Introduction
Recently, an increasing number of studies have
focused on stem cells for cell therapy (1–3). The
application of stem cell to the field of regenerative medicine
provides novel strategies to induce tissue repair (4,5). Studies
have documented the effect of mesenchymal stem or stromal cells
(MSCs) and endothelial progenitor cells (EPCs) on postnatal
vasculogenesis (6,7). MSCs act as precursors of mesenchymal
tissue cells. Friedenstein et al (8) first described MSCs as fibroblast
precursors from bone marrow (BM) and Caplan (9) first proposed the term ‘mesenchymal stem
cell’. MSCs exhibit a self-renewing capacity, ability to
differentiate into multiple lineages and immunomodulatory potential
(10). EPCs are precursors of
endothelial cells and can mature into cells that line the lumen of
blood vessels. Since Asahara et al (11) first detected EPCs in adult peripheral
blood, more findings have indicated that EPCs serve an important
role in endothelium maintenance and thus are involved in
re-endothelialisation and neovascularisation (12,13). A
previous study by our group demonstrated that EPCs were possible
biological components of stem-cell niches and affected biological
processes of MSCs (14). BMs are
major sources of MSCs and EPCs in mice. Therefore, the current
study aims to obtain the two types of cells from murine BM.
Methods for isolation of MSCs and EPCs include
plastic adherence (15,16), density gradient centrifugation
(17,18), immunomagnetic selection (11,19,20) and
flow cytometry sorting (21).
However, no optimal method is available for retrieval of such cells
(22). In addition to fibroblastic
cells, primary cultures derived from BM contain fibroblasts,
macrophages, endothelial cells, adipocytes, hematopoietic stem
cells (HSCs), EPCs and red cells. These cells in BM exhibit
different adherent capacities; in particular macrophages and mature
endothelial cells easily attach to dish wall, followed by
fibroblasts and fibroblastic cells, finally adipocytes, HSCs and
EPCs adhere poorly to dish walls (23,24).
Based on the plastic adherent property, MSCs and EPCs were isolated
simultaneously. Purification of MSCs and EPCs was also conducted
since MSCs differentiate into a trypsin-sensitive population,
whereas EPCs differentiate into a trypsin-resistant population
(25).
The present study aimed to demonstrate an improved
method of plastic adherence to isolate homogenous populations of
MSCs with good proliferation and differentiation capacities.
Furthermore, it was explored whether EPCs could also be obtained
while avoiding the sacrifice of numerous mice.
Materials and methods
Isolation and culture of MSCs and EPCs
derived from BM
A total of 20 male C57BL/6 mice (6–8 weeks old,
25–35 g) were purchased from the Laboratory Animal Center of
Xinjiang Medical University (Urumqi, China). Mice were maintained
under a 12 h light/dark cycle at 25±2°C with 50±5% humidity. Food
and water were available ad libitum. The experimental animal
protocol used in the present study was approved by the Animal
Experimental Ethics Committee of Shihezi University (Shihezi,
China). The mice were euthanized by trained personnel using
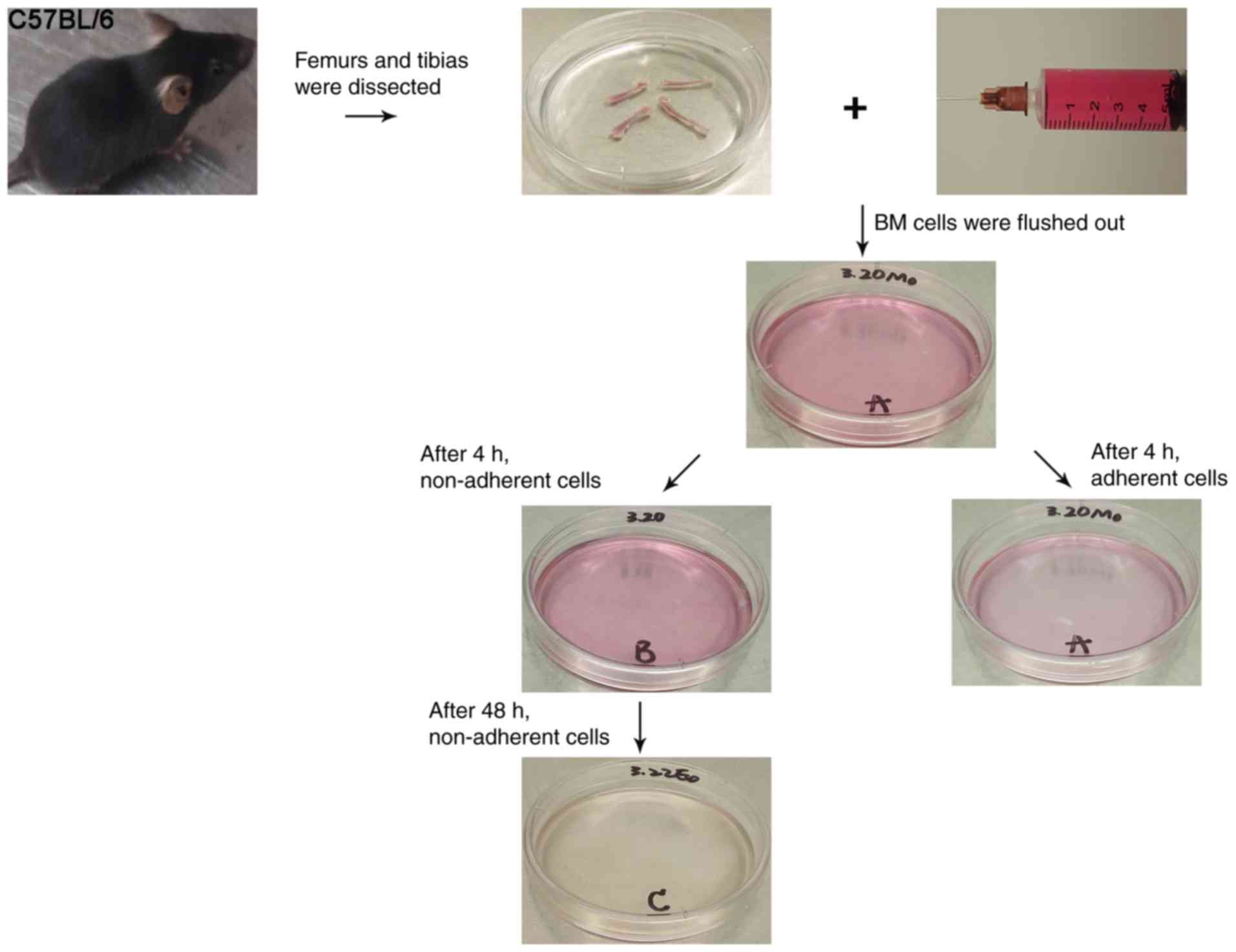
CO2 inhalation. Femurs and tibias were separated, and
muscles and connective tissues were manually removed (Fig. 1). Then, a 5 ml syringe was filled
with complete Dulbecco's modified Eagle's medium (DMEM), which
consisted of low-glucose DMEM (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), 10% foetal bovine serum (FBS; Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA), 100 U/ml penicillin and
