Introduction
Liver fibrosis is a common pathological occurrence
and increases the risk of cirrhosis, hepatic carcinoma and liver
failure (1). Liver fibrosis may be
triggered by several types of etiologies include viral hepatitis,
chronic alcoholism, nonalcoholic steatohepatitis, toxicants or
drugs, autoimmune liver disease, genetic and metabolic diseases,
and liver congestion (2). It is a
complex disease process that requires various different therapies
for effective treatment (3).
Improved knowledge concerning the initiation and progression of
liver fibrosis has resulted in the development of novel therapies.
Liver transplantation is the only treatment for patients with
cirrhosis and clinical complications (4). Removal of pathogens is the most
effective treatment of liver fibrosis; this strategy has been
proved to be effective in most of the causes of chronic liver
disease (5). Blockade of the
tyrosine kinase appears to be a prospect treatment of liver
fibrosis. Protein tyrosine kinase inhibitors have been considered
to be effective anti-schistosomal and anti-fibrotic drugs, which
inhibit and reverse liver fibrosis induced by Schistosoma
mansoni (6). Vatalanib is a
tyrosine kinase inhibitor that reduces liver fibrosis and sinus
capillary vascularization in CCl4-induced fibrotic mice (7). The use of anti-inflammatory drugs has
been suggested because inflammation precedes and promotes the
progress of liver fibrosis (8).
Glucocorticoids are used only for the treatment of liver fibrosis
in patients with autoimmune hepatitis and acute alcoholic hepatitis
(8). Antioxidants, including vitamin
E, silymarin, phosphatidylcholine and methionine, are beneficial in
the treatment of alcoholic liver disease and non-alcoholic
steatohepatitis (9). However,
monotherapy with tyrosine kinase inhibitors or antioxidants exhibit
limited therapeutic effects (10,11). In
addition, previous results have suggested that the use of any
single pharmacological agent at high doses may induce severe side
effects (12). Subsequently,
research into the identification of novel effective therapies that
treat hepatic fibrosis with fewer side effects is warranted.
Combination therapies involving the use of multiple pharmacological
agents may be superior to monotherapy due to their potential
synergistic effects and limited side effects (13,14).
Combination therapies consist of multiple pharmacological agents
that target various cellular sites of action and may intervene at
different stages of fibrogenesis (15). Therefore, the use of combination
therapies may be an effective therapeutic method of treating liver
fibrosis.
During the course of fibrogenesis, hepatic stellate
cells (HSCs) undergo activation and transdifferentiate into
myofibroblast-like cells that proliferate and synthesize excess
levels of extracellular matrix (ECM) (16,17).
Therefore, experimental analysis of HSC activation is an important
aspect of research into the pathogenesis hepatic fibrosis. Our
group previously reported that combination therapy with taurine,
epigallocatechin gallate (EGCG) and genistein exhibits a protective
effect against alcohol-induced liver fibrosis and also reduces cell
proliferation and the expression of fibrogenic factors in the rat
HSC-T6 cell line (15,18).
To determine the mechanism by which combination
treatment with taurine, EGCG and genistein protects against hepatic
fibrosis in the present study, an isobaric tag for relative and
absolute quantification (iTRAQ) was used to analyze the proteome of
HSC-T6 cells following treatment with combination therapy. The use
of cutting-edge iTRAQ-based proteomic profiling with
two-dimensional liquid chromatography coupled with tandem mass
spectrometry allows for multidimensional protein identification
with high sensitivity (19–21). This approach aids the analysis of the
entire proteome of a given biological sample in a single experiment
(22,23). This advanced proteomic approach was
used in the present study to improve mechanistic understanding of
hepatic fibrogenesis.
Materials and methods
Pharmacological agents and
reagents
Genistein, taurine, 3-[(3-cholamidopropyl)
dimethylammonio]-1-propanesulfonate (CHAPS), Tris, SDS,
DL-dithiothreitol (DTT), trichloroacetic acid (TCA), Dulbecco's
modified Eagle's medium (DMEM), sodium dihydrogen phosphate
(NaH2PO4), potassium chloride (KCl), acetone
and acetonitrile (ACN) were all purchased from Sigma-Aldrich; Merck
KGaA (Darmstadt, Germany). EGCG was acquired from Sichuan Yuga Tea
Development Co., Ltd. (Sichuan, China). Reagents for iTRAQ were
obtained from AB Sciex Pte., Ltd. (Concord, ON, Canada). Phosphate
buffered saline (PBS; pH 7.4), ampholine and apotinin were
purchased from Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
Furthermore, radioimmunoprecipitation assay lysis buffer was
purchased from Beyotime Institute of Biotechnology (Shanghai,
China) and Trypsin Gold was acquired from Promega Corporation
(Madison, WI, USA).
Cell lines
The HSC-T6 cell line, an immortalized rat HSC line
exhibiting a stable phenotype and biochemical characteristics of
liver fibrosis, was purchased from the Xiangya School of Medicine,
Central South University (Hunan, China). HSC-T6 cells were cultured
in DMEM supplemented with heat-inactivated 10% fetal bovine serum
(both Gibco; Thermo Fisher Scientific, Inc.) at 37°C in a
humidified atmosphere containing 5% CO2. Cells were
passaged via trypsinization every 3 days.
Protein extraction and
preparation
HSC-T6 cells were seeded into 50-ml culture flasks
at a density of 5×104 cells/ml. Following 24 h culture,
cells were treated with or without combined treatment with 0.03
mg/ml taurine, 0.035 mg/ml EGCG and 0.007 mg/ml genistein for 24 h,
as previously reported (15).
Subsequently cells were trypsinized, and centrifuged at 12,000 × g
and 4°C for 20 min to obtain cell pellets. Pellets were washed
three times with ice-cold PBS, resuspended in 1 ml lysis buffer
(containing 9 mol/l urea, 4% CHAPS, 40 mmol/l Tris, 1% DTT, 0.8%
ampholine and 0.002% apotinin), placed on ice and vortexed every 5
min for a total of 20 min. Cell supernatants were collected
following centrifugation at 12,000 × g for 20 min at 4°C.
Supernatants were then precipitated with pre-chilled acetone
(containing 10% v/v TCA) for 2 h and centrifuged at 12,000 × g for
15 min at 4°C. The resulting pellets were washed with pre-chilled
acetone, incubated at 4°C for 2 h and centrifuged again at 12,000 g
for 15 min at 4°C. Subsequently, cells were washed with pre-chilled
acetone a further three times. The final pellets were lyophilized
and protein concentration was determined using bovine serum albumin
(Wuhan Boster Biological Technology, Ltd., Wuhan, China) as the
standard (24).
Protein digestion and peptide iTRAQ
labeling
A total of 100 µg processed protein was reduced,
blocked with cysteine (25) and
digested with Trypsin Gold at a ratio of protein to trypsin of 20:1
at 37°C for 12 h. Following trypsin digestion, peptides were dried
using vacuum centrifugation (250 × g, −20°C, 1 h). Subsequently,
peptides were reconstituted in 0.5 mol/l tetraethylammonium bromide
and processed using 4-plex iTRAQ (both AB Sciex Pte., Ltd.)
following the manufacturer's protocol. One unit of iTRAQ reagent
was thawed and reconstituted in 150 µl 100% isopropanol. Peptides
were individually labeled with iTRAQ tags as follows: Control
group, 114; and the combination treatment group, 115. Following
incubation for 2 h at room temperature with the iTRAQ reagent,
labeled samples were mixed equally prior to further analysis
(25).
