Introduction
For several thousand years, Ganoderma
(G.) lucidum (Ling-Zhi in Chinese and Reishi in
Japanese) has been widely used as a traditional medication for the
prevention and treatment of various human diseases in Asia
(1). The major biologically active
components of G. lucidum are polysaccharides and the
secondary metabolites ganoderic acids (GAs), including GA-A, -B,
-C, -D, -E and -F. Most GAs have a significant pharmacological
potential; they were demonstrated to exhibit significant medicinal
value with activities including inhibition of histamine release and
cholesterol synthesis as well as antitumor and anti-inflammatory
effects (2).
It was demonstrated that inflammation participates
in the mediation of acute and chronic neurological disorders,
including epilepsy and seizure (3).
In human epilepsy patients and in experimental models of epilepsy,
inflammatory processes, including activation of microglia and
release of proinflammatory cytokines, have been described (4–6).
Emerging evidence thus supports the hypothesis that inflammation
may contribute to epileptogenesis. Microglia are intimately
associated with diverse neuronal functions, such as modulation of
synaptic function and plasticity, regulation of the delivery of
energy substrates and enforcement of cellular immunity in the brain
to restore function and promote healing, which helps to maintain
tissue homeostasis (7). However,
dysregulation of microglial functions may cause seizures or promote
epileptogenesis. Activation of microglia through increases of
excitability and inflammation is a prominent feature of epileptic
foci in the human brain and in experimental epilepsy models
(8–10). Uncontrolled microglia-mediated
immunity may cause sustained release of inflammatory cytokines,
including interleukin (IL)-1β, IL-6 and tumor necrosis factor
(TNF)-α, to facilitate epileptogenesis (11).
GAs, the triterpenoid components of G.
lucidum mushroom extracts, are attractive sources of
anti-inflammation products. Although numerous studies have reported
that GAs exhibit anti-inflammatory and antitumor properties in
animal models mainly through the induction of cytokines such as
IL-1, IL-6, interferon-γ and TNF-α in monocytes/macrophages and T
lymphocytes (12–14), encouraging the potential use of GAs
in combination therapy against inflammation and cancer, little
information is available on their in vitro and in
vivo effect on microglia-mediated inflammation. The present
study aimed to investigate the role of GA-A on microglia-mediated
inflammation in vitro. In addition, the effect of GA-A on
lipopolysaccharide (LPS)-evoked alterations in mitochondrial
metabolic activity of microglia was evaluated.
Materials and methods
Reagents and chemicals
GA-A, poly-L-ornithine hydrobromide, dimethyl
sulphoxide (DMSO), deoxyribonuclease I from bovine pancreas
(DNaseI), penicillin-streptomycin, MTT and LPS were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Mouse monoclonal
anti-CD68 antibody conjugated to phycoerythrin (PE) was purchased
from eBioscience (San Diego, CA, USA; cat. no. 12-0681-82). Mouse
monoclonal anti-CD11b antibody conjugated to fluorescein
isothiocyanate was purchased from ProSpec-Tany Technogene Ltd.
(East Brunswick, NJ, USA; cat. no. ANT-136). High-glucose
Dulbecco's modified Eagle's medium (DMEM), PBS, 0.05% (w/v)
trypsin-EDTA and fetal bovine serum (FBS) were obtained from Thermo
Fisher Scientific, Inc. (Waltham, MA, USA).
Isolation and culture of primary
microglia
The primary microglia cultures were established as
follows: Mixed glial cultures from male C57BL/6 mice, purchased
from the Animal Centre of the Xiangya Third Hospital (Changsha,
China) were established from neonatal cortices (postnatal day 0–1;
n=5; mean weight, 1.4 g) (15).
Cells were cultured in high-glucose DMEM supplemented with 10% FBS,
penicillin and streptomycin in a humidified atmosphere of 95% air
and 5% CO2 at 37°C. The culture medium was replaced with
fresh medium 24 h after the initial preparation and every 3 days
thereafter. Following 1 week of culture, microglia were obtained by
mechanical shaking of the mixed glial cell cultures for 1 h. Cells
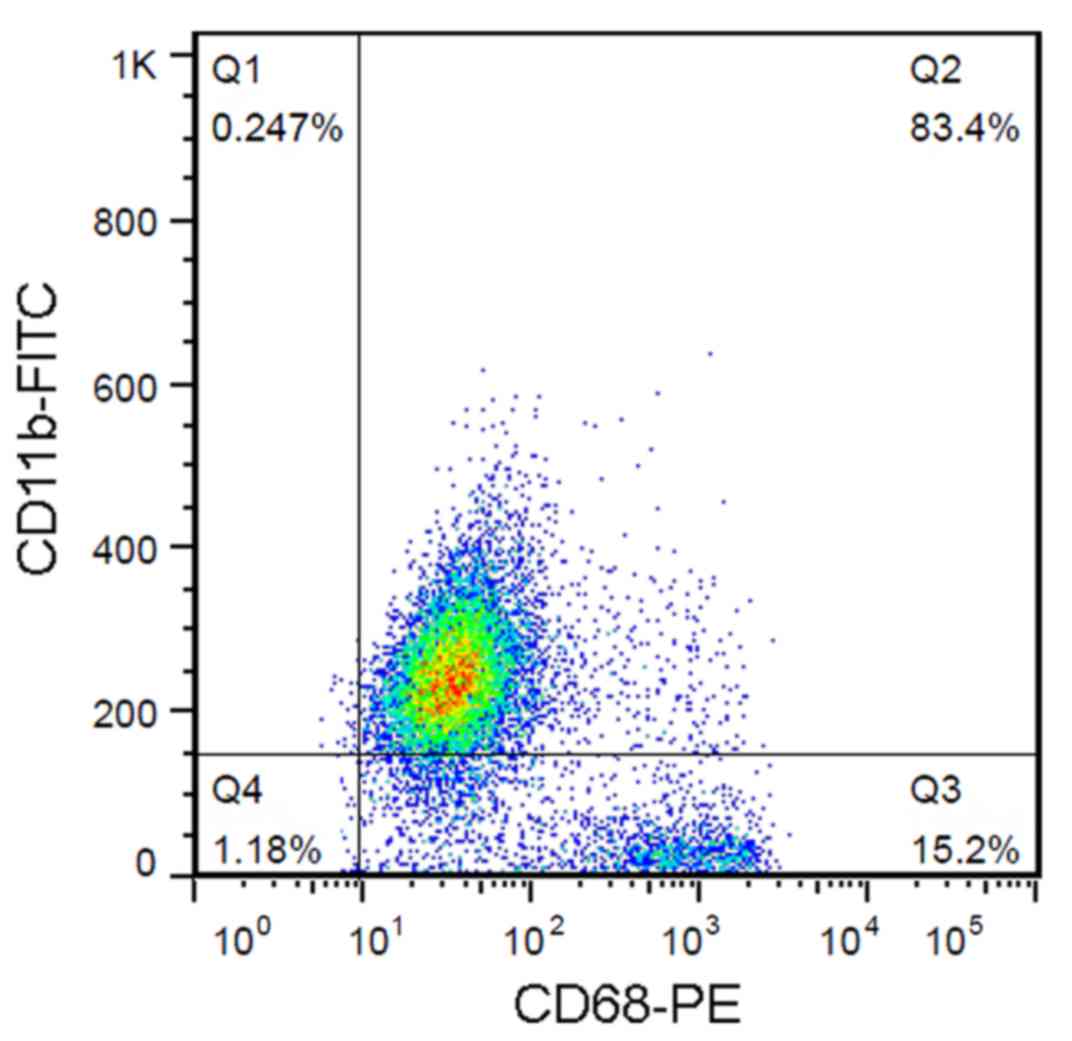
were routinely monitored for purity by fluorescence-activated cell
