Introduction
Allergic rhinitis (AR) is characterized by the
inflammation of the nasal mucosa and is one of the most common
chronic diseases in the world, with its prevalence rapidly
increasing over the past few decades (1). AR induces inflammation of the upper
respiratory tract, which is associated with mediators released by
several types of hypersensitive immune cells, including antigen
presenting cells, eosinophils, B cells and mast cells (2). Many patients with AR often exhibit
complications including chronic sinusitis and asthma (3). AR seriously affects the lives of
patients as the disease greatly increases the familial and
socio-economic burden (4). There are
several commonly used AR treatments, including intranasal steroids,
antihistamines, leukotriene receptor antagonists and immunotherapy
(5,6). However, patients with AR always have
unsatisfactory results (7). It is
therefore necessary to assess novel and effective AR treatments
(8).
Within AR, genetic and environmental factors work in
congruence (9). The consensus is
that AR is an immunoglobulin E (IgE)-mediated specific type I
hypersensitivity reaction, which is induced by the imbalance of
T-helper (Th) 1 and Th2 immune responses in the body, and the nasal
mucosal Th2 immune response (10).
Regulating the balance of Th1 and Th2 immune responses therefore
serve preventive effects on AR. In 1988, Hashimoto et al
(11) identified a Toll gene in
Drosophila and in 1996, the Toll gene was revealed to serve
a role in Drosophila immunity (12). Medzhitov et al (13) then identified toll-like receptors
(TLRs) in humans and mammals. TLRs are pattern recognition
receptors that serve a very important role in innate and acquired
immune responses (14,15). TLRs participate in the innate immune
response but also affect the type and intensity of the acquired
immune response, stimulate immune cells to synthesize immune
factors and regulate the differentiation of T cells (16). Within the TLR family, the most
studied is TLR4, which localizes to the cell membrane and the
cytoplasm and is assessed primarily in immune cells (17). TLR4 is activated by and recognizes,
bacterial lipopolysaccharide (LPS), which is the main molecular
component of the cell wall in gram-negative bacteria (18,19).
Upon cell membrane receptor dimerization, the TLR4 receptor system
initiates a cascade of protein-protein interactions, resulting in
the production of pro-inflammatory cytokines and interferons
(17,20). These events initiate the inflammation
and immune response (17,20,21).
When TLR4 binds to its ligand, it induces Th0 cells to
differentiate into Th2 cells and therefore promotes the occurrence
of Th2-associated allergic diseases (22–24).
Therefore, TLR4 may serve an important role in the pathogenesis of
AR. A previous study has revealed that TLR4 is highly expressed in
the nasal mucosa of patients with AR (25). However, the role of TLR4 in AR
remains unclear.
The aim of the current study was to investigate the
precise role and molecular mechanism of TLR4 in the mouse model of
AR and to explore.
Materials and methods
Ovalbumin (OVA)-induced AR
establishment
A total of 40, 6-week-old BALB/c mice (~20 g; 20
male and 20 female) were obtained from Charles River Laboratories,
Inc. Mice were maintained under a 12 h dark/light cycle, 20±1°C
room temperature, and 55±5% humidity with free access to food and
water. All animal experiments were performed in accordance with the
protocol approved by the Care and Use of Laboratory Animals
Committee. The current study was approved by the Committee on the
Use and Care of Animals of Taizhou Central Hospital (Taizhou
University Hospital, Taizhou, China). Mice were randomly divided
into four groups (n=10): A control group; an AR group; an
AR+control-short hairpin RNA (shRNA) group and an AR+TLR4-shRNA
group. The AR mouse model was constructed as previously described
(26). Briefly, mice were sensitized
with an intraperitoneal injection of 25 µg OVA and 2 mg aluminum
hydroxide (Sigma-Aldrich; Merck KGaA) on days 0, 7 and 14 to
promote primary sensitization. One week after the last
intraperitoneal injection, mice were intranasally challenged with
3% OVA daily for a week for secondary immunization.
Intranasal administration of
TLR4-shRNA
A total of 20 µl control-shRNA (cat. no. sc-108080;
Santa Cruz Biotechnology, Inc.) or 20 µl TLR4-shRNA (cat. no.
sc-40261-v; Santa Cruz Biotechnology, Inc.) was intranasally
administrated to mice 3 h prior to every daily OVA challenge (once
a day) on days 28–34. AR group mice were treated with 20 µl saline
3 h prior to every daily OVA challenge (once a day) on days
28–34.
