Introduction
Adaptive immune responses are classified into two
categories: T helper (Th) 1-type response (or cellular immune
response) and Th2 response (or humoral immune response). These
subsets of helper cluster of differentiation (CD)4+ T
cells differ in their cytokine secretion patterns and effector
functions, and counter-regulate one another (1). The majority of allergic diseases,
including atopic dermatitis (AD), are considered to be due to the
abnormal activation of Th2 cells, namely the polarization of
allergen-specific T-cell memory towards the Th2 instead of the Th1
immune response (2). Interleukin
(IL)-4, which is a representative cytokine of Th2 cells, induces
immunoglobulin E (IgE) isotype switching, augments IgE production
in B cells, and promotes Th2 differentiation (2). Interferon-γ (IFN-γ), a representative
cytokine of Th1 cells, is known to suppress the development of Th2
cells and promote Th1 differentiation (3,4).
Considering the evidence suggesting that the Th1 and Th2 types of
immune responses are reciprocally regulated in vivo
(5), modulation of the Th1/Th2
balance, namely shifting the balance from Th2 to Th1 dominance, may
be an effective strategy for the treatment of allergic diseases
associated with Th2 cells (6–9).
AD is a highly prevalent skin inflammatory disease
characterized by allergic signs and symptoms of the skin, such as
redness, edema, warmth, and pruritus (10–12). AD
is a multifactorial disease with genetic and environmental
components, and its pathogenesis has not yet been fully defined
(13). AD is associated with
elevated IgE and IL-4 production (13,14),
indicating the predominance of the Th2 response over the Th1
response. The current therapeutic management of AD aims to relieve
recurring skin inflammation and prevent flare-ups. Topical steroids
and immunosuppressive agents have typically been the standard
treatment for severe cases of AD (15,16).
However, these agents are often associated with severe adverse
effects and are not sufficiently effective in a substantial number
of patients with AD (15,16). There is therefore a need to develop
novel effective therapies for the treatment of AD. Notably, it has
been reported that certain strains of lactobacilli may offer
clinical benefits to patients with AD or other allergic diseases
through the enhancement of Th1 immunity and suppression of Th2
immunity (17–19).
Lactobacilli are used widely in industrial
and traditional fermentative food processes, and have a long
history of safe use (20,21). It has been reported that
Lactobacilli in a variety of food products may potentially
provide health benefits (22–24).
Certain Lactobacillus strains may potentially modulate the
immune response via the production of interleukin (IL)-12,
promoting the Th1/Th2 balance towards a Th1-dominant state in
systemic immunity (25–34). This effect may be of use in the
prevention and treatment of AD.
Jeotgal is a traditional Korean fermented
food, which is made with various salted seafood, such as shrimp,
oysters, shellfish, fish, and fish eggs, and is primarily used as a
condiment in the production of other fermented foods (35–41). It
has previously been reported that many Lactobacillus strains
are involved in the fermentation of Jeotgal, suggesting that
it is a good source of various Lactobacillus strains
(35–41).
The aim of present study was to examine the in
vivo anti-AD effects of two novel strains of
Lactobacillus isolated from Jeotgal using a 2,
4-dinitrofluorobenzene (DNFB)-induced AD-like skin inflammation
model. To explain a putative mechanism of action of the isolated
Lactobacilli, the ex vivo effects on the secretion of
cytokines by CD4+ T cells from lymph nodes isolated from
mice that were sensitized and challenged with DNFB were examined,
as well as the in vitro effects on IL-12 production in
macrophages.
Materials and methods
Materials
MRS Broth was obtained from BD Biosciences (Franklin
Lakes, NJ, USA). RPMI-1640, fetal bovine serum (FBS),
penicillin/streptomycin, and DNFB were purchased from Sigma-Aldrich
(Merck KGaA, Darmstadt, Germany). All other reagents were obtained
from Sigma-Aldrich (Merck KGaA).
Lactobacillus strains
The authors of the present study searched for novel
Lactobacillus strains that may be beneficial for AD
treatment, and isolated novel Lactobacillus strains with
conjugated linoleic acid (CLA)-producing ability, using a linoleic
acid tolerance method, as previously described (42). The Lactobacillus strains used
in the present study were isolated from five types of the Korean
traditional fermented food Jeotgal obtained from
Shinan-Suhyup Market (Shinan, Korea), of which the original sources
were baby octopus (Octopus ocellatus), tripe of pollack
(Pollachius pollachius), small octopus (Octopus
minor), scallop (Patinopecten yessoensis) and oyster
(Crassostrea gigas). The lactic acid bacteria that were able
to produce high-level CLA were selected using the linoleic acid
tolerance method (42) as shown in
Table I and identified by 16S rRNA
gene sequence analysis (43) as
Lactobacillus plantarum (JBCC105645 and JBCC105683) with
99–100% similarity. JBCC105683 exhibited the highest CLA producing
ability (748.8 µg/ml), and JBCC105645 produced 227.4 µg/ml
(Table I). Following cultivation in
MRS broth at 37°C for 24 h, the cells of both strains were
harvested, washed with sterile distilled water and lyophilized in
lyoprotectant consisting of 750 ml horse serum and 250 ml distilled
water supplemented with 7.5% sucrose (both from Sigma-Aldrich;
Merck KGaA), 0.63% nutrient broth (Sigma-Aldrich; Merck KGaA) and 1
mg/ml cysteine HCl. The cells were killed by heating at 105°C for
15 min, and the resultant heat-killed cells were lyophilized and
then suspended in PBS to be used in the following experiments.
 | Table I.Production of total CLA by
lactobacilli isolated from Jeotgal. |
Table I.
Production of total CLA by
lactobacilli isolated from Jeotgal.
| Strain | Linoleic acid
(µg/ml) | Total CLA
(µg/ml) |
|---|
| Lactobacillus
plantarum |
|
|
| (JBCC105683) |
100 | 334.475±1.797 |
|
|
200 | 553.118±5.635 |
|
|
600 | 748.807±1.919 |
|
| 1,000 | 412.929±1.919 |
| Lactobacillus
plantarum |
|
|
| (JBCC105645) |
100 | 227.403±13.067 |
|
|
200 | 185.449±5.513 |
|
|
600 | 192.061±0.000 |
|
| 1,000 | 167.230±13.067 |
Mice
A total of 100 female 5-week-old BALB/c mice
weighing 15–20 g were purchased from the Da-Mul Experimental Animal
Center (Daejeon, Korea) and were maintained conventionally in
plastic cages at a temperature of ~22°C and humidity of ~50% under
a 12-h light/dark cycle. The mice were provided with a standard
diet and access to autoclaved water ad libitum throughout
the experimental period. For experiments, mice were divided into 4
groups (n=6 in each group) as follows: Normal control group,
unchallenged with DNFB and administrated with PBS; DNFB model,
challenged with DNFB and administrated with PBS; JBCC195645-treated
group, challenged with DNFB and administrated with JBCC195645;
JBCC105683-treated group, challenged with DNFB and administrated
with JBCC105683. The mice were anesthetized with 2% isoflurane
(Sigma-Aldrich; Merck KGaA) in 98% oxygen before sacrifice by
decapitation. The present study was performed according to the
guidelines laid out by Ethics Committee for Animal Experiments of
Wonkwang University (Iksan, Korea) and was approved by the Ethics
Committee (WKU15-17).
