Introduction
As novel cancer-targeting drugs, cyclin-dependent
kinase (CDK) inhibitors are highly anticipated (1). Over the past decade, numerous studies
have implicated roscovitine as an anticancer drug with promising
therapeutic properties (2).
Roscovitine was found to inhibit cell division control protein 2,
cdk2 or cdk5 kinase activities (3,4), and was
shown to directly inhibit nuclear factor-κb activation in cancer
cell lines (5).
Additional studies have demonstrated further
properties of cyclin-dependent kinase inhibitors (6) or roscovitine (7); for instance, roscovitine downregulated
the expression of myeloid cell leukemia (Mcl)-1 protein, which is
likely involved in inflammatory processes (7). Other studies have shown that
upregulation of adenosine monophosphate-activated protein kinase
(AMPK) activation also inhibited the expression of Mcl-1 (8–10). AMPK
is an important regulator of energy homeostasis in cells. In recent
years, studies have demonstrated that AMPK has an important role in
modulating inflammation in inflammatory pulmonary diseases,
including asthma, pulmonary infectious diseases and pulmonary
fibrosis (11–14). These results suggested that
roscovitine may have anti-inflammatory effects; while it has
remained elusive whether it also inhibits inflammation in murine
Leydig cells.
Leydig cells are distributed in the loose connective
tissue between seminiferous tubules. The main function of Leydig
cells is the synthesis and secretion of androgen, which has
important roles in male reproductive function (15). Inflammation can affect the normal
function of Leydig cells (16).
Lipopolysaccharide (LPS) was used to establish a mouse model of
inflammation (17) and the testes
were collected to investigate the effects of roscovitine on murine
testis inflammation.
Materials and methods
Animals and ethics statement
A total of 16, 2-month-old male C57BL/6 mice (four
groups, n=4 each group, weight range from 20–25 g) were obtained
from the Shanghai Laboratory Animal Company (Shanghai, China). The
mice were housed at 21±2°C with 55±10% humidity under a 12-h
light/dark cycle. Chow and water were available ad libitum. All
animal procedures were reviewed and approved by the Animal Care
Committee of Tongji University (Shanghai, China).
Experimental groups
Mice were divided into four groups (n=4 each):
Control group, case group, LPS group and dimethylsulfoxide (DMSO)
group. In the case group, each mouse was injected i.p. with
roscovitine (S1153; Selleck Chemicals, Houston, TX, USA) at 5 mg/kg
dissolved in DMSO (Sigma-Aldrich, Merck Millipore, Darmstadt,
Germany), and LPS (Sigma-Aldrich, Merck Millipore) at 3 mg/kg
dissolved in saline (0.9% NaCl). Animals in the LPS group received
LPS only (5 mg/kg). Animals in the DMSO and control groups only
received equivalent volumes of DMSO and saline, respectively. After
12 h, mice were sacrificed by carbon dioxide anesthesia (SMQ-II;
Tianhuan Technology, Co., Ltd., Shanghai, China) with a final
concentration of 80–100% CO2 in a cage 730×560×600 mm in
size. Mortality was confirmed following 5 min of observation.
Venous blood was collected from the caudal vein and the testis was
removed for further examination.
Behavior and weight of mice
The behavior of the mice was observed and recorded
prior to and after drug treatment, and the mice were weighed using
an electronic balance at the same time-points.
Observation of histological
changes
Mouse testis were fixed at room temperature by
soaking overnight in 2% paraformaldehyde and transferred by
gradient ethanol (from 50 to 100%) and embedded into paraffin.
Sections were sliced to 5 µm and stained with hematoxylin and
eosin. Tissue sections were also stained immunohistochemically to
observe specific expression of 3β-hydroxysteroid dehydrogenase
(3β-HSD) in testis tissue after the indicated treatments. The
dilution ratio of Anti-HSD3B1 antibody (catalogue no. ab55268;
Abcam, Cambridge, UK) was 1:1,000. Histology of testicular tissues
was observed by light microscopy (Eclipse E100, Nikon, Tokyo,
Japan).
Serum testosterone levels
Venous blood was extracted from the mice and blood
was allowed to coagulate for 12 h at room temperature, followed by
centrifugation for 10 min at 4°C at 1006.2 × g (Centrifuge 5408R;
Eppendorf, Hamburg, Germany). The upper serum layer was assessed
using an ELISA kit (cat. no. YC30087; Yuanchuang Bio-Chemical Co.,
Ltd., Shanghai, China) to quantify testosterone levels according to
the manufacturer's instructions.
Cell culture and reagents
Mouse testes were collected to extract primary
Leydig cells for RNA and protein isolation. Furthermore, Leydig
cells were isolated and maintained in Dulbecco's modified Eagle's
medium (Sigma-Aldrich, Merck-Millipore) supplemented with 10% fetal
bovine serum (Gibco, Thermo Fisher Scientific, Inc., Waltham, MA,
USA), 100 U/ml penicillin and 100 µg/ml streptomycin (18).
RNA analysis by reverse-transcription
quantitative polymerase chain reaction (RT-qPCR)
Total RNA from cells was extracted using TRIzol
(Life Technologies, Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. RNA samples were
reverse-transcribed into complementary DNA using the Mx3005P qPCR
system (Agilent Technologies, Inc., Santa Clara, CA, USA). The PCR
reaction was completed as follows: 94°C for 5 min followed by 94°C
for 30 sec, 55–58°C for 30 sec, 72°C for 30 sec-1 min and 70°C for
10 min at 4°C for 30 cycles. PCR reaction mixtures contained 10 ml
SYBR Green Premix Ex Taq (Takara Bio Inc., Dalian, China). Relative
gene expression levels were normalized to GAPDH mRNA levels in each
sample. Melting curve analysis for each primer set revealed only
one peak for each product. The Applied Biosystems 7900HT fast
real-time system (Thermo Fisher Scientific, Inc.) was used. Primer
sequences are listed in Table I
(Takara, Dalian, China) (19) and
quantification was completed using the 2(−Delta Delta C(T)) method
(20).
 | Table I.Primer sequences used for PCR. |
Table I.
Primer sequences used for PCR.
| Gene | Primer (5′-3′) |
|---|
| Hsd17b3 | F:
ATTTTACCAGAGAAGACATCT |
|
| R:
GGGGTCAGCACCTGAATAATG |
| Cyp17a1 | F:
CCAGGACCCAAGTGTGTTCT |
|
| R:
CCTGATACGAAGCACTTCTCG |
| Cyp11a1 | F:
AGGTGTAGCTCAGGACTTCA |
|
| R:
AGGAGGCTATAAAGGACACC |
| Srd5a1 | F:
CACATCCTGCGGAATCTGA |
|
| R:
TGCTGCCTCGCTCTGGT |
| Cox2 | F:
TTCAACACACTCTATCACTGGC |
|
| R:
AGAAGCGTTTGCGGTACTCAT |
| iNOS | F:
GTTCTCAGCCCAACAATACAAGA |
|
| R:
GTGGACGGGTCGATGTCAC |
| GAPDH | F:
AGGTCGGTGTGAACGGATTTG |
|
| R:
TGTAGACCATGTAGTTGAGGTCA |
Western blot analysis
After the indicated treatments, Leydig cells were
lysed in lysis buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 5 mM
EDTA, 1 mM phenylmethane sulfonyl fluoride and 1% Triton X-100).
