Introduction
Diabetic nephropathy (DN) previously known as
idiopathic nodular glomerulosclerosis is the leading cause of
end-stage renal disease. It is characterized by increased blood
pressure, increased urinary albumin and glomerular lesions, leading
to the loss of glomerular filtration is the main microvascular
complications of diabetes (DM) (1,2). There
are a variety of mechanisms believed to be involved, including
non-enzymatic glycosylation, polyol pathway theory and protein
kinase C theory (3–5). What is the most widely accepted
hypothesis of DN is the non-enzymatic glycoside theory. In the
long-term hyperglycemic state, glucose molecules and protein in
vivo occur non-enzymatic glycosylation reaction and yield
advanced glycation end products (advanced glycation or
glycoxidation end-products, AGEs), leading to the occurrence of the
disease (6,7). Lots of studies have confirmed that
eliminating the effects of AGEs can reduce renal damage, inhibit
the progress of complications of diabetes and promote the
regeneration of renal cells, but there are few studies focus on
these (8,9). Some potential targets for
renoprotective therapies, including AGEs inhibitors, PKC
inhibitors, have been studied and tested in cells and animals
models, the development of diabetic nephropathy remains a major
concern. Therefore, more specific pathophysiologic mechanisms
underlying DN should be explored and identified to develop new
treatment and strategies for this devastating disease.
Previous study found that AGEs could induce
apoptosis and dysfunction of tubular cells may contribute in part
to glomerular hyperfiltration, an early renal dysfunction in
diabetes, so that late glomerulosclerosis (10,11).
Autophagy engages in complex interplay with apoptosis. Apoptosis
and autophagy occur simultaneously within the same cell under the
stimulation of various stress including starvation, hypoxia and ER
stress (12). In the present study,
we explored the relationship between autophagy and apoptosis under
AGEs treatment. Whether curcumin could reduce AGEs-induced cell
apoptosis and what role of autophagy in this process. Furthermore,
we determined whether curcumin exerts a protective effect through
autophagy pathway in response to AGEs.
Materials and methods
Materials
Curcumi, AGEs, Triton X-100, DMSO, LY294002 and
3-methyladenine (3-MA) were purchased from Sigma-Aldrich (Merck
KGaA, Darmstadt, Germany). DMEM and fetal bovine serum (FBS) were
both obtained from HyClone (Thermo Fisher Scientific, Inc., Logan,
UT, USA). Anti-GAPDH, anti-bax, anti-AIF, anti-caspase-3 and
anti-p-AKT were obtained (Santa Cruz Biotechnology, Inc., Dallas,
TX, USA). Anti-Beclin 1 and anti-LC3 were both obtained from Cell
Signaling Technology, Inc. (Beverly, MA, USA). ECL kit was
purchased (Pierce; Thermo Fisher Scientific, Inc.). Flow cytometer
(FACSCalibur; Becton-Dickinson, Franklin Lakes, NJ, USA). All
reagents used were trace element analysis grade. All water used was
glass distilled.
Cell culture
Rat kidney tubular epithelial cell line NRK-52E was
purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA). The cells were cultured in a 5% CO2
atmosphere in Dulbecco's modified Eagle's medium (DMEM; low
glucose), supplemented with 10% fetal bovine serum, 4 mM
L-glutamine and 1% penicillin/streptomycin at a density of 6×103
cells/well in six-well culture plates.
Cell viability
We use
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay to detect cell viability. Briefly, the sample medium were
added to 10 µl MTT (500 µg/ml) and incubated for 3 h at 37°C
following treatment. Subsequently, the MTT solution was removed and
100 µl dimethyl sulfoxide (DMSO) was added to dissolve the colored
formazan crystals. The absorbance of each aliquot at 490 nm was
measured using a Sunrise microplate reader (Tecan Group Ltd.,
Männedorf, Switzerland). The cell viability was determined as the
ratio of the signal.
Measurement of apoptosis by flow
aytometry and terminal uridine nick 3′ end labelling (TUNEL)
assays
We detected apoptosis by Annexin V labeled with
FITC. Propidium iodide was used to determine cell necrosis. After
exposure to various experimental conditions, cells were trypsinized
and labeled with fluorochromes at 37°C, and then cytofluorometric
analysis was performed with a FACS can (Becton-Dickinson).
Apoptosis was also evaluated by the TUNEL method.
For the TUNEL assays, cells grown on a coverslip were pretreated
with various experimental conditions. The TUNEL assay kit was used
to detect apoptotic cells under a fluorescence microscope (Leica
TCS SPE; Leica Microsystems GmbH, Wetzlar, Germany). After
treatment, the cells were washed with PBS, fixed in 4%
paraformaldehyde/PBS, and permeabilized with 0.2% Triton X-100 in
citrate buffer. Samples were incubated with TdT and
fluorescein-labeled dUTP, counterstained with
4′,6-diamidino-2-phenylindole (DAPI), and then observed under a
fluorescence microscope (Leica TCS SPE). Percentages of apoptotic
cells were estimated by counting 300 cells in random fields.