Evaluation of nasal symptoms
For 10 min, after the last administration of OVA
challenge, the frequency of sneezing and nose friction (scratching
of the nose by the mouse) in mice was calculated to assess early
allergic reactions.
Inflammation cell counting (27)
Inflammation cells (leucocytes, eosinophils,
neutrophils and lymphocytes) in the nasal lavage fluid (NLF) were
counted. Cells were re-suspended in NLF with 1 ml 100 mmol/l PBS
and 1% BSA (cat. no. ST023; Beyotime Institute of Biotechnology). A
hemocytometer (Mindray 3000; Shenzhen Mindray Bio-Medical
Electronics Co., Ltd.) was subsequently used to count the number of
leucocytes. Wright's-Giemsa staining was then performed at 37°C for
20 min to detect eosinophil, neutrophil and lymphocyte numbers
under a light microscope at a magnification, ×200.
ELISA
After the last TLR4-shRNA or control-shRNA
administration, the blood and NLF samples of mice were collected.
Serum was harvested from blood samples and NLF supernatant was
obtained by centrifugation for 10 min at 1,600 × g and 4°C; the
samples were then stored at −80°C until use. To determine the
protein levels of OVA-specific IgE (cat. no. 439807-1; BioLegend,
Inc.), eosinophil cation protein (ECP; cat. no. ABB-KTE71563-48T;
Abbkine Scientific Co., Ltd.), leukotriene C4 (LTC4; cat. no.
E-EL-M0753km-1; Shanghai Zhenyu Chemical Technology Co., Ltd.) and
prostaglandin D2 (PGD2; cat. no. ABB-KTE70766-48T; Abbkine
Scientific Co., Ltd.) in the plasma and NLF samples of mice from
different groups, ELISA was performed. The secretion of several
pro-inflammatory factors and anti-inflammatory factors including
interleukin (IL)-4 (cat. no. ab221833), IL-5 (cat. no. ab204523),
IL-13 (cat. no. ab219634), IL-17 (cat. no. ab100702), tumor
necrosis factor alpha (TNF-α; cat. no. ab208348), interferon-γ
(IFN-γ; cat. no. ab252352) and IL-2 (cat. no. ab223588; all Abcam)
in mice serum were assessed using an ELISA kit according to
manufacturer's protocol. Analysis of the ELISA detection limit for
each protein revealed that no samples were below the detection
limit.
Reverse transcription-quantitative PCR
(RT-qPCR)
An RNeasy Mini kit (Qiagen, Inc.) was used to
extract total RNA from nasal mucosa cells, according to the
manufacturer's protocol. The PrimeScript First Strand cDNA
Synthesis kit (Takara Biotechnology Co., Ltd.) was utilized for
cDNA synthesis as per the manufacturer's protocol. Synthesized
cDNAs were analyzed using qPCR with SYBR RT-PCR kit (Takara Bio,
Inc.) on the MiniOpticonTM RT PCR System (Bio-Rad Laboratories,
Inc.). The amplification conditions were as follows: 98°C for 1
min, followed by 40 cycles at 98°C for 10 sec, 56°C for 20 sec and
72°C for 30 sec. Primer sequences were as follows: TLR4, forward
5′-CCTGACACCAGGAAGCTTGAA-3′ and reverse
5′-TCTGATCCATGCATTGGTAGGT-3′; GAPDH, forward
5′-CTTTGGTATCGTGGAAGGACTC-3′ and reverse
5′-GTAGAGGCAGGGATGATGTTCT-3′. Relative gene expression levels were
calculated using the 2−ΔΔCq method (28). GAPDH was used as the internal
control. All experiments were performed in triplicate.
Western blot analysis
Total nasal mucosa proteins were extracted from mice
using an M-PER Mammalian Protein Extraction Reagent (ThermoFisher
Scientific, Inc.) according to manufacturer's protocol. Protein
concentrations were measured via a BCA kit according to
manufacturer's protocol. Protein samples (30 mg/lane) were
separated by 10% SDS-PAGE and subsequently transferred onto PVDF
membranes (Merck KGaA). Membranes were blocked in 5% skim milk at
room temperature for 2 h and then incubated with the following
primary antibodies: Phosphorylated (p)-p65 (cat. no. 3033), p65
(cat. no. 8242), TLR4 (cat. no. 14358) and β-actin (cat. no. 4970;
all dilution: 1:1,000; Cell Signaling Technology, Inc.), at 4°C
overnight. After membranes were washed five times with TBST, they
were incubated with a horseradish-peroxidase-conjugated anti-rabbit
secondary antibody (cat no. 7074; 1:5,000; Cell Signaling
Technology, Inc.) for 1 h at room temperature. Proteins were
detected using SignalFire™ Plus ECL Reagent (cat. no. 12630; Cell
Signaling Technology, Inc.) and imaged. Band density was quantified
using Gel-Pro Analyzer densitometry software (version 6.3; Media
Cybernetics, Inc.).