Samples were fractionated by 12% SDS-PAGE and transferred onto
polyvinylidene difluoride membranes (Millipore Corp., Bedford, MA,
USA). The membranes were blocked with 10% nonfat milk and incubated
at room temperature with primary antibodies for 2 h, followed by
incubation with horseradish peroxidase-conjugated secondary
antibodies (Li-cor Biosciences, Lincoln).
NE, USA; catalogue no. 926-32211, 926-68021) at a
dilution of 1:10,000 at room temperature for 1 h. Images were
obtained using Odyssey (LI-COR Biosciences, Lincoln, NE, USA).
Membranes were then stripped and re-probed with GAPDH antibody to
ensure equal protein loading. TNF-α (D2D4, 1:1,000) and AMPK
(catalogue no. 23A3, 1:1,000) antibodies were obtained from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Interleukin (IL)-1β
antibody (catalogue no. CC36131) at a dilution of 1:1,000 was
purchased from Bioworld Technology (Nanjing, China). Odyssey
infrared imaging system (version 3.0.21; LI-COR Biosciences,
Lincoln, NE, USA) was used for densitometry.
Statistical analysis
Values are expressed as the mean ± standard error of
the mean. Statistical differences were analyzed with Student's
t-test or one-way analysis of variance if there were more than two
groups. P<0.05 was considered to indicate a statistically
significant difference. Statistical analysis was performed using
Statistical Package for Social Sciences (SPSS) version 12.0 (SPSS
Inc., Chicago, IL, USA).
Results
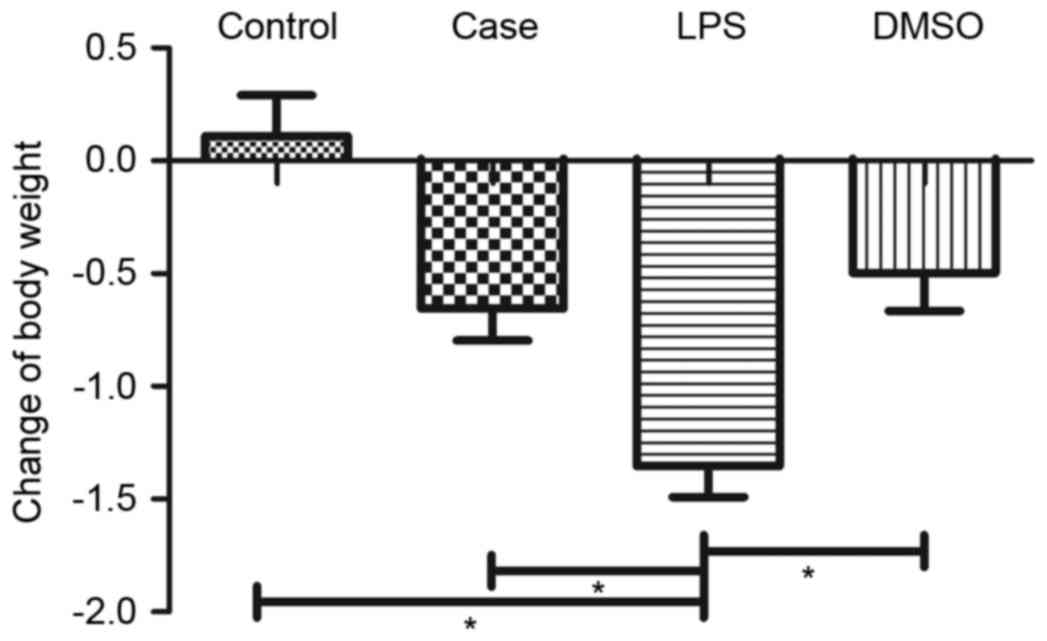
Behavior and body weight of mice
The behavior of mice in the LPS group showed obvious
changes at 12 h after injection of LPS. They showed less activity
and eating, and their body weight was reduced (Fig. 1). In the DMSO and case groups, only
minor weight reductions were observed, while no reduction was
present in the control group (Fig.
1).
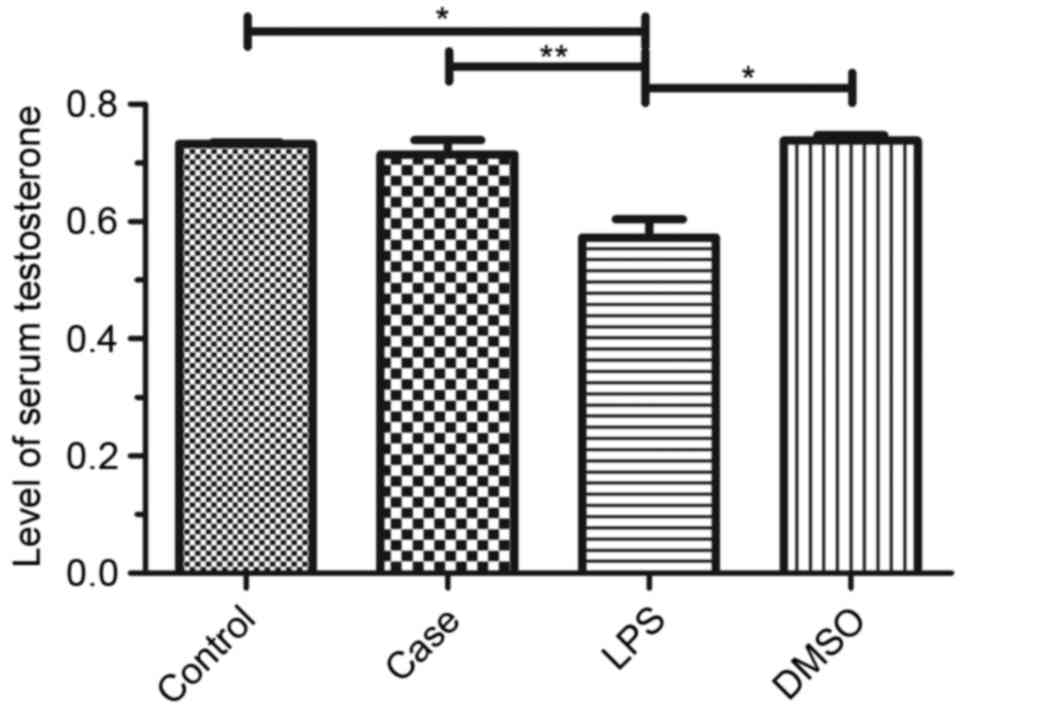
Serum testosterone levels
Venous blood was collected from the mice and serum
testosterone levels were detected using an ELISA kit. Testosterone
levels were significantly reduced in the LPS group compared to that
in the control, with only a minor reduction in the case group and
no reduction in the DMSO group (Fig.
2).
