macrophages and exerts its the anti-atherosclerotic
effects (Fig. 6).
Conclusions
Inflammation is a systemic protective response of
plants, animals and humans to the effect of pathogenic and stress
factors of the environment (bacteria, viruses and oxidative stress
inducers). In plants, the inflammatory reaction manifests in the
form of a hypersensitive response, while presenting as and immune
response in animals and humans. Numerous polyphenolic compounds,
including RA, LT and DSL, can be considered as endogenous
protective means involved in the control of an induced inflammatory
process in a producer organism.
Since inflammation has a crucial role in the
progression of T2D-associated pathologies, including insulin
resistance, cardiovascular diseases, obesity and certain immune
disorders, the positive therapeutic effects of PPC from Z.
seagrasses and its constituents, RA, LT and DSL, in T2D and
hyperlipidemia models, are plausible, as outlined in the present
review. RA, LT and DSL are used as therapeutic agents exhibit a
high antioxidant, anti-inflammatory and immunomodulatory activity.
It should be noted that, in spite of the lack of highly selective
activity, they have an effect on the functional activity of
numerous cell targets and signaling pathways with multiple
mechanisms of action by regulating the progression of various
inflammatory reactions through direct and indirect effects on ROS
via PPAR-dependent and -independent pathways.
When used for therapeutic purposes, RA, LT and DSL
have unique features regarding the mechanism of their
anti-inflammatory and antioxidant activity; of note, PPC exerted a
more pronounced pharmacological effect than its major components,
RA, LT and DSL, applied separately. For this reason, the use of
combined pharmaceutical agents as PPC (5,26),
acting on several different targets in cells, appears to be a
promising approach for development of remedies of complementary
therapy for preventing and treating multifactorial metabolic
diseases. This approach may overcome the abovementioned problems
regarding undesirable side effects of potent synthetic PPAR
agonists. Future studies are required to further clarify the
mechanisms of action of RA, LT and DSL in T2D and associated
diseases.
Clarification of the ‘intricate pattern’ of redox
reactions and features of interaction of RA, LT and DSL with
various cellular targets in normal and pathological cells and
tissues, depending on factors including their dose, cell type and
pathology, may provide more effective antidiabetic therapies. For
this purpose, the features and functioning of biochemical signaling
pathways and their interactions should be fully elucidated.
Understanding the molecular mechanisms of their antidiabetic action
will facilitate the effective application of RA, LT and DSL for the
prevention and treatment of T2D along with the widely used modern
pharmaceutical drugs.
Acknowledgments
This study was supported by the grant Far Eastern
Branch of the Russian Academy of Sciences (grant no.
N15-I-5-018).
Glossary
Abbreviations
Abbreviations:
|
T2D
|
type 2 diabetes
|
|
MS
|
metabolic syndrome
|
|
BAS
|
biologically active substances
|
|
PPC
|
polyphenol complex
|
|
RA
|
rosmarinic acid
|
|
LT
|
luteolin
|
|
DSL
|
7,3′-disulfate luteolin
|
|
PPARs
|
peroxisome proliferator-activated
receptors
|
|
FFAs
|
free fatty acids
|
|
TZDs
|
thiazolidinediones
|
|
LP
|
lipid peroxidation
|
|
ROS
|
reactive oxygen species
|
|
Nrf2
|
nuclear factor erythroid-2 p45-related
factor 2
|
|
NO
|
nitric oxide
|
|
iNOS
|
inducible nitric oxide synthase
|
|
cPLA2
|
cytosolic phospholipase A2
|
|
LPS
|
lipopolysaccharide
|
|
TLR4
|
Toll-like receptor 4
|
|
NF-κB
|
nuclear factor κB
|
|
JNK
|
c-Jun N-terminal kinases
|
|
TNF-α
|
tumor necrosis factor alpha
|
|
IL
|
interleukin
|
|
SR
|
scavenger receptor
|
|
oxLDL
|
oxidized low-density lipoprotein
|
|
NSAID
|
non-steroidal anti-inflammatory
agent
|
|
SAID
|
steroidal anti-inflammatory agent
|
|
PKC
|
protein kinase C
|
|
PTC
|
protein tyrosine kinase
|
|
MAPK
|
mitogen-activated protein kinase
|
|
COX
|
cyclooxygenase
|
|
LOX
|
lipoxygenase
|
References
|
1
|
Finucane MM, Danaei G and Ezzati M:
Bayesian estimation of population-level trends in measures of
health status. Statistical Sci. 29:18–25. 2014. View Article : Google Scholar
|
|
2
|
Palomer X, Barroso E, Zarei M, Botteri G
and Vázquez-Carrera M: 2PPARβ/δ and lipid metabolism in the heart.
