Introduction
Cryptococcus neoformans (C.
neoformans) is the most common fungus to cause
meningoencephalitis in the central nervous system (CNS) worldwide
(1). Each year, an estimated 1
million cases of cryptococcal meningitis are reported, with a
>60% mortality rate within the first 3 months of infection
(2,3). C. neoformans is typically
acquired by inhaling spores or desiccated yeast from the
environment (3). Following an
initial asymptomatic pulmonary infection, the organism is carried
in the bloodstream and subsequently disseminated to target organs,
including lung, skin and bone, which typically results in
lymphocytic meningitis (2–9). Results from experimental mouse models
and human cases of cryptococcal meningitis have indicated that
C. neoformans infection may also spread to the brain
(4–9). While the lungs are considered a common
site of infection, C. neoformans predominantly targets the
brain; however, this is dependent on its ability to overcome the
innate immune system of the host and cross the blood-brain barrier
(BBB) in the initial phase of infection (10).
To defend against C. neoformans infection in
the initial phase, the host employs several types of innate immune
cells, including macrophages, dendritic cells (DCs) and
neutrophils, which phagocytize invading fungi and generate reactive
oxygen species (ROS), nitrogen species (RNS) and chlorine species
to aid in host protection (11,12). In
response to the host innate immune response, C. neoformans
activates virulence factors, including polysaccharide capsules and
melanin pigment, to resist phagocytosis and avoid clearance
(13,14). Furthermore, C. neoformans
induces the activation of antioxidant enzymes, including superoxide
dismutase (SOD) and catalases, and the synthesis of antioxidants,
such as glutathione (GSH), to adapt to oxidative attack (14,15).
These anti-oxidative factors have been demonstrated to be important
for ROS and RNS resistance, repair of damage caused by oxidative
attack and survival in the host (13–15).
The present review summarized the current
understanding of the anti-innate immune response strategies
utilized by C. neoformans and the mechanism involved in
cryptococcal BBB traversal.
Innate immune system
Innate immune responses restrict the growth and
invasion of C. neoformans in mammalian hosts (16). Innate immune cells are the first
cells to encounter fungi and are the primary effector cells in the
destruction and clearance of cryptococcal infection (17–26)
(Table I). Furthermore, the
generation of oxidative products by phagocytic cells may directly
destroy the invading fungi (12).
 | Table I.Primary functions of host innate
immune cells. |
Table I.
Primary functions of host innate
immune cells.
| Cells | Primary
function | (Refs.) |
|---|
| Macrophages | Phagocytosis,
production of ROI, RNI, superoxide and nitric oxide | (17–20) |
| Neutrophils | Phagocytosis,
production of ROI, RNI, myeloperoxidase, defensins 1–4 and
lysozymes | (21–23) |
| Dendritic
cells | Fungal
internalization and destruction | (24) |
| Natural killer | Direct
destruction | (25,26) |
Macrophages
Macrophages are critical phagocytic cells within the
host innate immune system (17).
Complement and mannose receptors on the surface of macrophages
mediate the phagocytosis of C. neoformans (17). In addition, macrophages release high
levels of reactive oxygen intermediates (ROI), reactive nitrogen
intermediates (RNI), superoxide and nitric oxide, which damage DNA
and a number of chemical moieties (11). Furthermore, macrophages promote
Th1-like responses to induce fungal clearance (18,19).
However, C. neoformans is able to proliferate within the
macrophage phagosome, and may pass between macrophages (18–20).
Neutrophils
Similar to macrophages, neutrophils capture and
degrade pathogens and serve a specific role in the initiation of
inflammation in response to infection (21). Neutrophils enhance granuloma
formation to destroy C. neoformans by oxidative and
non-oxidative mechanisms (21).
Furthermore, a previous study indicated that myeloperoxidase, which
is located primarily in neutrophils, produces a strong oxidant
hypochlorous acid from hydrogen peroxide and chloride ions
(22). This is another predominant
mechanism that neutrophils use to defend against fungal infection
(22). In addition, neutrophils
contain defensins 1–4, which are cytotoxic to C. neoformans
(23).
DCs and natural killer (NK) cells
DCs phagocytize C. neoformans via complement
or anti-capsular antibody-mediated opsonization, which leads to
fungal internalization and destruction, ultimately resulting in
tumor necrosis factor-α secretion and DC activation (24). Once phagocytized, cryptococci are
degraded through oxidative and non-oxidative mechanisms following
passage through lysosomes (24). A
previous study indicated that NK cells bind and inhibit the growth
of C. neoformans in vitro and induce fungal clearance in
mice (25). In addition, previous
results suggest that NK cells may directly destroy C.
neoformans when mediated by perforin (26).
Innate immune system evasion
C. neoformans has a number of established virulence
factors, including polysaccharide capsules and melanin, which
function as non-enzymatic factors that potently influence the
overall pathogenicity and phagocytic resistance of C.
neoformans (27). A variety of
extracellular proteins, including phospholipases, proteases and
ureases (27), and enzymatic
components of the redox system, including thioredoxins (Trxs),
glutaredoxins (Grxs), peroxiredoxin (Prxs) and catalases, serve as
enzymatic factors for C. neoformans survival during innate
immune attack (27–66) (Table
II).
| Table II.Primary function of C.
neoformans antioxidant factors against host innate immune
cells. |
Table II.
Primary function of C.
neoformans antioxidant factors against host innate immune
cells.
| Antioxidant
factor | Function against
host innate immune cells | (Refs.) |
|---|
| Polysaccharide
capsules | Inhibition of
phagocytosis and resistance to phagosome digestion | (27–34) |
| Melanins | Scavenging ROS and
reactive nitrogen intermediates | (27,34–37) |
| Phospholipases | Promotion of
survival and replication in macrophages | (38–40) |
| Proteinase | Promotion of
replication in macrophages and damage to phagosomal membranes | (41,42) |
| Ureases | Scavenging
nitrogen | (33–35,43–45) |
| Peroxiredoxins | Metabolism of
peroxides and/or peroxynitrite | (46–48) |
| Thioredoxins,
glutaredoxins | Metabolism of ROS,
reduction of oxidized sulfhydryl groups and maintenance of cellular
redox homeostasis | (46,49–54) |
|
|
| (46,49,58–60) |
| Superoxide
dismutases | Conversion of
superoxide to hydrogen peroxide | (15,61–63) |
| Catalases | Conversion of
hydrogen peroxide to water and molecular oxygen | (64) |
| Cytochrome c
peroxidases | Degradation of
hydrogen peroxide | (65) |
| Alternative oxidase
genes | Interaction with
the classic oxidative pathway | (66) |
Roles of virulence factors in defense
against the innate immune response
Polysaccharide capsules
The polysaccharide capsule, which is composed of
90–95% glucuronxylomannan and 5% galactoxylomannan, is among the
most important virulence factors of C. neoformans that aids
the fungus to avoid recognition and phagocytosis by host phagocytes
(28). The capsule prevents the
phagocytosis of C. neoformans and resists phagosome
digestion to preserve C. neoformans survival (28,29).