Induction of AD-like skin
inflammation
AD-like skin inflammation was induced by repeated
treatment with DNFB solution on the surfaces of the right ear and
shaved abdomen in BALB/c mice. After administration of anesthesia
(1.2% isoflurane), shaved abdomen skin was coated with a total of
100 µl 0.5% DNFB in acetone/olive oil (4:1) once a day for 5 days.
Following this, mouse ears were coated with 25 µl 0.2% DNFB (25 µl)
for 2 days. Normal control mice were treated with acetone/olive oil
(4:1) in the absence of DNFB. Ear thickness was measured with a
spring-loaded micrometer (Oditest; Swiss Precision Instruments,
Inc., Garden Grove, CA, USA) prior to DNFB and 24 and 48 h
following the DNFB challenge.
Oral administration of
lactobacilli
A total of 200 µl PBS containing heat-killed
JBCC105645 or JBCC105683 strains [1×109 colony-forming
units (CFU)/ml] was orally administrated once a day for 7 days
before DNFB challenge. DNFB model mice were orally treated with 200
µl PBS in the absence of lactobacilli.
Histological study
Mice were anesthetized with 2% isoflurane
(Sigma-Aldrich; Merck KGaA) in 98% oxygen and sacrificed by
decapitation. Treated ears were harvested 48 h after the final
application of DNFB or acetone/olive oil. The tissue was fixed for
10 min at room temperature in a 10% neutral formalin solution and
embedded in paraffin wax by standard procedures (44). The sections (4–5 µm) were stained
with hematoxylin and eosin and observed using a light microscope.
Digital photomicrograph images were captured from five
representative areas at a fixed magnification of ×100.
Analysis of total serum IgE
production
Blood samples (1 ml) were collected from the heart
following completion of treatment, and the sera were separated by
centrifugation at 3,000 × g for 5 min at 4°C. Concentrations of
total serum IgE were measured using a Mouse IgE ELISA MAX™ Deluxe
kit (cat. no. 432406) purchased from BioLegend. Inc. (San Diego,
CA, USA), according to the manufacturer's protocols.
Isolation of CD4+ T cells
and analysis of IL-4 and IFN-γ production
On day 11 following the end of treatment, all mice
were sacrificed and their lymph nodes were isolated as previously
described (45). T cells were
isolated from the lymph nodes and CD4+ T cells were
purified using a CD4+ T cell isolation kit purchased
from Miltenyi Biotec GmbH (Bergisch Gladbach, Germany), according
to the manufacturer's protocol. Isolated CD4+ T cells
(1.5×105) were cultured for 2 h at 37°C in 96-well
flat-bottom culture plates in RPMI-1640 medium with 10% FBS, 50 µM
β-mercaptoethanol and 2 mM glutamine (both from Sigma-Aldrich;
Merck KGaA), and then stimulated for 24 h at 37°C with immobilized
anti-CD3 antibody (5 µg/ml; cat. no. 554829) and the soluble form
of purified anti-CD28 antibody (2 µg/ml; cat. no. 553294) (both
from BD Pharmingen, San Diego, CA, USA). Levels of IL-4 and IFN-γ
were quantified using IL-4 ELISA (cat. no. 431103) and IFN-γ ELISA
kits (cat. no. 430807) (both from BioLegend, Inc.), respectively,
following T cell activation.
Macrophage culture and analysis of
IL-12 production
RAW264.7 macrophages were obtained from American
Type Culture Collection (cat. no. ATCC® TIB-71™; ATCC,
Manassas, VA, USA). Cells were maintained for 18 h at 37°C in
RPMI-1640 medium supplemented with 10% heat-inactivated FBS, 100
U/ml penicillin, and 100 µg/ml streptomycin. Macrophages
(1.5×105) were cultured for 2 h at 37°C in 96-well
flat-bottom culture plates in RPMI-1640 medium with 10%
heat-inactivated FBS, and then stimulated for 48 h at 37°C with
5×107 CFU/ml heat-killed JBCC105645 or JBCC105683. The
production of IL-12 following stimulation was quantified using a
mouse IL-12 ELISA kit (cat. no. 433606; BioLegend Inc.).