Strong cation exchange (SCX)
chromatography
SCX chromatography was performed using a LC-20AB
high performance liquid chromatography pump system (Shimadzu
Corporation, Kyoto, Japan). iTRAQ-labeled peptide mixtures were
reconstituted with 4 ml buffer A (25 mmol/l
NaH2PO4 in 25% ACN, pH 2.7) and loaded onto a
Ultremex SCX column (4.6×250 mm, 5 µm; Phenomenex, Torrance, CA,
USA). Peptides were eluted with a gradient of buffer A for 10 min,
5–35% buffer B (25 mmol/l NaH2PO4, 1 mol/l
KCl in 25% ACN, pH 2.7) for 11 min and 35–80% buffer B for 1 min at
a flow rate of 1 ml/min. The system was maintained in 80% buffer B
for 3 min prior to equilibrating with buffer A for 10 min before
the next injection. Elution was measured at 214 nm and fractions
were collected every min. Eluted peptides were pooled into 20
fractions, desalted with a Strata X C18 column (Phenomenex) and
vacuum-dried.
Liquid chromatography-electrospray
ionization-tandem mass spectrometry (LC-ESI-MS/MS) analysis
Each fraction was resuspended in buffer A, which
consisted of 2% ACN, 0.1% formic acid (FA), and centrifuged at
20,000 rpm for 10 min. In each fraction, the final concentration of
peptide was ~0.25 µg/µl. Using an auto sampler, 9 µl supernatant
was loaded onto a Symmetry C18 column (180 µm ×20 mm, 5 µm).
LC-ESI-MS/MS was performed using a nanoACQuity UPLC system (Waters
Corporation, Milford, MA, USA). The sample was eluted with buffer A
at 2 µl/min for 15 min. Peptides were eluted onto a BEH130 C18
column (100 µm ×10 mm, 1.7 µm; Waters Corporation) for online
trapping, desalting and analytical separation. Samples were loaded
at a flow rate of 300 nl/min with 5% buffer B (98% ACN and 0.1% FA)
for 1 min, eluted with a 40-min gradient from 5–35% buffer B,
followed by a 5-min linear gradient to 80% buffer B and maintenance
with 80% buffer B for 5 min. Initial chromatographic conditions
were restored following 2 min.
A TripleTOF 5600 system fitted with a Nanospray III
source (both AB Sciex Pte., Ltd.) and a pulled quartz tip as the
emitter (New Objective, Inc., Woburn, MA, USA) was used for data
acquisition. Data were acquired using an ion spray voltage of 2.5
kV, curtain gas of 30 PSI, nebulizer gas of 5 PSI and an interface
heater (temperature, 150°C). Survey scans were acquired in 250 msec
and the top-30 product ion scans were collected if a threshold of
120 counts per sec was exceeded and a +2 to +5 charge state was
exhibited. Four time bins were summed for each scan at a pulse
frequency value of 11 kHz by monitoring the 40 GHz multichannel TDC
detector with 4-anode channel detection. A sweeping collision
energy setting of 35±5 eV was applied to all precursor ions for
collision-induced dissociation. Dynamic exclusion was set for the ½
peak width (~18 sec) and the precursor was refreshed off of the
exclusion list. All iTRAQ experiments were performed in
triplicate.
Western blot analysis
Changes in the expression of hexokinase-2 (HK2) and
lysosome-associated membrane glycoprotein 1 (LAMP 1) were
determined by western blot analysis following iTRAQ analysis.
Proteins were extracted in the iTRAQ experiment and quantified with
a bicinchonic acid protein assay kit (Wuhan Boster Biological
Technology, Ltd.). Subsequently, 50 µg total protein were separated
in each lane via a 10% SDS-PAGE gels (Wuhan Boster Biological
Technology, Ltd.) and transferred to polyvinylidene fluoride
membranes (EMD Millipore, Billerica, MA, USA). Membranes were
blocked for 1 h at room temperature with 5% non-fat dried milk in
Tris-buffered saline with Tween 20 (TBST). Subsequently, proteins
were washed and incubated with anti-HK2 (cat. no. ab227198),
anti-LAMP 1 (cat. no. ab62562; both 1:1,000; Abcam, Cambridge, MA,
USA) and anti-GAPDH (cat. no. A00227-1; 1:500; Wuhan Boster
Biological Technology, Ltd.) antibodies at room temperature for 1
h. Membranes were washed three times with TBST and incubated with
secondary antibodies (IRDye® 680LT Goat anti-Rabbit IgG;
cat. no. 926-68021; 1:10,000; LI-COR Biosciences; Lincoln, NE, USA)
for 1 h at room temperature. Blots were washed three times with
TBST and detected using an LI-COR Odyssey® Infrared
Imaging system (Odyssey System, Version 2.0.25, LI-COR
Biosciences). GAPDH was used as the loading control and total
protein content in each lane was quantified using densitometry.
Experiments were performed in triplicate.
Data analysis and bioinformatics
Data analyses were performed using Protein-Pilot
software 4.0 (AB Sciex Pte., Ltd.) and the search parameter for
cysteine was determined to be the carbamidomethylation of cysteine.
Resulting MS/MS spectra were searched against the International
Protein Index (IPI) rat sequence databases (version 3.87; 39,925
sequences; http://www.ebi.ac.uk/IPI/IPIhelp.html) using MASCOT
software (version 2.3.02; Matrix Science, London, UK). For protein
identification and quantification, a peptide mass tolerance of 8.6
ppm was used for intact peptide masses and 0.05 Da for fragmented
ions. One missed cleavage was accepted in the trypsin digests,
carbamidomethylation of cysteine was considered to be a fixed
modification and the conversion of N-terminal glutamine to
pyro-glutamic acid and methionine oxidation were considered
variable modifications. All identified peptides had an ion score
above the Mascot peptide identity threshold and a protein was
considered identified if at least one such unique peptide match was
apparent for the protein. For protein-abundance ratios measured
using iTRAQ, fold-changes >1.3 or <0.7 were set as the
threshold and P<0.05 was considered to indicate a statistically
significant difference. The accession numbers of proteins that were
significantly differentially expressed were converted to a gene
list using the Database for Annotation Visualization and Integrated
Discovery (DAVID) functional annotation tool (http://david.abcc.ncifcrf.gov/summary.jsp) and
biological processes, cell components and molecular functions were
analyzed using the Gene Ontology terms (26). The protein interaction network mode
was created using the Search Tool for the Retrieval of Interacting
Genes (STRING) database (http://string-db.org/), which quantitatively
integrated interaction data for a large number of organisms and
transferred information between these organisms where
applicable.
Statistical analysis
To determine the proteins that were differentially
expressed, another two separate experiments were performed as
aforementioned. Statistical analysis was performed using SPSS 16.0
for Windows (SPSS, Inc., Chicago, IL, USA). Data were presented as
the mean ± standard deviation from three independent experiments.
Quantitative variables were analyzed using Student's t-test.
Spearman's rank was used to determine whether there was a
correlation between parameters. P<0.05 was determined to
indicate a statistical significance.
Results
To investigate the response in the HSC-T6 proteome
to combination treatment, quantitative proteomic analysis was
performed and differentially expressed proteins were identified. A
total of 713 distinct proteins were identified and quantified using
the MASCOT search algorithm against the IPI rat protein database.
According to the set change ratio (fold-change, >1.3 or <0.7;
P<0.05), 166 proteins were identified as significantly
differentially expressed proteins in all three experiments (data
not shown).
Functional classifications of
proteins
Of the 166 differentially expressed proteins, the
expression of 76 were increased and 90 were decreased following
combination treatment. Details of the proteins associated with cell
signaling pathways associated with fibrosis are listed in Table I. Peptide Mass (https://web.expasy.org/peptide_mass/)
and Prot Param (https://web.expasy.org/protparam/) tools in ExPASy
were used to predict peptides produced by trypsin digestion of heme
oxygenase 1 and analyze physical and chemical properties, including
peptides molecular weight, isoelectric point, amino acid, atom,
molar absorptivity, half-life, instability coefficient, aliphatic
amino acid coefficient and average hydrophilicity coefficient.