Once engulfed by macrophages, the C. neoformans capsule may
release polysaccharides into vesicles surrounding the phagosome
that accumulate in the host cell cytoplasm, which promotes
macrophage dysfunction and lysis (29). In addition, the capsular material
suppresses the migration of phagocytes (30), interferes with cytokine secretion
(31), directly inhibits T-cell
proliferation (32), induces
macrophage apoptosis (33) and
delays the maturation and activation of human DCs (34). By contrast, acapsular mutants of
C. neoformans may be effectively recognized and induce
pro-inflammatory cytokine secretion in macrophages (29).
Melanin
Melanin is a negatively charged, hydrophobic pigment
that is located in the cell walls of C. neoformans, and
serves a key role in virulence and survival (34). A melanin gene disruption study has
indicated that wild-type melanin-producing C. neoformans are
more virulent (34). Melanin is
composed of aggregates of small particles or granules and exogenous
substrates (34,35). In the natural environment, melanin
protects fungi from ultraviolet light, high temperatures, freezing
and thawing (36). The potent
antioxidant activity of melanin provides protection against oxidant
concentrations similar to those produced by stimulated macrophages
(37). The oxidative burst that
follows phagocytosis is an important mechanism by which immune
effector cells mediate antimicrobial action, which suggests that
melanin may enhance virulence by protecting fungal cells against
immune system-stimulated oxidative attack (37).
Roles of extracellular proteins in
defense against the innate immune response
Phospholipases
Phospholipases are a heterogeneous group of enzymes
that cleave phospholipids to produce various biologically active
compounds, which alter the infection microenvironment and may favor
the survival of C. neoformans in the host (38). The action of phospholipases may
result in the destabilization of membranes, cell lysis and release
of lipid second messengers (39).
The secretion of PLB has been reported to promote the survival and
replication of C. neoformans in macrophages in vitro
(38). In addition, a previous study
demonstrated that disruption of the PLB1 gene led to reduced
virulence in vivo and growth inhibition in a macrophage-like
cell line (40).
Proteinase
Environmental and clinical isolates of C.
neoformans possess proteinase activity that has been
demonstrated to degrade host proteins, including collagen, elastin,
fibrinogen, immune-globulins and complement factors (41). Replication of C. neoformans
inside macrophages is accompanied by the production of enzymes,
including proteinases and phospholipases, which damage the
phagosomal membrane (42).
Therefore, cryptococcal proteinases may cause tissue damage,
providing nutrients to the pathogen and protection from the
host.
Ureases
As a nitrogen-scavenging enzyme, urease catalyzes
the hydrolysis of urea to ammonia and carbonate, and is an
important virulence factor of C. neoformans (43). C. neoformans may utilize
urease to invade the CNS via the BBB and cause life-threatening
meningoencephalitis (43,44). A previous study suggested that urease
promoted the sequestration of cryptococcal cells in the
microvasculature of the brain, while urease-negative strains seldom
penetrated the CNS or caused disease (45). Although the specific role of urease
protein in BBB invasion is unknown, it has been suggested that the
extracellular enzymatic degradation of urea to toxic ammonia may
damage endothelial cells and lead to an increase in barrier
permeability (33–35).
Roles of antioxidant systems in defense against
the innate immune response
Resistance to RNS and ROS through antioxidant
defense systems has been correlated with virulence in C.
neoformans clinical isolates, and has been associated with
in vitro and in vivo oxidative stress resistance
(46). There are several enzymatic
anti-oxidant systems that have been identified in C.
neoformans, including the Prx, Trx and Grx systems (Fig. 1).
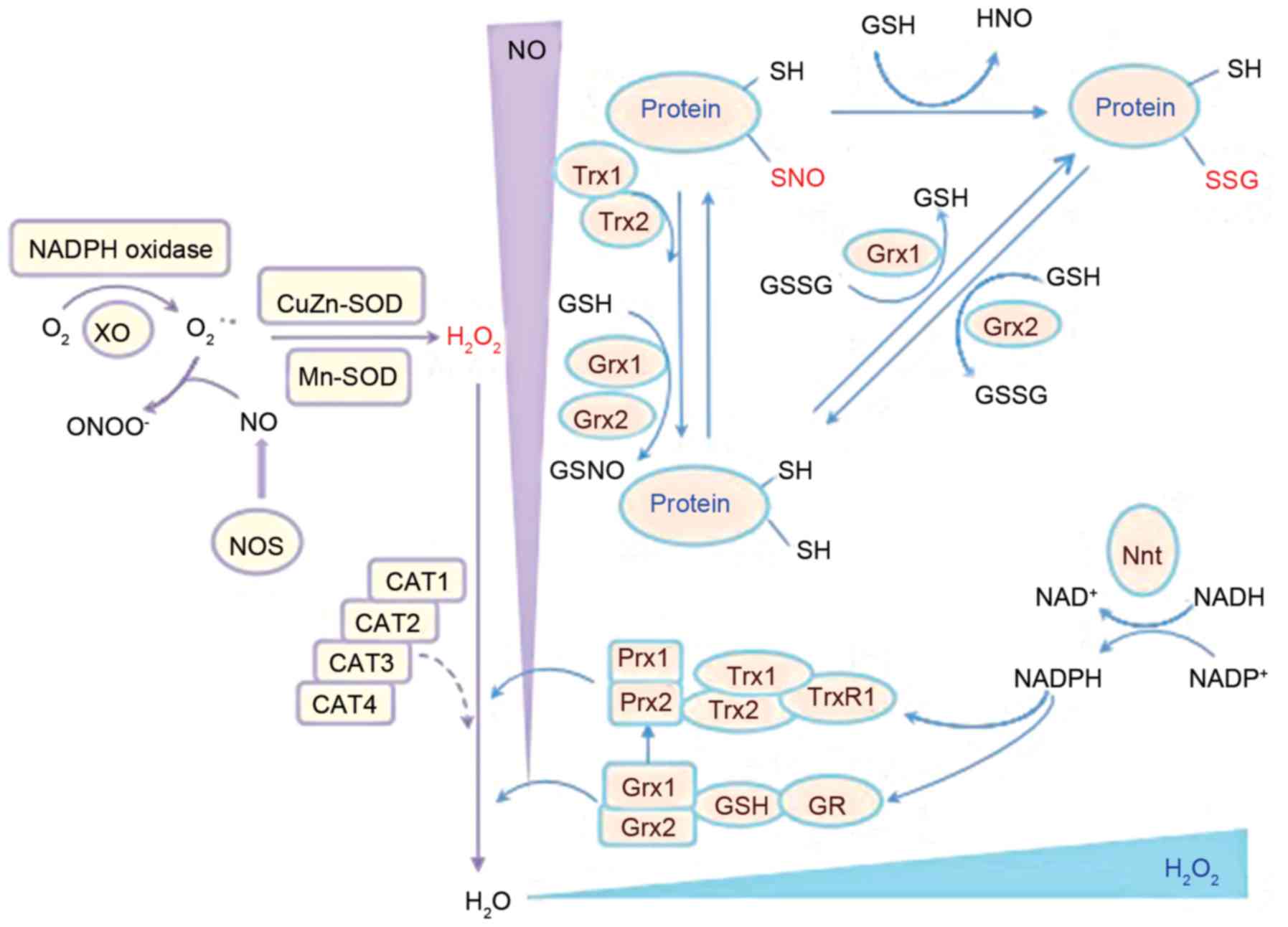 | Figure 1.Schematic representation of the
antioxidant chain of C. neoformans and the primary
antioxidant enzymes involved. Prx, Trx and Grx systems are major
enzymatic antioxidant systems in C. neoformans that regulate
redox balance (49–51) that are associated with 2 SODs,
Cu/Zn-SOD (SOD1) and Mn-SOD (SOD2), which may convert superoxide to
hydrogen peroxide (61). C.