Statistical analyses
All experiments were repeated three times. Data were
presented as the mean ± standard deviation. Data analyses were
performed using SPSS 18.0 statistical software (SPSS Inc.). A
one-way ANOVA, followed by a Tukey's test, was used to measure the
significance between groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
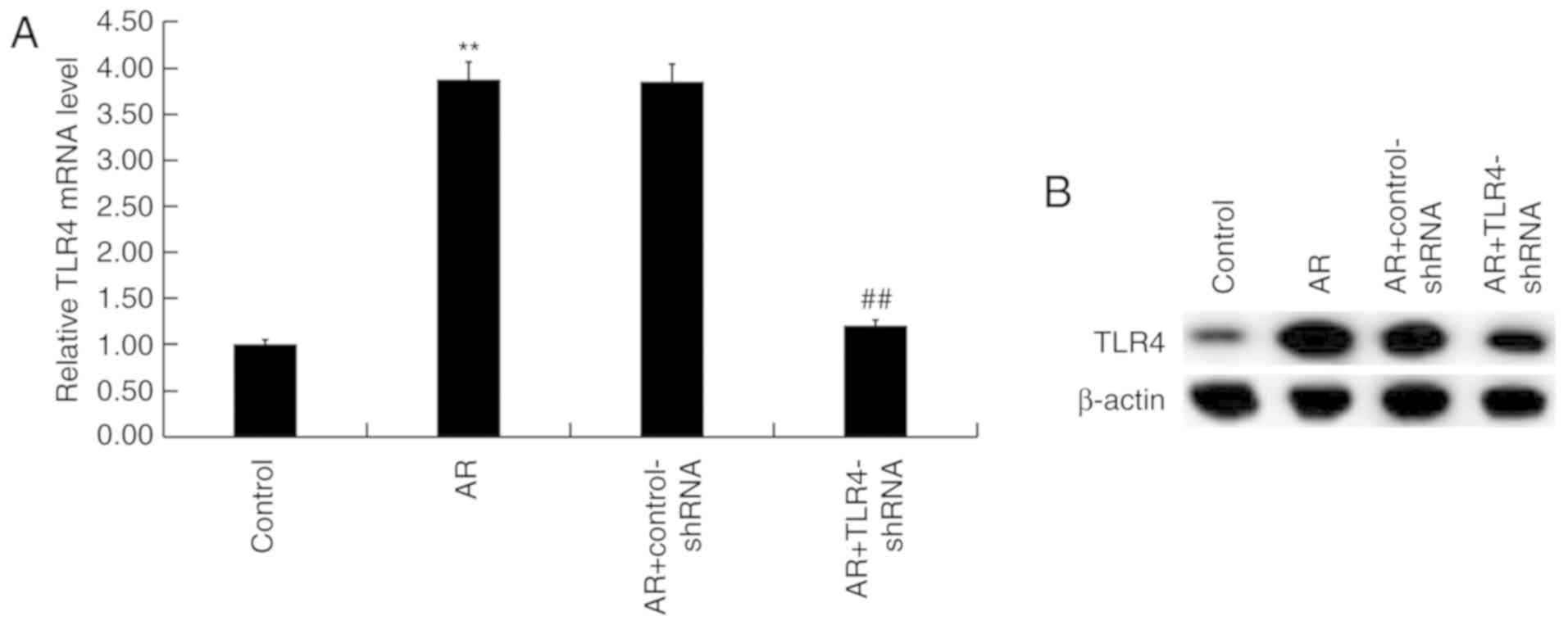
TLR4 is upregulated in the nasal
mucosa of AR mice
mRNA and protein levels of TLR4 were detected in the
nasal mucosa of AR mice using RT-qPCR and western blot analysis.
TLR4 mRNA levels were significantly upregulated in AR mice compared
with the control group, whereas TLR4-shRNA significantly reduced
levels compared with AR mice (Fig.