100 U/ml streptomycin (both Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany). The ends of the tibias and femurs, inferior to the
medullary cavity, were cut with scissors. The marrow in the
medullary cavity was flushed out by inserting a syringe needle
(27-gauge) attached to the 5 ml syringe into one end of the bones.
Finally, BM cells were filtered into 60 mm plastic culture dishes
through a 200-mesh filter. The culture was maintained at 37°C in a
humidified incubator containing 95% air and 5% CO2.
After 4 h, non-adherent cells that accumulated on the surface of
culture dish were pipetted into a new culture dish, as described
previously (26). Cells in the
original dish were cultured for an additional 8 h, followed by
gradual replacement of the medium with 1.5 ml fresh complete DMEM
to obtain MSCs. Thereafter, this step was repeated every 8 h for 72
h for culture initiation. Cells in the new culture dish were
cultured for 48 h to obtain EPCs, as described previously (18). Then, non-adherent cells from 2 mice
were collected, plated in a 60 mm culture dish coated with human
fibronectin (Gibco; Thermo Fisher Scientific, Inc.) and maintained
in endothelial growth medium (EGM), which contained endothelial
cell ‘basal medium-2, EGM™−2 MV SingleQuots™
(both Lonza Group, Ltd., Basel, Switzerland), 100 U/ml penicillin
and 100 U/ml streptomycin (27).
Following 72 h of culturing, non-adherent cells were removed. The
medium for MSCs and EPCs was replaced every 3–4 days. After 4, 7
and 14 days, cells were visualised under an inverted microscope
(Zeiss Axio Observer; Zeiss AG, Oberkochen, Germany) at
magnification, ×50.
To obtain homogenous populations of MSCs, the MSC
culture was treated with 1 ml StemPro Accutase (Gibco; Thermo
Fisher Scientific, Inc.) for 2 min when 90–100% confluence was
reached. Cells detached within 2 min, then were harvested and
sub-cultured. At 90–100% confluence, the EPC culture was similarly
treated with 1 ml StemPro Accutase to homogenise EPCs. In contrast
to MSCs, a number of cells detached after 2 min, whereas the cells
that had not detached were treated for another 3 min, harvested and
sub-cultured.
Fluorescence-activated cell sorting
(FACS) analysis
Passage 3 cells in complete DMEM were detached using
StemPro Accutase and counted. Then, 2×105 cells in 100
µl buffer containing PBS (Hyclone; GE Healthcare Life Sciences) and
2% FBS (Hyclone; GE Healthcare Life Sciences) were divided into
aliquots in 1.5 ml centrifuge tubes. Cells were stained with
fluorescent isothiocyanate (FITC)-conjugated, phycoerythrin
(PE)-conjugated or PE-Cyanine7-conjugated anti-mouse cluster of
differentiation CD29 (cat. no. 11-0291; dilution, 1:50), CD44 (cat.
no. 12-0441; dilution, 1:160), stem cell antigen-1 (Sca-1; cat. no.
11-5981-82; dilution, 1:100), CD45 (cat. no. 25-0451; dilution,
1:160), CD11b (cat. no. 11-0112-82; dilution, 1:100), CD133 (cat.
no. 11-1331-80; dilution, 1:100) and vascular endothelial growth
factor receptor 2 (VEGFR-2; cat. no. 12-5821-82; dilution, 1:40;
all eBioscience, Thermo Fisher Scientific, Inc.) antibodies in the
dark at 4°C for 30 min. Cells stained with FITC-conjugated (cat.
no. 11-4714-81; dilution, 1:100) or PE-conjugated (cat. no.
12-4724-42; dilution, 1:100) anti-mouse immunoglobulin (Ig)G (both
eBioscience, Thermo Fisher Scientific, Inc.) in the dark at 4°C for
30 min served as controls. Thereafter, cells were pelleted by
centrifugation at 400 × g at 4°C for 5 min. Following two washes
with PBS, cells were examined using the FACSCalibur flow cytometer
(BD Biosciences, Franklin Lakes, NJ, USA) and data were analysed
using FlowJo 7.6 software program (Tree Star, Inc., Ashland, OR,
USA).