Immunofluorescence staining
NRK-52E cells were fixed with 4% paraformaldehyde at
4°C for 30 min, incubated with 0.2% Triton X-100 for 10 min. The
cells were blocked at non-specific antibody binding sites by
incubating with 10% goat serum in PBS containing 0.3% Triton X-100
and 0.5% bovine serum albumin (BSA) for 30 min at room temperature,
followed by incubation with a mouse monoclonal antibody against
Beclin 1 and LC3 (1:400 in PBS; Cell Signaling Technology, Inc.)
overnight. Then TRITC and FITC-conjugated goat anti-mouse IgG
(1:100 in PBS) was used to incubate for 0.5 h at room temperature.
Heochst 33342 was added to the cells for 15 min. After washing
three times with PBS, cells were visualized under fluorescence
microscopy.
Western blot analysis
The NRK-52E cells were lysed in protein extraction
reagent (Tissue Protein Extraction kit; Pierce, Rockford, IL, USA)
and the lysates were extracted by centrifugation. Equal amounts of
protein were loaded per sample in each experiment, separated by
SDS-PAGE and transferred to polyvinylidene difluoride membranes by
electroblotting. The protein-bound membranes were blocked and
washed in Tris-buffered saline (TBS)-Tween-20. The membranes were
incubated overnight with primary antibodies. The primary antibodies
used in the present study were as follows: anti-GAPDH, anti-bax,
anti-AIF, anti-caspase-3 and anti-p-AKT (Santa Cruz Biotechnology,
Inc.). Anti-Beclin 1 and anti-LC3 (Cell Signaling Technology,
Inc.). After washing in TBS-0.1% Tween-20, the membranes were then
incubated with horseradish peroxidase-conjugated secondary
antibodies overnight at 4°C. Finally, the blots were developed
using an enhanced chemiluminescence system (Pierce). To compare the
levels of proteins, the density of each signal was evaluated by
image analysis software (CS Analyzer; ATTO, Tokyo, Japan).
Statistical analysis
Statistical comparison was made on the differences
in mean values among multiple groups by using one-way analysis of
variance with post hoc Scheffe's test. Statistical significance was
achieved if P-values were <0.05.
Results
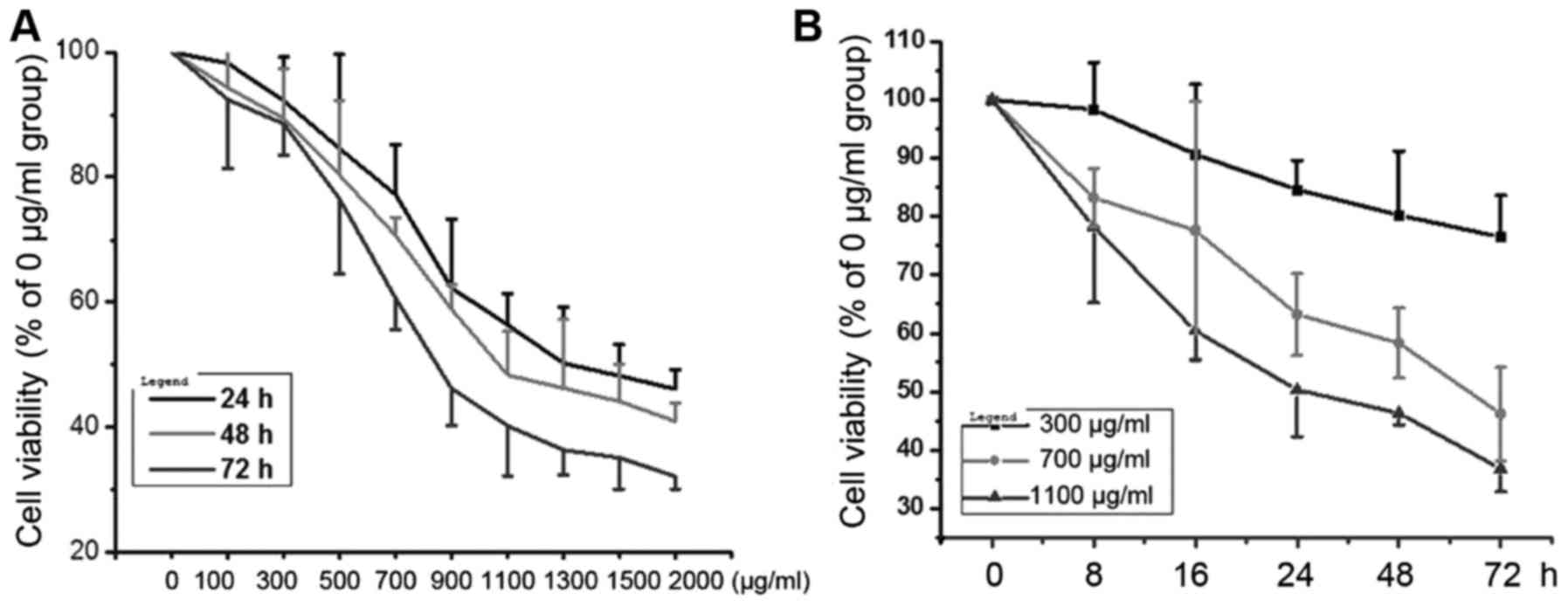
AGEs inhibits cell viability in
NRK-52E cells
The cell viability of NRK-52E cells under AGEs
conditions were assessed by MTT. As shown in (Fig. 1A), when the cells were treated with
0–2,000 µg/ml AGEs for 24, 48 and 72 h, AGEs significantly
inhibited NRK-52E cells viability in concentration-dependent
manner. Similarly, when the cells were treated with 300, 700 and
1,000 µg/ml AGEs for 0–72 h, AGEs inhibited NRK-52E cell viability
in time-dependent manner (Fig.