Peptides with good solubility and stability, and satisfying mass
spectrometry detectable mass-to-charge ratio were selected, and
analyzed for peptide homology using online software BlastP
(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome)
in NBCI database. Skyline (version 4.1; MacCoss Lab Software,
Washington, USA) was used to predict specific peptide scanning mass
spectrometry fragment information. Therefore, a representative mass
spectrum and quantitative information of the heme oxygenase 1
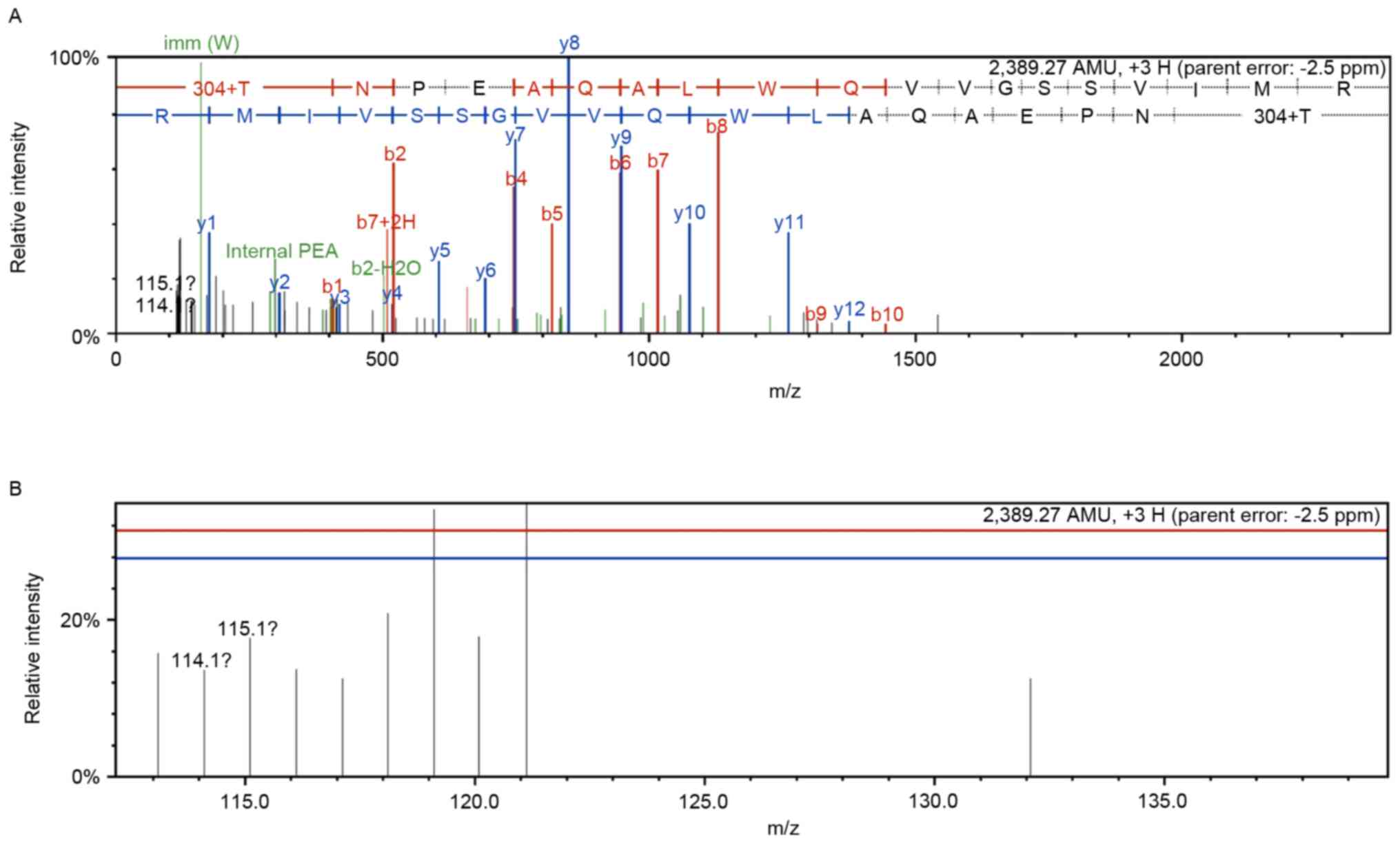
peptide (sequence: PSLFPAASGAFSSFR) is indicated in Fig. 1. First order mass and tandem mass
spectra of heme oxygenase 1 were presented in Fig. 1 (the control group, n=114 and the
combination treatment group, n=115).
 | Table I.Identification of differentially
expressed proteins for pathway and protein-protein interaction. |
Table I.
Identification of differentially
expressed proteins for pathway and protein-protein interaction.
| N | Accession no. | Gene symbol | Protein name | % Cov | Unique peptide | Ratio |
|---|
| 1 | IPI00215208 | Rpl8 | LOC100360117 60S
ribosomal protein L8 | 30.4 | 5 | 0.683±0.026 |
| 2 | IPI00230917 | Rpl18 | 60S ribosomal
protein L18 | 19.7 | 3 | 0.623±0.027 |
| 3 | IPI00203523 | Rpl23a | 60S ribosomal
protein L23a | 25.0 | 4 | 0.654±0.042 |
| 4 | IPI00231202 | Rps8 | 40S ribosomal
protein S8 | 45.7 | 8 | 0.697±0.073 |
| 5 | IPI00324983 | Rps17 | LOC100365810 40S
ribosomal protein S17 | 49.6 | 6 | 0.566±0.021 |
| 6 | IPI00358600 | RGD1559972 | Ribosomal protein
L27a-like | 17.6 | 2 | 0.541±0.051 |
| 7 | IPI00369491 | RGD1564744 | Ribosomal protein
P1-like | 29.2 | 2 | 0.510±0.060 |
| 8 | IPI00765221 | LOC683961 | Ribosomal protein
S13-like | 29.0 | 5 | 1.315±0.054 |
| 9 | IPI00206336 | Lamp1 | Lysosome-associated
membrane glycoprotein 1 | 5.4 | 2 | 1.585±0.013 |
| 10 | IPI00212731 | Ctsd | Cathepsin D | 28.0 | 8 | 1.412±0.010 |
| 11 | IPI00195160 | Psap | Prosaposin | 13.9 | 7 | 1.632±0.032 |
| 12 | IPI00230870 | Clta | Isoform Non-brain
of Clathrin light chain A | 13.3 | 2 | 0.687±0.016 |
| 13 | IPI00215580 | Atp6v0c | V-type proton
ATPase 16 kDa proteolipid subunit | 11.6 | 1 | 0.615±0.061 |
| 14 | IPI00201057 | Hk2 | Hexokinase-2 | 31.6 | 20 | 0.432±0.030 |
| 15 | IPI00951991 | Aldoa | 45 kDa protein | 58.6 | 13 | 1.313±0.012 |
| 16 | IPI00203690 | Aldh9a1 |
4-trimethylaminobutyraldehyde
dehydrogenase | 17.3 | 7 | 1.313±0.053 |
| 17 | IPI00364311 | Gpi | Glucose-6-phosphate
isomerase | 25.6 | 12 | 1.336±0.024 |
| 18 | IPI00231631 | Eno3 | Beta-enolase | 21.4 | 2 | 1.370±0.038 |
| 19 | IPI00209980 | Pmpcb |
Mitochondrial-processing peptidase subunit
beta | 12.1 | 4 | 0.567±0.057 |
| 20 | IPI00213245 | Stat1 | Signal transducer
and activator of transcription 1 isoform alpha | 15.9 | 10 | 0.603±0.098 |
| 21 | IPI00205805 | Timp1 | Metalloproteinase
inhibitor 1 | 40.1 | 6 | 0.529±0.030 |
| 22 | IPI00200610 | Gfm1 | Elongation factor
G, mitochondrial | 18.2 | 10 | 0.699±0.058 |