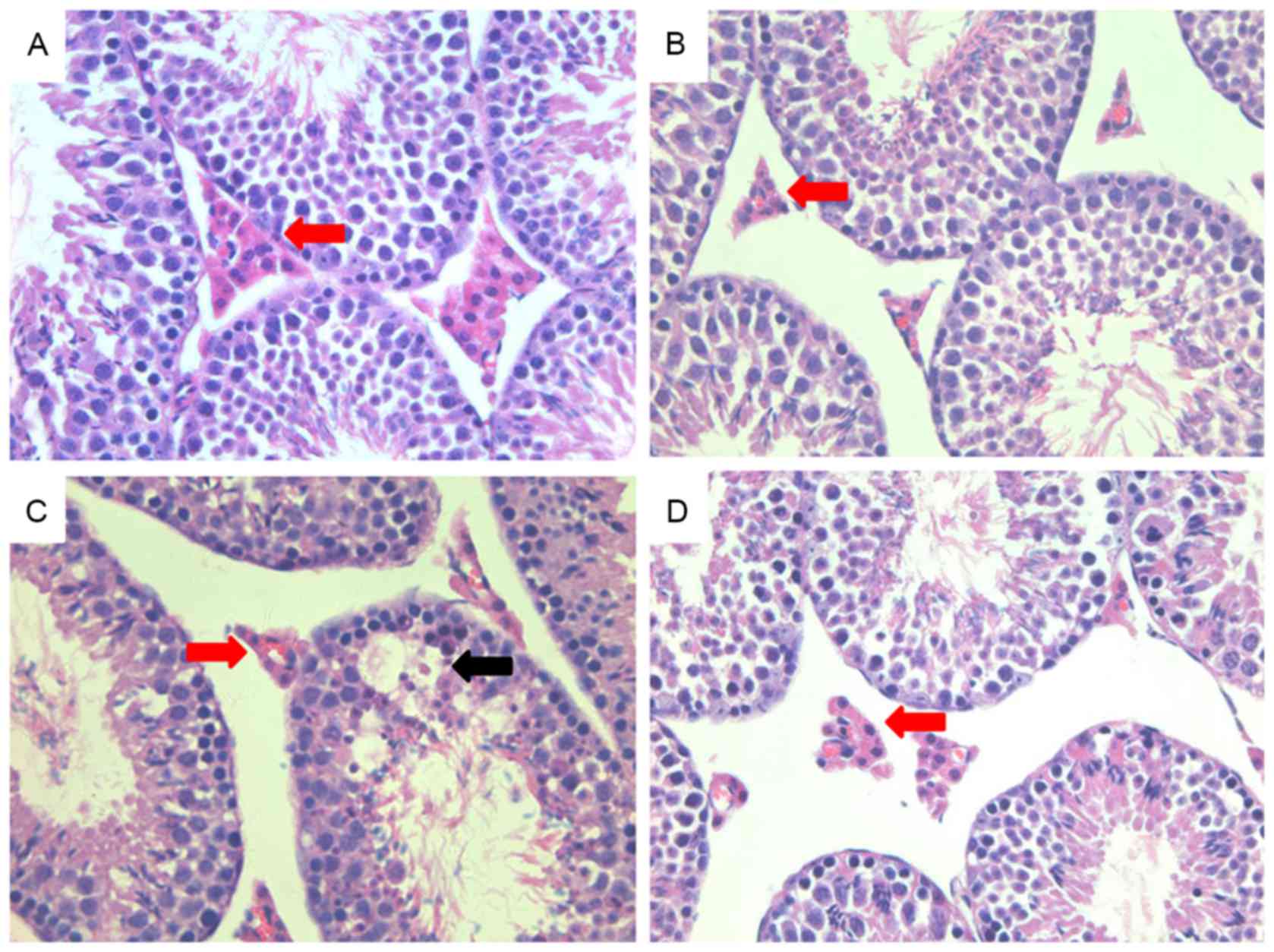
Histology of mouse testes
Hematoxylin and eosin staining revealed that the
structure of the testis was intact in the control animals (Fig. 3A), with a large number of Leydig
cells localized to the gaps between convoluted seminiferous
tubules. In the LPS group, the structure of the testis was altered
and fewer Leydig cells were present, while the case group treated
with roscovitine showed partial rescue of the changes induced by
LPS (Fig. 3B and C). There were no
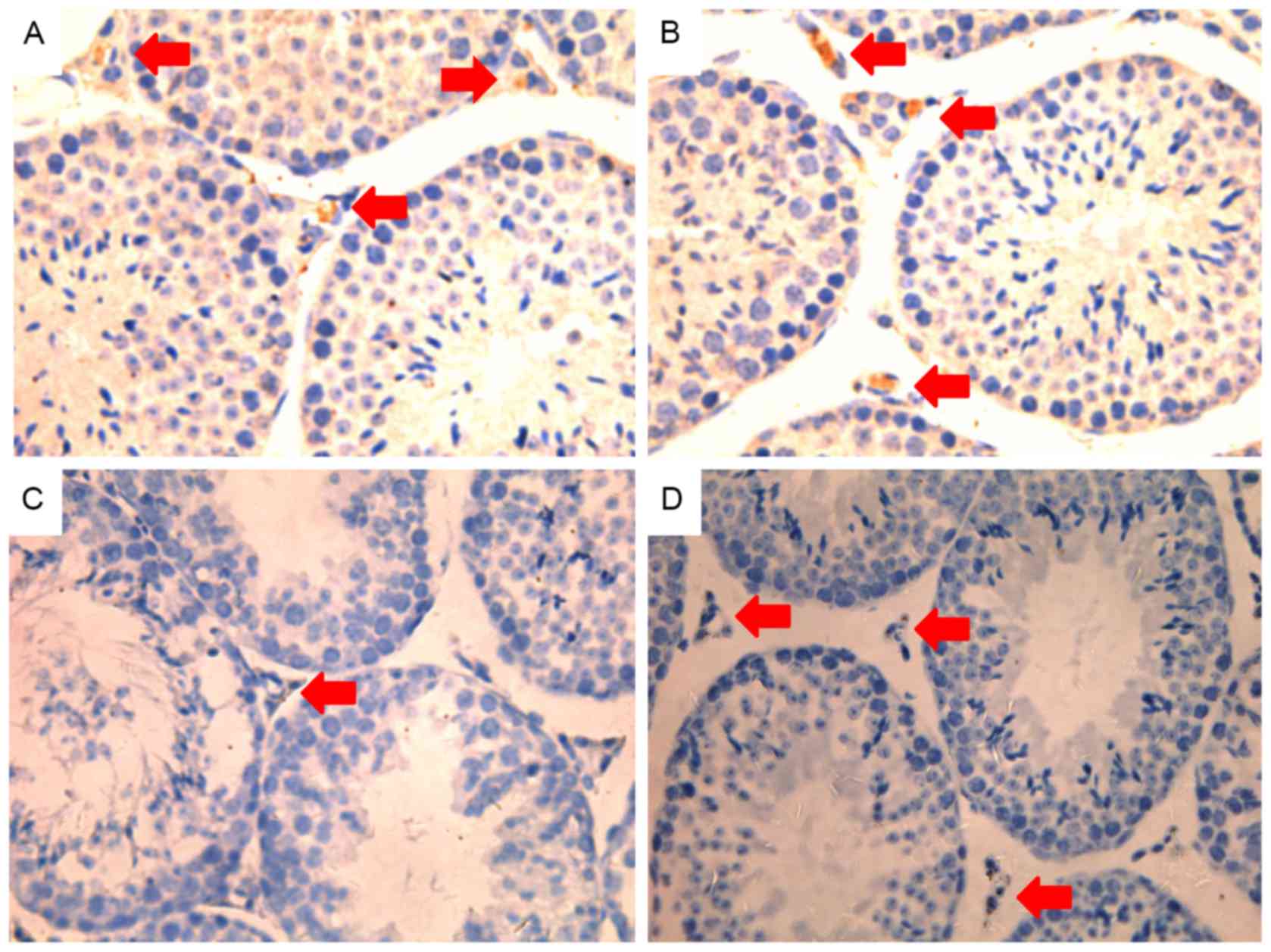
obvious morphological changes in the DMSO group (Fig. 3D). In addition, the expression of
3βHSD was observed by immunohistochemical staining (Fig. 4), revealing a marked decrease in the
LPS group compared to that in the other groups.