Biochim Biophys Acta. 1860:1569–1578. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tuttolomondo A, Maida C and Pinto A:
Diabetic foot syndrome: Immune-inflammatory features as possible
cardiovascular markers in diabetes. World J Orthop. 6:62–76. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Krylova NV, Popov AM and Leonova GN:
Antioxidants as potential antiviral agents for flavivirus
infections (Review). Antibiotiki i Khimioterapiya. 61:25–31.
2016.(In Russian).
|
|
5
|
Krivoshapko ON, Popov AM, Artyukov AA and
Kostetskiĭ EIa: Features of corrective actions polar lipids from
marine and aquatic bioantioxidants in disorders of lipid and
carbohydrate metabolism. Biomed Khim. 58:189–198. 2012.(In
Russian). View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Krivoshapko ON and Popov AM: Curative and
preventive properties of lipids and antioxidants isolated from
marine hydrobionts. Vopr Pitan. 80:4–8. 2011.(In Russian).
PubMed/NCBI
|
|
7
|
Popov AM and Krivoshapko ON: Protective
Effects of Polar Lipids and Redox-active compounds from marine
organisms at modeling of hyperlipidemia and diabetes. J Biomed Sci
Eng. 6:543–550. 2013. View Article : Google Scholar
|
|
8
|
Popov AM, Krivoshapko ON and Artyukov AA:
Protective mechanisms of the pharmacological activity of
flavonoids. Rus J Biopharm. 4:27–41. 2012.
|
|
9
|
Popov AM, Krivoshapko ON and Artyukov AA:
Comparative evaluation of the pharmacological activity of luteolin
and luteolin 7,3′-disulfate in modeling different pathologies. Rus
J Biopharm. 3:27–33. 2011.
|
|
10
|
Popov AM, Artyukov AA, Krivoshapko ON,
Krylova NV, Leonova GN and Kozlovskaja EP: Medication, processing
antioxidant, cardioprotective, antidiabetic, anti-inflammatory,
hepatoprotective, antitumoral and antiviral action. Patent Russian
Federation 2432959. 2011.
|
|
11
|
Gervois P, Fruchart JC and Staels B: Drug
Insight: Mechanisms of action and therapeutic applications for
agonists of peroxisome proliferator-activated receptors. Nat Clin
Pract Endocrinol Metab. 3:145–156. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mueller M, Lukas B, Novak J, Simoncini T,
Genazzani AR and Janqbauer A: Oregano: A source for peroxisome
proliferator-activated receptor gamma antagonists. J Agric Food
Chem. 56:11621–11630. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsybulsky AV, Popov AM and Artyukov AA:
The Comparative study of the medical action of lyuteolin,
rosmarinic acid and echinochrome A at experimental stress-induced
cardiovascular. Biomed Khim. 57:314–325. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu ZM, Hu M, Chan P and Tomlinson B:
Early investigational drugs targeting PPAR-α for the treatment of
metabolic disease. Expert Opin Investig Drugs. 24:611–621. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xagorari A, Papapetropoulos A, Mauromatis
A, Economou M, Fotsis T and Roussos C: Luteolin inhibits an
endotoxin-stimulated phosphorylation cascade and proinflammatory
cytokine production in macrophages. J Pharmacol Exp Ther.
296:181–187. 2001.PubMed/NCBI
|
|
16
|
Ding L, Jin D and Chen X: Luteolin
enhances insulin sensitivity via activation of PPARγ
transcriptional activity in adipocytes. J Nutr Biochem. 21:941–947.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Park HS, Kim SH, Kim YS, Ryu SY, Hwang JT,
Yang HJ, Kim GH, Kwon DY and Kim MS: Luteolin inhibits adipogenic
differentiation by regulating PPARgamma activation. Biofactors.
35:373–379. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Puhl AC, Bernardes A, Silveira RL, Yuan J,
Campos JL, Saidemberg DM, Palma MS, Cvoro A, Ayers SD, Webb P, et
al: Mode of peroxisome proliferator-activated receptor γ activation
by luteolin. Mol Pharmacol. 81:788–799. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lopez-Lázaro M: Distribution and
biological activities of the flavonoid luteolin. Mini Rev Med Chem.