1A). Similarly, protein levels of TLR4 were markedly
upregulated in AR mice compared with the control group, whereas
TLR4-shRNA markedly reduced levels compared with the AR group
(Fig. 1B).
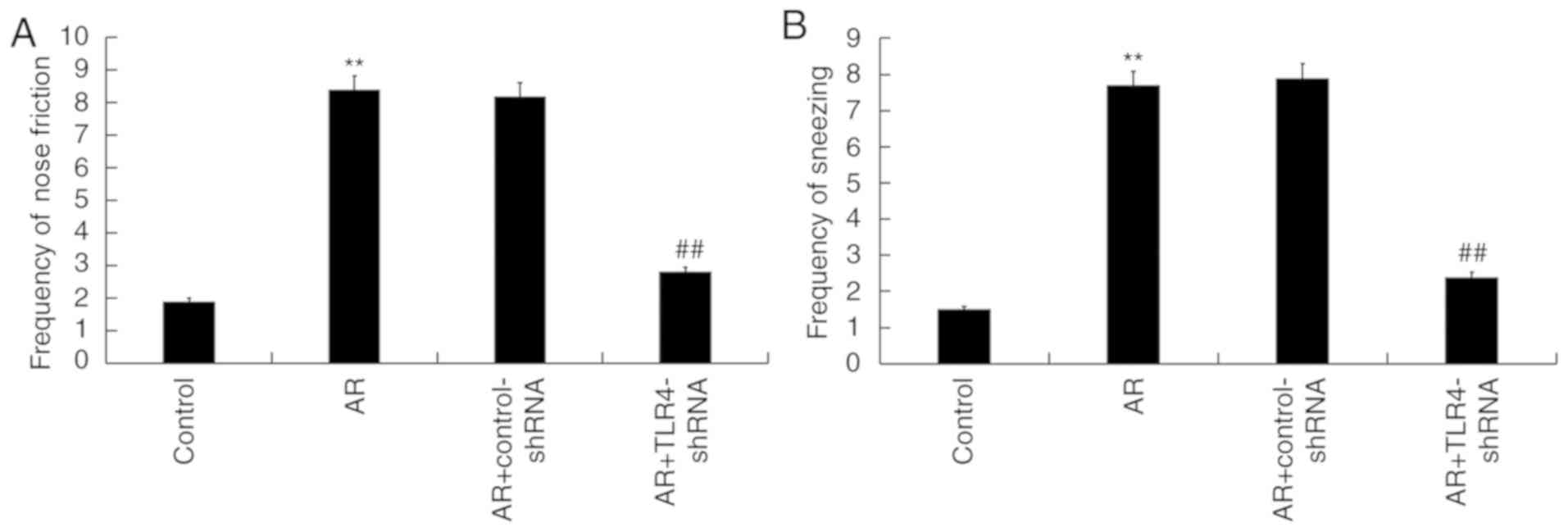
TLR4 downregulation alleviates AR
symptoms in mice
To assess the role of TLR4 in the regulation of AR,
TLR4-shRNA was administered into the nostril daily for 7 days, 3 h
before OVA challenge on days 28–34. The frequency of sneezing and
nose friction were detected after the last administration of OVA
challenge. Compared with the control group, OVA induction
significantly increased the frequency of nose friction and
sneezing, while TLR4-shRNA administration significantly reduced the
frequency of each when compared with the AR group (Fig. 2A and B).
TLR4 downregulation decreases allergic
inflammatory cytokine and OVA-specific IgE levels in AR mice
To assess the underlying mechanisms of TLR4-shRNA in
AR, levels of allergic inflammatory cytokines and immunoglobulins,
including OVA-specific IgE, ECP, LTC4 and PGD2, were determined in
the serum and NLF of mice. Enhanced levels of OVA-specific IgE,
LTC4 and ECP in the serum of AR mice were reduced by TLR4-shRNA
administration (Fig. 3A-C). The
upregulated OVA-specific IgE, LTC4 and PGD2 in the NLF of the mice
also decreased in the TLR4-shRNA treatment (Fig. 3D-F).