Roscovitine inhibits the expression of
proinflammatory cytokines
As roscovitine was observed to exert protective
effects on Leydig cells, it was hypothesized that roscovitine
inhibits inflammatory and proinflammatory signaling pathways. To
examine this hypothesis, western blot and RT-qPCR analyses were
performed to detect inflammatory proteins and mRNAs in Leydig cells
from mice of the various treatment groups. TNF-α and IL-1β were
significantly reduced in the case group compared to the LPS group
(Fig. 5A and B).
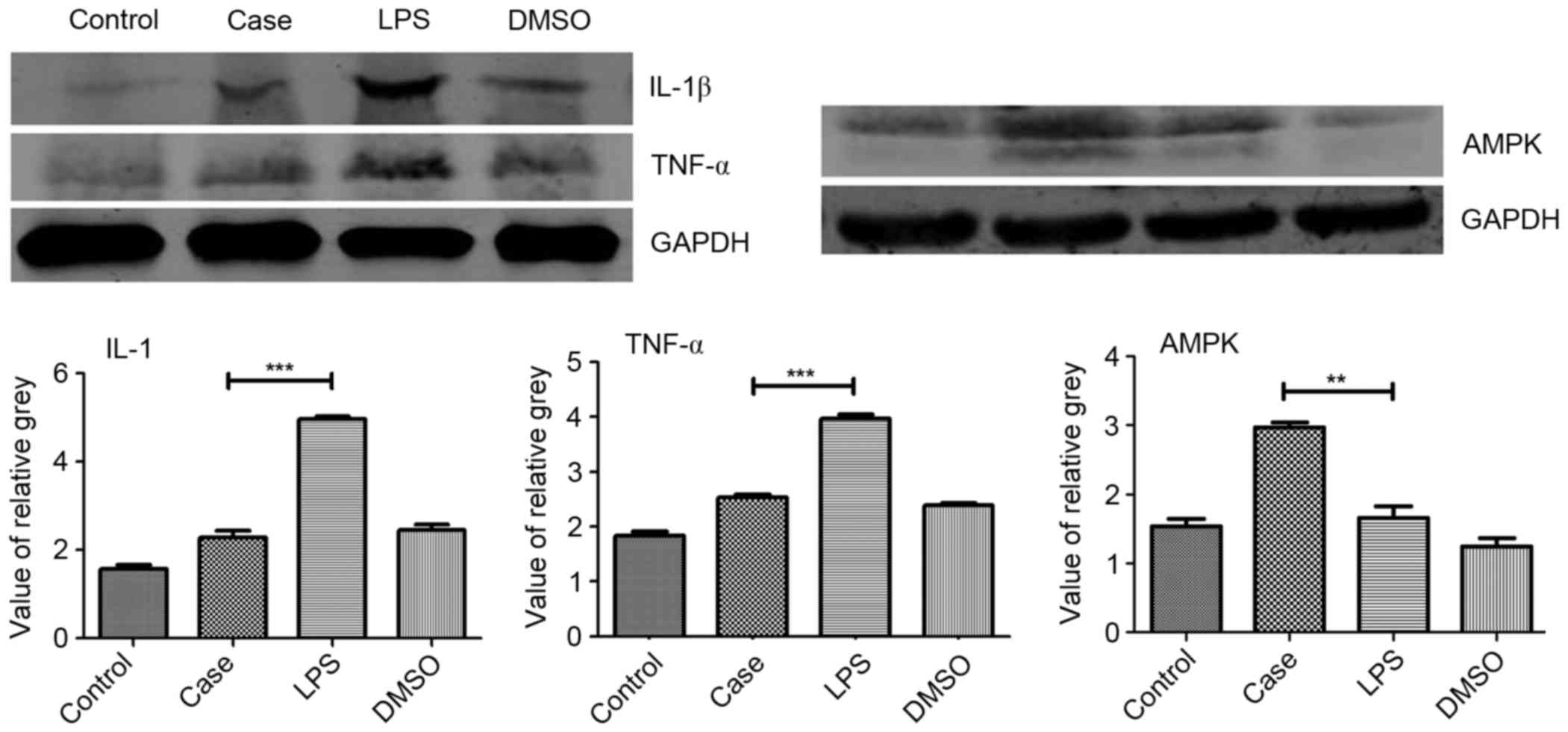 | Figure 5.Protein expression of IL-1β, TNF-α and
AMPK. Protein levels of IL-1β, TNF-α and AMPK were determined by
western blot analysis of testicular tissue from each group 12 h
after each treatment. Roscovitine was found to significantly
increase the expression of AMPK compared with that in the other
three groups. Furthermore, IL-1β and TNF-α expression in the case
group were significantly downregulated compared with that in the
LPS group. **P<0.01; ***P<0.001. Groups: Control, injected
with saline; case, injected with roscovitine (5 mg/kg) dissolved in
DMSO followed by injection of 5 mg/kg LPS in saline; LPS, injected
with LPS only; DMSO, injected with DMSO only; LPS,
lipopolysaccharide; DMSO, dimethyl sulfoxide; TNF, tumor necrosis
factor; IL, interleukin; AMPK, adenosine monophosphate-activated
protein kinase. |
Roscovitine inhibits inflammation by
upregulating AMPK
The western blot analysis results showed an obvious
upregulation of AMPK levels in the case group compared to that in
the other groups. Compared with the control group, a slight
upregulation of AMPK expression was observed in the LPS group as
well as a downregulation in the DMSO group (Fig. 5C).
Roscovitine inhibit the
pro-inflammatory gene
The protective effect of roscovitine in the case
group was further confirmed by the inhibition of the
pro-inflammatory genes cyclooxygenase (COX) −2 and inducible nitric
oxide synthase (iNOS; Fig. 6A and
B).