1B).
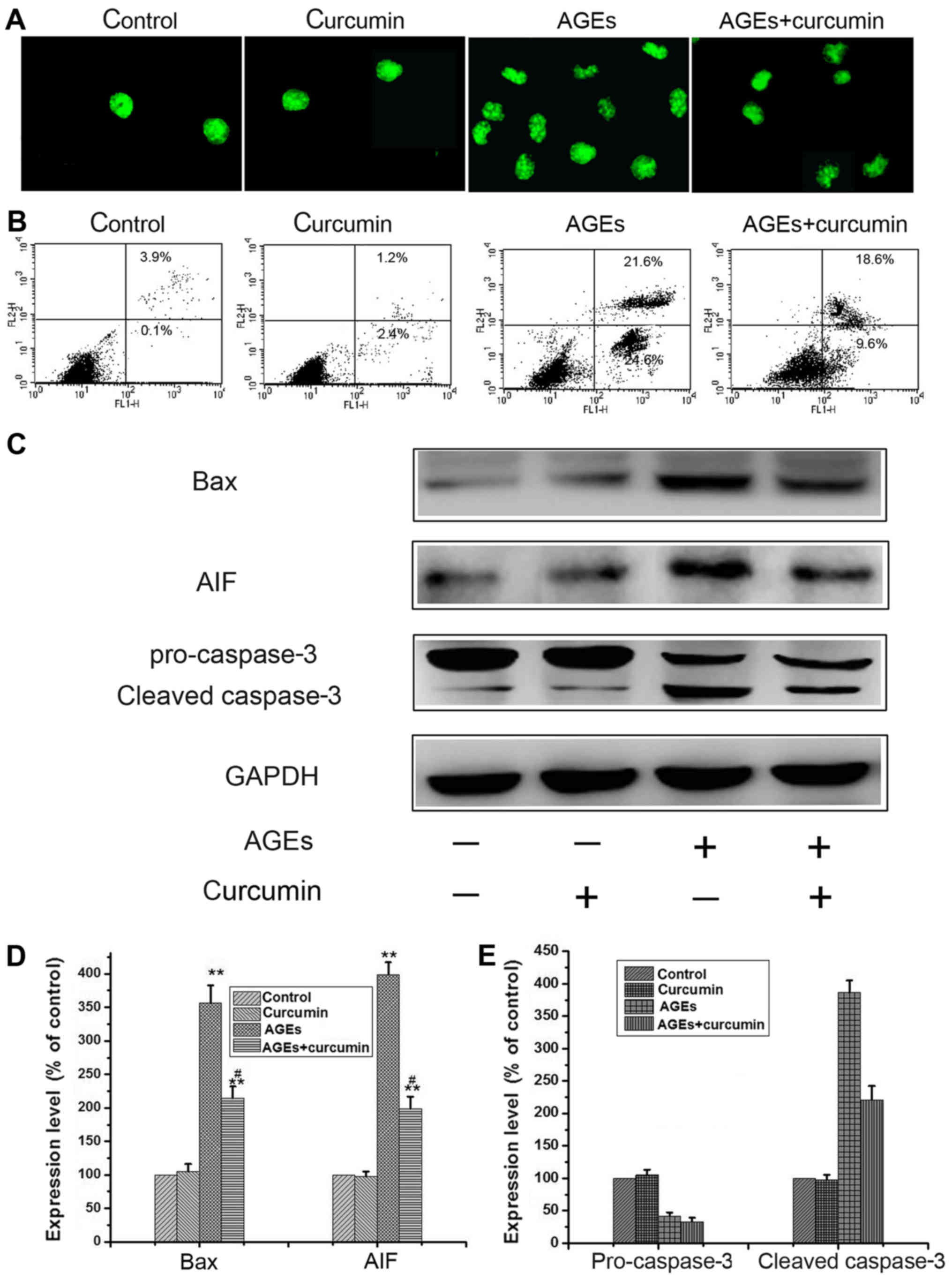
Curcumin inhibits AGEs-induced cell
apoptosis
Curcumin, the active ingredient from the spice
turmeric (Curcuma longa L.), has been demonstrated recently to
possess anti-apoptosis effects (13). This study aimed to investigate the
protective effect of curcumin on AGEs induced apoptosis and the
underlying mechanism in NRK-52E cells. We then further hypothesize
that curcumin may also play a role in the regulation of apoptosis
under AGEs. As shown in (Fig. 2),
the NRK-52E cells were treated with 700 µg/ml AGEs with or without
10 µM curcumin for 48 h. Firstly, AGEs-induced apoptosis was
determined by TUNEL analysis. As can be seen from (Fig. 2A), AGEs treatment alone led to a
significant increase in the apoptotic population, while curcumin
cotreatment resulted in a substantial decrease of AGEs-induced
NRK-52E cells apoptosis. We further evaluated the anti-apoptotic
effect of curcumin by directly assessing the percentage of
apoptotic cells using flow cytometry followed by Annexin V/PI
double staining. Addition of curcumin reduced the percentage of
AGEs-induced apoptotic cells from 46.2 to 28.2% (Fig. 2B). To further confirm the
anti-apoptotic effect of curcumin, we performed western blot
analysis. AGEs injury is associated with upregulation of Bax and
AIF, leading to caspase cascade activation (caspase-3), However,
co-incubation of NRK-52E cells with curcumin significantly reduced
AGEs-induced apoptosis-related proteins (Fig. 2C-E). These results indicate that
curcumin activates cytoprotective effect in AGEs induced
apoptosis.