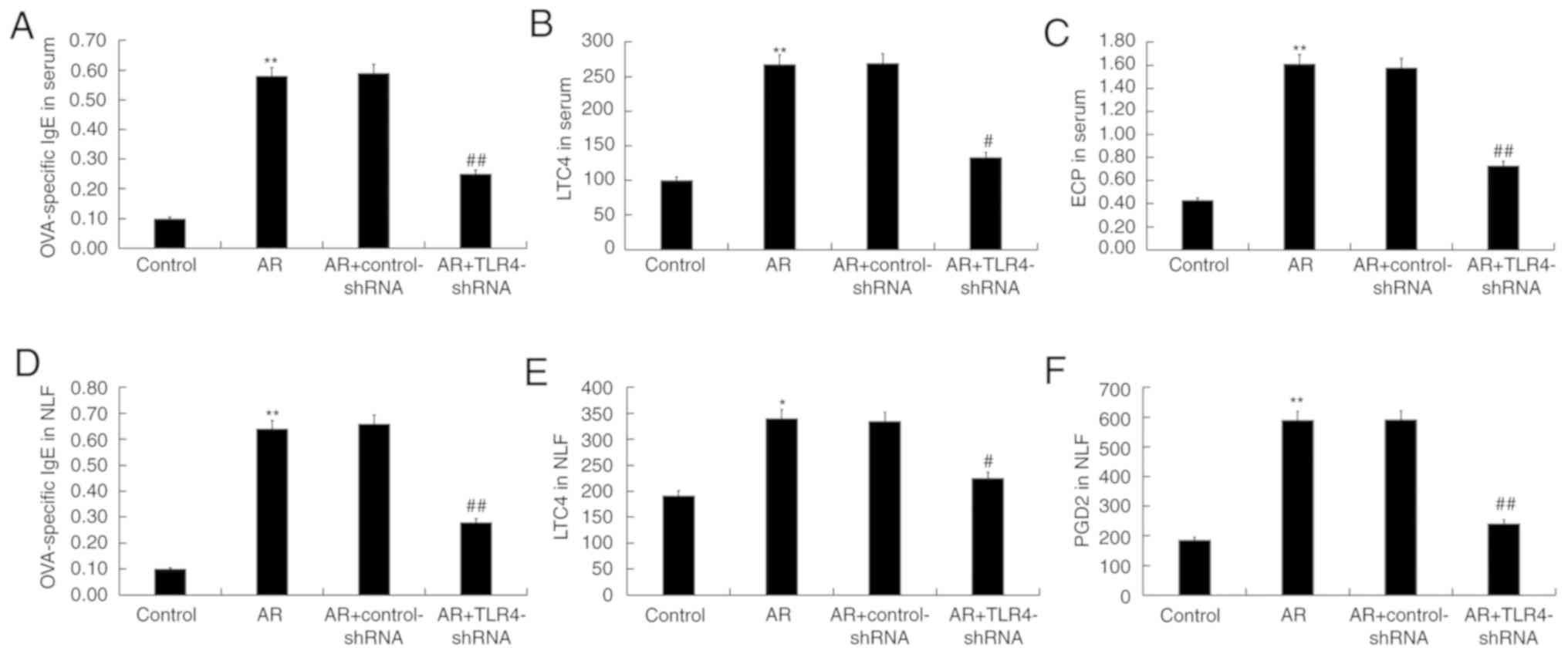 | Figure 3.TLR4 inhibition decreased
OVA-specific (A) IgE, (B) LTC4 and (C) ECP in serum, and
OVA-specific (D) IgE, (E) LTC4 and (F) PGD2 in the NLF of AR mice.