| 23 | IPI00192246 | Cox5a | Cytochrome c
oxidase subunit 5A, mitochondrial | 28.1 | 3 | 0.602±0.020 |
| 24 | IPI00210945 | Tpm1 | Tropomyosin alpha-1
chain isoform c | 31.3 | 1 | 0.425±0.014 |
| 25 | IPI00187731 | Tpm2 | Isoform 2 of
Tropomyosin beta chain | 31.3 | 3 | 0.499±0.026 |
| 26 | IPI00203832 | Plrg1 | Pleiotropic
regulator 1 | 4.7 | 2 | 0.679±0.034 |
| 27 | IPI00763263 | Thoc4 | Uncharacterized
protein | 23.4 | 4 | 0.574±0.059 |
| 28 | IPI00194222 | Cox4i1 | Cytochrome c
oxidase subunit 4 isoform 1, mitochondrial | 26.6 | 4 | 0.624±0.066 |
| 29 | IPI00200920 | Khsrp | Far upstream
element-binding protein 2 | 30.1 | 14 | 0.482±0.015 |
| 30 | IPI00464532 | Pycr2 |
Pyrroline-5-carboxylate reductase 2 | 25.3 | 3 | 1.483±0.023 |
| 31 | IPI00372370 | P4ha2 | Uncharacterized
protein | 23.7 | 9 | 1.317±0.034 |
| 32 | IPI00365868 | Acyp1 |
Acylphosphatase-1 | 47.5 | 4 | 1.416±0.094 |
| 33 | IPI00480766 | Acat3 | Acetyl-CoA
acetyltransferase, cytosolic | 22.2 | 4 | 1.595±0.028 |
| 34 | IPI00195860 | Cox7a2 | Cox7a2 Cytochrome c
oxidase subunit 7A2, mitochondrial | 12.0 | 1 | 3.396±0.034 |
| 35 | IPI00210920 | Got2 | Aspartate
aminotransferase, mitochondrial | 34.7 | 14 | 1.391±0.086 |
| 36 | IPI00214536 | Nop58 | Nucleolar protein
58 | 31.1 | 11 | 1.322±0.005 |
| 37 | IPI00197164 | Icam1 | Intercellular
adhesion molecule 1 | 10.8 | 5 | 1.419±0.010 |
| 38 | IPI00371124 | Srsf9 |
Serine/arginine-rich splicing factor
9 | 21.7 | 4 | 1.455±0.064 |
| 39 | IPI00411215 | Phf5a | PHD finger-like
domain-containing protein 5A | 18.2 | 2 | 1.379±0.021 |
| 40 | IPI00192936 | Magoh | Protein mago nashi
homolog | 51.4 | 7 | 1.369±0.039 |
| 41 | IPI00357978 | Srsf6 | Splicing factor,
arginine/serine-rich 6 | 11.8 | 3 | 1.354±0.029 |
| 42 | IPI00372819 | Snrpc | U1 small nuclear
ribonucleoprotein C | 18.9 | 2 | 1.741±0.036 |
| 43 | IPI00188158 | Hmgcs1 |
Hydroxymethylglutaryl-CoA synthase,
cytoplasmic | 17.1 | 8 | 1.733±0.025 |
| 44 | IPI00204738 | Aacs | Acetoacetyl-CoA
synthetase | 4.5 | 3 | 1.312±0.021 |
| 45 | IPI00766047 | LOC679203 | TH1-like isoform
4 | 7.3 | 4 | 1.756±0.057 |
The 166 differentially expressed proteins were
analyzed using the DAVID functional annotation tool to determine
their cellular components, molecular functions and participation in
biological processes (data not shown). The top three biological
processes included cellular processes (11.86%), single-organism
processes (10.09%) and metabolic processes (9.85%). The top three
molecular function categories included binding (49.82%), catalytic
activity (24.21%) and structural molecule activity (6.67%). The top
three associated cellular components included the cell (37.60%),
organelle (27.15%) and membrane (11.55%).
Protein-protein interaction
analysis
In order to characterize the protein-protein
interactions of the identified differentially expressed proteins,
the STRING database was assessed to establish a protein-protein
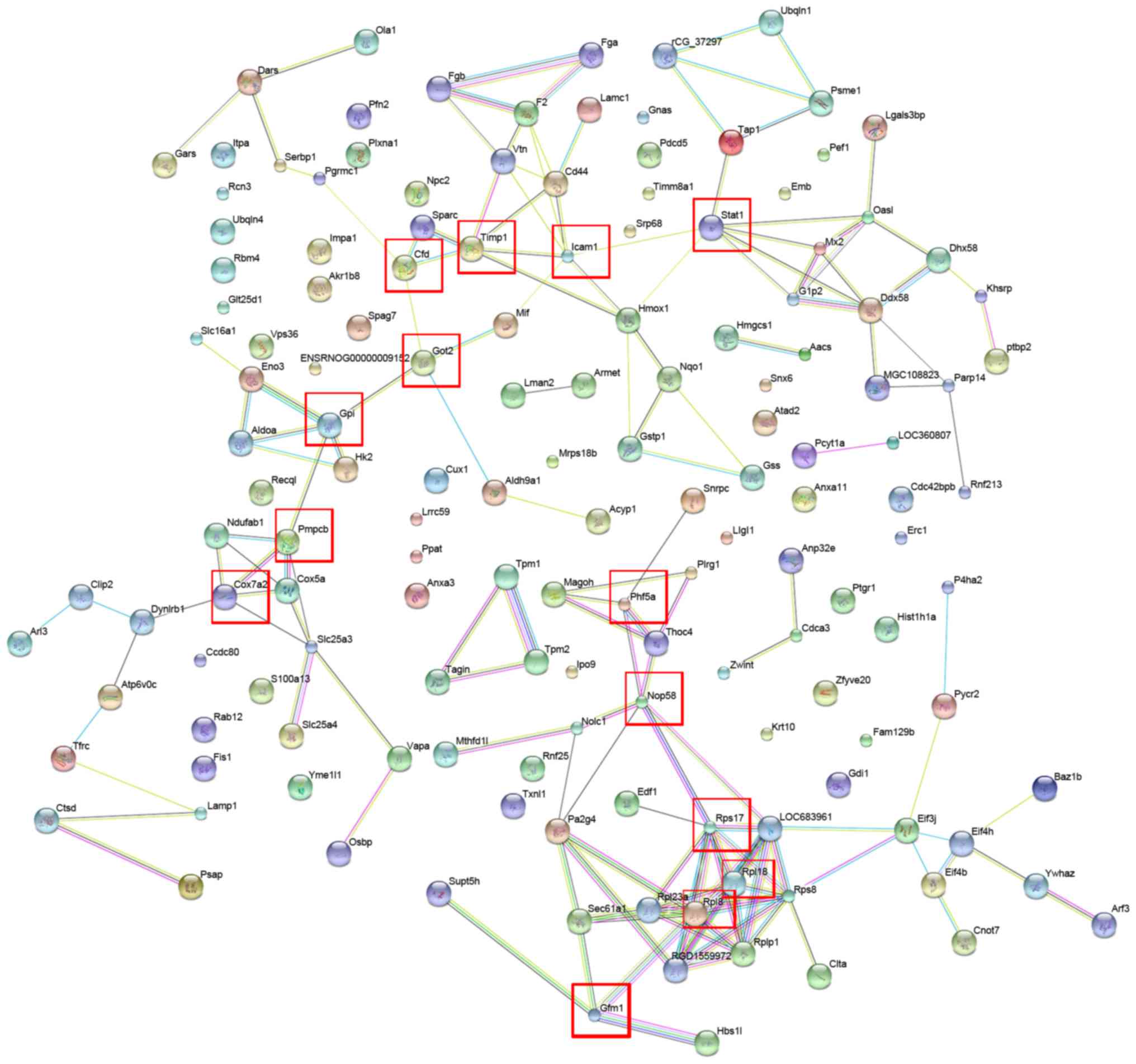
interaction network (Fig. 2).