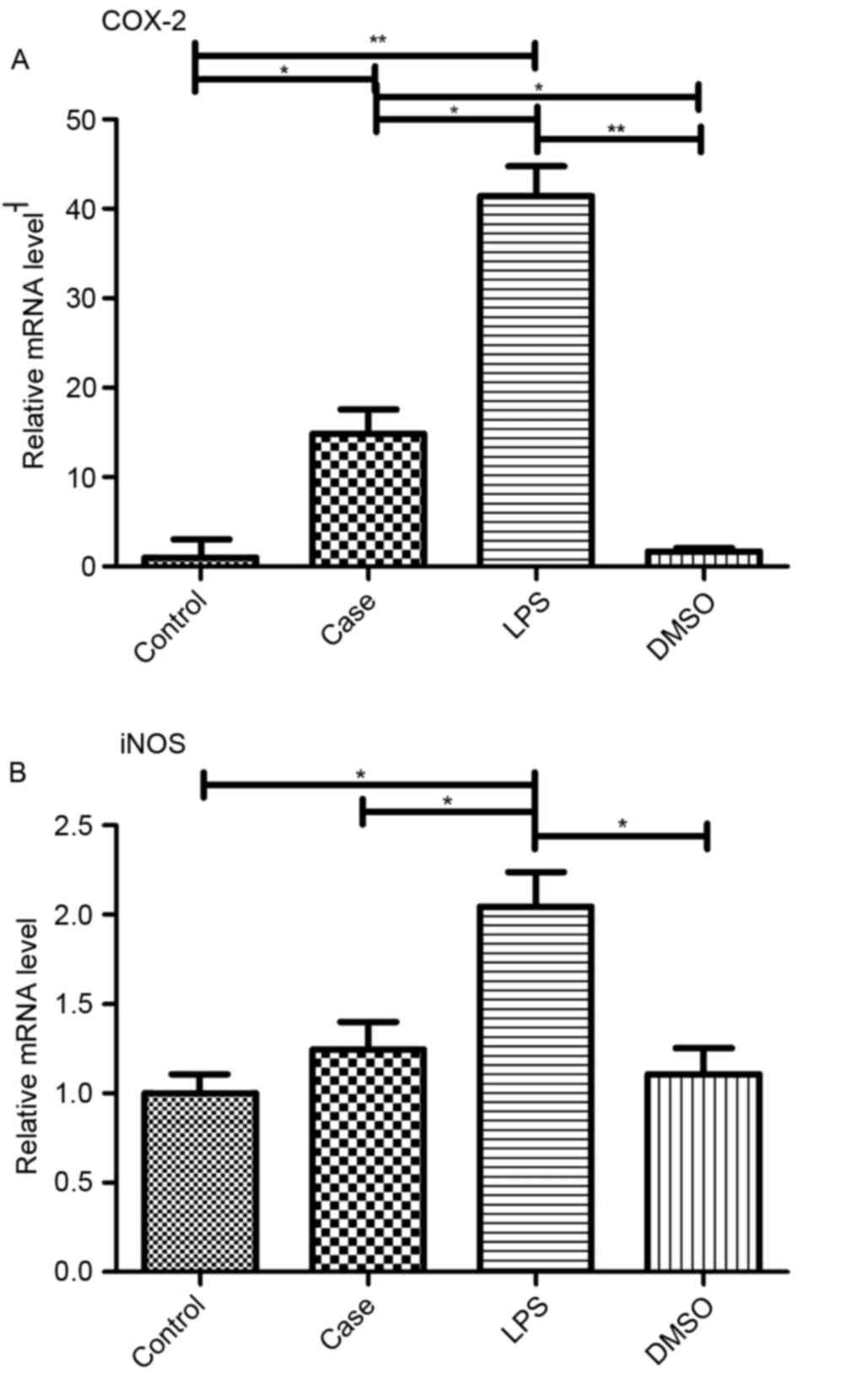 | Figure 6.Transcriptional regulation of COX-2
and iNOS in Leydig cells. Gene expression of (A) COX-2 and (B) iNOS
was examined by reverse-transcription quantitative polymerase chain
reaction analysis 12 h after the indicated treatments. LPS alone
significantly increased the expression of COX-2 and iNOS, which was
inhibited by roscovitine, while COX-2 mRNA levels were still higher
than those in the control and DMSO groups. *P<0.05; **P<0.01.
Groups: Control, injected with saline; case, injected with
roscovitine (5 mg/kg) dissolved in DMSO followed by injection of 5
mg/kg LPS in saline; LPS, injected with LPS only; DMSO, injected
with DMSO only; LPS, lipopolysaccharide; DMSO, dimethyl sulfoxide;
COX, cyclooxygenase; iNOS, inducible nitric oxide synthase. |
Roscovitine rescues LPS-induced
inhibition of testosterone synthesis gene transcription
RT-qPCR was used to assess the effect of roscovitine
on the transcription of the key reproduction-associated genes
Hsd17b3, cytochrome P450 (Cyp) 17a1, Cyp11a1 and steroid 5
alpha-reductase 1 (Srd5a1). As shown in Fig. 7, LPS treatment significantly reduced
the mRNA levels of each of these genes, while roscovitine partly
attenuated these decreases, which was significant for Hsd17b3 and
Cyp17a1 (Fig. 7A and B), but not for
Cyp11a1 and Srd5a1 (Fig. 7C and D).
There was a significant change in Srd5a1 expression in the DMSO
group compared with that in the control group (Fig. 7C).
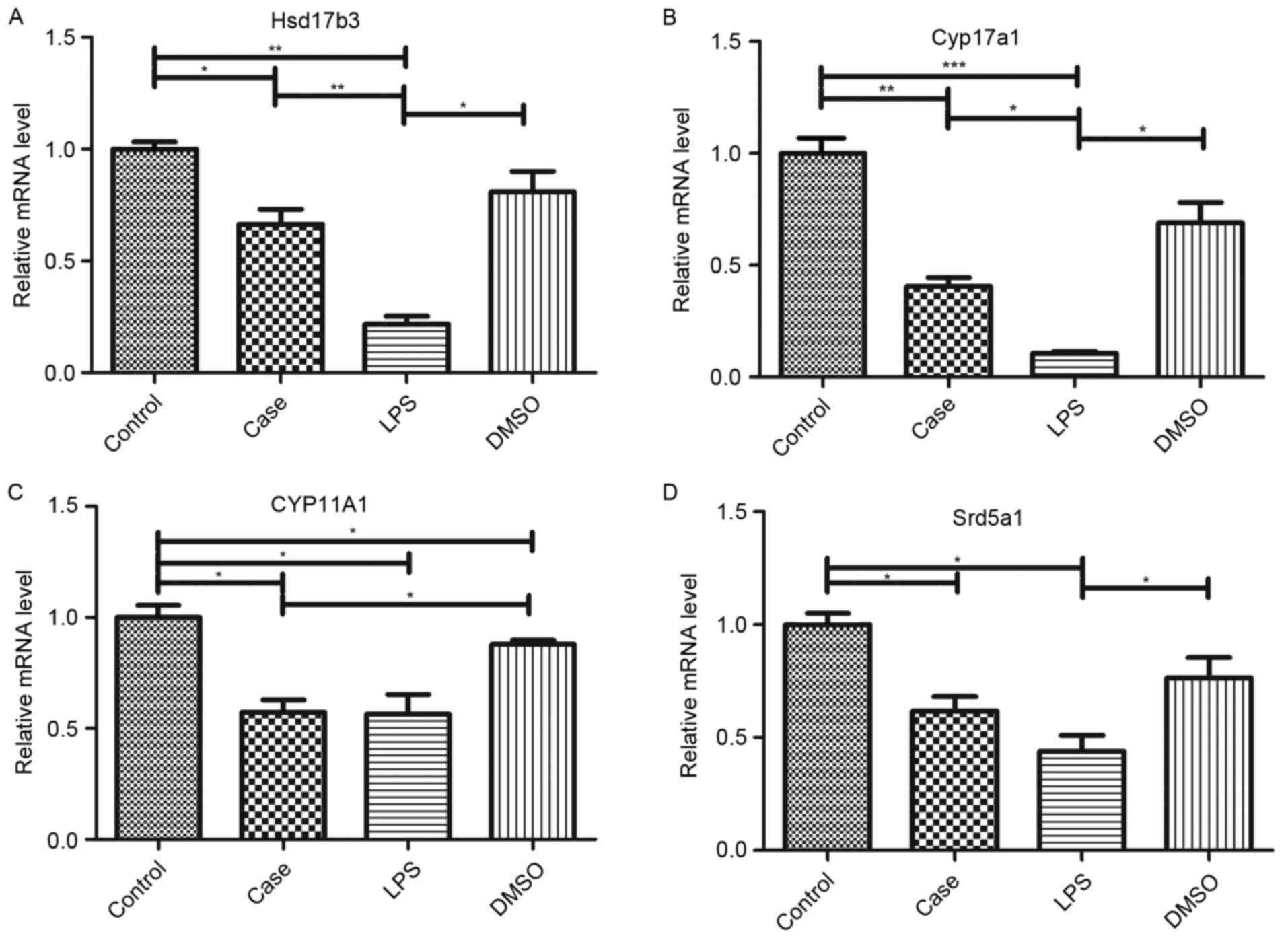 | Figure 7.mRNA expression of testosterone
synthesis genes. mRNA levels of (A) Hsd17b3, (B) Cyp17a1, (C)
Cyp11a1 and (D) Srd5a1 were determined by reverse-transcription
quantitative polymerase chain reaction analysis. While all genes
were downregulated in the LPS group, this was preserved by
co-treatment with roscovitine. The preserving effect was
significant for Hsd17b3 and Cyp17a1 but not for Cyp11a1 and Srd5a1.