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical differences between the means of treated and control
groups were analyzed using one-way analysis of variance and
Fisher's protected least significant difference and GraphPad Prism
software version 3.03 (GraphPad Software, Inc., La Jolla, CA, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of JBCC105645 and JBCC105683
on DNFB-induced ear swelling response
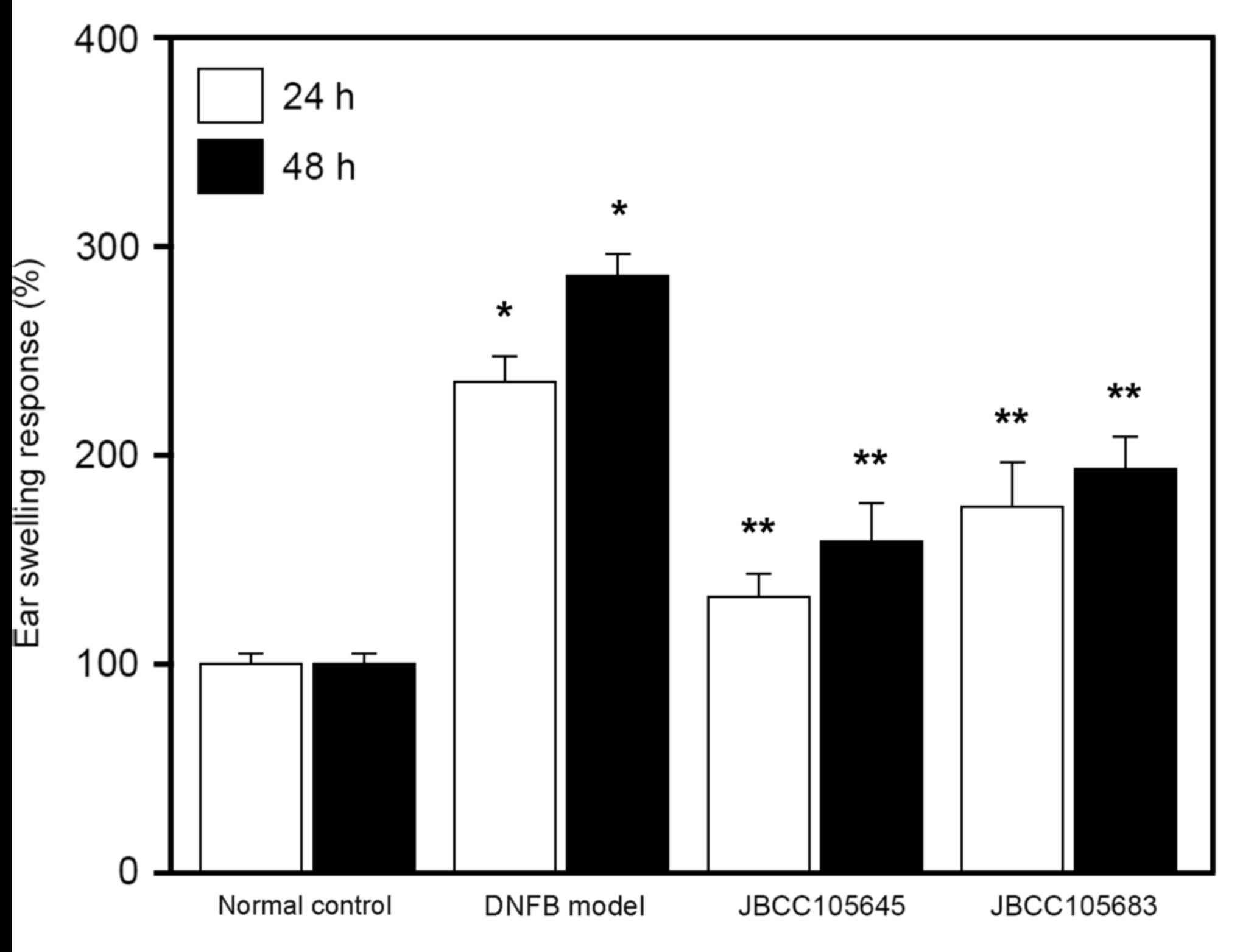
DNFB was used to induce a model of murine contact
hypersensitivity. In the skin following exposure to DNFB, murine
contact hypersensitivity is able to generate AD-like lesions and
induce Th2-type immune responses, and is therefore often used to
study the effects of drugs on AD pathogenesis (46,47). In
the present study, repeated applications of DNFB were used to
induce AD-like lesions in BALB/c mice. Following epidermal
application of DNFB to the skin of mouse ears (DNFB challenge),
significant ear swelling was induced in the DNFB model group
compared with the normal control group (P<0.05; Fig. 1). This DNFB-induced ear swelling was
significantly reduced in both the JBCC105645 and JBCC105683-treated
groups at 24 and 48 h following treatment (P<0.05; Fig. 1); JBCC105645 was markedly more
effective than JBCC105683 in reducing DNFB-induced edema.
Effects of JBCC105645 and JBCC105683
on histopathological findings of DNFB-challenged ear skin
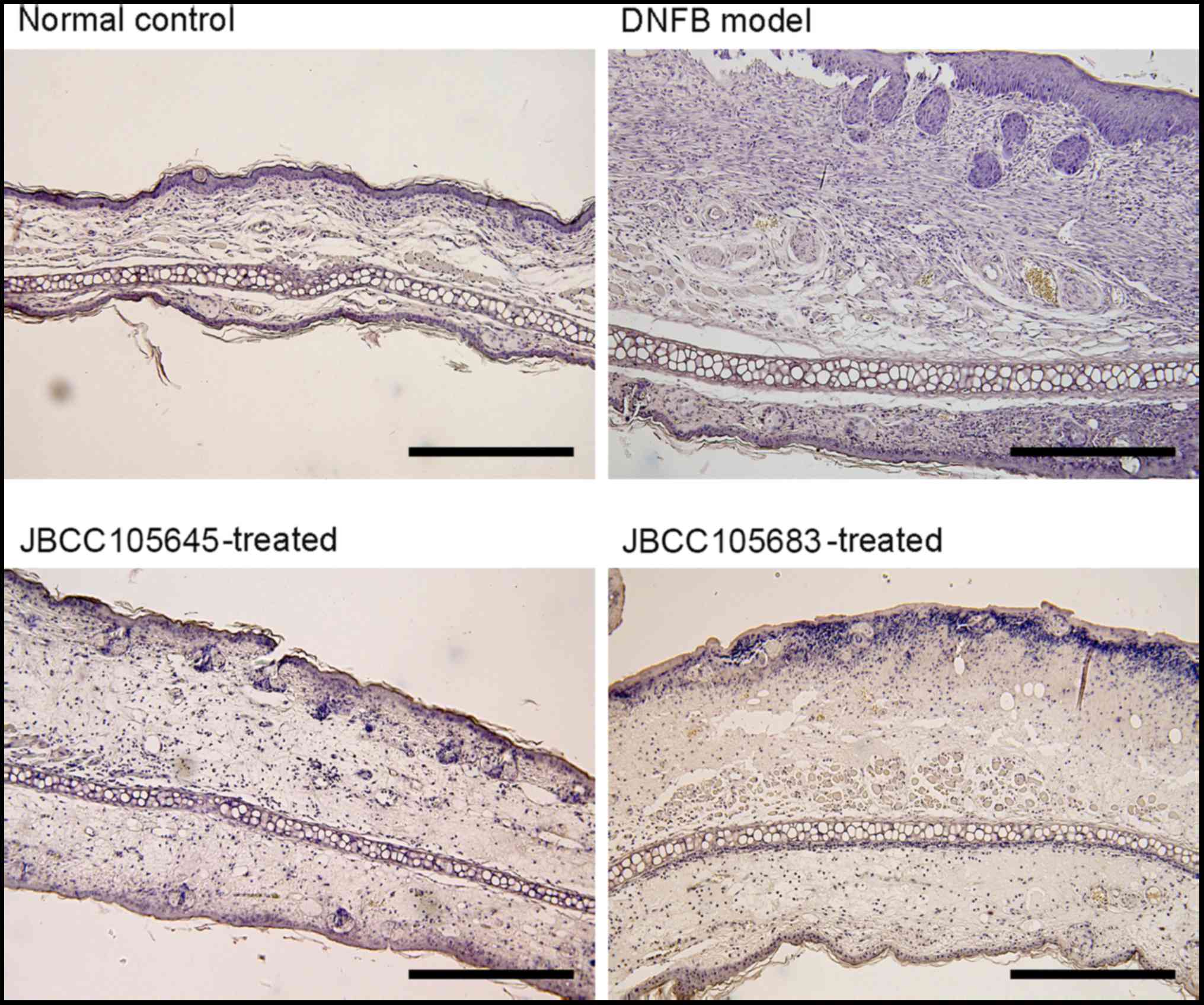
Histological analysis revealed spongiosis in the ear
skin of mice in the DNFB model group (Fig. 2). Additionally, marked infiltration
of inflammatory cells in the epidermis and dermis was also observed
in DNFB model group (Fig. 2).