sorting and the population of CD11b+ CD68+
cells was >80% (Fig. 1). All
experimental procedures were approved by the Xiangya Third Hospital
Ethics Committee for Experimentation on Animals (Changsha, China),
where the cultures were established.
Cell treatment
To induce the release of proinflammatory cytokines,
microglia were treated with DMEM containing 0.1 µg/ml LPS or
vehicle for 24 h. To investigate the role of GA-A in LPS-induced
release of cytokines, microglial cells were treated with a range of
concentrations of GA-A (10, 20, 50 or 100 µg/ml), or co-treated
with LPS. Cells treated with vehicle were used as a control.
RNA extraction and SYBR green
quantitative polymerase chain reaction (qPCR) analysis
Total RNA was extracted from cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
RevertAid™ First Strand cDNA Synthesis kit (Thermo
Fisher Scientific, Inc.) was used to reverse transcribe the mRNA to
cDNA according to the manufacturer's protocol. Briefly, 0.5 ng of
the template RNA, 1 µl Oligo(dT)18 Primer and 12 µl RNase free
water were mixed, and incubated at 65°C for 5 min, then cooled down
on ice. The mixture was added to Reaction Buffer (4 µl), RiboLock
RNase inhibitor (1 µl), 10 mM dNTP mix (2 µl) and RevertAid Reverse
Transcriptase (1 µl), and incubated at 42°C for 1 h. Finally, the
mixture was heated to 70°C for 5 min to obtain the cDNA. The
expression of IL-1β, IL-6 and TNF-α in cells was detected using a
SYBR Fast qPCR mix (cat. no. RR430A; Takara, Dalian, China) at
following condition: 95°C for 30 sec, followed by 40 cycles of 95°C
for 5 sec and 60°C for 10 sec. Expression of β-actin was assessed
as an endogenous control. The mRNA levels were quantified using the
2−ΔΔCq method (16).
Measurement of the release of IL-1β,
IL-6 and TNF-α
Levels of IL-1β, IL-6 and TNF-α were measured in
supernatants of the primary mouse microglia cell culture with
commercially available ELISA kits (Sigma-Aldrich; Merck KGaA)
according to the manufacturer's instructions. Measurements were
performed in three independent experiments.
Western blot analysis
Immunoblotting was performed to detect the
expression of IL-1β, IL-6 and TNF-α in the cell lysate, as well as
the levels of nuclear factor (NF)-κB and phosphorylated inhibitor
of NF-κB (p-IκBα) in microglial cells. Cultured or transfected
cells were lysed in radioimmunoprecipitation assay buffer with 1%
phenylmethane sulfonyl fluoride. The concentration of protein was
determined using the BCA Protein Assay kit (cat. no. P0011;
Beyotime Institute of Biotechnology, Haimen, China) according to
the manufacturer's protocol. A total of 60 µg/lane of protein was
loaded into a 12% SDS-PAGE minigel, electrophoresed and transferred
onto a polyvinylidene difluoride membrane (Wuhan Boster Biological
Technology, Ltd., Wuhan, China). The membranes were probed with
antibodies directed against IL-1β (cat. no. I3767; 1:1,000), IL-6
(cat. no. SAB4301665; 1:1,000) and TNF-α (cat. no. SAB4502982;
1:2,000) from Sigma-Aldrich (Merck KGaA); p-IκBα (cat. no. 9246S;
1:1,000) and NF-κB (cat. no. 8242; 1:3,000) from Cell Signaling
Technology, Inc. (Danvers, MA, USA); GAPDH (cat. no. ab9485;
1:3,000) from Abcam (Cambridge, UK) at 4°C overnight. The blots
were subsequently incubated with horseradish peroxidase-conjugated
secondary antibodies (cat. no. ab7090 and ab97040; 1:5,000; Abcam)
for 1 h at 37°C. Signals were visualized using enhanced
chemiluminescence substrate (EMD Millipore, Billerica, MA, USA).
GAPDH was used as an endogenous protein for normalization.
Measurement of the mitochondrial
metabolic activity
The mitochondrial activity of microglial cells was
determined by measurement of MTT reduction to MTT formazan by
cellular mitochondrial dehydrogenases. After incubation with GA-A
for 24 h, MTT (0.5 mg/ml) was added to the medium, followed by
incubation for 4 h at 37°C. Subsequently, dimethyl sulfoxide (150
µl) was added to dissolve the formazan crystals and the absorbance
was measured at 570 nm using a microplate reader (Paradigm
Detection Platform; Beckman Coulter, Brea, CA, USA); this value was
proportional to the number of viable cells with intact
mitochondria. Measurements were performed in three independent
experiments.