Data are presented as the mean ± standard deviation. *P<0.05,
**P<0.01 vs. the control group; #P<0.05,
##P<0.01 vs. the AR group. TLR4, toll-like receptor
4; OVA, ovalbumin; IgE, immunoglobulin E; LTC4, leukotriene C4;
ECP, eosinophil cation protein; PGD2, prostaglandin D2; NLF, nasal
lavage fluid; AR, allergic rhinitis; shRNA, short hairpin RNA. |
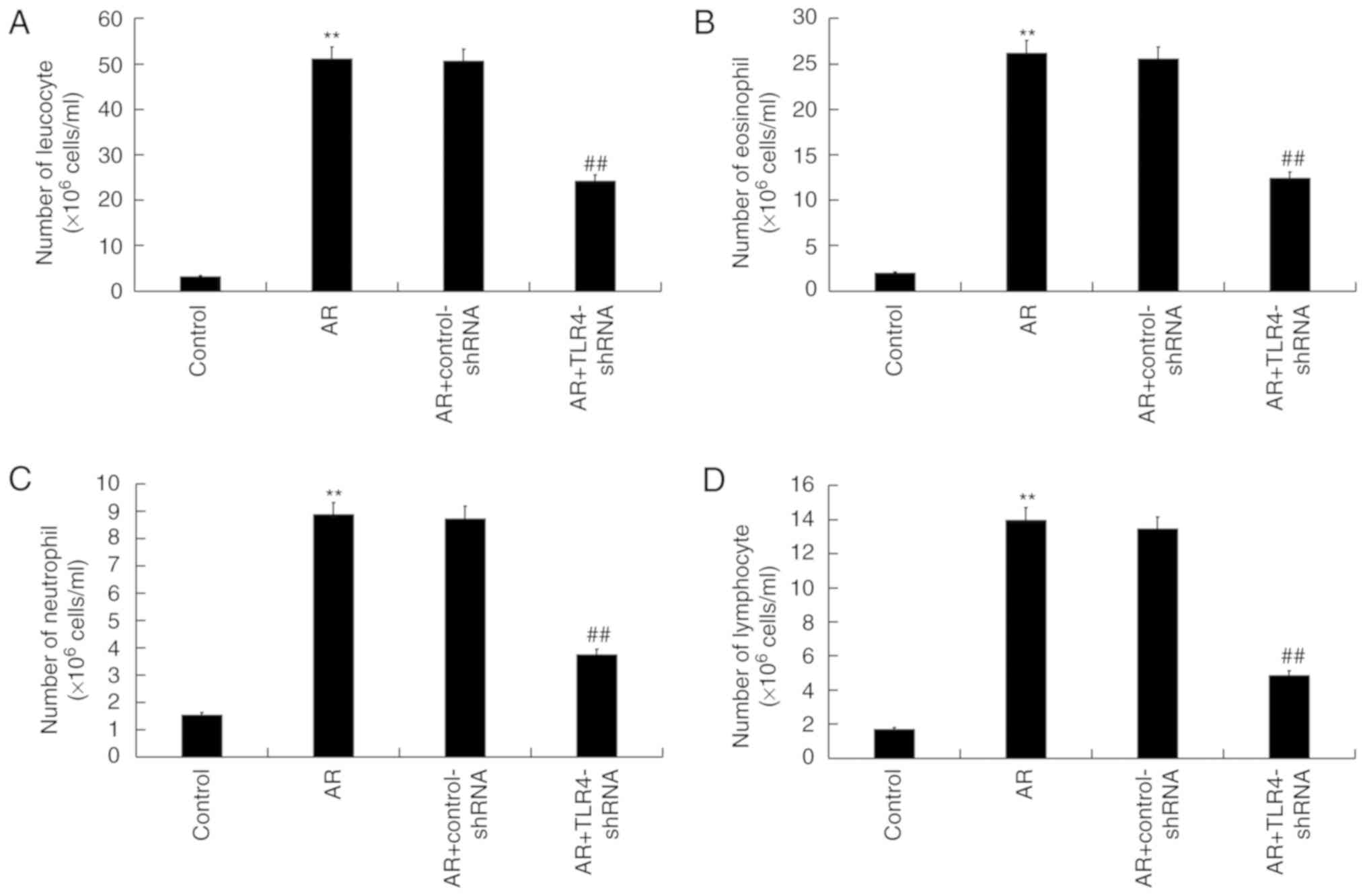
TLR4 downregulation decreases
inflammatory cell numbers in the NLF of AR mice
Compared with the control group, the total number of
cell leucocytes (Fig. 4A),
eosinophils (Fig. 4B), neutrophils
(Fig. 4C) and lymphocytes (Fig. 4D) were significantly increased in AR
mice, and were markedly reduced by TLR4-shRNA treatment.
TLR4 downregulation affects
inflammatory cytokine secretion in the serum of AR mice
Pro-inflammatory factors and anti-inflammatory
factors, including IL-4, IL-5, IL-13, IL-17, TNF-α, IFN-γ and IL-2,
in the serum of mice were measured. Compared with control mice,
levels of IL-4 (Fig. 5A), IL-5
(Fig. 5B), IL-13 (Fig. 5C), IL-17 (Fig. 5D) and TNF-α (Fig. 5E) in the serum of AR mice
significantly increased, while the levels of IFN-γ (Fig. 5F) and IL-2 (Fig. 5G) significantly decreased. TLR4-shRNA
administration significantly reversed each affect in AR mice.