neoformans, Cryptococcus neoformans; CAT, catalase; GR,
glutathione reductase; Grx, glutaredoxin; GSH, glutathione; GSSG,
glutathione disulfide; H2O2, hydrogen
peroxide; NADPH, nicotinamide adenine dinucleotide phosphate; Nnt,
nicotinamide nucleotide transhydrogenase; NO, nitric oxide; NOS,
nitric oxide synthase; protein-SNO, protein nitrosylation;
protein-SSG, protein glutathionylation; Prx1/2, peroxiredoxin 1/2;
SH, thiol; SOD, superoxide dismutase; Trx, thioredoxin; TrxR,
thioredoxin reductase. |
Prx systems
The Prx system is important for cellular processes
associated with disulfide bond formation, the anti-oxidative stress
response and pathogenesis of C. neoformans infection
(47). Prxs, also known as thiol
peroxidases, are 20–30 kDa-sized molecules that provide antioxidant
protection by removing peroxides (47). Prxs may be classified into 1-Cys and
2-Cys subgroups (48). Following
peroxidation, typical 2-Cys Prxs form homodimers through an
intersubunit disulfide bridge, whereas atypical 2-Cys Prxs form an
intramolecular disulfide bridge (48). By contrast, 1-Cys Prxs assume a
monomeric form with a single active cysteine site (48). In a previous study, two Prxs, TSA1
and TSA3, were discovered in C. neoformans, of which TSA1 is
highly conserved (48). In addition,
the findings of Missall et al (48) indicated that Prxs were induced under
oxidative and nitrosative stress and were critical for C.
neoformans virulence in mice. Furthermore, their study
demonstrated that deletion of TSA1, but not TSA3,
abolished virulence of the pathogen, which indicated that the
TSA1-mediated Prx system is a core antioxidant system in C.
neoformans (48).
Trx systems
The downstream component of Prx is the Trx system,
which is comprised of NADPH, Trx and thioredoxin reductase (TrxR),
and is involved in the regulation of DNA synthesis, gene
transcription, cell growth and apoptosis (49–51). Trx
is a small dithiol oxidoreductase that serves as a major carrier of
redox potential in cells (52) and a
cofactor for essential enzymes, and is involved in protein repair
via methionine sulphoxide reductase (53) and the reduction of protein
disulphides (54). This redox
control of the Trx system is considered to regulate the expression
of multiple stress defensive enzymes and protect cells against
oxidative stress (54).
C. neoformans contains two Trx proteins (TRX1
and TRX2) and one TrxR protein (TRXR1) (50), which are involved in the reduction
and recycling of the oxidized, inactive form of Prxs (55). TRX1 promotes normal growth and a
healthy oxidative state (50,53).
TRX2, though dispensable for vegetative growth, is important for
resistance to nitrosative stress (50,53).
TRXR1 is stimulated during oxidative stress induced by hydrogen
peroxide and nitrosative stress induced by nitric oxide (56). In C. neoformans, deletion of
these genes renders them sensitive to oxidative stress and results
in decreased survival in macrophage culture (50,57).
Grx systems
The GSH/Grxs system in C. neoformans is
another major thiol-dependent antioxidant system that participates
in the defense against oxidization (49). GSH is an important antioxidant for
fungi. Strains that lack or possess altered GSH are sensitive to
particular types of oxidative stress components, including
peroxides, superoxide anions and the toxic products of lipid
peroxidation (49). Exposure of
yeast cells to hydrogen peroxide caused a reduction in GSH levels
and a shift in the redox balance towards an oxidized state
(49). In C. neoformans, Grxs
are small heat-stable proteins encoded by two Grx genes
(GRX1 and GRX2), and are reduced in a manner similar
to Prx (49). GSH and Grxs may
regulate protein function by reversible protein S-glutathionylation
under oxidative stress (58). C.
neoformans contains two glutathione peroxidases (GPX1 and GPX2)
that respond differently to various stressors (59). A strain of C. neoformans that
lacked the GRX1 gene was sensitive to oxidative stress
induced by the superoxide anion, whereas a GRX2 mutant was
sensitive to oxidative stress generated by hydrogen peroxide
(59). Furthermore, GPX1 and
GPX2 deletion mutants have been reported as only mildly
sensitive to oxidant-induced destruction by macrophages and
exhibited no change in virulence in a murine model (59). GPX1 and GPX2 are involved in the
defense against organic peroxides, including tert-butyl
hydroperoxide (59). Although both
Gpx proteins are required for survival in macrophages, deletion of
GPX1 and GPX2 does not affect the virulence of C.
neoformans in mice (57),
suggesting that other peroxidases or antioxidant systems may
compensate for the loss. In addition, the GSH Prxs and GSH
S-transferases are involved in the breakdown of organic
hydroperoxides with GSH as a reductant or in the conjugation of
toxic lipophilic compounds to GSH, respectively (60).
Additional antioxidant systems
The primary function of SOD is to convert superoxide
to hydrogen peroxide (61). There
are four classes of SODs: Mn, Fe, Ni and Cu/Zn (15). C. neoformans has two SODs,
Cu/Zn-SOD (SOD1) and Mn-SOD (SOD2) (61). Cytosolic SOD1 has been demonstrated
to serve a pivotal role in the defense against ROS and contributed
to virulence in a mouse model (61).
Furthermore, in a previous study, a C. neoformans SOD1
mutant strain exhibited slower growth in macrophages and greater
susceptibility to neutrophil destruction (62). In addition, Narasipura et al
(63) reported that a C.
neoformans SOD1 mutant strain exhibited defects in
phospholipase, urease and laccase expression, which severely
attenuated virulence and the anti-phagocytotic activity of the
pathogen.