Statistical analysis
Values are expressed as the mean ± standard error.
Statistical analysis was performed using SPSS software (version 20;
IBM Corp., Armonk, NY, USA). Student's t-test was performed for
two-group comparisons, while one-way analysis of variance with
Tukey's post-hoc test was performed for multiple-group comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of GA-A on LPS-induced release
of IL-1β, IL-6 and TNF-α from cortical microglial cells in
culture
To investigate the role of GA-A on LPS-induced
release of IL-1β, IL-6 and TNF-α from mouse cortical microglial
cells, the cells were treated with 0.1 µg/ml LPS for 24 h.
Treatment with LPS resulted in a potent, 80-, 42- and 110-fold
increase in IL-1β, IL-6 and TNF-α release, respectively (Fig. 2A). In addition, the microglial cells
were treated with a range of concentrations of GA-A (10, 20, 50 and
100 µg/ml) for 24 h. The results indicated that GA-A treatment
slightly but not significantly altered the release of IL-1β, IL-6
and TNF-α (Fig. 2B-D). GA-A
increased IL-1β at 20 µg/ml, and IL-6 at 10 and 20 µg/ml; GA-A at
100 µg/ml decreased the release of IL-1β, IL-6 and TNF-α to ~75% of
that in the control group, but this effect was not statistically
significant (Fig. 2B-D).
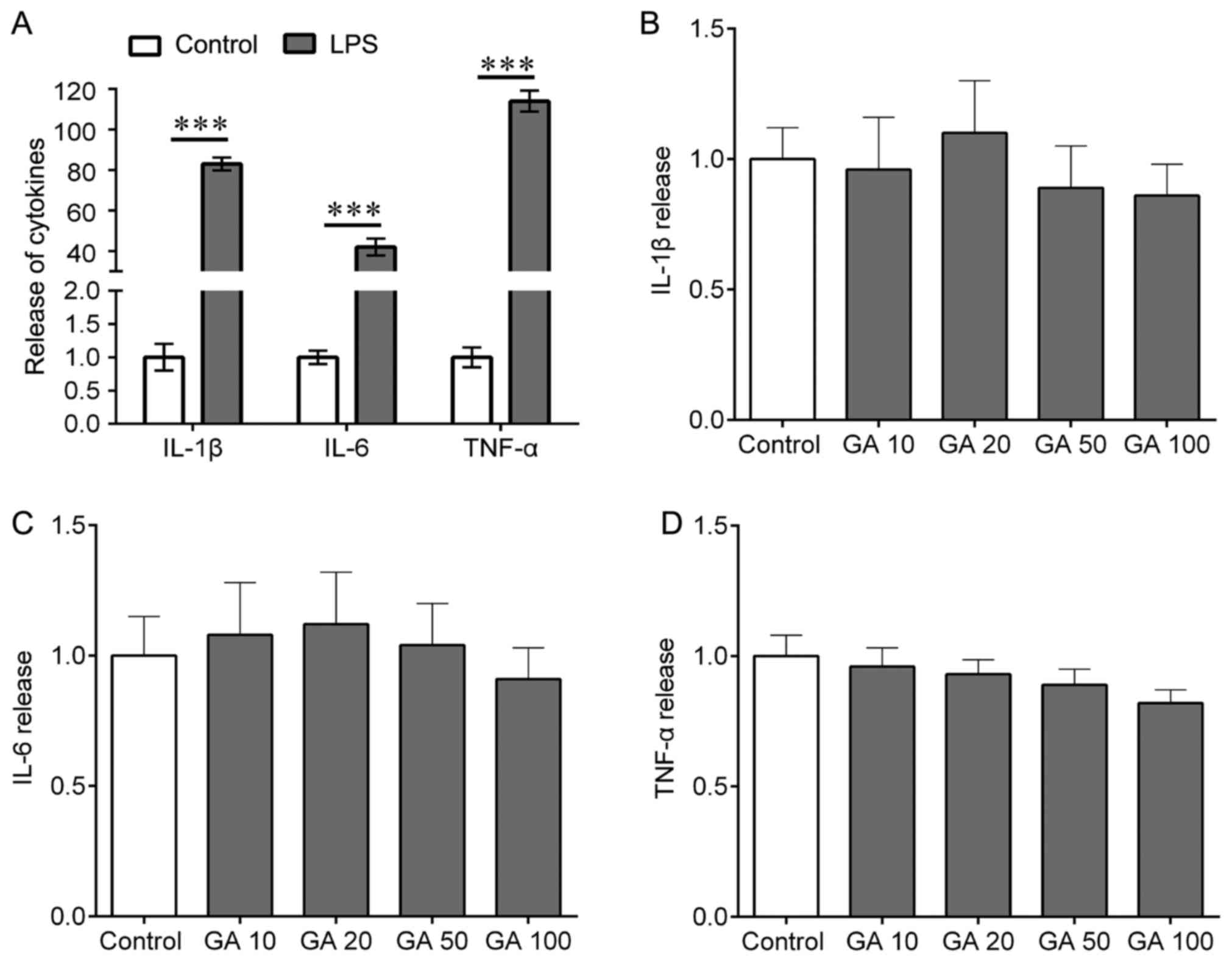 | Figure 2.Effects of LPS on the release of
IL-1β, IL-6 and TNF-α from mouse cortical microglial cells in
culture. (A) The cells were treated with LPS (0.1 µg/ml) for 24 h,
and ELISA were used to determine the release of IL-1β, IL-6 and
TNF-α. (B-D) The cells were treated with GA (10, 20, 50 or 100
µg/ml) for 24 h and ELISA was used to determine the release of (B)
IL-1β, (C) IL-6 and (D) TNF-α. Values are expressed as the mean ±
standard error of the mean and relative to the control. Three
independent experiments were performed. ***P<0.001 vs. control.