Proteins that responded significantly to combination treatment and
interacted with each other included Stat1, Icaml, Timp1, Cfd, Got2,
Gpi, Pmpcb, Cox7a2, Phf5a, Nop58, Rps17, Rpl18, Rp18 and Gfm1
(Fig. 2). These seed proteins serve
important roles in catalytic activity, biological regulation and
coagulation systems (27–33). Furthermore, pathway annotation
indicated that certain differentially expressed proteins, including
glucose-6-phosphate isomerase (GPI) and ribosomal protein L8
(RPL8), were involved in eight different Kyoto Encyclopedia of
Genes and Genomes (KEGG) pathways (glycolysis and gluconeogenesis;
lysosome metabolism; ribosome metabolism; spliceosome metabolism;
butanoate metabolism; arginine and proline metabolism; cardiac
muscle contraction; and pyruvate metabolism signaling pathways;
Table II). Notably, GPI is a
crucial enzyme that catalyzes the interconversion of
glucose-6-phosphate and fructose-6-phosphate in glycolysis and
gluconeogenesis (34). Rp18 is a
component of the 60S subunit of the ribosome that is involved in
the protein synthesis pathway. Depletion of Rp18 impairs drosophila
development and is associated with apoptosis (35). The results suggest that the
interactions and signaling pathways associated with the
differentially expressed proteins may serve important roles in the
pathogenesis of liver fibrosis.
| Table II.Pathway annotation of the
differentially expressed proteins involved in eight different Kyoto
Encyclopedia of Genes and Genomes pathways. |
Table II.
Pathway annotation of the
differentially expressed proteins involved in eight different Kyoto
Encyclopedia of Genes and Genomes pathways.
| Pathway | Differentially
expressed proteins with pathway annotation |
|---|
| Glycolysis and
gluconeogenesis | Hk2, Aldoa,
Aldh9a1, Gpi, Eno3 |
| Lysosome | Atp6v0c, Clta,
Psap, Ctsd, Lamp1 |
| Ribosome | Rpl8, Rpl18, Rps8,
Rps17, Rpl23a, LOC683961 RGD1559972, RGD1564744 |
| Spliceosome | Magoh, Thoc4,
Snrpc, Srsf9, Phf5a, Srsf6, Plrg1 |
| Butanoate
metabolism | Hmgcs1, Aldh9a1,
Aacs, LOC679203 |
| Arginine and
proline metabolism | Aldh9a1, Khsrp,
Pycr2, P4ha2 |
| Cardiac muscle
contraction | RGD1559972, Cox5a,
Tpm1, Tpm2 |
| Pyruvate
metabolism | Aldh9a1, Acyp1,
Acat3 |
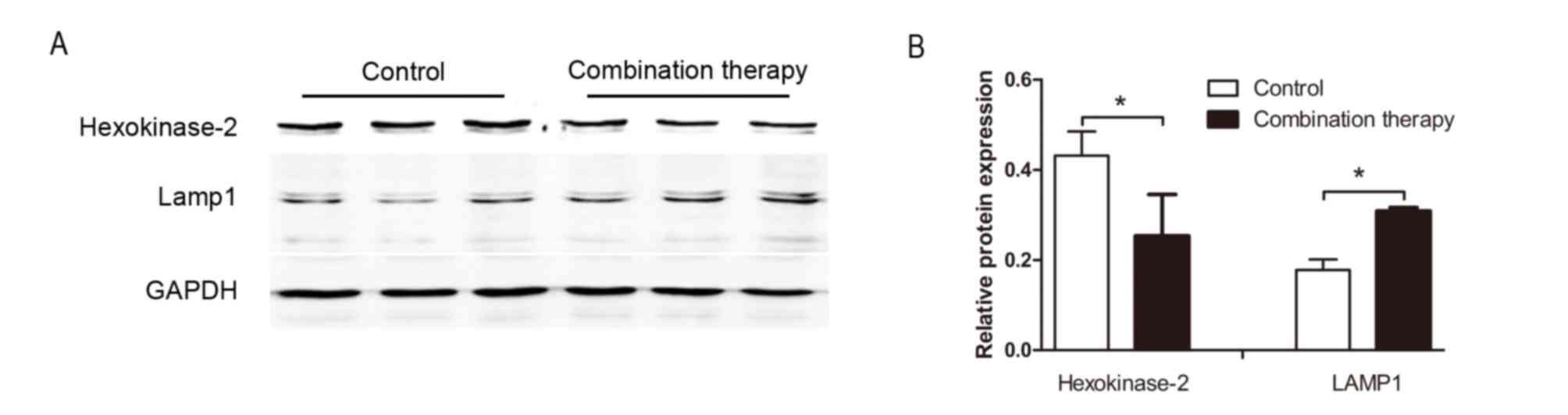
Confirmation of differentially
expressed proteins using western blot analysis
To further investigate the expression of the
differentially expressed proteins obtained from the iTRAQ-based
quantitative proteomics study, western blot analysis was performed
to assess the expression of HK2 and LAMP 1. Levels of HK2 and LAMP
1 were significantly downregulated and upregulated in HSC-T6 cells
following combination therapy, respectively (each, P<0.05;
Fig. 3). This pattern was similar to
the changes in the expression of these proteins obtained following
iTRAQ (Table I), supporting the
validity of the proteomic analysis performed in the present
study.
Discussion
A variety of biological factors are involved in the
pathogenesis of liver fibrosis. As an approach to investigate the
multiple signaling pathways that may contribute to liver fibrosis,
our group previously reported that combination therapy with
taurine, EGCG and genistein exhibits a protective effect against
liver fibrosis in vivo and in vitro (15,18). In
the present study, the mechanistic underpinnings of this effect in
HSC-T6 cells were evaluated using a proteomic approach. Proteomic
analysis is a powerful tool for revealing the molecular mechanisms
of disease development, and is useful for the identification of
disease-specific biomarkers and evaluating biological networks for
drug therapy (36,37). Previous studies have also revealed
that the optimal approach for developing novel therapies is to
characterize key regulatory pathways and networks (38,39).
One of the primary objectives of treatments for
liver fibrosis is the inhibition of proliferation or the induction
of apoptosis of activated HSC cells (40). Therefore, in the present study,
activated HSC-T6 cells were used to study the anti-fibrotic effects
of combination treatment with taurine, EGCG and genistein.
iTRAQ-based proteomics was used to analyze the molecular targets of
combination treatment with taurine, EGCG and genistein in HSC-T6
cells. A total of 166 differentially expressed proteins were
identified, which represent a diverse array of molecular weights,
isoelectric point and protein functions. In a previous study by our
group, it was demonstrated that changes in the expression of
transforming growth factor (TGF)-β1, tissue inhibitor of
metalloproteinases (TIMP)-1 and −2, matrix
metalloproteinase (MMP)-2 and collagen type (Col)-I
mRNA were associated with hepatic fibrogenesis in HSC-T6 cells
(10). The present study also
revealed similar results regarding the expression of TGF-β1,
TIMP-1, MMP-2, TIMP-2 and Col-1 proteins. In the present study, due
to the set change ratio (fold-change >1.3 or <0.7), the
proteins TIMP-2 and Col-1 were not regarded as significantly
differentially expressed. Furthermore, bioinformatics analysis
revealed that all 166 differentially expressed proteins are
involved in various biological processes, including metabolic
processes, single-organism processes, cellular processes, response
to stimuli, cell necrosis, reproduction and cell apoptosis (data
not shown). Additionally, several identified significantly
differentially expressed proteins, including Hk2, Aldoa, Aldh9a1,
Gpi, Eno3, Atp6v0c, Clta, Psap, Ctsd and Lamp1, were associated
with eight different KEGG pathways that connected with each other
to form a network (Table II).