DNFB-induced pathophysiological parameters, such as spongiosis and
infiltration of inflammatory cells, were markedly reduced following
oral treatment with JBCC105645 or JBCC105683 (Fig. 2).
Effects of oral administration of
JBCC105645 and JBCC105683 on total serum IgE levels
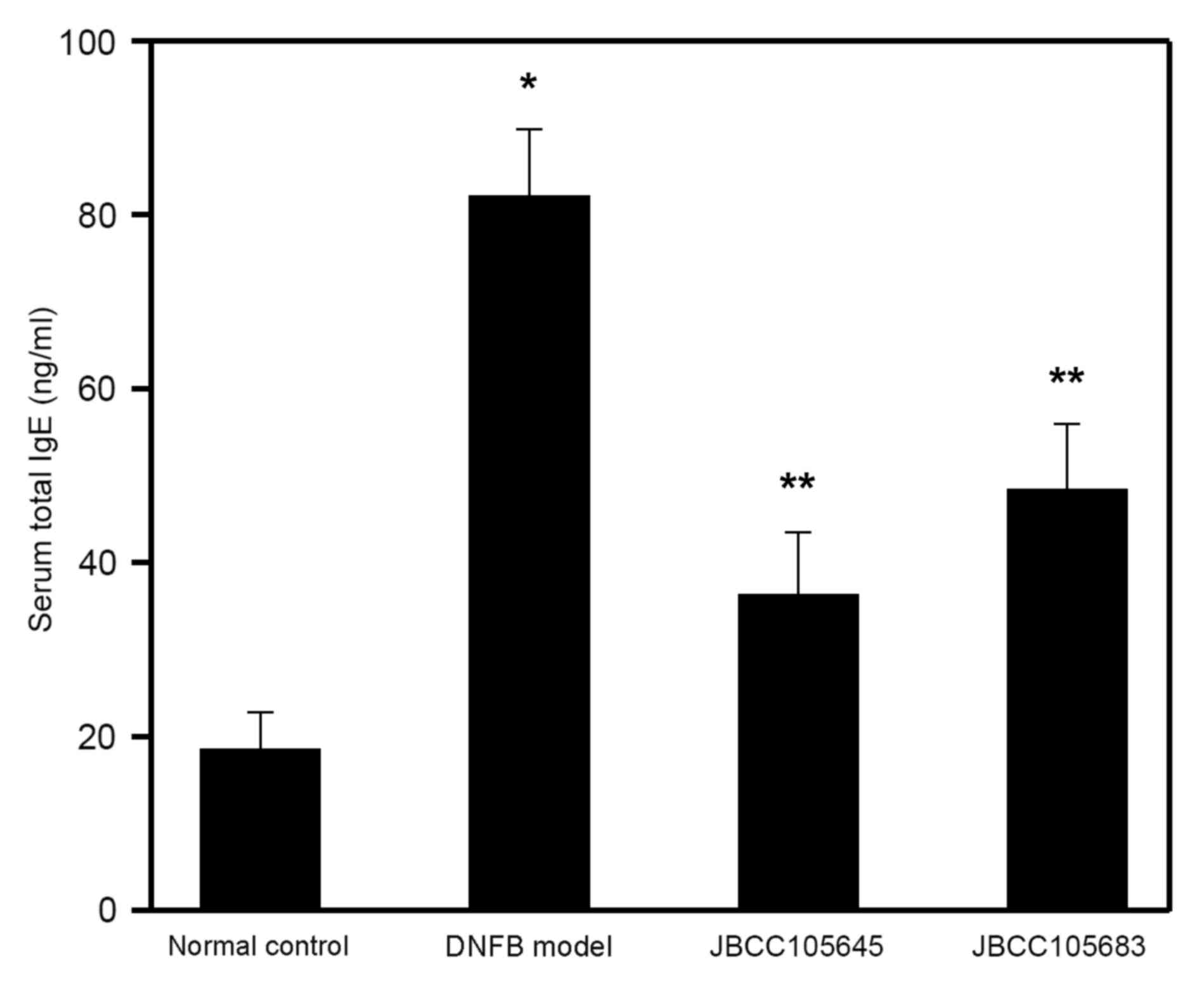
In addition to the clinical features, the levels of
total serum IgE, which is one of the most common indicators of AD
(13,14), were also assessed to characterize the
immunological response during disease progression. Following the
completion of DNFB treatment, serum samples were obtained, and
total serum IgE levels were determined by ELISA. Total serum IgE
levels were significantly increased by DNFB challenge compared with
the normal control group (P<0.05; Fig. 3). The DNFB-induced elevation in serum
IgE levels was significantly decreased by oral treatment with
JBCC105645 or JBCC105683 (P<0.05; Fig. 3); JBCC105645 was markedly more
effective than JBCC105683 in serum IgE inhibition.