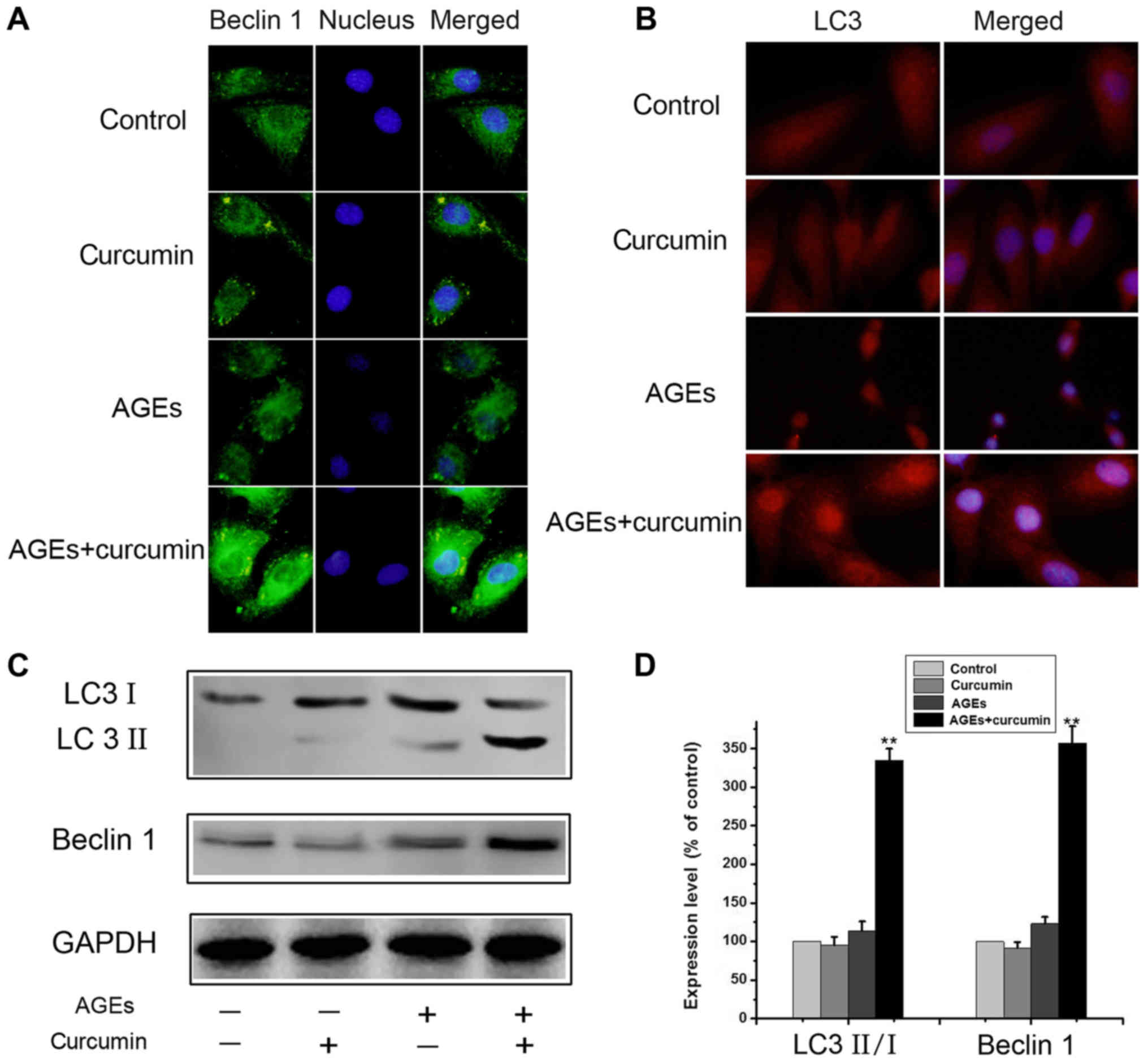
Curcumin enhances autophagy in NRK-52E
cells
Apoptosis and autophagy are two evolutionarily
conserved processes that maintain homeostasis during stress. The
crosstalk between apoptosis and autophagy is complex, as autophagy
can function to promote cell survival under various cellular
conditions. In this text, we explored the role of autophagy in the
cytoprotective effect of curcumin. As above, the NRK-52E cells were
treated with 700 µg/ml AGEs with or without 10 µM curcumin for 48
h. After both curcumin and AGEs treatment, the number of LC3 and
Beclin 1 fluorescent dots dramatically increased (Fig. 3A and B), suggesting that autophagic
vacuolization occurs in response to curcumin treatment, compared to
the AGEs alone group, From the results of fluorescence microscope.