After 14 days, cells in EGM were detached, counted
and separate using the aforementioned protocol. The cells were
stained with FITC-conjugated or PE-conjugated anti-mouse Sca-1
(dilution, 1:100), VEGFR-2 (dilution, 1:40) and CD133 (dilution,
1:100; all eBioscience, Thermo Fisher Scientific, Inc.) in the dark
at 4°C for 30 min. Cells stained with FITC-labelled or PE-labelled
anti-mouse IgG served as controls. Cells were pelleted and analysed
using the aforementioned protocol.
Differentiation of MSC assays
Passage 3 cells in complete DMEM were seeded in
6-well plates at 1×105 cells/well. When the cells
reached 60–70% confluence, DMEM complete medium was carefully
aspirated from each well, and 2 ml osteogenic differentiation
medium was added. Osteogenic differentiation medium consisted of
DMEM complete medium, 10 mM β-glycerol phosphate, 50 µM ascorbate
and 10−7 M dexamethasone (all Cyagen Biosciences Inc.,
Guangzhou, China) as described previously (27,28). The
medium was changed twice per week for 3 weeks. Thereafter, cells
were rinsed twice with PBS, fixed with 4% paraformaldehyde for 30
min and stained with alizarin red S for 5 min. Cells were
visualised under an inverted microscope at magnification, ×100.
For adipogenic differentiation, passage 3 cells in
complete DMEM were seeded in 6-well plates at 1×105
104 cells/well. When cells reached 100% confluence or
during the post-confluent stage, complete DMEM was carefully
aspirated from each well and 2 ml adipogenic differentiation medium
was added. The medium consisted of complete DMEM, 200 µM
indomethacin, 10−7 M dexamethasone, 0.5 mM
3-isobutyl-1-methylxanthine and 10 µM insulin (all Cyagen
Biosciences Inc.) as previously described (27,28).
After 3 days, the medium was replaced with complete DMEM. After 24
h, the medium was changed back to adipogenic differentiation
medium. Following 3–5 cycles of induction and maintenance, the
cells were cultured in adipogenic differentiation medium until
lipid droplets were sufficiently large and round. Then, the cells
were fixed with 4% paraformaldehyde for 30 min and stained with oil
red O for 30 min at room temperature. Cells were visualised under
an inverted microscope at magnification, ×50.
For chondrocytic differentiation, passage 3 cells in
complete DMEM were seeded in 6-well plates at 1×105
104 cells/well. When cells reached 60–70% confluence,
complete DMEM was carefully aspirated from each well and 2 ml
chondrocytic differentiation medium was added. Chondrocytic
differentiation medium consisted of DMEM with 1% FBS (Hyclone; GE
Healthcare Life Sciences), 1 mM sodium pyruvate, 50 µM ascorbate,
50 mg/ml proline, 20 ng/ml TGF-β3, 1% ITS supplement and
10−7 M dexamethasone (all Cyagen Biosciences, Inc.) as
previously described (2,28). The medium was changed twice per week
for 3 weeks. Thereafter, cells were rinsed twice with PBS, fixed
with 4% paraformaldehyde for 30 min and stained with alcian blue
for 30 min at room temperature. Cells were visualised under an
inverted microscope at magnification, ×100.
Fluorescent co-staining assay
After 7 days in culture, to examine the presence of
specific scavenger receptors for acetylated low-density lipoprotein
(acLDL) and murine endothelial cell markers, attached cells in EGM
underwent dual binding with
1,1′-dioctadecyl-3,3,3′,3′-tetramethylidocarbocyanine perchlorate
(DiI)-labelled acLDL (Molecular Probes; Thermo Fisher Scientific,
Inc.) and FITC-labelled Bandeiraea simplicifolia lectin I
(BS I; Sigma-Aldrich; Merck KGaA). Cells were initially incubated
in EGM containing 5 µg/ml DiI-acLDL for 4 h at 37°C and then fixed
with 4% paraformaldehyde for 10 min at room temperature. Following
washing with PBS, cells were stained with 10 µg/ml FITC-labelled
BS-I lectin for 1 h at 37°C. Samples were viewed by confocal laser
scanning microscopy (Zeiss LSM 510 Meta; Zeiss AG) at
magnification, ×100. Double-labelled fluorescent cells were
identified as differentiating EPCs.
Tube-like structure formation
assay
A 24-well plate was coated with Matrigel (BD
Biosciences), which was melted into liquid at 4°C overnight.
Subsequently, the plate was placed on ice and incubated for 30 min
at 37°C in a 5% CO2 humidified incubator to allow
solidification of Matrigel. Following 14 days in culture,
6×104 EPCs without any staining were seeded in the plate
and cultured for 6–8 h at 37°C in a 5% CO2 humidified
incubator. Finally, images were randomly captured using an inverted
microscope at magnification, ×200.
Results
Culturing BM cells produce typical
MSCs and EPC-derived endothelial cells
Following 72 h of culture initiation, non-adherent
hematopoietic cells were removed with frequent medium changes.