Catalases are antioxidant metalloenzymes that
promote the conversion of hydrogen peroxide to water and molecular
oxygen (64). C. neoformans
contains four catalases (CAT1, CAT2, CAT3 and CAT4), among which
CAT2 and CAT4 are the closest orthologs of yeast
CAT1 and CAT3 respectively (64). However, previous results have
suggested that deletion of all four CAT genes did not affect
the sensitivity to ROS or virulence of the pathogen (64), indicating that other
peroxide-detoxifying systems may have a complementary role in C.
neoformans. In addition, C. neoformans contains other
enzymatic factors that protect against oxidative stress. For
instance, cytochrome c peroxidase (CCP1) may degrade
hydrogen peroxide (65).
Furthermore, alternative oxidase gene (AOX1) is an enzyme that
forms part of the electron transport chain in the mitochondria in
C. neoformans, and its AOX1 mutant strain was demonstrated
to be more sensitive to the oxidative stressor tert-butyl
hydroperoxide; however, its target in the classic oxidative pathway
remains unknown (66).
Traversal of the BBB
After surviving phagocytosis and oxidative attack
initiated by the innate immune system, C. neoformans may be
carried in the bloodstream and disseminated to target organs,
including the brain (67,68).
The BBB ensures that the brain is protected, and
provides limited access to circulating macromolecules and
microorganisms (67,68). In order to infect the brain, C.
neoformans may use one of three potential traversal pathways to
cross the BBB: Transcellular traversal, the paracellular pathway
and Trojan horse dissemination (69–71)
(Fig. 2).
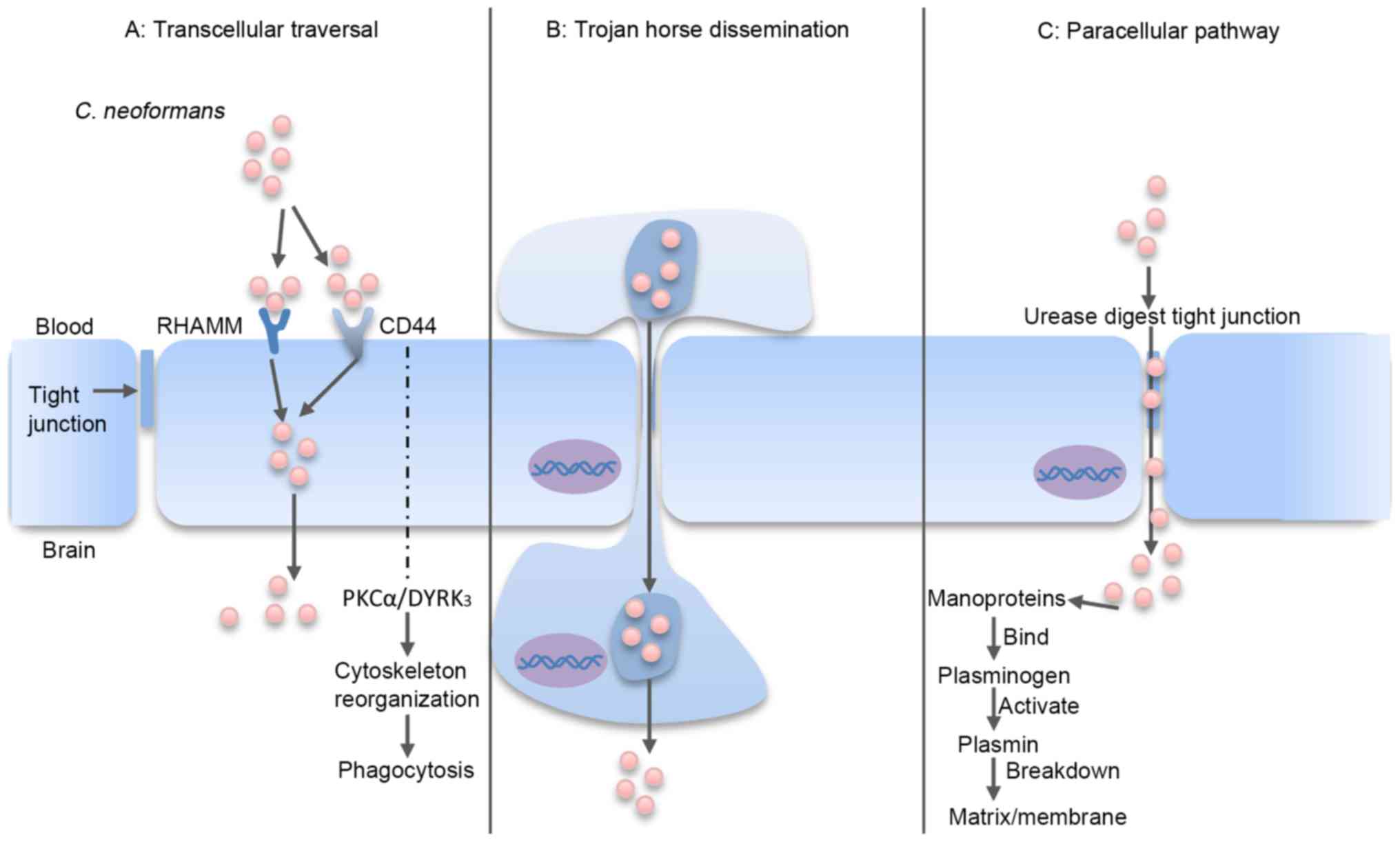 | Figure 2.C. neoformans traversal of the
BBB. (A) Transcellular traversal: C. neoformans binds to
receptors on the endothelial cell, which triggers cellular
endocytosis (72–79). (B) Trojan horse dissemination: C.
neoformans is phagocytosed by a macrophage, which is able to
cross the BBB, resulting in pathogen transportation into the brain
(85–92). (C) Paracellular pathway: C.
neoformans damages and weakens the intercellular tight
junctions, which facilitates passage of the organism between the
endothelial cells (80–82). RHAMM, receptor of hyaluronan-mediated
motility; CD44, cluster of differentiation 44; PKCα, protein kinase
Cα; DYRK3, dual-specificity tyrosine phosphorylation-regulated
kinase 3; BBB, blood-brain barrier; C. neoformans,
Cryptococcus neoformans. |
Transcellular traversal
Transcellular traversal refers to the penetration of
C. neoformans through barrier cells via the exploitation of
cellular endocytosis (72,73). Transcellular traversal of the BBB has
been widely studied using in vitro models, which have
demonstrated an ability of C. neoformans to adhere to one or
more receptors on the endothelial cell barrier (67,74).
Previous in vitro and in vivo results
have demonstrated that the glycoprotein cluster of differentiation
(CD)44 receptor on the surface of brain endothelial cells has a key
role in transcellular traversal invasion of C. neoformans
(75,76). CD44 is the endothelial cell receptor
for hyaluronic acid, and is located in lipid rafts/caveolae on the
endothelial cell surface (77,78).