IL, interleukin; TNF, tumor necrosis factor; LPS,
lipopolysaccharide; GA, ganoderic acid A. |
Furthermore, it was determined whether GA-A
treatment was able to abolish the LPS-induced release of IL-1β,
IL-6 and TNF-α from mouse cortical microglial cells. First, RT-qPCR
was performed to detect whether the levels of IL-1β, IL-6 and TNF-α
were affected at the transcriptional level, revealing that GA-A
treatment (50 µg/ml) did not significantly alter the cellular mRNA
expression of IL-1β, IL-6 and TNF-α (Fig. 3A). However, ELISAs indicated that
GA-A treatment (10, 20, 50 or 100 µg/ml) for 24 h caused a
statistically significant reduction of LPS-stimulated release of
IL-1β from primary mouse microglial cells in a
concentration-dependent manner (Fig.
3B). At lower concentrations (10 µg/ml), the drug did not cause
any significant decrease of IL-6 and TNF-α release (Fig. 3C and D). However, IL-6 and TNF-α
release were markedly reduced at higher concentrations of the drug
(50 and 100 µg/ml; Fig. 3C and D).
The most prominent effect was observed at a dose of 100 µg/ml (a
decrease by 30, 32 and 22% for IL-1β, IL-6 and TNF-α,
respectively). These results were confirmed by western blot
analysis for IL-1β, IL-6 and TNF-α expression in cell lysate
(Fig. 3E).
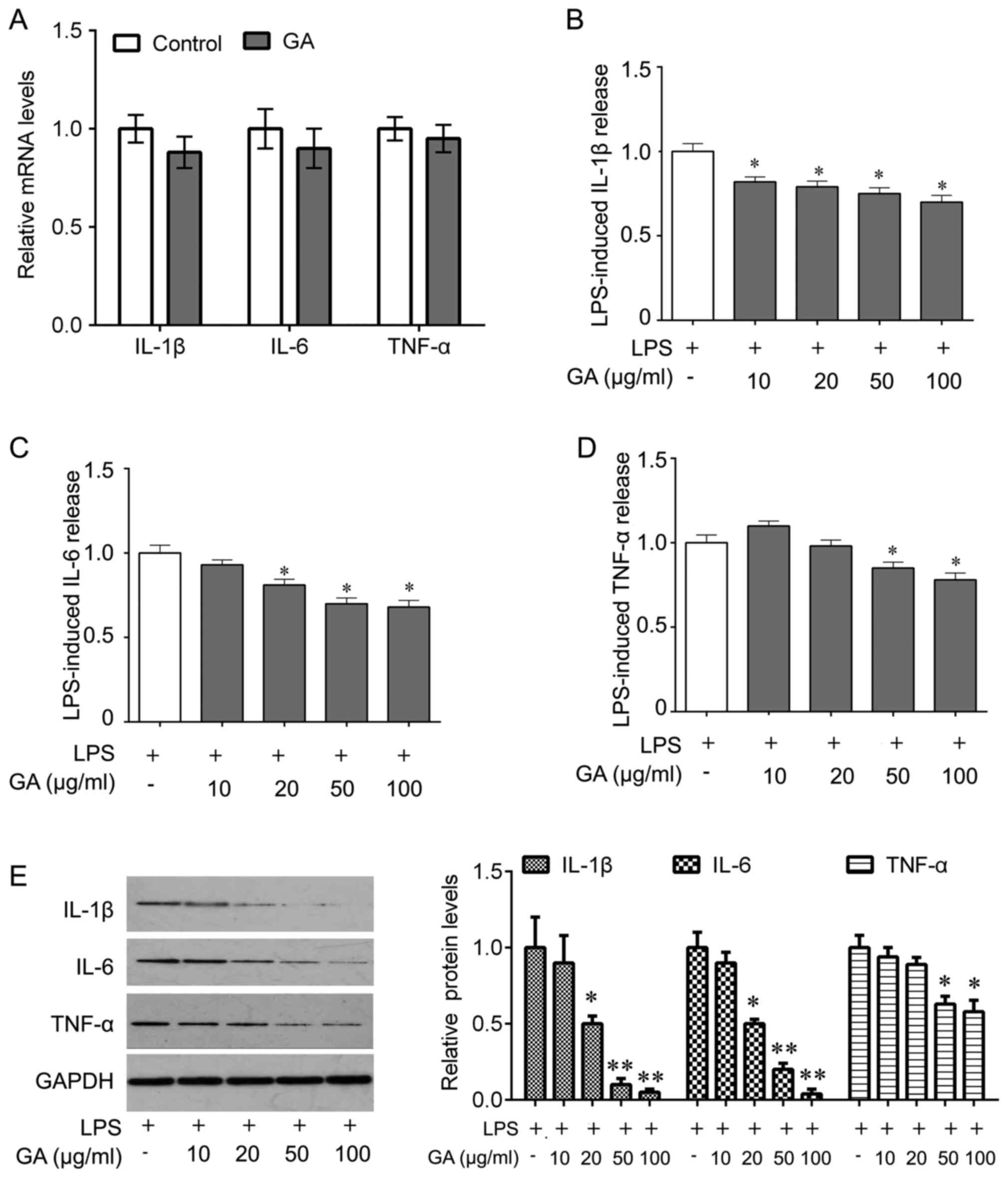 | Figure 3.Effects of GA on LPS-induced release
of IL-1β, IL-6 and TNF-α from mouse cortical microglial cells in
culture. (A) The cells were treated with GA (50 µg/ml) for 24 h,
and total RNA was then extracted. Quantitative polymerase chain
reaction was used for analysis of the mRNA levels of IL-1β, IL-6
and TNF-α. (B-D) The cells were treated with LPS alone or
co-treated with LPS and GA (10, 20, 50 and 100 µg/ml) for 24 h, and
ELISA was used to determine the release of (B) IL-1β, (C) IL-6 and
(D) TNF-α. (E) Western blot was performed to analyze the expression
of IL-1β, IL-6 and TNF-α in cell lysate after the indicated
treatments (left) and quantification (right). Values are expressed
as the mean ± standard error of the mean and relative to the
control that was treated with LPS alone. Three independent
experiments were performed. *P<0.05, **P<0.01 vs. control.