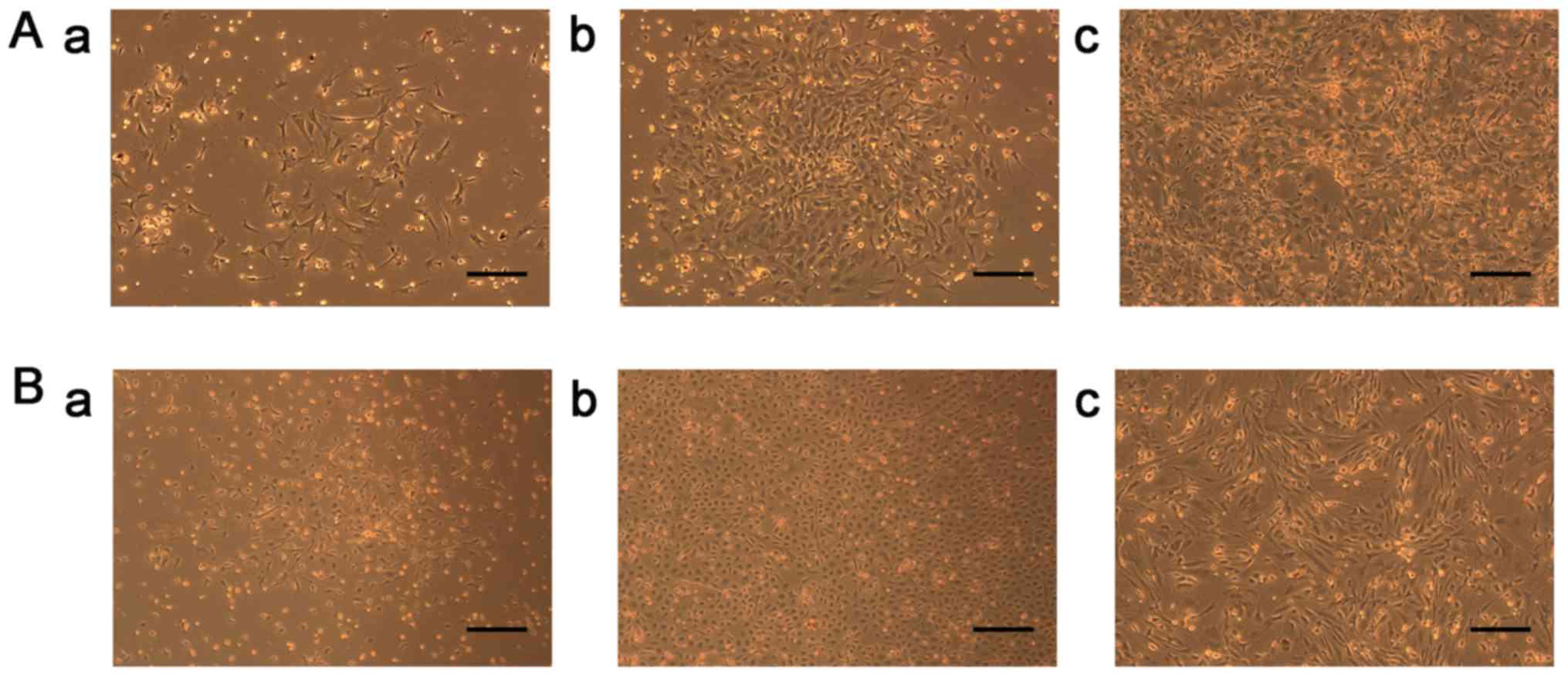
Adherent cells appeared as individual cells; they proliferated and
gradually formed small colonies at approximately day 4 (Fig. 2Aa). During the 7-day culture, typical
colonies of fibroblastic cells appeared (Fig. 2Ab), as described by Ji et al
(29). The number of cellular
colonies with different sizes markedly increased, cells reached
near 100% confluence within 14 days and were triangle in shape
(Fig. 2Ac). After 1 min of enzyme
digestion, numerous cells with triangle-like morphology were
detached (data not shown). With further passages, colonies became
more homogeneous (data not known).
Following 3 days of isolation under
endothelial-specific conditions, cells formed colonies, which were
composed of a centre of round cells with elongated spindle-shaped
cells sprouting at the periphery (Fig.
2Ba). A total of 2 weeks of isolation yielded colonies of
outgrowth cells (late EPCs), which exhibited a ‘cobblestone’
morphology and a monolayer growth pattern at confluence (Fig. 2Bb). Following culture for 21 days,
cells were fusiform, which is typical of EPC-derived endothelial
cells (Fig. 2Bc), as described by
Ingram et al (30).
MSCs and EPCs exhibit typical MSC- and
EPC-markers, respectively, following the novel protocol
Cells were analysed for cell surface antigens by
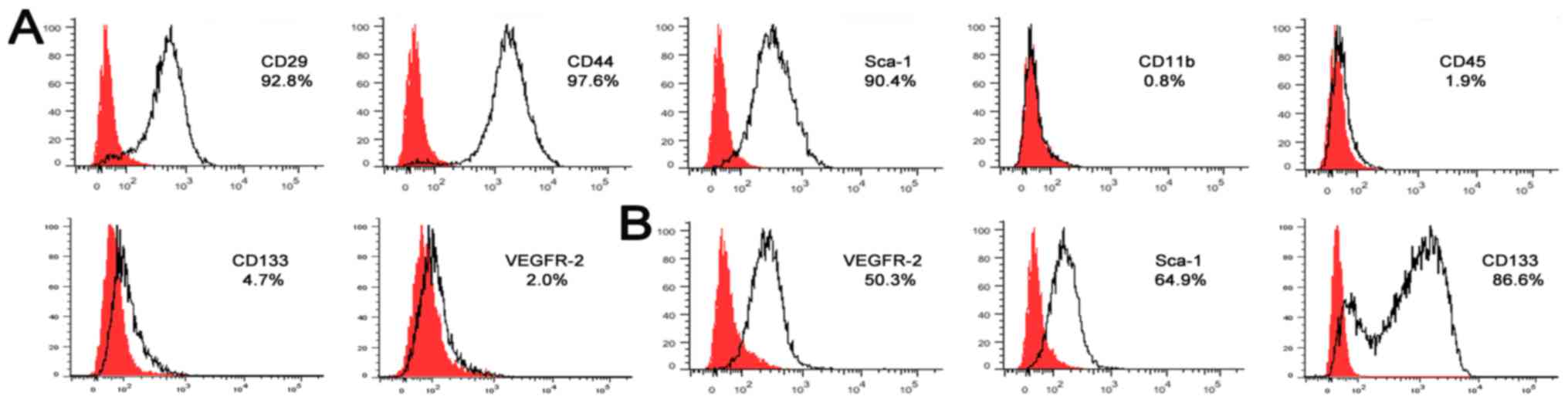
FACS. The results revealed that cells in complete DMEM were
homogenously positive for MSC markers CD29 (92.8%), CD44 (97.6%)
and Sca-1 (90.4%), but negative for hematopoietic markers CD11b
(0.8%) and CD45 (1.9%) (Fig. 3A).