*P<0.05; **P<0.01; ***P<0.001. Groups: Control, injected
with saline; case, injected with roscovitine (5 mg/kg) dissolved in
DMSO followed by injection of 5 mg/kg LPS in saline; LPS, injected
with LPS only; DMSO, injected with DMSO only; LPS,
lipopolysaccharide; DMSO, dimethyl sulfoxide; hsd, hydroxysteroid
dehydrogenase; CYP, cytochrome P450; Srd5a1, steroid 5
alpha-reductase 1. |
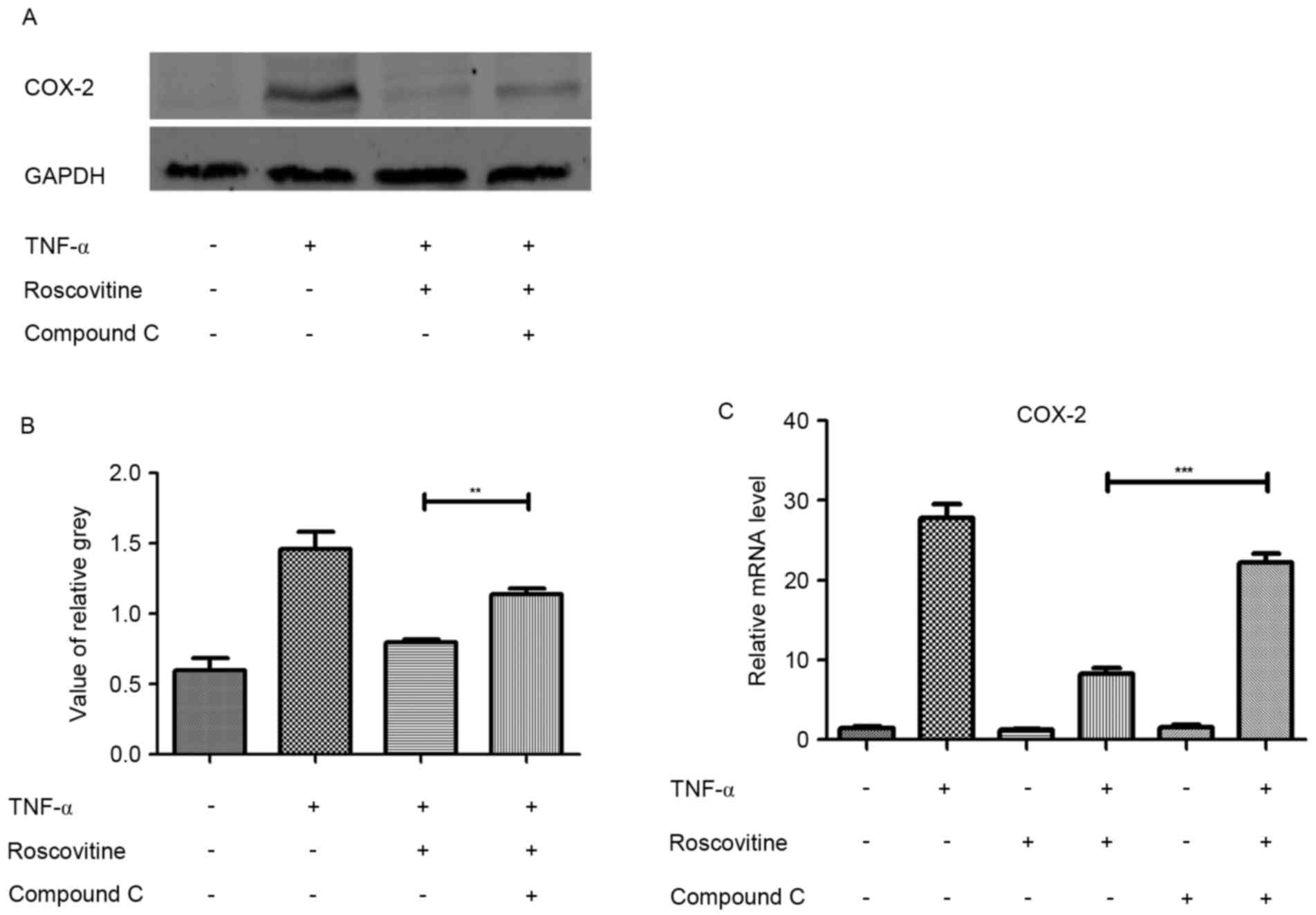
AMPK inhibitor reverses the effect of
roscovitine
Leydig cells were induced with TNF-α and pretreated
with or without AMPK inhibitor (compound C) for 1 h. Subsequently
western blot and RT-qPCR were used to detect changes of the
inflammatory factor COX-2. The results revealed that AMPK inhibitor
reversed the inhibitory effect of roscovitine (Fig. 8).
Discussion
Inflammation in the male reproductive system has
been considered to be one of the causes of male infertility
(21–23). Infection of the male reproductive
system can affect the growth of the testis through direct damage to
testicular sperm tissue or lead to the obstruction of sperm
delivery ducts (24). Leydig cells
are important for the maintenance of male reproductive function.
Hong et al (15) found that
inflammation can inhibit the function of Leydig cells. Thus, it is
important to further study inflammation and anti-inflammatory
mechanisms. To explore possible treatments, the present study
established an experimental inflammation model in mice employing
LPS as the most commonly used reagent for producing inflammation
(17). The present study was using
this mode following the successful experience of a previous study
(19).