IL, interleukin; TNF, tumor necrosis factor; LPS,
lipopolysaccharide; GA, ganoderic acid A. |
GA-A inhibits LPS-induced NF-κB
pathway activation
Microglial cells were then treated with GA-A (0, 10
or 50 µg/ml) and stimulated with LPS (0.1 µg/ml). Western blot
analysis of the total protein extract indicated that treatment with
GA-A at 10 and 50 µg/ml reduced LPS-induced p-IκBα and NF-κB (p65)
expression, with 50 µg/ml being more effective (Fig. 4A). These results revealed that GA-A
inhibited LPS-induced NF-κB pathway activation, which may suggest
that that non-toxic suppression of IL-1β, IL-6 and TNF-α production
by GA-A is, at least in part, due to suppression of the NF-κB
signaling pathway.
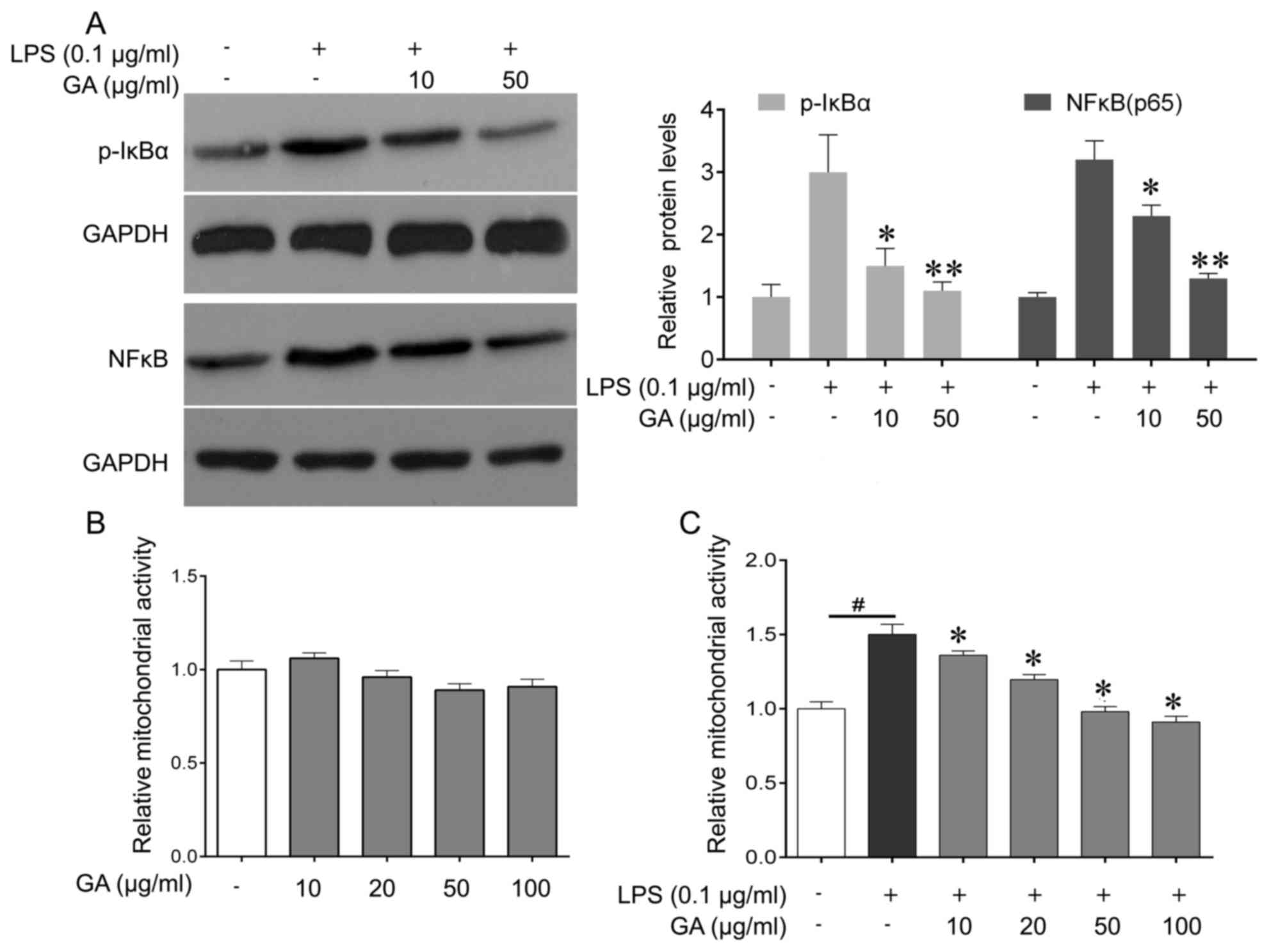 | Figure 4.Effect of GA on LPS (0.1
µg/ml)-induced changes in the NF-κB signaling pathway and
mitochondrial activity of mouse cortical microglial cells in
culture. (A) Western blot was performed to analyze the expression
of p-IκB and p-p65 in microglial cells after the indicated
treatments (left), and quantification (right). (B) The cells were
treated with GA (10, 20, 50, 100 µg/ml), or vehicle (control) for
24 h, and MTT were used to determine the mitochondrial activity of
microglia cells. (C) The cells were treated with LPS alone, or
co-treated with LPS and GA (10, 20, 50 or 100 µg/ml), or vehicle
(control) for 24 h, and the MTT assay was used to determine the
mitochondrial activity of microglial cells. Values are expressed as
the mean ± standard error of the mean and relative to the control
treated with vehicle. Three independent experiments were performed.
#P<0.05 vs. control, *P<0.05, **P<0.01 vs. LPS
alone. GA, ganoderic acid A; LPS, lipopolysaccharide; NF-κB,
nuclear factor κB; p-IκBα, phosphorylated inhibitor of NF-κB. |
GA-A reduces LPS-induced increases in
mitochondrial activity of mouse cortical microglia cells
GA-A treatment (10, 20, 50 or 100 µg/ml for 24 h)
did not significantly alter the mitochondrial activity of
microglial cells (Fig. 4B), while
cells stimulated with LPS (0.1 µg/ml) for 24 h exhibited a
significant increase in their mitochondrial activity by 50%
(Fig. 4C). The increase of
mitochondrial activity induced by LPS was markedly attenuated by
treatment of the cells with GA-A used at 10 and 20 µg/ml, and was
abolished by higher concentrations of the drug (50 and 100 µg/ml;
Fig. 4C).