Effects of JBCC105645 and JBCC105683
on IL-4/IFN-γ production by ex vivo CD4+ T cells
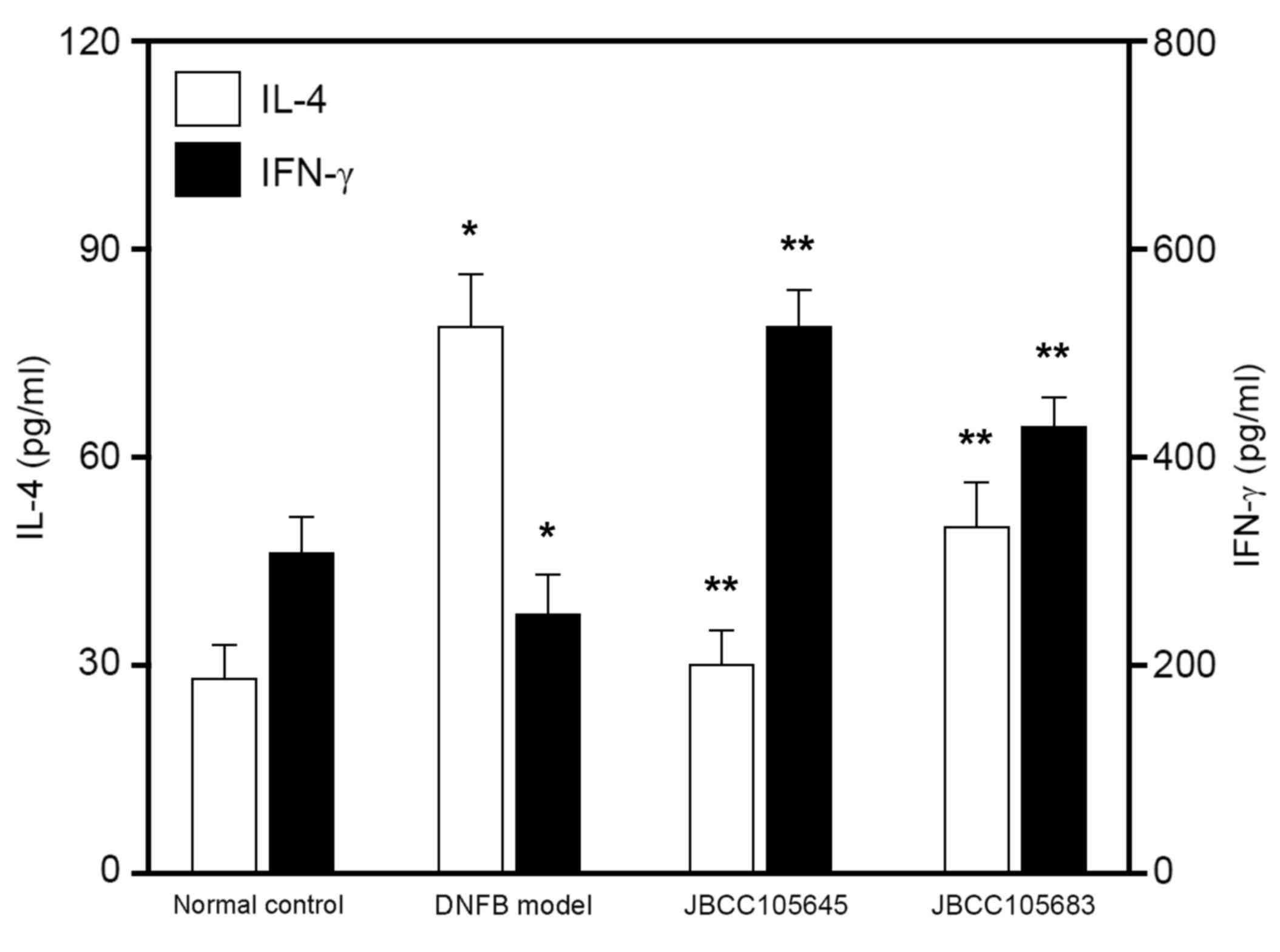
To evaluate whether oral administration of
lactobacilli would affect IL-4 and IFN-γ production,
CD4+ T cells were isolated from the lymph nodes of mice
following exposure to DNFB and re-stimulated for 48 h with anti-CD3
and anti-CD28 antibodies. IL-4 and IFN-γ concentrations in the
culture supernatant were assessed using ELISA. Both IL-4 and IFN-γ
production was measured in CD4+ T cells. IL-4 production
in ex vivo re-stimulated CD4+ T cells was
significantly increased in the DNFB model compared with the
control, while IFN-γ production was significantly decreased (both
P<0.05; Fig. 4). IL-4 production
was significantly attenuated by JBCC105645 and JBCC105683 compared
with the DNFB model (P<0.05; Fig.
4), while IFN-γ production was significantly augmented by these
treatments (P<0.05; Fig. 4).
JBCC105645 was markedly more effective in suppressing IL-4
production and enhancing IFN-γ production than JBCC105683.
Effects of JBCC105645 and JBCC105683
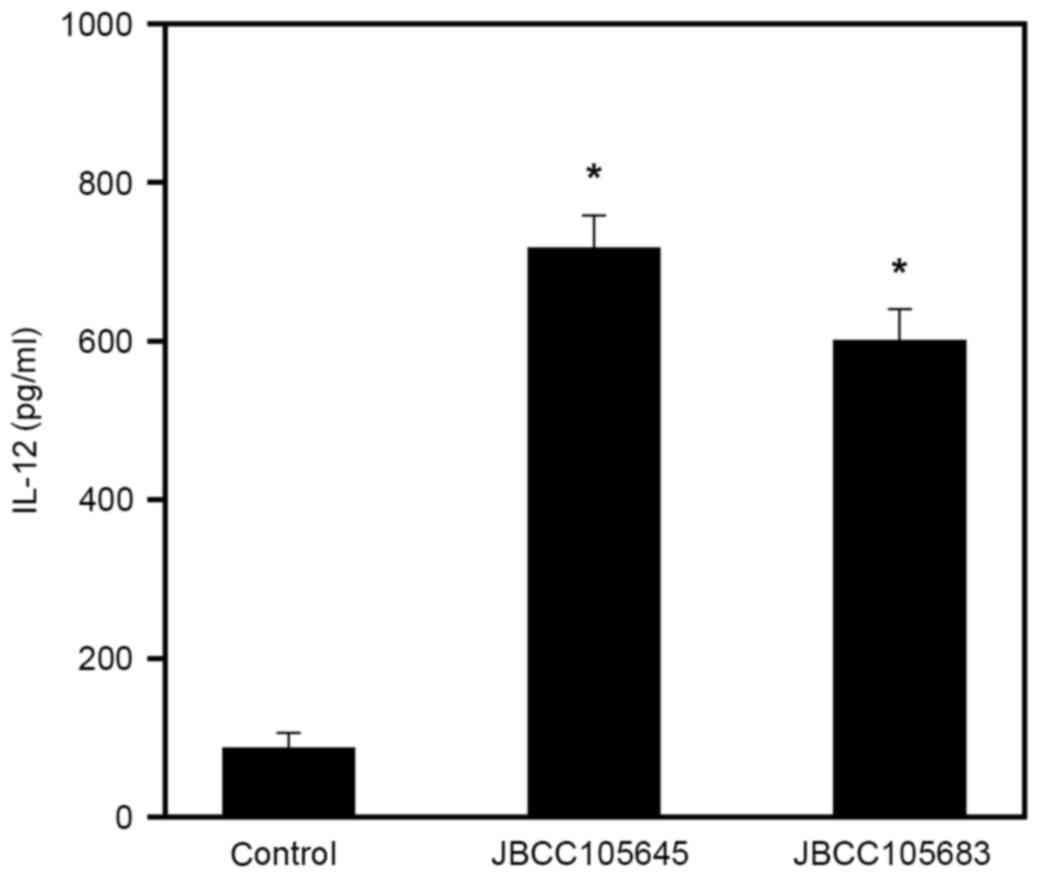
on IL-12 production by in vitro macrophages
In the present study, in vitro cell culture
studies were performed by testing whether lactobacilli stimulated
macrophages to produce IL-12, which is a Th1
differentiation-stimulatory cytokine (48). RAW264.7 macrophages were stimulated
with heat-killed lactobacilli for 24 h, and levels of IL-12 in
supernatants were subsequently measured using ELISA. Macrophages
stimulated with JBCC105645 and JBCC105683 had significantly
increased levels of IL-12 production compared with untreated cells
(P<0.05; Fig. 5). At a given
concentration (5×107 CFU/ml), JBCC105645 was markedly
more effective in induction of IL-12 production than
JBCC105683.