However, treatment of curcumin alone did not increase autophagy in
NRK-52E cells (Fig. 3A and B).
Similar changes were observed on western blot results. The
expressions of Beclin 1 and the ratio of LC3II and LC3I were
significantly increased in AGEs + curcumin group compared to AGEs
group (P<0.01) (Fig. 3C and D).
Theses results indicate that curcumin promotes autophagy in NRK-52E
cells.
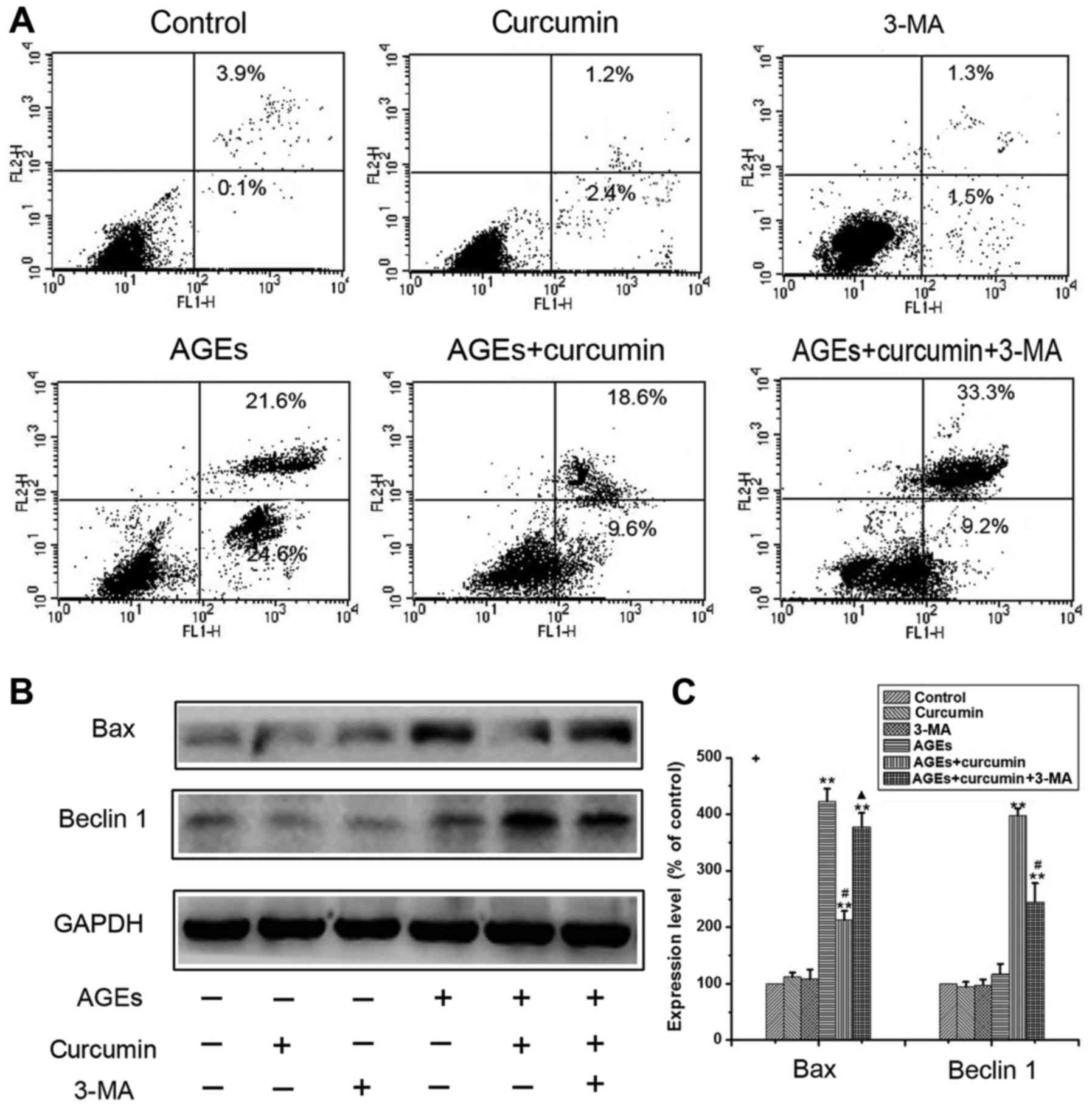
Curcumin inhibits apoptosis through
protective autophagy
To investigate whether the mechanism of curcumin
anti-apoptotic effects was the induction of chondrocyte autophagy,
3-Methyladenine (3-MA), a type of potent pharmacological inhibitors
on autophagy, was used to suppress the autophagy. As (Fig. 4A) demonstrated that pretreatment with
2 mM 3-MA could block autophagy in NRK-52E cells without
significant cytotoxicity. From the results of (Fig. 4A), curcumin significantly inhibited
AGEs-induced apoptosis, however, the protective role of curcumin
was greatly reduced when co-cultured with autophagy inhibitor 3-MA.