The majority of the cells in complete DMEM did not express CD133
(4.7%) and VEGFR-2 (2.0%).
Following 14 days in the EGM culture, FACS
demonstrated that the cells expressed endothelial cell lineage
antigens VEGFR-2 (50.3%), Sca-1 (64.9%) and CD133 (86.6%) (Fig 3B). These findings confirmed that cells
possessed typical characteristics of EPCs.
MSCs can differentiate into
osteoblasts, adipocytes and chondrocytes
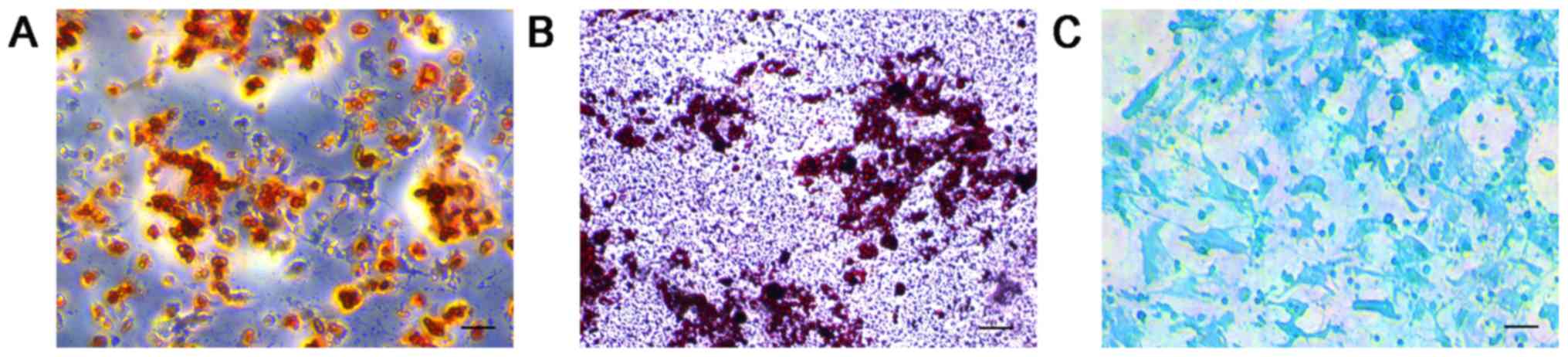
Following 3 weeks of osteogenic induction, calcium
deposits were stained using alizarin red S, demonstrating that MSCs
were undergoing osteogenesis (Fig.
4A). Similarly, following supplementation with adipogenic
differentiation medium for 3 weeks, MSCs stained with oil red O
were positive for adipocyte globules, indicating that they
expressed an adipocyte phenotype (Fig.
4B). The MSCs stained with alcian blue demonstrated an
accumulation of cartilaginous proteoglycans, which suggested that
the cells had differentiated into chondrocytes (Fig. 4C). MSCs cultured in complete DMEM
retained their osteoblastic, adipocytic and chondrocytic
differentiation potentials until passage 10 (data not shown).
EPCs demonstrate endothelial cell
characteristics and are able to form tube-like structures
Following supplementation with EGM for 7 days,
uptake of acLDLs and binding of BS-1 lectin was exhibited in
attached EPCs, demonstrating their endothelial cell characteristics
(Fig. 5A). Tube-like structures were
observed following the culturing of EPCs on Matrigel (Fig. 5B).