Discussion
It has previously been reported that intestinal
microflora serve an important role in Th1/Th2 balance (29,49),
which is the primary mechanism associated with allergic diseases.
Furthermore, human epidemiological data suggest that altered
microbial exposure during childhood influences the outcome of
allergic sensitization (50–52); therefore, the role of probiotics in
allergic disease has been highlighted. There are a substantial
number of reports that demonstrate the beneficial effects of
probiotics in the management of AD and other allergic responses
(53–55). A large number of organisms in
probiotics have previously been studied, and certain strains of
lactobacilli and bifidobacteria are now believed to have
therapeutic effects on the extent and severity of allergic
diseases, including AD (25–34,56). In
the present study, it was addressed whether oral administration of
lactobacilli (JBCC105645 and JBCC105683 strains) isolated from
Jeotgal, a Korean traditional fermented food (35–41), was
able to reduce the severity of AD-like skin lesions induced by DNFB
in BALB/c mice. The results demonstrated that two lactobacillus
strains significantly suppressed AD-like skin inflammation with
varying effectiveness. JBCC105645 was more potent than JBCC105683,
thus suggesting that the beneficial effects of the lactobacilli on
the extent and severity of AD may be strain-specific.
The upregulation of total serum IgE is a hallmark of
AD (13,14). Elevated serum IgE levels are known to
cause both acute and chronic skin inflammation (13,14), and
DNFB exposure increases serum IgE levels in several animal models
(46,47). In line with this, a significant
increase in total serum IgE levels was also observed in the present
AD mouse model. However, oral treatment with the lactobacilli
isolated from Jeotgal significantly reduced DNFB-induced IgE
levels. Notably, the potency for IgE inhibition by the lactobacilli
was similar to that for their suppression of AD-like skin
inflammation.
The balance of the Th2 cytokine IL-4 and the Th1
cytokine IFN-γ has previously been reported to be critical for
regulating IgE production (2–5). IL-4
produced by Th2 cells induces a switch to IgE production by
differentiating B cells, resulting in an increase in IgE
production, whereas IFN-γ produced by Th1 cells is able to inhibit
that switch, consequently preventing IgE production (2–5). In this
context, it is possible that the inhibition of serum IgE levels by
JBCC105645 and JBCC105683 may result from downregulation of IL-4
production and/or upregulation of IFN-γ production. To test this
possibility, it was investigated whether oral administration of
lactobacillus was able to influence the balance of IL-4/IFN-γ
production in ex vivo re-stimulated CD4+ T cells
obtained from lymph nodes of DNFB-challenged mice. Oral
administration of the lactobacilli induced a significant decrease
in IL-4 production with concomitant increase in IFN-γ production.
Interestingly, JBCC105645 was more effective in IL-4 inhibition and
IFN-γ enhancement than JBCC105683, which was similar to their
effects on IgE inhibition. These results suggest that oral
administration of lactobacilli may reduce the levels of serum IgE,
at least in part, via modulation of IL-4/IFN-γ production in
DNFB-challenged mice. The results of the present study are also in
agreement with previous reports that found that other lactobacillus
strains are able to modulate IL-4/IFN-γ production in re-stimulated
CD4+ T cells (28,57).
At present, the mechanism(s) by which the
lactobacilli modulate IL-4/IFN-γ production remains unclear. It has
been reported that IL-12, which is primarily produced by
antigen-presenting cells such as macrophages and dendritic cells,
typically induces differentiation of IFN-γ-producing Th1 cells,
thereby preventing differentiation of IL-4-producing Th2 cells
(25,28). Furthermore, it has been reported that
various lactobacillus strains are able to induce IL-12 production
in mouse macrophages and stimulate IFN-γ production in mouse
splenocytes (25); which prompted
the examination of whether JBCC105645 and JBCC105683 were also able
to stimulate macrophages to produce IL-12 in the present study. The
results demonstrated that the lactobacilli directly stimulated
macrophages in vitro to induce IL-12 production, and
JBCC105645 was more effective in IL-12 production than JBCC105683.
This suggests that the enhancement of IFN-γ production by
lactobacilli may be a result of the stimulating effects of
JBCC105645 and JBCC105683 on IL-12 release from the
antigen-presenting cell macrophages. The mechanism(s) by which the
lactobacilli reduce IL-4 production in re-stimulated
CD4+ T cells has yet to be elucidated. IFN-γ is able to
prevent IL-4 production via the inhibition of Th2 differentiation
(3–5), and so it may be assumed that inhibition
of IL-4 production is due to enhanced IFN-γ production. Future
studies should aim to produce a detailed study of the relevant
mechanisms.