Discussion
The present study demonstrated, for the first time,
to the best of our knowledge, the effects of GA-A on LPS-stimulated
release of proinflammatory cytokines from primary mouse microglia
cultures. To date, only a few studies have examined the effects of
GA-A on microglia-mediated inflammation (17). Microglial cells are brain-resident
macrophage-like cells that contribute to innate immune mechanisms
(18). Microglia-mediated
inflammation was reported to have an important role in the
pathogenesis of epilepsy (19). In
response to stressors, microglia are activated, leading to the
release of proinflammatory mediators that may promote seizures and
epileptogenesis, particularly when uncontrolled inflammation occurs
(5). Of note, the extent of
microglia activation correlates with the frequency and duration of
seizures (20).
Activated microglial cells release cytokines that
induce transcriptional and post-transcriptional signaling. For
instance, microglia release a variety of pro-inflammatory and
cytotoxic soluble factors, including IL-1β and IL-6, and subsequent
activation of the proinflammatory IL-1 receptor/Toll-like receptor
(IL1R/TLR) system (21). In epilepsy
models, this IL1R/TLR signaling is activated, which promotes the
onset and recurrence of seizures (22). Of note, pharmacological blockade or
genetic inactivation of the IL1R/TLR system drastically reduces
seizure activity (23). IL-1β
increases glutamate release via TNF-α production, resulting in
elevated extracellular glutamate levels and hyperexcitability
(24). Furthermore, IL-1β also
stimulates IL-6 release (25). After
a febrile seizure, children had significantly higher serum IL-6
levels than a healthy control group (26), and IL-6 levels had a descending trend
during the time of recovery from the seizure in the intractable
epilepsy group and the non-intractable epilepsy group (27), indicating that high IL-6 levels may
be a pathogenetic factor in epilepsy.
LPS is a component of the wall of Gram-negative
bacteria, which may induce immediate focal epileptic-type
discharges in the mouse neocortex mediated by IL-1β release
(28). LPS activates microglia via
TLR4. Endogenous ligands of TLR4, including IL-1β, may be generated
by microglia following brain injury, mimicking the effect of LPS
(29). An animal study indicated
that activated microglia release proinflammatory molecules to
decrease the seizure threshold (30). Consequently, microglia may help
generate seizures by releasing, and responding to, endogenous
inflammatory mediators, including IL-1β and TNF-α (31). For instance, chemokine-activated
microglia cooperated with astrocytes to release TNF-α and other
cytokines, thereby contributing to cell loss and seizures (32).
Anti-inflammatory molecules then help to resolve the
inflammatory tissue response. Clinical anti-inflammatory or
immunosuppressive treatments may control seizures in certain
epileptic syndromes. For instance, intravenous immunoglobulin
(IVIG) increases circulating levels of IL-1 receptor antagonist and
blocks IL-1β signaling (33,34). IVIG suppresses seizures, which may be
partially mediated by the reduction of proinflammatory cytokines
and suppression of microglia activation (35).
Numerous studies have indicated that GAs enhance the
immune system. Akihisa et al (36) identified four GAs isolated from the
fruiting bodies of the fungus G. lucidum and identified that
these GAs inhibit 12-O-tetradecanoylphorbol-13-acetate-induced
inflammation in mice. The triterpene extract from G. lucidum
markedly suppressed the secretion of the inflammatory cytokines
TNF-α and IL-6, as well as the inflammatory mediator nitric oxide
from LPS-stimulated murine RAW264.7 cells (37). In addition, natural killer cell
activity was significantly enhanced by intraperitoneal
administration of GA-Me (38). The
anti-inflammatory properties of purified GAs encourage the
potential use of GAs in combination therapy against
inflammation.
The present study demonstrated that GA-A
significantly decreased LPS-induced IL-1β, IL-6 and TNF-α release
from primary mouse cortical microglial cells in a
concentration-dependent manner. Furthermore, GA treatment did not
alter the mRNA expression of IL-1β, IL-6 and TNF-α, but decreased
the release of IL-1β, IL-6 and TNF-α in the cell culture medium,
indicating that GA may target the release of cytokines. LPS induces
proinflammatory cytokines, including IL-1β, IL-6 and TNF-α, through
the activation of several intracellular signaling pathways such as
NF-κB (39). A previous study
demonstrated that G. lucidum extracts significantly and
non-toxically suppressed TNF-α production by murine macrophages
induced by peripheral blood mononuclear cells from asthma patients
(12). Furthermore, the inhibitory
effect of G. lucidum extracts on LPS-induced TNF-α
production by macrophages was associated with the suppression of
NF-κB signaling. In line with these previous studies, the results
of the present study indicated that GA treatment reduced
LPS-induced p-IκBα and NF-κB (p65) expression, suggesting that
non-toxic suppression of IL-1β, IL-6 and TNF-α production by GA is,
at least in part, due to suppression of NF-κB signaling. Although
the results of the present study indicate GA regulates the IL-1β,
IL-6 and TNF-α production through NF-κB at transcriptional level,
other mechanism by which GA control the cytokines protein
production cannot be excluded, such as the ubiquitin pathway
(40).
Since the alterations of microglial function have an
impact on neuronal excitability and epileptic activity, the present
study also examined the effect of GA-A on the metabolic activity in
mitochondria. In line with other studies (41,42), the
results of the present study indicated that LPS stimulation
resulted in significant increase in mitochondrial activity of
microglial cells, and this effect was abolished by co-treatment
with GA-A. A previous study identified that GA-T treatment resulted
in a reduction of mitochondrial membrane potential and release of
cytochrome C, as well as subsequent apoptosis in lung cancer
cells, suggesting that the apoptosis induction of GA-T may be
mediated by mitochondrial dysfunction (43). However, incubation in
GAC2-conditioned media attenuated mitochondrial defects
in a 3-nitropropionic acid (3-NP) cell model, and G. lucidum
treatment therapeutically restored neuronal loss in mice with
3-NP-induced behavioral impairment and striatal degeneration
(44). Whether the effects of GA-A
on LPS-induced changes in mitochondrial activity of microglial
cells are involved in the drug's action on the release of
proinflammatory cytokines remains to be elucidated.