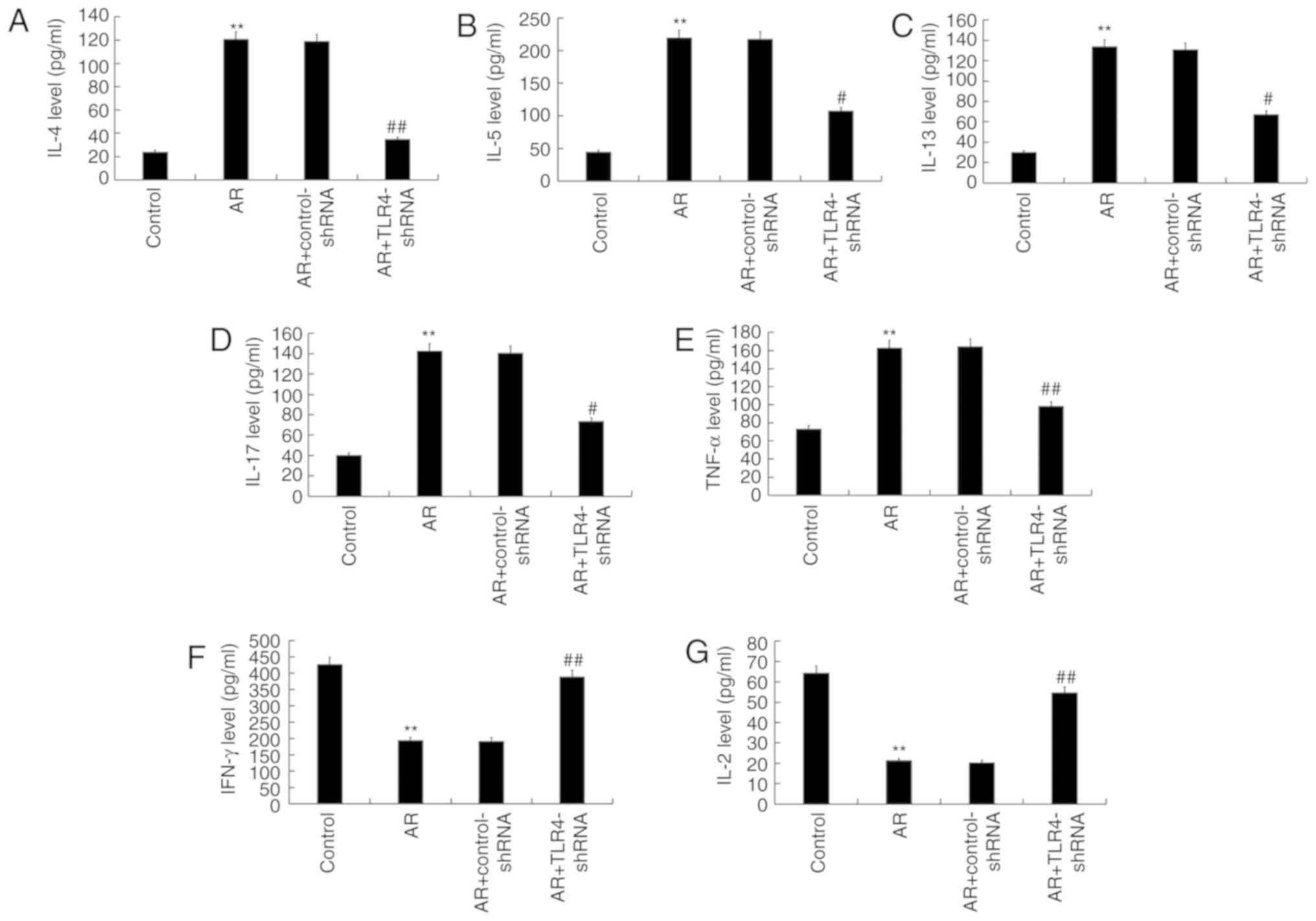 | Figure 5.TLR4 inhibition impacted inflammatory
cytokine secretion in the serum of AR mice. (A) IL-4, (B) IL-5, (C)
IL-13, (D) IL-17, (E) TNF-α (F) IFN-γ and (G) IL-2 secretion. Data
are presented as the mean ± standard deviation. **P<0.01 vs.
control group; #P<0.05 and ##P<0.01 vs.