These results show that autophagy is an important pathway for a
protective role of curcumin. In order to further validate this
conclusion, we further carried out western blot experiment. The
western blot analysis offered another evidence. The upregulation of
the expression of apoptosic protein AIF, Bax and cleaved caspase-3
were decreased when cells were co-treated with curcumin but this
effect was attenuated by co-treatment with 3-MA (Fig. 4B and C). These results indicated that
autophagy induced by curcumin serve in a protective manner, and
blockage of autophagy interrupted the protection effect of curcumin
in NRK-52E cells.
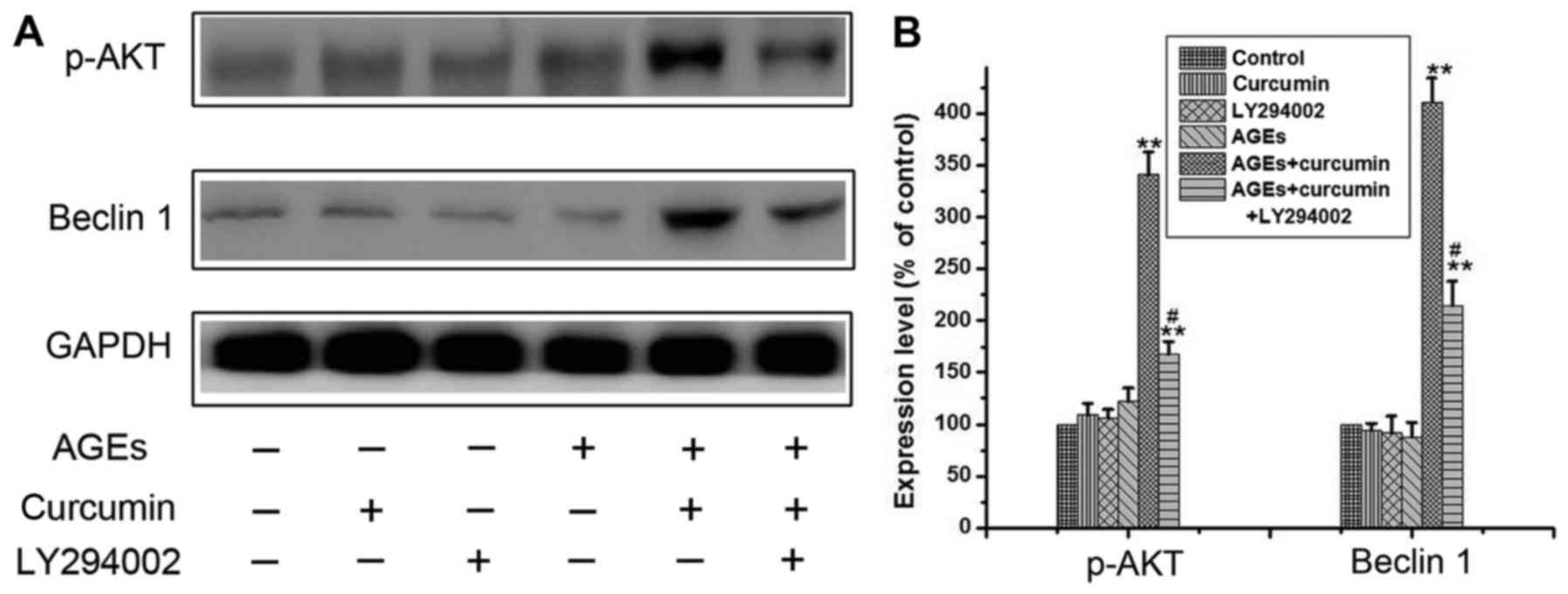
Curcumin upregulates autophagy through
PI3k/AKT pathway
Given that PI3K/AKT pathway activation promotes
autophagy in different cell lines (14). To investigate the involvement of
PI3k/AKT pathway in curcumin-induced autophagy, 10 µM LY294002 (the
inhibitor of PI3K/AKT) was added to the medium of NRK-52E cells. As
the results of Fig. 5A and B,
preincubation of the cells with both curcumin and AGEs activated
the PI3k/AKT signaling pathway and significantly increased the
expression of Beclin 1. However, the expression of Beclin 1 was
greatly reduced when co-cultured with PI3k/AKT inhibitor LY294002.