As a CDK inhibitor, roscovitine is a compound that
competes for adenosine triphosphate binding sites on CDK (25). It has been demonstrated that
roscovitine has significant anti-tumor effects (26–28) and
can downregulate Mcl-1 expression (7). Other studies have confirmed that
upregulation of AMPK activation inhibits the expression of Mcl-1
(8–10). In the present study, it was observed
that roscovitine protected mice and their testes from inflammatory
injury. Compared with the LPS group, mice in the case group were
more active and had less weight reduction following drug treatment.
Moreover, LPS induced damage in mouse testes and reduced 3β-HSD
expression, which was attenuated by roscovitine. 3β-HSD is a key
enzyme regulating dihydrotestosterone generation and is a good
marker for Leydig cells (29–31).
Serum testosterone levels were then determined in
the experimental groups, revealing LPS significantly decreased
testosterone levels (P<0.05), with no significant change in the
other three groups. Xue et al (32) previously reported that inflammation
induced by LPS affects the levels of androgen in the
microenvironment. It was therefore speculated that roscovitine may
have a protective effect on androgen production in the testis.
Further study using the present inflammation model
revealed that roscovitine downregulated the expression of
inflammatory factors and proinflammatory genes, including TNF-α,
IL-1β, COX-2 and iNOS. Inflammatory cytokines are mainly produced
by activated lymphocytes and monocytes and are important in
mediating inflammatory responses. Inflammatory cytokines such as
IL-1β and TNF-α, secreted by endotoxin and immune complexes, may
cause damage via autocrine and paracrine effects. COX-2 is an
inducible enzyme similar to ring oxidase. It is highly expressed in
inflammatory cells and catalyzes the synthesis of prostaglandin E2
and other products at inflammatory sites to promote an inflammatory
reaction resulting in tissue damage. Chen et al (33) found that LPS induced inflammatory
reactions in RAW264.7 cells, as well as overexpression of COX-2 and
iNOS. Yang et al (34) used
LPS to treat microglial cell lines and observed overexpression of
COX-2 and iNOS, which was inhibited by application of curcumin.
The pathways via which roscovitine inhibits
inflammation were further examined, revealing that the
roscovitine-induced overexpression of AMPK was inversely correlated
with the production of inflammatory factors. AMPK is a cellular
energy regulator. Studies have found that AMPK has a significant
inhibitory effect on inflammatory cells, inflammatory factors and
inflammation pathways (35–40). Therefore, AMPK and its associated
signaling may be important pharmacological targets in the treatment
of inflammatory diseases.
The present also investigated the expression of
reproduction-associated genes in the experimental groups. Hsd17b3
and Cyp17a1 are key enzymes in testosterone synthesis. Yu et
al (40) used dibutylphthalate
to inhibit the expression of the Cyp17a1 gene, which reduced
testicular rostenedione levels and inhibited testosterone synthesis
in Leydig cells. A reduction in testosterone was also detected in
male mice treated with LPS, and it was suggested that LPS treatment
inhibited the expression of testosterone synthase. The results of
the present study, showing diminished mRNA levels of Hsd17b3 and
Cyp17a1 after LPS exposure, are consistent with this hypothesis. No
significant affect was observed for Cyp11a1 and Srd5a1. However,
roscovitine was demonstrated to inhibit these LPS-induced
reductions.
In conclusion, to the best of our knowledge, the
present study was the first to suggest that roscovitine has a
protective role against inflammation in murine Leydig cells. The
mechanism for these anti-inflammatory actions is likely to involve
upregulation of AMPK. Future studies will further explore the
specific roles of AMPK and its upstream and downstream signaling in
mediating the development of testicular interstitial cell
inflammation and examine the effect of roscovitine on these
pathways.
Acknowledgements
This study was supported a grant from the National
Natural Science Foundation of China (no. 81370699).
References
|
1
|
Krystof V and Uldrijan S: Cyclin-dependent
kinase inhibitors as anticancer drugs. Curr Drug Targets.
11:291–302. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lapenna S and Giordano A: Cell cycle
kinases as therapeutic targets for cancer. Nat Rev Drug Discov.
8:547–566. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Meijer L, Borgne A, Mulner O, Chong JP,
Blow JJ, Inagaki N, Inagaki M, Delcros JG and Moulinoux JP:
Biochemical and cellular effects of roscovitine, a potent and
selective inhibitor of the cyclin-dependent kinases cdc2, cdk2 and
cdk5. Eur J Biochem. 243:527–536. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Payton M, Chung G, Yakowec P, Wong A,
Powers D, Xiong L, Zhang N, Leal J, Bush TL, Santora V, et al:
Discovery and evaluation of dual CDK1 and CDK2 inhibitors. Cancer
Res. 66:4299–4308. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dey A, Wong ET, Cheok CF, Tergaonkar V and
Lane DP: R-Roscovitine simultaneously targets both the p53 and
NF-kappaB pathways and causes potentiation of apoptosis:
Implications in cancer therapy. Cell Death Differ. 15:263–273.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rossi AG, Sawatzky DA, Walker A, Ward C,
Sheldrake TA, Riley NA, Caldicott A, Martinez-Losa M, Walker TR,
Duffin R, et al: Cyclin-dependent kinase inhibitors enhance the
resolution of inflammation by promoting inflammatory cell
apoptosis. Nat Med. 12:1056–1064. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Leitch AE, Riley NA, Sheldrake TA, Festa
M, Fox S, Duffin R, Haslett C and Rossi AG: The cyclin-dependent
kinase inhibitor R-roscovitine down-regulates Mcl-1 to override
pro-inflammatory signalling and drive neutrophil apoptosis. Eur J
Immunol. 40:1127–1138. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim SM, Yun MR, Hong YK, Solca F, Kim JH,
Kim HJ and Cho BC: Glycolysis inhibition sensitizes non-small cell
lung cancer with T790M mutation to irreversible EGFR inhibitors via
translational suppression of Mcl-1 by AMPK activation. Mol Cancer
Ther. 12:2145–2156. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yue W, Zheng X, Lin Y, Yang CS, Xu Q,
Carpizo D, Huang H, DiPaola RS and Tan XL: Metformin combined with
aspirin significantly inhibits pancreatic cancer cell growth in
vitro and in vivo by suppressing anti-apoptotic proteins Mcl-1 and
Bcl-2. Oncotarget. 6:21208–21224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang ST, Huang SW, Kao JK, Liang SM, Chen
YJ, Chen YY, Wu CY and Shieh JJ: Imiquimod-induced AMPK activation
causes translation attenuation and apoptosis but not autophagy. J
Dermatol Sci. 78:108–116. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nunes AK, Rapôso C, Rocha SW, Barbosa KP,
Luna RL, da Cruz-Höfling MA and Peixoto CA: Involvement of AMPK,
IKβα-NFκB and eNOS in the sildenafil anti-inflammatory mechanism in
a demyelination model. Brain Res. 1627:119–133. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liong S and Lappas M: Activation of AMPK
improves inflammation and insulin resistance in adipose tissue and
skeletal muscle from pregnant women. J Physiol Biochem. 71:703–717.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hoogendijk AJ, Diks SH, van der Poll T,
Peppelenbosch MP and Wieland CW: Kinase activity profiling of
pneumococcal pneumonia. PLoS One. 6:e185192011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hoogendijk AJ, Pinhancos SS, van der Poll
T and Wieland CW: AMP-activated protein kinase activation by
5-aminoimidazole-4-carbox-amide-1-β-D-ribofuranoside (AICAR)
reduces lipoteichoic acid-induced lung inflammation. J Biol Chem.