In conclusion, the results of the present study
demonstrated that GA-A reduced the LPS-induced release of
proinflammatory cytokines, IL-1β, IL-6 and TNF-α from mouse
cortical microglial cells in culture at least in part via
suppression of the NF-κB signaling pathway, and suppressed the
LPS-stimulated increase in mitochondrial metabolic activity of the
cells. Thus, treatment with GA-A may be potential therapeutic
strategy for epilepsy via suppression of microglia-derived
proinflammatory mediators.
Acknowledgements
This study was supported by the Natural Science
Foundation of Heilongjiang province, China (grant no. B2015013) and
the Youth Fund of Jiamusi University (grant no. Sq2013-025).
References
|
1
|
Bishop KS, Kao CH, Xu Y, Glucina MP,
Paterson RR and Ferguson LR: From 2000 years of Ganoderma lucidum
to recent developments in nutraceuticals. Phytochemistry.
114:56–65. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferreira IC, Heleno SA, Reis FS, Stojkovic
D, Queiroz MJ, Vasconcelos MH and Sokovic M: Chemical features of
Ganoderma polysaccharides with antioxidant, antitumor and
antimicrobial activities. Phytochemistry. 114:38–55. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dey A, Kang X, Qiu J, Du Y and Jiang J:
Anti-inflammatory small molecules to treat seizures and epilepsy:
From bench to bedside. Trends Pharmacol Sci. 37:463–484. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang B, Zou J, Han L, Rensing N and Wong
M: Microglial activation during epileptogenesis in a mouse model of
tuberous sclerosis complex. Epilepsia. 57:1317–1325. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pfluger P, Viau CM, Coelho VR, Berwig NA,
Staub RB, Pereira P and Saffi J: Gamma-decanolactone inhibits iNOS
and TNF-alpha production by lipopolysaccharide-activated microglia
in N9 cells. Eur J Pharmacol. 780:38–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Avignone E, Lepleux M, Angibaud J and
Nägerl UV: Altered morphological dynamics of activated microglia
after induction of status epilepticus. J Neuroinflammation.
12:2022015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mecha M, Carrillo-Salinas FJ, Feliú A,
Mestre L and Guaza C: Microglia activation states and cannabinoid
system: Therapeutic implications. Pharmacol Ther. 166:40–55. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kiyomoto M, Shinoda M, Honda K, Nakaya Y,
Dezawa K, Katagiri A, Kamakura S, Inoue T and Iwata K: p38
phosphorylation in medullary microglia mediates ectopic orofacial
inflammatory pain in rats. Mol Pain. 11:482015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jebelli J, Su W, Hopkins S, Pocock J and
Garden GA: Glia: Guardians, gluttons, or guides for the maintenance
of neuronal connectivity? Ann N Y Acad Sci. 1351:1–10. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Riazi K, Galic MA, Kuzmiski JB, Ho W,
Sharkey KA and Pittman QJ: Microglial activation and TNFalpha
production mediate altered CNS excitability following peripheral
inflammation. Proc Natl Acad Sci USA. 105:pp. 17151–17156. 2008;
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dambach H, Hinkerohe D, Prochnow N,
Stienen MN, Moinfar Z, Haase CG, Hufnagel A and Faustmann PM: Glia
and epilepsy: Experimental investigation of antiepileptic drugs in
an astroglia/microglia co-culture model of inflammation. Epilepsia.
55:184–192. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu C, Yang N, Song Y, Wang L, Zi J, Zhang
S, Dunkin D, Busse P, Weir D, Tversky J, et al: Ganoderic acid C1
isolated from the anti-asthma formula, ASHMI™ suppresses TNF-α
production by mouse macrophages and peripheral blood mononuclear
cells from asthma patients. Int Immunopharmacol. 27:224–231. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu C, Dunkin D, Lai J, Song Y, Ceballos
C, Benkov K and Li XM: Anti-inflammatory effects of Ganoderma
lucidum triterpenoid in human crohn's disease associated with
downregulation of NF-κB signaling. Inflamm Bowel Dis. 21:1918–1925.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jin X, Ruiz BJ, Sze DM and Chan GC:
Ganoderma lucidum (Reishi mushroom) for cancer treatment. Cochrane
Database Syst Rev. 4:CD0077312016.PubMed/NCBI
|
|
15
|
Gordon R, Hogan CE, Neal ML, Anantharam V,
Kanthasamy AG and Kanthasamy A: A simple magnetic separation method
for high-yield isolation of pure primary microglia. J Neurosci
Methods. 194:287–296. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yoon HM, Jang KJ, Han MS, Jeong JW, Kim
GY, Lee JH and Choi YH: Ganoderma lucidum ethanol extract inhibits
the inflammatory response by suppressing the NF-κB and toll-like
receptor pathways in lipopolysaccharide-stimulated BV2 microglial
cells. Exp Ther Med. 5:957–963. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rangarajan P, Karthikeyan A and Dheen ST:
Role of dietary phenols in mitigating microglia-mediated
neuroinflammation. Neuromolecular Med. 18:453–464. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zattoni M, Mura ML, Deprez F, Schwendener
RA, Engelhardt B, Frei K and Fritschy JM: Brain infiltration of
leukocytes contributes to the pathophysiology of temporal lobe
epilepsy. J Neurosci. 31:4037–4050. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Johnson AM, Sugo E, Barreto D, Hiew CC,
Lawson JA, Connolly AM, Somerville E, Hasic E, Bye AM and
Cunningham AM: The severity of gliosis in hippocampal sclerosis
correlates with pre-operative seizure burden and outcome after
temporal lobectomy. Mol Neurobiol. 53:5446–5456. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vezzani A, Maroso M, Balosso S, Sanchez MA
and Bartfai T: IL-1 receptor/Toll-like receptor signaling in
infection, inflammation, stress and neurodegeneration couples
hyperexcitability and seizures. Brain Behav Immun. 25:1281–1289.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maroso M, Balosso S, Ravizza T, Liu J,
Bianchi ME and Vezzani A: Interleukin-1 type 1 receptor/Toll-like
receptor signalling in epilepsy: The importance of IL-1beta and
high-mobility group box 1. J Intern Med. 270:319–326. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Noe FM, Polascheck N, Frigerio F,
Bankstahl M, Ravizza T, Marchini S, Beltrame L, Banderó CR, Löscher
W and Vezzani A: Pharmacological blockade of IL-1β/IL-1 receptor
type 1 axis during epileptogenesis provides neuroprotection in two
rat models of temporal lobe epilepsy. Neurobiol Dis. 59:183–193.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dolga AM, Granic I, Blank T, Knaus HG,
Spiess J, Luiten PG, Eisel UL and Nijholt IM: TNF-alpha-mediates
neuroprotection against glutamate-induced excitotoxicity via
NF-kappaB-dependent up-regulation of K2.2 channels. J Neurochem.