When hyaluronic acid in the C. neoformans capsule binds to
CD44, a downstream signaling pathway mediated by protein kinase Cα
and dual-specificity tyrosine phosphorylation-regulated kinase 3 is
initiated, which triggers actin cytoskeleton reorganization and
phagocytosis (5,71,79–81).
Notably, Jong et al (79)
documented that CD44−/− mice exhibited only a 2-fold
reduction in cryptococcal meningitis compared with wild-type mice
following intravenous injection of C. neoformans.
Furthermore, knockdown of CD44 and the hyaluronan-mediated motility
(RHAMM) receptor in mice conferred significantly higher protection
and inhibited the invasion of C. neoformans in the brain
when compared with knockdown of either receptor alone (79). These results suggest that CD44 and
RHAMM serve as receptors for C. neoformans on the surface of
brain endothelial cells through binding to hyaluronic acid. In
addition, Maruvada et al (40) reported that C. neoformans PLB1
may interact with lipid mediators in the endothelial cell membrane
of the brain to convert GDP-Ras-related C3 botulinum toxin
substrate 1 (Rac1) to GTP-Rac1, which may then associate with
signal transducer and activator of transcription 3 (40). A recent study also indicated that
metalloprotease 1, belonging to the M36 class of proteases,
promoted the migration of C. neoformans across the brain
endothelium and into the CNS by facilitating cryptococcal
attachment to the endothelium surface, thus underscoring the
critical role of M36 proteases in BBB permeation (80).
Paracellular pathway
The paracellular pathway involves the entry of
C. neoformans through damaged or weakened tight junctions
between the intercellular spaces of endothelial cells (81,82).
Neuropil edema is a characteristic of endothelium damage (83,84), and
sequestration of C. neoformans in the brain microvessels and
cryptococcal binding to the endothelium has been demonstrated to
induce tight junction alterations (83,84). In
addition, cryptococcal mannoproteins bind and activate host
plasminogens (82). Activated
plasmins may subsequently bind and break down the extracellular
matrix and membrane, which increases the likelihood of paracellular
invasion (82).
A standard in vivo approach to investigate
the mechanisms involved in the paracellular pathway is intravenous
incubation of mice with C. neoformans and imaging of the
blood vessels in the brain (67).
Shi et al (71) imaged the
cerebral blood vessels of mice infected with a fluorescently
labeled C. neoformans urease mutant strain, and identified a
markedly reduced capacity of the mutant to traverse to the brain
(71). It is possible that urease
participates in the enzymatic digestion of tight junctions between
endothelial cells, and thus facilitates brain invasion via the
transcellular route (43).
Collectively, these findings indicate the possibility that C.
neoformans uses a paracellular entry mechanism by weakening the
brain endothelial tight junctions.
Trojan horse dissemination
The Trojan-horse dissemination of C.
neoformans traversal involves the transport of pathogens into
the brain within parasitized phagocytes (67,85).
Previous results support the existence of a Trojan horse mechanism
for BBB traversal (85). C.
neoformans is a facultative intracellular pathogen that may
survive and multiply inside phagocytes (86,87).
Furthermore, C. neoformans may infect other phagocytes
following their escape from phagocytic cells by vomocytosis,
leaving the host macrophage unharmed (19,88).
This direct cell-to-cell spread potentially explains how
cryptococci may exploit phagocytes to penetrate the BBB in a Trojan
horse manner (89). In previous
studies, C. neoformans was identified inside phagocytes on
the outer side of a meningeal capillary, which suggests that C.
neoformans may have been transported within circulating
phagocytes (90–92). These findings suggest that C.
neoformans may use the Trojan-horse dissemination model to
traverse into the brain.
Conclusions
The successful traversal of C. neoformans
across the BBB relies on failure of the defense mechanisms imposed
by the host innate immune system in the first phase of infection.
Various innate immune constituents, including macrophages and
neutrophils, contribute to phagocytosis, oxidative stress and
clearance of C. neoformans (27). However, C. neoformans contains
redundant layers of anti-phagocytic and anti-oxidative factors to
resist the innate immune cells of the host, including
polysaccharide capsules, melanin, phospholipases, proteases,
ureases, TSA1, TSA3, TRX1, TRX2, TRXR1, GRX1, GRX2, GPX1, GPX2,
SOD1, SOD2, CAT1, CAT2, CAT3, CAT4, CCP1 and AOX1.
Following its subversion of the innate immune
response, C. neoformans may disseminate to the brain.
However, entry into the highly protected environment of the brain
requires C. neoformans to overcome the BBB. C.
neoformans may gain entry through direct engulfment by
endothelial cells (transcellular traversal), inducing damage to
tight junctions (paracellular pathway) or hiding within phagocytes
(Trojan-horse dissemination). Following successful evasion of the
innate immune response, the fungi may proliferate (67). Results have indicated that >0.6
million cryptococcal meningitis cases each year result in mortality
within 3 months of infection, even with treatment. In light of the
present findings, the primary challenge in the field will be to
obtain sufficient resources to identify Cryptococcus
biomarkers for the improved determination of disease risk,
treatment and prevention using therapies and vaccines, which may
boost the immunity of the host to C. neoformans.
Acknowledgements
The present study was supported by the Nature
Foundation of China (grant no. 81550028) received by Dr J. Wang,
the Hubei Office of Education Foundation (grant nos. Q20151204 and
B2016022) received by Dr LL. Zou and Dr J. Wang, and the Yi Chang
Office of Healthcare Foundation (grant nos. A12301-25 and
A16301-14) received by Mr CL. Yang and Dr J. Wang.
References
|
1
|
Loyse A, Thangaraj H, Eastervrook P, Ford
N, Roy M, Chiller T, Govender N, Harrison TS and Bicanic T:
Cryptococcal meningitis: Improving access to essential antifungal
medicines in resource-poor countries. Lancet Infect Dis.
13:629–637. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Beardsley J, Wolbers M, Kibengo FM, Ggayi
AB, Kamali A, Cuc NT, Binh TQ, Chau NV, Farrar J, Merson L, et al:
Adjunctive dexamethasone in HIV-associated cryptococcal meningitis.
N Engl J Med. 374:542–554. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fang W, Fa Z and Liao W: Epidemiology of
Cryptococcus and cryptococcosis in China. Fungal Genet Biol.
78:7–15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu TB, Kim JC, Wang Y, Toffaletti DL,
Eugenin E, Perfect JR, Kim KJ and Xue C: Brain inositol is a novel
stimulator for promoting Cryptococcus penetration of the
blood-brain barrier. PLoS Pathog. 9:e10032472013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jong A, Wu CH, Prasadarao NV, Kwon-Chung
KJ, Chang YC, Ouyang Y, Shackleford GM and Huang SH: Invasion of
Cryptococcus neoformans into human brain microvascular
endothelial cells requires protein kinase C-alpha activation. Cell
Microbiol. 10:1854–1865. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Panackal AA, Wuest SC, Lin YC, Wu T, Zhang
N, Kosa P, Komori M, Blake A, Browne SK, Rosen LB, et al:
Paradoxical immune responses in non-HIV Cryptococcal meningitis.