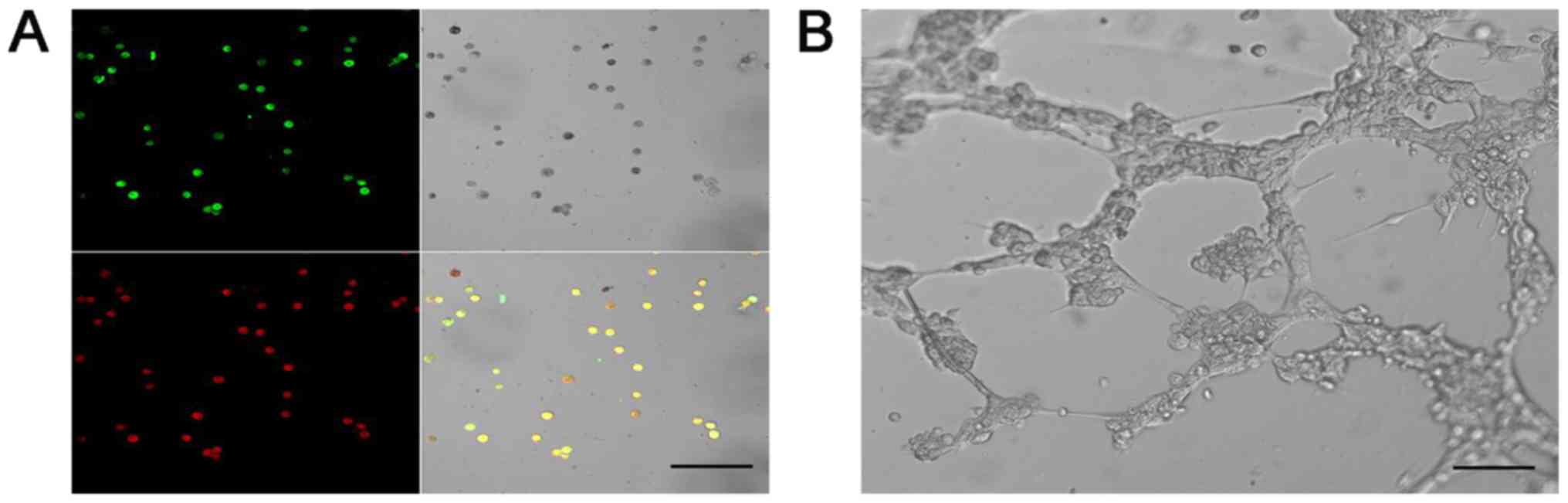 | Figure 5.EPCs demonstrate endothelial cell
characteristics and are able to form tube-like structures. (A)
Fluorescence confocal microscopy illustrates the uptake of
DiI-acLDL (red), bonding of FITC BS-I lectin (green), double
staining with DiI-acLDL, FITC BS-I (yellow), and the cellular
morphology in bright field (grey) after 7 days of culturing. Scale
bar, 100 µm. (B) EPCs after culturing on Matrigel for 8 h. Scale
bar, 50 µm. EPCs, endothelial progenitor cells; DiI-acLDL,
1,1′-dioctadecyl-3,3,3′,3′-tetramethylidocarbocyanine
perchlorate-labelled acetylated low-density lipoprotein;
FITC-labelled BS-I lectin, fluorescent isothiocyanate-labelled
Bandeiraea simplicifolia-I lectin. |
Discussion
Previous studies have revealed that highly purified
MSCs and EPCs can be obtained by employing three methods: Density
gradient centrifugation (17,18),
immunomagnetic selection (11,19,20) and
flow cytometry sorting (21).
However, these three types of experimental process significantly
affect cellular activity. There are a small number of BM cells in
small animals, such as small, 1-week-old mice, and fibroblastic
cells comprise 0.001–0.01% BM cells (22). Thus, technical difficulties may arise
during isolation of fibroblastic cells from mice. By contrast,
cells can easily be obtained based on their plastic adherence
characteristics, but this method cannot be used to retrieve pure
cells (31). In the present study,
pure MSCs and EPCs were simultaneously isolated and obtained from
murine BM. The protocol employed in the current study featured
three important points: Firstly, the medium was frequently changed
to prevent the adherence of non-MSCs to the culture dish; secondly,
the attachment time for MSCs and EPCs differed; thirdly, the
duration of trypsinisation was well controlled.
MSCs resembled fibroblasts in terms of their
morphology and due to their colony formation; these characteristics
were identified in MSCs from numerous species, including humans
(32), rats (33), mice (34) and rabbits (35). However, the expandability of MSCs
in vitro varied significantly among different species, and
different methodologies for isolation and plating of cells. In the
present study, isolated cells were identified as MSCs and EPCs on
the two bases: i) MSCs are fibroblast-like clonogenic cells
(colony-forming unit-fibroblast) with high replicative capacity
in vitro (36) and ii) EPCs
exhibit the ability to form colonies or small clusters of cells
based on differences in proliferative and/or differentiation
potentials (37). MSCs and EPCs
formed colonies, demonstrating their stem and/or progenitor cell
characteristics.
These observations were further supported by
detecting expression levels of cell surface antigens. Surface
antigens for murine-derived MSCs are not well defined (38). Generally, MSCs are characterized by
an immunophenotype depicted as positive for Sca-1, CD29 and CD44,
and negative for CD11b and CD45 because CD11b is expressed by
monocytes, granulocytes and natural killer cells, and CD45 is
expressed in all lymphohematopoietic cell lineages (34). The results of FACS analysis revealed
that the cells scarcely expressed surface markers CD11b and CD45,
but expressed high levels of CD29, CD44 and Sca-1. This finding is
consistent with previously published data on MSC surface markers
and implies that the proposed method obtains pure MSCs. Surface
markers for murine-derived EPCs also remain unclear (39). It was hypothesised that CD133 is a
reliable phenotypic marker that can be used to isolate bona fide
EPCs (40). In addition, VEGFR-2 is
an endothelial-specific marker and Sca-1 is expressed only in
murine species (27,41). Thus, VEGFR-2, Sca-1 and CD133 are
considered surface markers of murine-derived EPCs. In the present
study, CD133 and VEGFR-2 expression was also evaluated in MSCs in
the current study to determine if EPCs differentiated from MSCs.
The results revealed that CD133 and VEGFR-2 expression was low in
MSCs, which may disprove the EPC differentiation from MSCs. FACS
analysis indicated that EPCs expressed VEGFR-2, Sca-1 and
CD133.
Finally, functional analysis was performed for
prospective identification and further characterisation of
progenitors. Multilineage differentiation potential is considered
an important quality of MSCs (42).
In the present study, cells cultured in complete DMEM successfully
differentiated into osteogenic, adipogenic and chondrogenic
lineages in the presence of tissue-specific induction media. EPCs
cultured in EGM medium incorporated acLDL and bound BS-1 lectin,
which characterise endothelial function.