107:1158–1167. 2008.PubMed/NCBI
|
|
25
|
Hwang JS, Jung EH, Kwon MY and Han IO:
Glioma-secreted soluble factors stimulate microglial activation:
The role of interleukin-1β and tumor necrosis factor-α. J
Neuroimmunol. 298:165–171. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Azab SF, Abdalhady MA, Almalky MA, Amin
EK, Sarhan DT, Elhindawy EM, Allah MA, Elhewala AA, Salam MM,
Hashem MI, et al: Serum and CSF adiponectin, leptin, and
interleukin 6 levels as adipocytokinesin Egyptian children with
febrile seizures: A cross-sectional study. Ital J Pediatr.
42:382016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lei HY, Yang DQ, Li YX, Wang LQ and Zheng
M: Association between human cytomegalovirus and onset of epilepsy.
Int J Clin Exp Med. 8:20556–20564. 2015.PubMed/NCBI
|
|
28
|
Galic MA, Riazi K, Heida JG, Mouihate A,
Fournier NM, Spencer SJ, Kalynchuk LE, Teskey GC and Pittman QJ:
Postnatal inflammation increases seizure susceptibility in adult
rats. J Neurosci. 28:6904–6913. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rodgers KM, Hutchinson MR, Northcutt A,
Maier SF, Watkins LR and Barth DS: The cortical innate immune
response increases local neuronal excitability leading to seizures.
Brain. 132:2478–2486. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li X, Han X, Bao J, Liu Y, Ye A, Thakur M
and Liu H: Nicotine increases eclampsia-like seizure threshold and
attenuates microglial activity in rat hippocampus through the α7
nicotinic acetylcholine receptor. Brain Res. 1642:487–496. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kosonowska E, Janeczko K and Setkowicz Z:
Inflammation induced at different developmental stages affects
differently the range of microglial reactivity and the course of
seizures evoked in the adult rat. Epilepsy Behav. 49:66–70. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang F, Liu J and Shi JS:
Anti-inflammatory activities of resveratrol in the brain: Role of
resveratrol in microglial activation. Eur J Pharmacol. 636:1–7.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cattepoel S, Schaub A, Ender M, Gaida A,
Kropf A, Guggisberg U, Nolte MW, Fabri L, Adlard PA, Finkelstein
DI, et al: Intravenous immunglobulin binds beta amyloid and
modifies its aggregation, neurotoxicity and microglial phagocytosis
in vitro. PLoS One. 8:e631622013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Novak P, Williams A, Ravin P, Zurkiya O,
Abduljalil A and Novak V: Treatment of multiple system atrophy
using intravenous immunoglobulin. BMC Neurol. 12:1312012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Janke AD and Yong VW: Impact of IVIg on
the interaction between activated T cells and microglia. Neurol
Res. 28:270–274. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Akihisa T, Nakamura Y, Tagata M, Tokuda H,
Yasukawa K, Uchiyama E, Suzuki T and Kimura Y: Anti-inflammatory
and anti-tumor-promoting effects of triterpene acids and sterols
from the fungus Ganoderma lucidum. Chem Biodivers. 4:224–231. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dudhgaonkar S, Thyagarajan A and Sliva D:
Suppression of the inflammatory response by triterpenes isolated
from the mushroom Ganoderma lucidum. Int Immunopharmacol.
9:1272–1280. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang G, Zhao J, Liu J, Huang Y, Zhong JJ
and Tang W: Enhancement of IL-2 and IFN-gamma expression and NK
cells activity involved in the anti-tumor effect of ganoderic acid
Me in vivo. Int Immunopharmacol. 7:864–870. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rummel C: Inflammatory transcription
factors as activation markers and functional readouts in
immune-to-brain communication. Brain BehavImmun. 54:1–14. 2016.
|
|
40
|
Lin TY and Hsu HY: Ling Zhi-8 reduces lung
cancer mobility and metastasis through disruption of focal adhesion
and induction of MDM2-mediated Slug degradation. Cancer Lett.
375:340–348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ma B, Yu J, Xie C, Sun L, Lin S, Ding J,
Luo J and Cai H: Toll-like receptors promote mitochondrial
translocation of nuclear transcription factor nuclear factor of
activated T-cells in prolonged microglial activation. J Neurosci.
35:10799–10814. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen H, Wang C, Wei X, Ding X and Ying W:
Malate-aspartate shuttle inhibitor aminooxyacetate acid induces
apoptosis and impairs energy metabolism of both resting microglia
and LPS-activated microglia. Neurochem Res. 40:1311–1318. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tang W, Liu JW, Zhao WM, Wei DZ and Zhong
JJ: Ganoderic acid T from Ganoderma lucidum mycelia induces
mitochondria mediated apoptosis in lung cancer cells. Life Sci.
80:205–211. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen LW, Horng LY, Wu CL, Sung HC and Wu
RT: Activating mitochondrial regulator PGC-1α expression by
astrocytic NGF is a therapeutic strategy for Huntington's disease.
Neuropharmacology. 63:719–732. 2012. View Article : Google Scholar : PubMed/NCBI
|