PLoS Pathog. 11:e10048842015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Graciela Agar CH, Orozco Rosalba V, Macias
Ivan C, Agnes F, Juan Luis GA and José Luis SH: Cryptococcal
choroid plexitis an uncommon fungal disease. Case report and
review. Can J Neurol Sci. 36:117–122. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kumari R, Raval M and Dhun A: Cryptococcal
choroid plexitis: Rare imaging findings of central nervous system
cryptococcal infection in an immunocompetent individual. Br J
Radiol. 83:e14–e17. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schwerk C, Tenenbaum T, Kim KS and
Schroten H: The choroid pleus-a multi-role player during infectious
diseases of the CNS. Front Cell Neurosci. 9:802015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ngamskulrungroj P, Chang Y, Sionov E and
Kwon-Chung KJ: The primary target organ of Cryptococcus
gattii is different from that of Cryptococcus neoformans
in a murine model. MBio. 3:e00103–e00112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nathan C and Shiloh MU: Reactive oxygen
and nitrogen inter-mediates in the relationship between mammalian
hosts and microbial pathogens. Proc Natl Acad Sci USA. 97:pp.
8841–8848. 2000, View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Almeida F, Wolf JM and Casadevall A:
Virulence-associated enzymes of Cryptococcus neoformans.
Eukaryot Cell. 14:1173–1185. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
de Jesús-Berríos M, Liu L, Nussbaum JC,
Cox GM, Stamler JS and Heitman J: Enzymes that counteract
nitrosative stress promote fungal virulence. Curr Biol.
13:1963–1968. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kwon-Chung KJ, Fraser JA, Doering TL, Wang
Z, Janbon G, Idnurm A and Bahn YS: Cryptococcus neoformans
and Cryptococcus gattii, the etiologic agents of
cryptococcosis. Cold Spring Harb Perspect Med. 4:a0197602014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Upadhya R, Campbll LT, Donlin MJ, Aurora R
and Lodge JK: Global transcriptome profile of Cryptococcus
neoformans during exposure to hydrogen peroxide induced
oxidative stress. PLoS One. 8:e551102013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Voelz K, Lammas DA and May RC: Cytokine
signaling regulates the outcome of intracellular macrophage
parasitism by Cryptococcus neoformans. Infect Immun.
77:3450–3457. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Redlich S, Ribes S, Schütze S, Eiffert H
and Nau R: Toll-like receptor stimulation increases phagocytosis of
Cryptococcus neoformans by microglial cells. J
Neuroinflammation. 10:712013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chrisman CJ, Albuquerque P, Guimaraes AJ,
Nieves E and Casadevall A: Phospholipids trigger Cryptococcus
neoformans capsular enlargement during interactions with
amoebae and macrophages. PLoS Pathog. 7:e10020472011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Alvarez M and Casadevall A: Cell-to-cell
spread and massive vacuole formation after Cryptococcus
neoformans infection of murine macrophages. BMC Immunol.
8:162007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rohatgi S and Pirofski LA: Host immunity
to Cryptococcus neoformans. Future Microbiol. 10:565–581.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mambula SS, Simons ER, Hastey R, Selsted
ME and Levitz SM: Human neutrophil-mediated nonoxidative antifungal
activity against Cryptococcus neoformans. Infect Immun.
68:6257–6264. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Winterbourn CC, Vissers MC and Kettle AJ:
Myeloperoxidase. Curr Opin Hematol. 7:53–58. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Alcouloumre MS, Ghannoum MA, Ibrahim AS,
Selsted ME and Edwards JE Jr: Fungicidal properties of defensin
NP-1 and activity against Cryptococcus neoformans in vitro.
Antimicrob Agents Chemother. 37:2628–2632. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wozniak KL and Levitz SM: Cryptococcus
neoformans enters the endolysosomal pathway of dendritic cells
and is killed by lysosomal components. Infect Immun. 76:4764–4771.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Islam A, Li SS, Oykhman P, Timm-McCann M,
Huston SM, Stack D, Xiang RF, Kelly MM and Mody CH: An acidic
microenvironment increases KN cell killing Cryptococcus
neoformans and Cryptococcus gattii by enhancing perforin
degtanulation. PLoS Pathog. 9:e10034392013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marr KJ, Jones GJ, Zheng C, Huston SM,
Timm-McCann M, Islam A, Berenger BM, Ma LL, Wisenman JC and Mody
CH: Cryptococcus neoformans directly stimulates perforin
production and rearms NK cells for enhanced anticryptococcal
microbicidal activity. Infect Immun. 77:2436–2446. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rodrigues ML, Nakayasu ES, Oliveira DL,
Nimrichter L, Nosanchuk JD, Almeida IC and Casadevall A:
Extracellular vesicles produced by Cryptococcus neoformans
contain protein components associated with virulence. Eukaryot
Cell. 7:58–67. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Doering TL: How sweet it is! Cell wall
biogenesis and polysaccharide capsule formation in Cryptococcus
neoformans. Annu Rev Microbiol. 63:223–247. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Leopold Wager CM, Hole CR, Wozniak KL,
Olszewski MA and Wormley FL Jr: STAT1 signaling is essential for
protection against Cryptococcus neoformans infection in
mice. J Immunol. 193:4060–4071. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Haynes BC, Skowyra ML, Spencer SJ, Gish
SR, Williams M, Held EP, Brent MR and Doering TL: Toward an
integrated model of capsule regulation in Cryptococcus
neoformans. PLoS Pathog. 7:e10024112011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Villena SN, Pinheiro RO, Pinheiro CS,
Nunes MP, Takiya CM, Dosreis GA, Previato JO, Mendonça-Previato L
and Freire-de-Lima CG: Capsular polysaccharides galactoxylomannan
and glucuronoxylomannan from Cryptococcus neoformans induce
macrophage apoptosis mediated by Fas ligand. Cell Microbiol.
10:1274–1285. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yauch LE, Lam JS and Levitz SM: Direct
inhibition of T-cell responses by the Cryptococcus capsular
polysaccharide glucuronoxylomannan. PLoS Pathog. 2:e1202006.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lupo P, Chang YC, Kelsall BL, Farber JM,
Pietrella D, Vecchiarelli A, Leon F and Kwon-Chung KJ: The presence
of capsule in Cryptococcus neoformans influences the gene
expression profile in dendritic cells during interaction with the
fungus. Infect Immun. 76:1581–1589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chatterjee S, Prados-Rosales R, Itin B,
Casadevall A and Stark RE: Solid-state NMR reveals the carbon-based
molecular architecture of Cryptococcus neoformans fungal
eumelanins in the cell wall. J Biol Chem. 290:13779–13790. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sundaram C, Shantveer GU, Umabala P and
Lakshmi V: Diagnostic utility of melanin production by fungi: Study
on tissue sections and culture smears with Masson-Fontana stain.