In summary, compared with other approaches for
separating MSCs and EPCs, plastic adherence is a simple and
efficient method. Through the principle of adhesion, homogeneous
MSCs and EPCs can be simultaneously obtained as potential resources
for the basic study of stem cell therapy and regenerative
medicine.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81760570
and 31271458), and the Science and Technology Program of Xinjiang
Production and Construction Corps (grant no. 2014AB047). It was
also partially supported by grants from the Basic Research Program
of Xinjiang Production and Construction Corps (grant no.
2016AG019), and Shihezi University high-level personnel scientific
research project (grant no. RCZX201538).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XWa isolated cells, and was a major contributor in
writing the manuscript. ZZ analyzed and interpreted the results of
FACS. HZ performed differentiation of MSC assays. JH conducted the
fluorescent co-staining assay. WF performed the tube-like structure
formation assay. MZ fed the mice and was involved in the isolation
of MSCs. JG participated in isolating EPCs. JX assisted with the
differentiation of MSC assays. QG prepared test reagents and was
involved in writing the Materials and methods section. XC made
substantial contributions to the acquisition of FACS data,
including the selection of fluorescent antibodies and operation of
the test. XWu conceived and designed the experiments. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental animal protocol used in the present
study was approved by the Animal Experimental Ethics Committee of
Shihezi University (Shihezi, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sabry D, Noh O and Samir M: Comparative
evaluation for potential differentiation of endothelial progenitor
cells and mesenchymal stem cells into endothelial-like cells. Int J
Stem Cells. 9:44–52. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yin C, Liang Y, Zhang J, Ruan G, Li Z,
Pang R and Pan X: Umbilical cord-derived mesenchymal stem cells
relieve hindlimb ischemia through enhancing angiogenesis in tree
shrews. Stem Cells Int. 2016:1–9. 2016. View Article : Google Scholar
|
|
3
|
Qadura M, Terenzi DC, Verma S, AL-Omran M
and Hess DA: Concise review: Cell therapy for critical limb
ischemia: An integrated review of preclinical and clinical studies.
Stem Cells. 36:161–171. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jameel MN and Zhang J: Stem cell therapy
for ischemic heart disease. Antioxid Redox Signal. 13:1879–1897.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Phillips MI, Tang YL and Pinkernell K:
Stem cell therapy for heart failure: The science and current
progress. Future Cardiol. 4:285–298. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ball SG, Shuttleworth CA and Kielty CM:
Mesenchymal stem cells and neovascularization: Role of
platelet-derived growth factor receptors. J Cell Mol Med.
11:1012–1030. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kawamoto A, Asahara T and Losordo DW:
Transplantation of endothelial progenitor cells for therapeutic
neovascularization. Cardiovasc Radiat Med. 3:221–225. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Friedenstein AJ, Deriglasova UF, Kulagina
NN, Panasuk AF, Rudakowa SF, Luriá EA and Ruadkow IA: Precursors
for fibroblasts in different populations of hematopoietic cells as
detected by the in vitro colony assay method. Exp Hematol. 2:83–92.
1974.PubMed/NCBI
|
|
9
|
Caplan AI: Mesenchymal stem cells. J
Orthop Res. 9:641–650. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu S, de Becker A, van Camp B,
Vanderkerken K and van Riet I: An improved harvest and in vitro
expansion protocol for murine bone marrow-derived mesenchymal stem
cells. J Biomed Biotechnol. 2010:1059402010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Asahara T, Murohara T, Sullivan A, Silver
M, Van Der Zee R, Li T, Witzenbichler B, Schatteman G and Isner JM:
Isolation of putative progenitor endothelial cells for
angiogenesis. Science. 275:964–967. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rehman J, Li JL, Orschell CM and March KL:
Peripheral blood ‘endothelial progenitor cells’ are derived from
monocyte/macrophages and secrete angiogenic growth factors.
Circulation. 107:1164–1169. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kawamoto A, Gwon HC, Iwaguro H, Yamaguchi
JI, Uchida S, Masuda H, Silver M, Ma H, Kearney M, Isner JM and
Asahara T: Therapeutic potential of ex vivo expanded endothelial
progenitor cells for myocardial ischemia. Circulation. 103:634–637.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang H, Xian L, Lin Z, Yang C, Zhang M,
Feng W, Peng X, Chen X and Wu X: Endothelial progenitor cells as a
possible component of stem cell niche to promote self-renewal of
mesenchymal stem cells. Mol Cell Biochem. 397:235–243. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Thomas MG, Stone L, Evill L, Ong S, Ziman
M and Hool L: Bone marrow stromal cells as replacement cells for
Parkinson's disease: Generation of an anatomical but not functional
neuronal phenotype. Tansl Res. 157:56–63. 2011. View Article : Google Scholar
|
|
16
|
Nadri S, Soleimani M, Hosseni RH, Massumi
M, Atashi A and Izadpanah R: An efficient method for isolation of
murine bone marrow mesenchymal stem cells. Int J Dev Biol.
51:723–729. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Eslaminejad MB, Nikmahzar A, Taghiyar L,
Nadri S and Massumi M: Murine mesenchymal stem cells isolated by
low density primary culture system. Dev Growth Differ. 48:361–370.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hill JM, Zalos G, Halcox JPJ, Schenke WH,
Waclawiw MA, Quyyumi AA and Finkel T: Circulating endothelial
progenitor cells, vascular function, and cardiovascular risk. N
Engl J Med. 348:593–600. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu J and Chao B: MRI-based visualization
of iron-labeled CD133+ human endothelial progenitor cells. Bio
Trace Elem Res. 126:83–91. 2008. View Article : Google Scholar
|
|
20
|
Kimura T, Boehmler AM, Seitz G, Kuçi S,
Wiesner T, Brinkmann V, Kanz L and Möhle R: The sphingosine
1-phosphate receptor agonist FTY720 supports CXCR4-dependent
migration and bone marrow homing of human CD34+
progenitor cells. Blood. 103:4478–4486. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen WC, Saparov A, Corselli M, Crisan M,
Zheng B, Péault B and Huard J: Isolation of blood-vessel-derived
multipotent precursors from human skeletal muscle. J Vis Exp.