Based on the present data, it was speculated that
the antifibrotic activity associated with combination treatment
with taurine, EGCG and genistein in HSC-T6 cells was associated
with multiple biological processes and signaling pathways. From
these indicated associations, the cellular processes of glycolysis
and gluconeogenesis, and the signaling pathways of lysosome and
ribosome synthesis were suggested to be of particular importance.
Glycolysis and gluconeogenesis serve important roles in supplying
ATP for hypoxic metabolism, cellular proliferation and apoptosis
(41). Several enzymes and proteins,
including hexokinase-2, aldolase A and glucose-6-phosphate
isomerase, associated with glycolysis and gluconeogenesis may be
potential molecular targets in pancreatic, breast and
gastrointestinal tumors (42,43). The
ribosome signaling pathway has been implicated in a wide variety of
biological functions, including cell cycle progression, apoptosis
and DNA damage responses (44–47).
Furthermore, the lysosomal signaling pathway may be involved in a
series of pathological processes, including cell death, necrosis,
apoptosis and autophagy (48).
In the early phase of liver damage or inflammation,
the hepatocyte microenvironment becomes hypoxic, leading to a
failure in oxidative energy generation and a switch to the
glycolysis and gluconeogenesis pathway to produce ATP (3). HK2 is the rate-limiting enzyme in the
glycolysis and gluconeogenesis pathway and its expression is
significantly enhanced in different types of cancer, including
metastatic colorectal cancer, lung cancer and gastric cancer
(49–52) and rapidly proliferating cells,
including those in the developing embryo (53). Previous studies have reported that
HK2 binding in the mitochondrial membrane may promote tumor growth
by inhibiting apoptosis and promoting cell proliferation (54,55).
This suggests that HK2 may be a potential molecular target in the
treatment of various diseases (56,57). The
results of the present study indicate that combination treatment
with taurine, EGCG and genistein significantly inhibits the
expression of HK2. This may potentially lead to improved
oxygenation in the cell microenvironment and induce the apoptosis
of HSC-T6 cells. Thus, downregulation of HK2 may contribute to the
antifibrotic effects associated with combination therapy.
GPI is a crucial enzyme that catalyzes the
interconversion of glucose-6-phosphate and fructose-6-phosphate in
glycolysis and gluconeogenesis (34). Furthermore, GPI is secreted by tumor
cells and stimulates cell migration (58). In hepatocellular carcinoma cells, GPI
may promote cell movement and activate the synthesis of MMP-3
(59). Upregulated GPI expression in
response to combination treatment with taurine, EGCG and genistein
in HSC-T6 cells, as identified in the current study, may induce
MMP-3 synthesis. MMP-3 is associated with the degradation of the
ECM (60); therefore, this
upregulation may promote ECM degradation and contribute to the
reversal of hepatic fibrosis.
Autophagy is a degradation and recirculation system
in eukaryotic cells that is required for the induction of apoptosis
(61,62). During autophagy, lysosomes serve an
important role in the degradation and recycling of material within
the terminal organelle (63). The
protein cathepsin D is a major incision enzyme in lysosomes that
maintains cell metabolism by degrading intracellular substances
(64). The overexpression of
cathepsin D may stimulate a series of pathological processes,
including cell necrosis, apoptosis and autophagy (48,65).
Another important lysosomal protein is LAMP 1, which may induce
autophagy and apoptosis (65). A
previous study indicated that LAMP 1 was upregulated and induced
apoptosis in human glioblastoma cells following treatment with
cisplatin (66). In the present
study, cathepsin D and LAMP 1 were upregulated in HSC-T6 cells
following combination treatment. In Table I, relative LAMP1 protein expression
was increased at a ratio of 1.585±0.013 after treatment and
cathepsin D was increased at a ratio of 1.412±0.010. Relative HK2
protein expression was decreased at a ratio of 0.432±0.030
following treatment.
These results suggest that the upregulation of
cathepsin D may lead to the interaction with LAMP 1 in the lysosome
pathway, initiating HSC autophagy and apoptosis, thus contributing
to the antifibrotic effect of the combination treatment. Ribosome
proteins are crucial components of the ribosomal subunits that
function in ribosome assembly and protein synthesis (67,68). The
abnormal expression of specific ribosome proteins may be
responsible for several human conditions, including
Diamond-Blackfan anemia (69),
Turner syndrome (70), hearing loss
(71) and cancer (72).
In conclusion, combination treatment with taurine,
EGCG and genistein induced antifibrotic effects in HSC-T6 cells in
at least three ways: i) Regulation of energy metabolism in HSCs via
glycolysis and gluconeogenesis; ii) protection of the liver via the
ribosome signaling pathway, with subsequent effects on numerous
biological functions, including cell cycle progression, protein
synthesis and DNA damage responses; and iii) regulation of
apoptosis and autophagy via the lysosomal signaling pathway.
Although the exact roles of these pathways remain to be elucidated,
the present study has enhanced understanding of the molecular
mechanism by which combination therapy alleviates liver fibrosis.
Furthermore, the present study also demonstrated that protein
profiling using iTRAQ-based proteomics is a powerful method of
performing quantitative proteome analysis in cell models in
response to therapy. Furthermore, the results suggest that
iTRAQ-based proteomics may be used to identify underlying
mechanisms of action and potential targets for novel therapies.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81160063,
81360078 and 81460128); the Guangxi Natural Science Foundation
(grant no. 2013GXNSFBA019167) and the Foundation of
High-Incidence-Tumor Prevention & Treatment, Guangxi Medical
University, Ministry of Education (grant no.
GK2013-13-A-01-03).
Availability of data and materials
All data generated and/or analyzed during the
present study are included in this published article.
Authors' contributions
YL analyzed data and wrote the manuscript. MZ and YH
performed the experiments. ML and XZ designed the study. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The Animal Experimentation Ethics Committee of
Guangxi Medical University (Nanning, China) approved the
experimental animal protocol of the present study.
Patient consent for publication
Not applicable.
Competing interest
The authors declare that they have no competing
interests.
References
|
1
|
Friedman SL: Liver fibrosis-from bench to
bedside. J Hepatol. 38 Suppl 1:S38–S53. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ahmad A and Ahmad R: Understanding the
mechanism of hepatic fibrosis and potential therapeutic approaches.
Saudi J Gastroenterol. 18:155–167. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bonis PAL, Friedman SL and Kaplan MM: Is
liver fibrosis reversible? N Engl J Med. 344:452–454. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Berenguer M: Hepatitis C virus and liver
transplantation. Springer; 2014, View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hammel P, Couvelard A, O'Toole D, Ratouis
A, Sauvanet A, Fléjou JF, Degott C, Belghiti J, Bernades P, Valla
D, et al: Regression of liver fibrosis after biliary drainage in
patients with chronic pancreatitis and stenosis of the common bile
duct. N Engl J Med. 344:4182001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sobhy MMK, Mahmoud SS, El-Sayed SH, Rizk
EMA, Raafat A and Negm MSI: Impact of treatment with a protein
tyrosine kinase inhibitor (Genistein) on acute and chronic
experimental Schistosoma mansoni infection. Exp Parasitol.