In conclusion, the results of the present study
demonstrate that oral treatment with JBCC105645 and JBCC105683
strains isolated from Jeotgal inhibits AD-like skin lesions
and reduces serum IgE levels in DNFB-treated mice. The cytokine
production pattern of ex vivo CD4+ T cells
demonstrated that administration of these lactobacillus strains
induced a significant decrease in levels of the Th2-related
cytokine IL-4 with a concomitant increase in the Th1-related
cytokine IFN-γ. Additionally, lactobacilli stimulate macrophages to
produce IL-12 in vitro. These results suggest that the
lactobacillus strains isolated from Jeotgal are capable of
reducing the severity of AD-like skin lesions, presumably by
switching the immune response from Th2 type toward Th1 type via the
activation of macrophages. Future studies should be directed
towards identifying which components of the lactobacilli are
responsible for their immunomodulatory effects.
Acknowledgements
The present study was supported by the High
Value-Added Food Technology Development Program, (Korea Institute
of Planning and Evaluation for Technology in Food, Agriculture,
Forestry and Fisheries, and the Ministry for Food, Agriculture,
Forestry, and Fisheries of Republic of Korea; project no.
313037-3).
References
|
1
|
Yacoub MR, Colombo G, Marcucci F, Caminati
M, Sensi L, Di Cara G, Frati F and Incorvaia C: Effects of
sublingual immunotherapy on allergic inflammation: An update.
Inflamm Allergy Drug Targets. 11:285–291. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Di Lernia V: Therapeutic strategies in
extrinsic atopic dermatitis: Focus on inhibition of IL-4 as a new
pharmacological approach. Expert Opin Ther Targets. 19:87–96. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Romagnani S: Regulation of the T cell
response. Clin Exp Allergy. 36:1357–1166. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Teixeira LK, Fonseca BP, Barboza BA and
Viola JP: The role of interferon-gamma on immune and allergic
responses. Mem Inst Oswaldo Cruz. 100 Suppl 1:S137–S144. 2005.
View Article : Google Scholar
|
|
5
|
Mucida D and Cheroutre H: The many
face-lifts of CD4 T helper cells. Adv Immunol. 107:139–152. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
McFadden JP, Thyssen JP, Basketter DA,
Puangpet P and Kimber I: T helper cell 2 immune skewing in
pregnancy/early life: Chemical exposure and the development of
atopic disease and allergy. Br J Dermatol. 172:584–591. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang Y, Zhang Y, Gu W, He L and Sun B:
Th1/Th2 cell's function in immune system. Adv Exp Med Biol.
841:45–65. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nguyen TH and Casale TB: Immune modulation
for treatment of allergic disease. Immunol Rev. 242:258–271. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kumazawa Y, Takimoto H, Matsumoto T and
Kawaguchi K: Potential use of dietary natural products, especially
polyphenols, for improving type-1 allergic symptoms. Curr Pharm
Des. 20:857–863. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Werfel T and Biedermann T: Current novel
approaches in systemic therapy of atopic dermatitis: Specific
inhibition of cutaneous Th2 polarized inflammation and itch. Curr
Opin Allergy Clin Immunol. 15:446–452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Montes-Torres A, Llamas-Velasco M,
Pérez-Plaza A, Solano-López G and Sánchez-Pérez J: Biological
treatments in atopic dermatitis. J Clin Med. 4:593–613. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee J, Park CO and Lee KH: Specific
immunotherapy in atopic dermatitis. Allergy Asthma Immunol Res.
7:221–229. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu LC and Scheerens H: Targeting IgE
production in mice and humans. Curr Opin Immunol. 31:8–15. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamanaka K and Mizutani H: The role of
cytokines/chemokines in the pathogenesis of atopic dermatitis. Curr
Probl Dermatol. 41:80–92. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Svensson A, Chambers C, Gånemo A and
Mitchell SA: A systematic review of tacrolimus ointment compared
with corticosteroids in the treatment of atopic dermatitis. Curr
Med Res Opin. 27:1395–1406. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Silverberg JI: Atopic dermatitis: An
evidence-based treatment update. Am J Clin Dermatol. 15:149–164.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lomax AR and Calder PC: Probiotics, immune
function, infection and inflammation: A review of the evidence from
studies conducted in humans. Curr Pharm Des. 15:1428–11518. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ozdemir O: Various effects of different
probiotic strains in allergic disorders: An update from laboratory
and clinical data. Clin Exp Immunol. 160:295–304. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vitaliti G, Pavone P, Guglielmo F, Spataro
G and Falsaperla R: The immunomodulatory effect of probiotics
beyond atopy: An update. J Asthma. 51:320–332. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vogel RF and Ehrmann M: Genetics of
lactobacilli in food fermentations. Biotechnol Annu Rev. 2:123–150.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Derrien M and van Hylckama Vlieg JE: Fate,
activity, and impact of ingested bacteria within the human gut
microbiota. Trends Microbiol. 23:354–366. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pandey V, Berwal V, Solanki N and Malik
NS: Probiotics: Healthy bugs and nourishing elements of diet. J Int
Soc Prev Community Dent. 5:81–87. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mitsuoka T: Development of functional
foods. Biosci Microbiota Food Health. 33:117–128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kemgang TS, Kapila S, Shanmugam VP and
Kapila R: Cross-talk between probiotic lactobacilli and host immune
system. J Appl Microbiol. 117:303–319. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Won TJ, Kim B, Song DS, Lim YT, Oh ES, Lee
do I, Park ES, Min H, Park SY and Hwang KW: Modulation of Th1/Th2
balance by lactobacillus strains isolated from Kimchi via
stimulation of macrophage cell line J774A.1 in vitro. J Food Sci.
76:H55–H61. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hazebrouck S, Przybylski-Nicaise L,
Ah-Leung S, Adel-Patient K, Corthier G, Langella P and Wal JM:
Influence of the route of administration on immunomodulatory
properties of bovine beta-lactoglobulin-producing Lactobacillus
casei. Vaccine. 27:5800–5805. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chuang L, Wu KG, Pai C, Hsieh PS, Tsai JJ,
Yen JH and Lin MY: Heat-killed cells of lactobacilli skew the
immune response toward T helper 1 polarization in mouse splenocytes
and dendritic cell-treated T cells. J Agric Food Chem.
55:11080–11086. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mohamadzadeh M, Olson S, Kalina WV, Ruthel
G, Demmin GL, Warfield KL, Bavari S and Klaenhammer TR:
Lactobacilli activate human dendritic cells that skew T cells
toward T helper 1 polarization. Proc Natl Acad Sci USA. 102:pp.
2880–2885. 2005; View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cross ML, Mortensen RR, Kudsk J and Gill
HS: Dietary intake of Lactobacillus rhamnosus HNOO1 enhances
production of both Th1 and Th2 cytokines in antigen-primed mice.