These results show that curcumin promotes autophagy through
PI3K/AKT signaling pathway.
Discussion
Curcumin as a kind of plant polyphenol extracted
from turmeric is the most important active ingredient of turmeric
to play the pharmacological effect (15). Curcumin exhibits a variety of
biological activities such as anti-inflammatory, antioxidation,
anti-human immunodeficiency virus, anti-fibrosis and anticancer
effects (16–18). During recent years, the defensive
role of curcumin against apoptosis has been experimentally
documented (19). To investigate
whether curcumin could protect rat's kidney against diabetes
damage, we assessed NRK-52E cells for apoptosis, autophagy and
apoptotic factor activities. Firstly, our study confirmed that
curcumin inhibited AGE-induced apoptosis and downregulated the
expression of Bax, AIF and caspase-3. This results obtained from
this study suggested that curcumin could reduce cytotoxic response
of AGEs. Furthermore, we observed that the NRK-52E cells were
treated with curcumin showed a significant activation of protective
autophagy and upregulated the level of Beclin 1 and the ratio of
LC3II and LC3I. Previous study found that curcumin could protect
cells against oxidative stress-induced damage through inhibiting
apoptosis and inducing autophagy (20). Our findings also confirm this
conclusion. However, the role of autophagy in this process has not
been reported.
The relationship between autophagy and apoptosis is
complex and can be induced by both processes in response to similar
stimuli, through which superfluous, damaged, or aged cells or
organelles are eliminated (21).
Autophagy as a double-edged sword is evolutionarily conserved
mechanism for maintaining cellular homeostasis and plays a positive
and negative role in promoting apoptosis (22,23).
Previous study highlighted that autophagy played protective roles
in the prevention of diabetes through their ability to provide an
adaptive response to insulin resistant states (24). Moreover, study also revealed that
curcumin caused some novel cellular mechanisms to promote autophagy
as a protective effect (25). Our
findings also confirm this conclusion that curcumin both greatly
attenuates AGE-induced apoptosis by activating autophagy and
increases autophagy by activation of PI3K/AKT signaling pathway (a
classical autophagic pathway).
In conclusion, we have shown the protective
potential of curcumin for AGEs-mediated apoptosis. Autophagy caused
by the activation of PI3K/AKT pathway plays a significant role in
this process. Although the present study is limited to in
vitro studies, it suggests that curcumin may control autophagy
and play a protective role to resistance to diabetic kidney damage.
The results also suggest that the controlling autophagy may have
the therapeutic potential to inhibit cell apoptosis and prevent
diabetic nephropathy.
Acknowledgements
This study was supported by The International
Science and Technology Cooperation Program of China (2010DFB33260;
2010DFB33260).
Glossary
Abbreviations
Abbreviations:
|
NRK-52E cells
|
normal rat kidney tubular epithelial
cells
|
|
EMT
|
epithelial-to-mesenchymal
transition
|
|
MTT
|
3-[4,5-dimethylthiazol-2-y]-2,5-diphenyltetrazolium bromide
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
DMSO
|
dimethyl sulfoxide
|
|
BSA
|
bovine serum albumin
|
|
PBS
|
phosphate-buffered saline
|
|
TBS
|
Tris-buffered saline
|
|
DN
|
diabetic nephropathy
|
|
AGEs
|
advanced glycation or glycoxidation
end-products
|
|
DM
|
diabetes mellitus
|
|
3-MA
|
3-methyladenine
|
|
FBS
|
fetal bovine serum
|
|
TUNEL
|
the terminal uridine nick 3′ end
labeling
|
References
|
1
|
Yamamoto Y, Kato I, Doi T, Yonekura H,
Ohashi S, Takeuchi M, Watanabe T, Yamagishi S, Sakurai S, Takasawa
S, et al: Development and prevention of advanced diabetic
nephropathy in RAGE-overexpressing mice. J Clin Invest.
108:261–268. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Elsherbiny NM and Al-Gayyar MM: The role
of IL-18 in type 1 diabetic nephropathy: The problem and future
treatment. Cytokine. 81:15–22. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kalia K, Sharma S and Mistry K:
Non-enzymatic glycosylation of immunoglobulins in diabetic
nephropathy. Clin Chim Acta. 347:169–176. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hashimoto Y, Yamagishi S, Mizukami H,
Yabe-Nishimura C, Lim SW, Kwon HM and Yagihashi S: Polyol pathway
and diabetic nephropathy revisited: Early tubular cell changes and
glomerulopathy in diabetic mice overexpressing human aldose
reductase. J Diabetes Investig. 2:111–122. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Teng B, Duong M, Tossidou I, Yu X and
Schiffer M: Role of protein kinase C in podocytes and development
of glomerular damage in diabetic nephropathy. Front Endocrinol
(Lausanne). 5:1792014.PubMed/NCBI
|
|
6
|
Fukami K, Yamagishi S, Ueda S and Okuda S:
Role of AGEs in diabetic nephropathy. Curr Pharm Des. 14:946–952.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kumar Pasupulati A, Chitra PS and Reddy
GB: Advanced glycation end products mediated cellular and molecular
events in the pathology of diabetic nephropathy. Biomol Concepts.