Indian J Pathol Microbiol. 57:217–222. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nosanchuk JD, Rosas AL, Lee SC and
Casadevall A: Melanisation of Cryptococcus neoformans in
human brain tissue. Lancet. 355:2049–2050. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mauch RM, Cunha Vde O and Dias AL: The
copper interference with the melanogenesis of Cryptococcus
neoformans. Rev Inst Med Trop Sao Paulo. 55:117–120. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Santangelo R, Zoellner H, Sorrell T,
Wilson C, Donald C, Djordjevic J, Shounan Y and Wright L: Role of
extracellular phospholipases and mononuclear phagocytes in
dissemination of cryptococcosis in a murine model. Infect Immun.
72:2229–2239. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Evans RJ, Li Z, Hughes WS, Djordjevic JT,
Nielsen K and May RC: Cryptococcal phospholipase B1 is required for
intracellular proliferation and control of titan cell morphology
during macrophage infection. Infect Immun. 83:1296–1304. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Maruvada R, Zhu L, Pearce D, Zheng Y,
Perfect J, Kwon-Chung KJ and Kim KS: Cryptococcus neoformans
phospholipase B1 activates host cell Rac1 for traversal across the
blood-brain barrier. Cell Microbiol. 14:1544–1553. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rawson RB: The site-2 protease. Biochim
Biophys Acta. 1828:2801–2807. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Desalermos A, Tan X, Rajamuthiah R,
Arvanitis M, Wang Y, Li D, Kourkoumpetis TK, Fuchs BB and Mylonakis
E: A multi-host approach for the systematic analysis of virulence
factors in Cryptococcus neoformans. J Infect Dis.
211:298–305. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Morrow CA and Fraser JA: Is the
nickel-dependent urease complex of Cryptococcus the pathogen's
Achilles' heel? MBio. 4:e00408–e00413. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Singh A, Panting RJ, Varma A, Saijo T,
Waldron KJ, Jong A, Ngamskulrungroj P, Chang YC, Rutherford JC and
Kwon-Chung KJ: Factors required for activation of urease as a
virulence determinant in Cryptococcus neoformans. MBio.
4:e00220–13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Olszewski MA, Noverr MC, Chen GH, Toews
GB, Cox GM, Perfect JR and Huffnagle GB: Urease expression by
Cryptococcus neoformans promotes microvascular
sequestration, thereby enhancing central nervous system invasion.
Am J Pathol. 164:1761–1771. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Breitenbach M, Weber M, Rinnerthaler M,
Karl T and Breitenbach-Koller L: Oxidative stress in fungi: Its
function in signal transduction, interaction with plant hosts, and
lignocellulose degradation. Biomolecules. 5:318–342. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Toledano MB, Delaunay-Moisan A, Outten CE
and Igbaria A: Functions and cellular copartmentation of the
thioredoxin and glutathione pathways in yeast. Antioxid Redox
Signal. 18:1699–1711. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Missall TA, Pusateri ME and Lodge JK:
Thiol peroxidase is critical for virulence and resistance to nitric
oxide and peroxide in the fungal pathogen, Cryptococcus
neoformans. Mol Microbiol. 51:1447–1458. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Benhar M, Shytaj IL, Stamler JS and
Savarino A: Dual targeting of the thioredoxin and glutathione
systems in cancer and HIV. J Clin Invest. 126:1630–1639. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Missall TA and Lodge JK: Function of
thioredoxin proteins in Cryptococcus neoformans during stress or
virulence and regulation by putative transcriptional modulators.
Mol Microbiol. 57:847–858. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Benhar M: Nitric oxide and the thioredoxin
system: A complex interplay in redox regulation. Biochim Biophys
Acta. 1850:2476–2484. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Netto LE and Antunes F: The roles of
peroxiredoxin and thioredoxin in hydrogen peroxide sensing and in
signal transduction. Mol Cells. 39:65–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lu J and Holmgren A: The thioredoxin
antioxidant system. Free Radic Biol Med. 66:75–87. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yoshioka J: Thioredoxin superfamily and
its effects on cardiac physiology and pathology. Compr Physlol.
5:513–530. 2015. View Article : Google Scholar
|
|
55
|
Ianiri G and Idnurm A: Essential gene
discovery in the basidiomycete Cryptococcus neoformans for
antifungal drug target prioritization. MBio. 6:e02334–14. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Missall TA and Lodge JK: Thioredoxin
reductase is essential for viability in the fungal pathogen
Cryptococcus neoformans. Eukaryot Cell. 4:487–489. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kalinina EV, Chernov NN and Novivhkova MD:
Role of glutathione transferase, and glutaredoxin in regulation of
redox-dependent processes. Biochemistry (Mosc). 79:1562–1583. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lu J and Holmgren A: The thioredoxin
antioxidant system. Free Radic Biol Med. 66:75–87. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Grant CM: Role of the
glutathione/glutaredoxin and thioredoxin systems in yeast growth
and response to stress conditions. Mol Microbiol. 39:533–541. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Hanschmann EM, Godoy JR, Berndt C,
Hudemann C and Lilling CH: Thioredoxins, glutaredoxins, and
peroxiredoxins-molecular mechanisms and health significance: From
cofactors to antioxidants to redox signaling. Antioxid Redox
Signal. 19:1539–1605. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Narasipura SD, Chaturvedi V and Chaturvedi
S: Characterization of Cryptococcus neoformans variety
gattii SOD2 reveals distinct roles of the two superoxide dismutases
in fungal biology and virulence. Mol Microbiol. 55:1782–1800. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Cox GM, Harrison TS, McDade HC, Taborda
CP, Heinrich G, Casadevall A and Perfect JR: Superoxide dismutase
influences the virulence of Cryptococcus neoformans by
affecting growth within macrophages. Infect Immun. 71:173–180.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Narasipura SD, Ault JG, Behr MJ,
Chaturvedi V and Chaturvedi S: Characterization of Cu, Zn
superoxide dismutase (SOD1) gene knock-out mutant of
Cryptococcus neoformans var. gattii: Role in biology and
virulence. Mol Microbiol. 47:1681–1694. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Giles SS, Stajich JE, Nichols C, Gerrald
QD, Alspaugh JA, Dietrich F and Perfect JR: The Cryptococcus
neoformans catalase gene family and its role in antioxidant
defense. Eukaryot Cell. 5:1447–1459. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Giles SS, Perfect JR and Cox GM:
Cytochrome c peroxidase contributes to the antioxidant
defense of Cryptococcus neoformans. Fungal Genet Biol.