185:115–123. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kong LJ, Li H, Du YJ, Pei FH, Hu Y, Zhao
LL and Chen J: Vatalanib, a tyrosine kinase inhibitor, decreases
hepatic fibrosis and sinusoidal capillarization in CCl4-induced
fibrotic mice. Mol Med Rep. 15:2604–2610. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Czaja AJ and Carpenter HA: Progressive
fibrosis during corticosteroid therapy of autoimmune hepatitis. J
Hepatol. 39:1631–1638. 2004. View Article : Google Scholar
|
|
9
|
Lieber CS: Role of oxidative stress and
antioxidant therapy in alcoholic and nonalcoholic liver diseases.
Adv Pharmacol. 38:601–628. 1996. View Article : Google Scholar
|
|
10
|
Sánchez-Valle V, Chávez-Tapia NC, Uribe M
and Méndez-Sánchez N: Role of oxidative stress and molecular
changes in liver fibrosis: A review. Curr Med Chem. 19:4850–4860.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Distler JH and Distler O: Tyrosine kinase
inhibitors for the treatment of fibrotic diseases such as systemic
sclerosis: Towards molecular targeted therapies. Ann Rheum Dis. 69
Suppl 1:i48–i51. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gebhardt R: Oxidative stress,
plant-derived antioxidants and liver fibrosis. Planta Med.
68:289–296. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Carragher NO, Unciti-Broceta A and Cameron
DA: Advancing cancer drug discovery towards more agile development
of targeted combination therapies. Future Med Chem. 4:87–105. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kumar M and Sarin SK: Systematic review:
Combination therapies for treatment-naive chronic hepatitis B.
Aliment Pharmacol Ther. 27:11872008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhuo L, Liao M, Zheng L, He M, Huang Q,
Wei L, Huang R, Zhang S and Lin X: Combination therapy with
taurine, epigallocatechin gallate and genistein for protection
against hepatic fibrosis induced by alcohol in rats. Biol Pharm
Bull. 35:1802–1810. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Friedman SL: Mechanism of hepatic
fibrogenesis. Gastroenterology. 134:1655–1669. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bataller R and Brenner DA: Liver fibrosis.
J Clin Invest. 115:209–218. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li Y, Luo Y, Zhang X, Lin X, He M and Liao
M: Combined taurine, epigallocatechin gallate and genistein therapy
reduces HSC-T6 cell proliferation and modulates the expression of
fibrogenic factors. Int J Mol Sci. 14:20543–20554. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mallick P and Kuster B: Proteomics: A
pragmatic perspective. Nat Biotechnol. 28:695–709. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Song X, Bandow J, Sherman J, Baker JD,
Brown PW, McDowell MT and Molloy MP: iTRAQ experimental design for
plasma biomarker discovery. J Proteome Res. 7:2952–2958. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Glen A, Gan CS, Hamdy FC, Eaton CL, Cross
SS, Catto JW, Wright PC and Rehman I: iTRAQ-facilitated proteomic
analysis of human prostate cancer cells identifies proteins
associated with progression. J Proteome Res. 7:897–907. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hanash SM, Bobek MP, Rickman DS, Williams
T, Rouillard JM, Kuick R and Puravs E: Integrating cancer genomics
and proteomics in the post-genome era. Proteomics. 2:69–75. 2015.
View Article : Google Scholar
|
|
23
|
Srinivas PR, Kramer BS and Srivastava S:
Trends in biomarker research for cancer detection. Lancet Oncol.
2:698–704. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Peterson GL: A simplification of the
protein assay method of Lowry et al. which is more generally
applicable. Anal Biochem. 83:346–356. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cao W, Zhou Y, Li Y, Zhang X, He M, Zang
N, Zhou Y and Liao M: iTRAQ-based proteomic analysis of combination
therapy with taurine, epigallocatechin gallate, and genistein on
carbon tetrachloride-induced liver fibrosis in rats. Toxicol Lett.
232:233–245. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cao W, Li Y, Li M, Zhang X and Liao M:
Txn1, Ctsd and Cdk4 are key proteins of combination therapy with
taurine, epigallocatechin gallate and genistein against liver
fibrosis in rats. Biomed Pharmacother. 85:611–619. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang Z, Meng Q, Zhao Y, Han R, Huang S, Li
M, Wu X, Cai W and Wang H: Resveratrol promoted interferon-induced
growth inhibition and apoptosis of SMMC7721 cells by activating the
SIRT/STAT1. J Interferon Cytokine Res. 38:261–271. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zheng YZ, Xue MZ, Shen HJ, Li XG, Ma D,
Gong Y, Liu YR, Qiao F, Xie HY, Lian B, et al: PHF5A epigenetically
inhibits apoptosis to promote breast cancer progression. Cancer
Res. 78:3190–3206. 2018.PubMed/NCBI
|
|
29
|
Wingren AG, Parra E, Varga M, Kalland T,
Sjögren HO, Hedlund G and Dohlsten M: T cell activation pathways:
B7, LFA-3, and ICAM-1 shape unique T cell profiles. Crit Rev
Immunol. 15:235–253. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang K, Lin B, Brems JJ and Gamelli RL:
Hepatic apoptosis can modulate liver fibrosis through TIMP1
pathway. Apoptosis. 18:566–577. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sookoian S, Castaño GO, Scian R, Gianotti
Fernández T, Dopazo H, Rohr C, Gaj G, Martino San J, Sevic I,
Flichman D and Pirola CJ: Serum aminotransferases in nonalcoholic
fatty liver disease are a signature of liver metabolic
perturbations at the amino acid and Krebs cycle level. Am J Clin
Nutr. 103:422–434. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Swan EJ, Maxwell AP and Mcknight AJ:
Distinct methylation patters in genes that affect mitochondrial
function are associated with kidney disease in blood-derived DNA
from individuals with type 1 diabetes. Diabet Med. 32:1110–1115.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kenney SP and Meng XJ: Identification and
fine mapping of nuclear and nucleolar localization signals within
the human ribosomal protein S17. Plos One. 10:e01243962015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Watanabe H, Takehana K, Date M, Shinozaki
T and Raz A: Tumor cell autocrine motility factor is the
neuroleukin/phosphohexose isomerase polypeptide. Cancer Res.
56:2960–2963. 1996.PubMed/NCBI
|
|
35
|
Lindsley DL, Sandler L, Baker BS,
Carpenter AT, Denell RE, Hall JC, Jacobs PA, Miklos GL, Davis BK,
Gethmann RC, et al: Segmental aneuploidy and the genetic gross
structure of the Drosophila genome. Genetics. 71:157–184.
1972.PubMed/NCBI
|
|
36
|
Hogeweg P: The roots of bioinformatics in
theoretical biology. PLoS Comput Biol. 7:e10020212011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dix M and Cravatt B: Global mapping of the
topography and magnitude of proteolytic events in apoptosis. Cell.
134:679–691. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee MJ, Ye AS, Gardino AK, Heijink AM,
Sorger PK, MacBeath G and Yaffe MB: Sequential application of
anticancer drugs enhances cell death by rewiring apoptotic
signaling networks. Cell. 149:780–794. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fu LL, Zhou CC, Yao S, Yu JY, Liu B and
Bao JK: Plant lectins: Targeting programmed cell death pathways as
antitumor agents. Int J Biochem Cell B. 43:1442–1449. 2011.