7:293–309. 2016.PubMed/NCBI
|
|
8
|
Kishore L, Kaur N and Singh R:
Renoprotective effect of Bacopa monnieri via inhibition of advanced
glycation end products and oxidative stress in
STZ-nicotinamide-induced diabetic nephropathy. Ren Fail.
38:1528–1544. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen C, Huang K, Hao J, Huang J, Yang Z,
Xiong F, Liu P and Huang H: Polydatin attenuates AGEs-induced
upregulation of fibronectin and ICAM-1 in rat glomerular mesangial
cells and db/db diabetic mice kidneys by inhibiting the activation
of the SphK1-S1P signaling pathway. Mol Cell Endocrinol. 427:45–56.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maeda S, Matsui T, Takeuchi M and
Yamagishi S: Sodium-glucose cotransporter 2-mediated oxidative
stress augments advanced glycation end products-induced tubular
cell apoptosis. Diabetes Metab Res Rev. 29:406–412. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qi W, Niu J, Qin Q, Qiao Z and Gu Y:
Glycated albumin triggers fibrosis and apoptosis via an NADPH
oxidase/Nox4-MAPK pathway-dependent mechanism in renal proximal
tubular cells. Mol Cell Endocrinol. 405:74–83. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Masini M, Bugliani M, Lupi R, del Guerra
S, Boggi U, Filipponi F, Marselli L, Masiello P and Marchetti P:
Autophagy in human type 2 diabetes pancreatic beta cells.
Diabetologia. 52:1083–1086. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jaroonwitchawan T, Chaicharoenaudomrung N,
Namkaew J and Noisa P: Curcumin attenuates paraquat-induced cell
death in human neuroblastoma cells through modulating oxidative
stress and autophagy. Neurosci Lett. 636:40–47. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zeng Y, Yang X, Wang J, Fan J, Kong Q and
Yu X: Aristolochic acid I induced autophagy extenuates cell
apoptosis via ERK 1/2 pathway in renal tubular epithelial cells.
PLoS One. 7:e303122012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Araújo CC and Leon LL: Biological
activities of Curcuma longa L. Mem Inst Oswaldo Cruz. 96:723–728.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Menon VP and Sudheer AR: Antioxidant and
anti-inflammatory properties of curcumin. Adv Exp Med Biol.
595:105–125. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu PY: Curcumin: Another potential
translational candidate for anti-fibrosis on heart failure? Acta
Cardiol Sin. 30:483–484. 2014.PubMed/NCBI
|
|
18
|
Sufi SA, Adigopula LN, Syed SB, Mukherjee
V, Coumar MS, Rao HS and Rajagopalan R: In-silico and in-vitro
anti-cancer potential of a curcumin analogue (1E, 6E)-1, 7-di
(1H-indol-3-yl) hepta-1, 6-diene-3, 5-dione. Biomed Pharmacother.
85:389–398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo S, Long M, Li X, Zhu S, Zhang M and
Yang Z: Curcumin activates autophagy and attenuates oxidative
damage in EA.hy926 cells via the Akt/mTOR pathway. Mol Med Rep.
13:2187–2193. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Delou JM, Biasoli D and Borges HL: The
complex link between apoptosis and autophagy: A promising new role
for RB. An Acad Bras Cienc. 88:2257–2275. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mizushima N: Autophagy and apoptosis.
Nihon Rinsho Meneki Gakkai Kaishi. 23:527–530. 2000.(In Japanese).
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nie C, Zhou J, Qin X, Shi X, Zeng Q, Liu
J, Yan S and Zhang L: Reduction of apoptosis by
proanthocyanidin-induced autophagy in the human gastric cancer cell
line MGC-803. Oncol Rep. 35:649–658. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim KY, Kim SH, Yu SN, Park SG, Kim YW,
Nam HW, An HH, Yu HS, Kim YW, Ji JH, et al: Lasalocid induces
cytotoxic apoptosis and cytoprotective autophagy through reactive
oxygen species in human prostate cancer PC-3 cells. Biomed
Pharmacother. 88:1016–1024. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fujitani Y, Kawamori R and Watada H: The
role of autophagy in pancreatic beta-cell and diabetes. Autophagy.
5:280–282. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Han J, Pan XY, Xu Y, Xiao Y, An Y, Tie L,
Pan Y and Li XJ: Curcumin induces autophagy to protect vascular
endothelial cell survival from oxidative stress damage. Autophagy.
8:812–825. 2012. View Article : Google Scholar : PubMed/NCBI
|