9:31–59. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kure A, Nakagawa K, Kondo M, Kato S,
Kimura F, Watanabe A, Shoji N, Hatanaka S, Tsushida T and Miyazawa
T: Metabolic fate of Luteolin in rats: Its relationship to
anti-inflammatory effect. J Agric Food Chem. 64:4246–4254. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gamo K, Shiraki T, Matsuura N and Miyachi
H: Mechanism of peroxisome proliferator-activated receptor gamma
(PPARγ) transactivation by hesperetin glucuronides is distinct from
that by a thiazolidine-2,4-dione agent. Chem Pharm Bull (Tokyo).
62:491–493. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Weidner C, Wowro SJ, Freiwald A, Kodelja
V, Abdel-Aziz H, Kelber O and Sauer S: Lemon balm extract causes
potent antihyperglycemic and antihyperlipidemic effects in
insulin-resistant obese mice. Mol Nutr Food Res. 58:903–907. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Molavi B, Rassouli N, Bagwe S and Rasouli
N: A review of thiazolidinediones and metformin in the treatment of
type 2 diabetes with focus on cardiovascular complications. Vasc
Health Risk Manag. 3:967–973. 2007.PubMed/NCBI
|
|
24
|
Lien EJ, Ren S, Bui HH and Wang R:
Quantitative structure-activity relationship analysis of phenolic
antioxidants. Free Radic Biol Med. 26:285–294. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nimnual AS, Taylor LJ and Bar-Sagi D:
Redox-dependent downregulation of Rho by Rac. Nat Cell Biol.
5:236–241. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Popov AM, Krivoshapko ON, Klimovich AA and
Artyukov AA: Biological activity and mechanisms of therapeutic
action of rosmarinic acid, luteolin and its sulphated derivatives.
Biomed Khim. 62:22–30. 2016.(In Russian). View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Popov AM, Osipov AN, Korepanova EA,
Krivoshapko ON, Artyukov AA and Klimovich AA: Study of antioxidant
and membrane activity of luteoline using different model systems.
Biophysics (Russian Federation). 61:1079–1087. 2016.
|
|
28
|
Nagao A, Seki M and Kobayashi H:
Inhibition of xanthine oxidase by flavonoids. Biosci Biotechnol
Biochem. 63:1787–1790. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sen N, Das BB, Ganguly A, Banerjee B, Sen
T and Majumder HK: Leishmania donovani: Intracellular ATP level
regulates apoptosis-like death in luteolin induced dyskinetoplastid
cells. Exp Parasitol. 114:204–214. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Manju V and Nalini N: Chemopreventive
potential of luteolin during colon carcinogenesis induced by
1,2-dimethylhydrazine. Ital J Biochem. 54:268–275. 2005.PubMed/NCBI
|
|
31
|
Robak J, Shridi F, Wolbís M and
Królikowska M: Screening of the influence of flavonoids on
lipoxygenase and cyclooxygenase activity, as well as on nonenzymic
lipid oxidation. Pol J Pharmacol Pharm. 40:451–458. 1988.PubMed/NCBI
|
|
32
|
Paredes-Gonzalez X, Fuentes F, Jeffery S,
Saw CL, Shu L, Su ZY and Kong AN: Induction of Nrf2-mediated gene
expression by dietary phytochemical flavones apigenin and luteolin.
Biopharm Drug Dispos. 36:440–451. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Brown JE and Rice-Evans CA: Luteolin-rich
artichoke extract protects low density lipoprotein from oxidation
in vitro. Free Radic Res. 29:247–255. 1988. View Article : Google Scholar
|
|
34
|
Popov AM, Osipov AN, Korepanova EA,
Krivoshapko ON and Artyukov AA: Study of antioxidant and membrane
activity of rosmarinic acid using different model systems.