Med Microbiol Immunol. 191:49–53. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Drago L, Nicola L, Iemoli E, Banfi G and
De Vecchi E: Strain-dependent release of cytokines modulated by
Lactobacillus salivarius human isolates in an in vitro model. BMC
Res Notes. 3:442010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mileti E, Matteoli G, Iliev ID and
Rescigno M: Comparison of the immunomodulatory properties of three
probiotic strains of Lactobacilli using complex culture systems:
Prediction for in vivo efficacy. PLoS One. 4:e70562009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hart AL, Lammers K, Brigidi P, Vitali B,
Rizzello F, Gionchetti P, Campieri M, Kamm MA, Knight SC and Stagg
AJ: Modulation of human dendritic cell phenotype and function by
probiotic bacteria. Gut. 53:1602–1609. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Christensen HR, Frøkiaer H and Pestka JJ:
Lactobacilli differentially modulate expression of cytokines and
maturation surface markers in murine dendritic cells. J Immunol.
168:171–178. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Miettinen M, Matikainen S, Vuopio-Varkila
J, Pirhonen J, Varkila K, Kurimoto M and Julkunen I: Lactobacilli
and streptococci induce interleukin-12 (IL-12), IL-18 and gamma
interferon production in human peripheral blood mononuclear cells.
Infect Immun. 66:6058–6062. 1998.PubMed/NCBI
|
|
35
|
Lee JM, Kim YR, Kim JK, Jeong GT, Ha JC
and Kong IS: Characterization of salt-tolerant β-glucosidase with
increased thermostability under high salinity conditions from
Bacillus sp. SJ-10 isolated from jeotgal, a traditional Korean
fermented seafood. Bioprocess Biosyst Eng. 38:pp. 1335–1346. 2015;
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Han KI, Kim YH, Hwang SG, Jung EG, Patnaik
BB, Han YS, Nam KW, Kim WJ and Han MD: Bacterial community dynamics
of salted and fermented shrimp based on denaturing gradient gel
electrophoresis. J Food Sci. 79:M2516–M2522. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jeong DW, Han S and Lee JH: Safety and
technological characterization of Staphylococcus equorum isolates
from jeotgal, a Korean high-salt-fermented seafood, for starter
development. Int J Food Microbiol. 188:108–115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee KW, Park JY, Sa HD, Jeong JH, Jin DE,
Heo HJ and Kim JH: Probiotic properties of Pediococcus strains
isolated from jeotgals, salted and fermented Korean sea-food.
Anaerobe. 28:199–206. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jung J, Lee SH, Jin HM, Jeon CO and Park
W: Pyrosequencing-based analysis of bacterial community and
metabolites profiles in Korean traditional seafood fermentation: A
flatfish-fermented seafood. Biosci Biotechnol Biochem. 78:908–910.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim MS and Park EJ: Bacterial communities
of traditional salted and fermented seafoods from Jeju Island of
Korea using 16S rRNA gene clone library analysis. J Food Sci.
79:M927–M934. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim YR, Kim EY, Lee JM, Kim JK and Kong
IS: Characterisation of a novel Bacillus sp. SJ-10
β-1,3-1,4-glucanase isolated from jeotgal, a traditional Korean
fermented fish. Bioprocess Biosyst Eng. 36:721–727. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Romero-Pérez GA, Inoue R, Ushida K and
Yajima T: A rapid method of screening lactic acid bacterial strains
for conjugated linoleic acid production. Biosci Biotechnol Biochem.
77:648–650. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Luo W, Chen M, Chen A, Dong W, Hou X and
Pu B: Isolation of lactic acid bacteria from pao cai, a Chinese
traditional fermented vegetable, with inhibitory activity against
Salmonella associated with fresh-cut apple, using a modelling
study. J Appl Microbiol. 118:998–1006. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vinod KR, Jones D and Udupa V: A simple
and effective heat induced antigen retrieval method. MethodsX.
3:315–319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Broggi MA, Schmaler M, Lagarde N and Rossi
SW: Isolation of murine lymph node stromal cells. J Vis Exp.
90:e518032014.
|
|
46
|
Yamashita H, Ito T, Kato H, Asai S, Tanaka
H, Nagai H and Inagaki N: Comparison of the efficacy of tacrolimus
and cyclosporine A in a murine model of
dinitrofluorobenzene-induced atopic dermatitis. Eur J Pharmacol.
645:171–176. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yun Y, Kim K, Choi I and Ko SG: Topical
herbal application in the management of atopic dermatitis: a review
of animal studies. Mediators Inflamm. 2014:7521032014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Schmitt N and Ueno H: Regulation of human
helper T cell subset differentiation by cytokines. Curr Opin
Immunol. 34:130–136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cross ML and Gill HS: Can immunoregulatory
lactic acid bacteria be used as dietary supplements to limit
allergies? Int Arch Allergy Immunol. 125:112–119. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Vernocchi P, Del Chierico F, Fiocchi AG,
El Hachem M, Dallapiccola B, Rossi P and Putignani L: Understanding
probiotics' role in allergic children: The clue of gut microbiota
profiling. Curr Opin Allergy Clin Immunol. 15:495–503. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Panzer AR and Lynch SV: Influence and
effect of the human microbiome in allergy and asthma. Curr Opin
Rheumatol. 27:373–380. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Putignani L, Del Chierico F, Petrucca A,
Vernocchi P and Dallapiccola B: The human gut microbiota: A dynamic
interplay with the host from birth to senescence settled during
childhood. Pediatr Res. 76:2–10. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Madonini ER: Probiotics and allergies:
Myth or reality? Eur Ann Allergy Clin Immunol. 46:196–200.
2014.PubMed/NCBI
|
|
54
|
Foolad N and Armstrong AW: Prebiotics and
probiotics: The prevention and reduction in severity of atopic
dermatitis in children. Benef Microbes. 5:151–160. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Żukiewicz-Sobczak W, Wróblewska P,
Adamczuk P and Silny W: Probiotic lactic acid bacteria and their
potential in the prevention and treatment of allergic diseases.
Cent Eur J Immunol. 39:104–108. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mansfield JA, Bergin SW, Cooper JR and
Olsen CH: Comparative probiotic strain efficacy in the prevention
of eczema in infants and children: A systematic review and
meta-analysis. Mil Med. 179:580–592. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Won TJ, Kim B, Lim YT, Song DS, Park SY,
Park ES, Lee DI and Hwang KW: Oral administration of Lactobacillus
strains from Kimchi inhibits atopic dermatitis in NC/Nga mice. J
Appl Microbiol. 110:1195–1202. 2011. View Article : Google Scholar : PubMed/NCBI
|