View Article : Google Scholar
|
|
40
|
Pellicoro A, Ramachandran P and Iredale
JP: Reversibility of liver fibrosis. Fibrogenesis Tissue Repair. 5
Suppl 1:S262012.PubMed/NCBI
|
|
41
|
Rui L: Energy metabolism in the liver.
Compr Physiol. 4:177–197. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhou YY, Cheng CL, Baranenko D, Wang JP,
Li YZ and Lu WH: Effects of acanthopanax senticosus on brain injury
induced by simulated spatial radiation in mouse model based on
pharmacokinetics and comparative proteomics. Int J Mol Sci.
19:E1592018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wan XP, Xie P, Bu Z and Zou XT: Changes in
hepatic glucose and lipid metabolism-related parameters in domestic
pigeon (Columba livia) during incubation and chick rearing. J Anim
Physiol Anim Nutr (Berl). 102:e558–e568. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Volarević S and Thomas G: Role of S6
phosphorylation and S6 kinase in cell growth. Prog Nucleic Acid Res
Mol Biol. 65:101–127. 2000. View Article : Google Scholar
|
|
45
|
Volarevic S, Stewart MJ, Ledermann B,
Zilberman F, Terracciano L, Montini E, Grompe M, Kozma SC and
Thomas G: Proliferation, but not growth, blocked by conditional
deletion of 40S ribosomal protein S6. Science. 288:2045–2047. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lohrum MA, Ludwig RL, Kubbutat MH, Hanlon
M and Vousden KH: Regulation of HDM2 activity by the ribosomal
protein L11. Cancer Cell. 3:577–587. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen FW and Ioannou YA: Ribosomal proteins
in cell proliferation and apoptosis. Int Rev Immunol. 18:429–448.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Qin AP, Zhang HL and Qin ZH: Mechanisms of
lysosomal proteases participating in cerebral ischemia-induced
neuronal death. Neurosci Bull. 24:117–123. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang H, Peng R, Chen X, Jia R, Huang C,
Huang Y, Xia L and Guo G: Effect of HK2, PKM2 and LDHA on Cetuximab
efficacy in metastatic colorectal cancer. Oncol Lett. 15:5553–5560.
2018.PubMed/NCBI
|
|
50
|
Wolf A, Agnihotri S, Munoz D and Guha A:
Developmental profile and regulation of the glycolytic enzyme
hexokinase 2 in normal brain and glioblastoma multiforme. Neurobiol
Dis. 44:84–91. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ong LC, Jin Y, Song IC, Yu S, Zhang K and
Chow PK: 2-[18F]-2-deoxy-D-glucose (FDG) uptake in human tumor
cells is related to the expression of GLUT-1 and hexokinase II.
Acta Radiol. 49:1145–1153. 2009. View Article : Google Scholar
|
|
52
|
Goel A, Mathupala SP and Pedersen PL:
Glucose metabolism in cancer. Evidence that demethylation events
play a role in activating type II hexokinase gene expression. J
Biol Chem. 278:15333–15340. 2015. View Article : Google Scholar
|
|
53
|
Wolf A, Agnihotri S, Micallef J, Mukherjee
J, Sabha N, Cairns R, Hawkins C and Guha A: Hexokinase 2 is a key
mediator of aerobic glycolysis and promotes tumor growth in human
glioblastoma multiforme. J Exp Med. 208:313–326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Pedersen PL, Mathupala S, Rempel A,
Geschwind JF and Ko YH: Mitochondrial bound type II hexokinase: A
key player in the growth and survival of many cancers and an ideal
prospect for therapeutic intervention. Biochim Biophys Acta.
1555:14–20. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pastorino JG and Hoek JB: Hexokinase II:
The integration of energy metabolism and control of apoptosis. Curr
Med Chem. 10:1535–1551. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mathupala SP, Ko YH and Pedersen PL:
Hexokinase II: Cancer's double-edged sword acting as both
facilitator and gatekeeper of malignancy when bound to
mitochondria. Oncogene. 25:4777–4786. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Jae HJ, Jin WC, Park HS, Lee MJ, Lee KC,
Kim HC, Yoon JH, Chung H and Park JH: The antitumor effect and
hepatotoxicity of a hexokinase II inhibitor 3-bromopyruvate: In
vivo investigation of intraarterial administration in a rabbit VX2
hepatoma model. Korean J Radiol. 10:596–603. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Liotta LA, Mandler R, Murano G, Katz DA,
Gordon RK, Chiang PK and Schiffmann E: Tumor cell autocrine
motility factor. Proc Natl Acad Sci USA. 83:3302–3306. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ho JC, Cheung ST, Patil M, Chen X and Fan
ST: Increased expression of glycosyl-phosphatidylinositol anchor
attachment protein 1 (GPAA1) is associated with gene amplification
in hepatocellular carcinoma. Int J Cancer. 119:1330–1337. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yu FL, Liao MH, Lee JW and Shih WL:
Induction of hepatoma cells migration by phosphoglucose
isomerase/autocrine motility factor through the upregulation of
matrix metalloproteinase-3. Biochem Biophys Res Commun. 314:76–82.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Scott RC, Juhász G and Neufeld TP: Direct
induction of autophagy by Atg1 inhibits cell growth and induces
apoptotic cell death. Curr Biol. 17:1–11. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Crighton D, Wilkinson S, O'Prey J, Syed N,
Smith P, Harrison PR, Gasco M, Garrone O, Crook T and Ryan KM:
DRAM, a p53-induced modulator of autophagy, is critical for
apoptosis. Cell. 126:121–134. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Mizushima N, Levine B, Cuervo AM and
Klionsky DJ: Autophagy fights disease through cellular
self-digestion. Nature. 451:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Liang ZQ, Wang X, Li LY, Wang Y, Chen RW,
Chuang DM, Chase TN and Qin ZH: Nuclear factor-kappaB-dependent
cyclin D1 induction and DNA replication associated with
N-methyl-D-aspartate receptor-mediated apoptosis in rat striatum. J
Neurosci Res. 85:1295–1309. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Chen JW, Pan W, D'Souza MP and August JT:
Lysosome-associated membrane proteins: Characterization of LAMP-1
of macrophage P388 and mouse embryo 3T3 cultured cells. Arch
Biochem Biophys. 239:574–586. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Chen JW, Madamanchi N, Madamanchi NR,
Trier TT and Keherly MJ: Lamp-1 is upregulated in human
glioblastoma cell lines induced to undergo apoptosis. J Biomed Sci.
8:365–374. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Ray PS, Arif A and Fox PL: Macromolecular
complexes as depots for releasable regulatory proteins. Trends
Biochem Sci. 32:158–164. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Jewett MC, Fritz BR, Timmerman LE and
Church GM: In vitro integration of ribosomal RNA synthesis,
ribosome assembly, and translation. Mol Syst Biol. 9:6782013.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Draptchinskaia N, Gustavsson P, Andersson
B, Pettersson M, Willig TN, Dianzani I, Ball S, Tchernia G, Klar J,
Matsson H, et al: The gene encoding ribosomal protein S19 is
mutated in Diamond-Blackfan anaemia. Nat Genet. 21:169–175. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Fisher EMC, Beer-Romero P, Brown LG,
Ridley A, McNeil JA, Lawrence JB, Willard HF, Bieber FR and Page
DC: Homologous ribosomal protein genes on the human X and Y
chromosomes: Escape from X inactivation and possible implications
for turner syndrome. Cell. 63:1205–1218. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
O'Brien TW, O'Brien BJ and Norman RA:
Nuclear MRP genes and mitochondrial disease. Gene. 354:147–151.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Ruggero D and Pandolfi PP: Does the
ribosome translate cancer? Nat Rev Cancer. 3:179–192. 2003.
View Article : Google Scholar : PubMed/NCBI
|