42:20–29. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Akhter S, McDade HC, Gorlach JM, Heinrich
G, Cox GM and Perfect JR: Role of alternative oxidase gene in
pathogenesis of Cryptococcus neoformans. Infect Immun.
71:5794–5802. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Vu K, Eigenheer RA, Phinney BS and Gelli
A: Cryptococcus neoformans promotes its transmigration into
the central nervous system by inducing molecular and cellular
changes in brain endothelial cells. Infect Immun. 81:3139–3147.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zou LL, Ma JL, Wang T, Yang TB and Liu CB:
Cell-penetrating peptide-mediated therapeutic molecule delivery
into the central nervous system. Curr Neuropharmacol. 11:197–208.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Tseng HK, Liu CP, Price MS, Jong AY, Chang
JC, Toffaletti DL, Betancourt-Quiroz M, Frazzitta AE, Cho WL and
Perfect JR: Identification of genes from the fungal pathogen
Cryptococcus neoformans related to transmigration into the
central nervous system. PLoS One. 7:e450832012. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Stie J and Fox D: Blood-brain barrier
invasion by Cryptococcus neoformans is enhanced by
functional interactions with plasmin. Microbiology. 158:240–258.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Shi M, Li SS, Zheng C, Jones GJ, Kim KS,
Zhou H, Kubes P and Mody CH: Real-time imaging of trapping and
urease-dependent transmigration of Cryptococcus neoformans
in mouse brain. J Clin Invest. 120:1683–1693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Charlier C, Chrétien F, Baudrimont M,
Mordelet E, Lortholary O and Dromer F: Capsule structure changes
associated with Cryptococcus neoformans crossing of the
blood-brain barrier. Am J Pathol. 166:421–432. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Chang YC, Wang Z, Flax LA, Xu D, Esko JD,
Nizet V and Baron MJ: Glycosaminoglycan binding facilitates entry
of a bacterial pathogen into central nervous systems. PLoS Pathog.
7:e10020822011. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Vu K, Weksler B, Romero I, Couraud PO and
Gelli A: Immortalized human brain endothelial cell line HCMEC/D3 as
a model of the blood-brain barrier facilitates in vitro studies of
central nervous system infection by Cryptococcus neoformans.
Eukaryot Cell. 8:1803–1807. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Jong A, Wu CH, Gonzales-Gomez I,
Kwon-Chung KJ, Chang YC, Tseng HK, Cho WL and Huang SH: Hyaluronic
acid receptor CD44 deficiency is associated with decreased
Cryptococcus neoformans brain infection. J Biol Chem.
287:15298–15306. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Jong A, Wu CH, Chen HM, Luo F, Kwon-Chuang
KJ, Chang YC, Lamunyon CW, Plaas A and Huang SH: Identification and
characterization of CPS1 as a hyaluronic acid synthase contributing
to the pathogenesis of Cryptococcus neoformans infection.
Eukaryot Cell. 6:1486–1496. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Huang SH, Long M, Wu CH, Kwon-Chung KJ,
Chang YC, Chi F, Lee S and Jong A: Invasion of Cryptococcus
neoformans into human brain microvascular endothelial cells is
mediated through the lipid rafts-endocytic pathway via the dual
specificity tyrosine phosphorylation-regulated kinase 3 (DYRK3). J
Biol Chem. 286:34761–34769. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Long M, Huang SH, Shackleford GM and Jong
A: Lipid raft/caveolae signaling is required for Cryptococcus
neoformans invasion into human brain microvascular endothelial
cells. J Biomed Sci. 19:192012. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Jong A, Wu CH, Shackleford GM, Kwon-Chung
KJ, Chang YC, Chen HM, Quyang Y and Huang SH: Involvement of human
CD44 during Cryptococcus neoformans infection of brain
microvascular endothelial cells. Cell Microbiol. 10:1313–1326.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Vu K, Tham R, Uhrig JP, Thompson GR III,
Pombejra Na S, Jamklang M, Bautos JM and Gelli A: Invasion of the
central nervous system by Cryptococcus neoformans requires a
secreted fungal metalloprotease. MBio. 5:e01101–e1114. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Kim KS: Mechanisms of microbial traversal
of the blood-brain barrier. Nat Rev Microbiol. 6:625–634. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Stie J, Bruni G and Fox D:
Surface-associated plasminogen binding of Cryptococcus
neoformans promotes extracellular matrix invasion. PLoS One.
4:e57802009. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Charlier C, Chrétien F, Lortholary O and
Dromer F: Early capsule structure changes associated with
Cryptococcus neoformans crossing of the blood-brain barrier. Med
Sci (Paris). 21:685–687. 2005.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Chen SH, Stins MF, Huang SH, Chen YH,
Kwon-Chung KJ, Chang Y, Kim KS, Suzuki K and Jong AY:
Cryptococcus neoformans induces alterations in the
cytoskeleton of human brain microvascular endothelial cells. J Med
Microbiol. 52:961–970. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Charlier C, Nielsen K, Daou S, Brigitte M,
Chretien F and Dromer F: Evidence of a role for monocytes in
dissemination and brain invasion by Cryptococcus neoformans.
Infect Immun. 77:120–127. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Alvarez M and Casadevall A: Phagosome
extrusion and host-cell survival after Cryptococcus
neoformans phagocytosis by macrophages. Curr Biol.
16:2161–2165. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Coelho C, Souza AC, Derengowski Lda S, de
Leon-Rodriguez C, Wang B, Leon-Rivera R, Bocca AL, Gonçalves T and
Casadevall A: Macrophage mitochondrial and stress response to
ingestion of Cryptococcus neoformans. J Immunol.
194:2345–2357. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Sorrell TC, Juillard PG, Djordjevic JT,
Kaufman-Francis K, Dietman A, Milonig A, Combes V and Grau GE:
Cryptococcal transmigration across a model brain blood-barrier:
Evidence of the Trojan horse mechanism and differences between
Cryptococcus neoformans var. grubii strain H99 and
Cryptococcus gattii strain R265. Microbes Infect. 18:57–67.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Ma H, Croudace JE, Lammas DA and May RC:
Direct cell-to-cell spread of a pathogenic yeast. BMC Immunol.
8:152007. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Davis MJ, Eastman AJ, Qiu Y, Greorka B,
Kozel TR, Osterholzer JJ, Curtis JL, Swanson JA and Olszewski MA:
Cryptococcus neoformans-induced macrophage lysosome damage
crucially contributes to fungal virulence. J Immunol.
194:2219–2231. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Liu TB, Perlin DS and Xue C: Molecular
mechanisms of cryptococcal meningitis. Virulence. 3:173–181. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Alanio A, Vernel-Pauillac F,
Sturny-Leclère A and Dromer F: Cryptococcus neoformans host
adaptation: Toward biological evidence of dormancy. MBio.
6:e02580–14. 2015. View Article : Google Scholar : PubMed/NCBI
|