288:7047–7052. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hong CY, Park JH, Ahn RS, Im SY, Choi HS,
Soh J, Mellon SH and Lee K: Molecular mechanism of suppression of
testicular steroidogenesis by proinflammatory cytokine tumor
necrosis factor alpha. Mol Cell Biol. 24:2593–2604. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dobashi M, Fujisawa M, Yamazaki T, Okuda
Y, Kanzaki M, Tatsumi N, Tsuji T, Okada H and Kamidono S:
Inhibition of steroidogenesis in Leydig cells by exogenous nitric
oxide occurs independently of steroidogenic acute regulatory
protein (star) mRNA. Arch Androl. 47:203–209. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liew SH, Meachem SJ and Hedger MP: A
stereological analysis of the response of spermatogenesis to an
acute inflammatory episode in adult rats. J Androl. 28:176–185.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tiancheng X and Yunfei X: Research
progress in separation and culture of mice leydig cells. J Clin
Pathol Res. 35:569–572. 2015.
|
|
19
|
Wu L, Xu B, Fan W, Zhu X, Wang G and Zhang
A: Adiponectin protects Leydig cells against proinflammatory
cytokines by suppressing the nuclear factor-κB signaling Pathway.
FEBS J. 280:3920–3927. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shchelochkov AM, Nefedova IF, Chernova SN
and Vartanova OV: The regional indicators of male fertility in the
Samara oblast and causative factors of their alterations. Klin Lab
Diagn. 25–29. 2012.(In Russian). PubMed/NCBI
|
|
22
|
Carlsen E, Andersson AM, Petersen JH and
Skakkebaek NE: History of febrile illness and variation in semen
quality. Hum Reprod. 18:2089–2092. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qureshi ST, Larivière L, Leveque G,
Clermont S, Moore KJ, Gros P and Malo D: Endotoxin-tolerant mice
have mutations in Toll-like receptor 4 (Tlr4). J Exp Med.
189:615–625. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Parvinen M, Söder O, Mali P, Fröysa B and
Ritzén M: In vitro stimulation of stage-specific deoxyribonucleic
acid synthesis in rat seminiferous tubule segments by interleukin-1
alpha. Endocrinology. 129:1614–1620. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Meijer L and Raymond E: Roscovitine and
other purines as kinase inhibitors. From starfish oocytes to
clinical trials. Acc Chem Res. 36:417–425. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cicenas J, Kalyan K, Sorokinas A,
Stankunas E, Levy J, Meskinyte I, Stankevicius V, Kaupinis A and
Valius M: Roscovitine in cancer and other diseases. Ann Transl Med.
3:1352015.PubMed/NCBI
|
|
27
|
Kolodziej M, Goetz C, Di Fazio P,
Montalbano R, Ocker M, Strik H and Quint K: Roscovitine has
anti-proliferative and pro-apoptotic effects on glioblastoma cell
lines: A pilot study. Oncol Rep. 34:1549–1556. 2015.PubMed/NCBI
|
|
28
|
Arisan ED, Akkoç Y, Akyüz KG, Kerman EM,
Obakan P, Çoker-Gürkan A and Palavan Ünsal N: Polyamines modulate
the roscovitine-induced cell death switch decision autophagy vs.
apoptosis in MCF-7 and MDA-MB-231 breast cancer cells. Mol Med Rep.
11:4532–4540. 2015.PubMed/NCBI
|
|
29
|
El-Alfy M, Luu-The V, Huang XF, Berger L,
Labrie F and Pelletier G: Localization if type 5
17beta-hydroxysteroid dehydrogenase, 3beta-hydroxysteroid
dehydrogenase, and androgen receptor in the human prostate by in
situ hybridization and immunocytochemistry. Endocrinology.
140:1481–1491. 1999. View Article : Google Scholar
|
|
30
|
Gingras S, Moriggl R, Groner B and Simard
J: Induction of 3beta-hydroxysteroid dehydrogenase/delta5-delta4
isomerase type 1 gene transcription in human breast cancer cell
lines and in normal mammary epithelial cells by interleukin-4 and
interleukin-13. Mol Endocrinol. 13:66–81. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gingras S and Simard J: Induction of
3beta-hydroxysteroid dehydrogenase/isomerase type 1expression by
interleukin-4 in human normal prostate epithelial cells,
immortalized keratinocytes, colon, and cervix cancer cell lines.
Endocrinology. 140:4573–4584. 1999. View Article : Google Scholar
|
|
32
|
Xue LY, Li J, Wang GX, Xiu HM and Sun HC:
The functional change of the rats testis in the condition of
undifferential orchitis induced by lipopolysaccharide. Reproduction
& Contraception. 26:77–80. 2006.
|
|
33
|
Chen L, Lai YS and Ji H: Effect of
COX/5-LOX dual inhibitor ZLJ-6 on the expression of LPS-induced
inflammation factors in macrophage cells. Journal of China
Pharmaceutical University. 43:70–74. 2012.
|
|
34
|
Yang KY, Gu JL, Yin DM and Shen Q: Effect
of Curcumin on iNOS Expression in LPS-activated Microglia Cells and
Ant-i oxidation. Chinese Journal of Biochemistry and Molecular
Biology. 23:938–945. 2007.
|
|
35
|
Ewart MA, Kohlhaas CF and Salt IP:
Inhibition of tumor necrosis factor alpha-stimulated monocyte
adhesion to human aortic endothelial cells by AMP-activated protein
kinase. Arterioscler Thromb Vasc Biol. 28:2255–2257. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Giri S, Nath N, Smith B, Viollet B, Singh
AK and Singh I:
5-Aminoimidazole-4-carboxamide-1-beta-4-ribofuranoside inhibits
proinflammatory response in glial cells: A possible role of
AMP-activated protein kinase. J Neurosci. 24:479–487. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yeung F, Hoberg JE, Ramsey CS, Keller MD,
Jones DR, Frye RA and Mayo MW: Modulation of NF-kappaB-dependent
transcription and cell survival by the SIRT1 deacetylase. EMBO J.
23:2369–2380. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ak P and Levine AJ: P53 and NF-κB:
Different strategies for responding to stress lead to a functional
antagonism. FASEB J. 24:3643–3652. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Komarova EA, Krivokrysenko V, Wang K,
Neznanov N, Chernov MV, Komarov PG, Brennan ML, Golovkina TV,
Rokhlin OW, Kuprash DV, et al: p53 is a suppressor of inflammatory
response in mice. FASEB J. 19:1030–1032. 2005.PubMed/NCBI
|
|
40
|
Yu M, Zhang LY, Qiao PH and Chang B:
Testicular oxidative stress and downregulation of CYP17a1 induced
by di(n-butyl) phthalate inhibit synthesis of testosterone. Wei
Sheng Yan Jiu. 44:364–370. 2015.PubMed/NCBI
|