Biofizika. 58:775–785. 2013.(In Russian). PubMed/NCBI
|
|
35
|
Sotnikova R, Okruhlicova L, Vlkovicova J,
Navarova J, Gajdacova B, Pivackova L, Fialova S and Krenek P:
Rosmarinic acid administration attenuates diabetes-induced vascular
dysfunction of the rat aorta. J Pharm Pharmacol. 65:713–723. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Harris GK, Qian Y, Leonard SS, Sbarra DC
and Shi X: Luteolin and chrysin differentially inhibit
cyclooxygenase-2 expression and scavenge reactive oxygen species
but similarly inhibit prostaglandin-E2 formation in RAW 264.7
cells. J Nutr. 136:1517–1521. 2006.PubMed/NCBI
|
|
37
|
Fadel O, El Kirat K and Morandat S: The
natural antioxidant rosmarinic acid spontaneously penetrates
membranes to inhibit lipid peroxidation in situ. Biochim Biophys
Acta. 1808:2973–2980. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kawai Y: Immunochemical detection of
food-derived polyphenols in the aorta: Macrophages as a major
target underlying the anti-atherosclerotic activity of polyphenols.
Biosci Biotechnol Biochem. 75:609–617. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lee J, Jung E, Kim Y, Lee J, Park J, Hong
S, Hyun CG, Park D and Kim YS: Rosmarinic acid as a downstream
inhibitor of IKK-beta in TNF-alpha-induced upregulation of CCL11
and CCR3. J Pharmacol. 148:366–375. 2006.
|
|
40
|
Ando C, Takahashi N, Hirai S, Nishimura K,
Lin S, Uemura T, Goto T, Yu R, Nakagami J, Murakami S and Kawada T:
Luteolin, a food-derived flavonoid, suppresses adipocyte-dependent
activation of macrophages by inhibiting JNK activation. FEBS Lett.
583:3649–3654. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hirai S, Takahashi N, Goto T, Lin S,
Uemura T, Yu R and Kawada T: Functional food targeting the
regulation of obesity-induced inflammatory responses and
pathologies. Mediators Inflamm. 2010:3678382010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Köhle C and Bock KW: Activation of coupled
Ah receptor and Nrf2 gene batteries by dietary phytochemicals in
relation to chemoprevention. Biochem Pharmacol. 72:795–805. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Govindaraj J and Pillai Sorimuthu S:
Rosmarinic acid modulates the antioxidant status and protects
pancreatic tissues from glucolipotoxicity mediated oxidative stress
in high-fat diet: Streptozotocin-induced diabetic rats. Mol Cell
Biochem. 404:143–159. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hirsch HA, Ilipoulos D and Struhl K:
Metformin inhibits the inflammatory response associated with
cellular transformation and cancer stem cell growth. Proc Natl Acad
Sci USA. 110:972–977. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Watson JD: Types diabetes as a redox
disease. Lancet. 383:841–843. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ristow M, Zarse K, Oberbach A, Klöting N,
Birringer M, Kiehntopf M, Stumvoll M, Kahn CR and Blüher M:
Antioxidants prevent health-promoting effects of physical exercise
in humans. Proc Natl Acad Sci USA. 106:8665–8670. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shi RX, Ong CN and Shen HM: Luteolin
sensitizes tumor necrosis factor-alpha-induced apoptosis in human
tumor cells. Oncogene. 23:7712–7721. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ju W, Wang X, Shi H, Chen W, Belinsky SA
and Lin Y: A critical role of luteolin-induced reactive oxygen
species in blockage of tumor necrosis factor-activated nuclear
factor-kappaB pathway and sensitization of apoptosis in lung cancer
cells. Mol Pharmacol. 71:1381–1388. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mukinda JT, Syce JA, Fisher D and Meyer M:
Effect of the Plant Matrix on the Uptake of Luteolin
Derivatives-containing Artemisia afra Aqueous-extract in Caco-2
cells. J Ethnopharmacol. 130:439–449. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hoshino J, Park EJ, Kondratyuk TP, Marler
L, Pezzuto JM, van Breemen RB, Mo S, Li Y and Cushman M: Selective
synthesis and biological evaluation of sulfate-conjugated
resveratrol metabolites. J Med Chem. 53:5033–5043. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Santner SJ, Feil PD and Santen RJ: In situ
estrogen production via estrone sulphatase pathway in breast
tumours: Relative importance versus the aromatase pathway. J Clin
Endocrinol Metab. 59:29–33. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Burckhardt G and Burckhardt BC: In vitro
and in vivo evidence of the importance of organic anion
transporters (OATs) in drug therapy. Handb Exp Pharmacol. 29–104.
2011. View Article : Google Scholar : PubMed/NCBI
|