21:e511952014.
|
|
22
|
Polisetti N, Chaitanya VG, Babu PP and
Vemuganti GK: Isolation, characterization and differentiation
potential of rat bone marrow stromal cells. Neurol India.
58:201–208. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Piersma AH, Brockbank KG, Ploemacher RE,
van Vliet E, Brakel-van Peer KM and Visser PJ: Characterization of
fibroblastic stromal cells from murine bone marrow. Exp Hematol.
13:237–243. 1985.PubMed/NCBI
|
|
24
|
Fang L, Du MJ, Li Y, Lin L, Zhang C and
Zhao ZM: Using the differential adhesion method to isolate and
culture mesenchymal stem cells and endothelial progenitor cells
from rat bone marrow. Biomed Res. 27:377–382. 2016.
|
|
25
|
Walia V and Elble RC: Enrichment for
breast cancer cells with stem/progenitor properties by differential
adhesion. Stem Cells Dev. 19:1175–1182. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Soleimani M and Nadri S: A protocol for
isolation and culture of mesenchymal stem cells from mouse bone
marrow. Nat Protoc. 4:102–106. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brunt KR, Hall SR, Ward CA and Melo LG:
Endothelial progenitor cell and mesenchymal stem cell isolation,
characterization, viral transduction. Methods Mol Med. 139:197–210.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kita K, Gauglitz GG, Phan TT, Herndon DN
and Jeschke MG: Isolation and characterization of mesenchymal stem
cells from the sub-amniotic human umbilical cord lining membrane.
Stem Cells Dev. 19:491–501. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ji M, Bai C, Li L, Fan Y, Ma C, Li X and
Guan W: Biological characterization of sheep kidney-derived
mesenchymal stem cells. Exp Ther Med. 12:3963–3971. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ingram DA, Mead LE, Tanaka H, Meade V,
Fenoglio A, Mortell K, Pollok K, Ferkowicz MJ, Gilley D and Yoder
MC: Identification of a novel hierarchy of endothelial progenitor
cells using human peripheral and umbilical cord blood. Blood.
104:2752–2760. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Phinney DG, Kopen G, Isaacson RL and
Prockop DJ: Plastic adherent stromal cells from the bone marrow of
commonly used strains of inbred mice: Variations in yield, growth,
and differentiation. J Cell Biochem. 72:570–585. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee HS, Huang GT, Chiang H, Chiou LL, Chen
MH, Hsieh CH and Jiang CC: Multipotential mesenchymal stem cells
from femoral bone marrow near the site of osteonecrosis. Stem
Cells. 21:190–199. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tang YL, Zhao Q, Qin X, Shen L, Cheng L,
Ge J and Phillips MI: Paracrine action enhances the effects of
autologous mesenchymal stem cell transplantation on vascular
regeneration in rat model of myocardial infarction. Ann Thorac
Surg. 80:229–237. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kinnaird T, Stabile E, Burnett MS, Shou M,
Lee CW, Barr S, Fuchs S and Epstein SE: Local delivery of
marrow-derived stromal cells augments collateral perfusion through
paracrine mechanisms. Circulation. 109:1543–1549. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tan SL, Ahmad TS, Selvaratnam L and
Kamarul T: Isolation, characterization and the multi-lineage
differentiation potential of rabbit bone marrow-derived mesenchymal
stem cells. J Anat. 222:437–450. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Afanasyev BV, Elstner E and Zander AR:
A.J. Friedenstein, founder of the mesenchymeal stem cell concept.
Cell Ther Transplant. 1:35–38. 2009.
|
|
37
|
Ingram DA, Caplice NM and Yoder MC:
Unresolved questions, changing definitions, and novel paradigms for
defining endothelial progenitor cells. Blood. 106:1525–1531. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
da Silva Meirelles L, Caplan AI and Nardi
NB: In search of the in vivo identity of mesenchymal stem cells.
Stem Cells. 26:2287–2299. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Aragona CO, Imbalzano E, Mamone F, Cairo
V, Lo Gullo A, D Ascola A, Sardo MA, Scuruchi M, Basile G, Saitta A
and Mandraffino G: Endothelial progenitor cells for diagnosis and
prognosis in cardiovascular disease. Stem Cells Int.
2016:80437922016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rafii S and Lyden D: Therapeutic stem and
progenitor cell transplantation for organ vascularization and
regeneration. Nat Med. 9:702–712. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao X, Qian D, Wu N, Yin Y, Chen J, Cui B
and Huang L: The spleen recruits endothelial progenitor cell via
SDF-1/CXCR4 axis in mice. J Recept Signal Transduct Res.
30:246–254. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Park JR, Kim E, Yang J, Lee H, Hong SH,
Woo HM, Park SM, Na S and Yang SR: Isolation of human dermis
derived mesenchymal stem cells using explants culture method:
Expansion and phenotypical characterization isolation of human
dermis derived mesenchymal stem cells using explants culture
method: Expansion and phenotypical characterization. Cell Tissue
Bank. 16:209–218. 2015. View Article : Google Scholar : PubMed/NCBI
|