AR group. TLR4, toll-like receptor 4; AR, allergic rhinitis; IL,
interleukin; TNF-α, tumor necrosis factor alpha; IFN- γ, interferon
γ; shRNA, short hairpin RNA. |
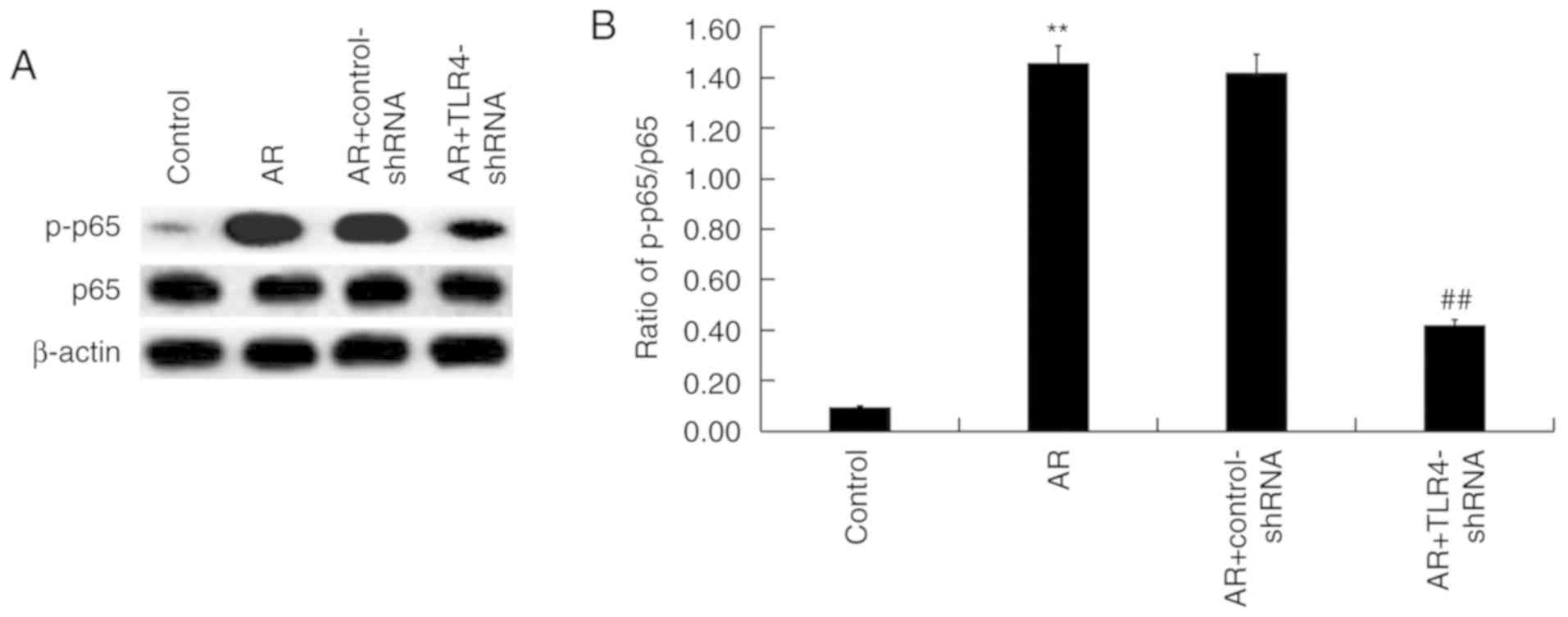
TLR4 downregulation regulates the
activation of the NFκB pathway in the nasal mucosa of AR mice
Whether the NF-κB pathway was involved in the
effects of TLR4 on AR mice was assessed. Protein levels of p-p65
and p65 were determined via western blot analysis (Fig. 6A) and the ratio of p-p65/p65 was
calculated and presented in Fig. 6B.
The increased protein level of p-p65 observed in AR mice was
significantly decreased via TLR4-shRNA administration (Fig. 6B).
Discussion
The present study demonstrated that TLR4 was
upregulated in the nasal mucosa of AR mice. Furthermore, TLR4
inhibition may significantly relieve AR symptoms, as indicated by
the decreased frequency of sneezing and nose friction, reduced
levels of OVA-specific IgE, ECP, LTC4, PGD2, fewer inflammatory
cells and decreased levels of Th2 type cytokines. The results
demonstrated that the activation of the NF-κB pathway, induced by
OVA, was repressed by TLR4-shRNA administration. Data in the
present study also revealed TLR4-shRNA administration exerted a
protective effect on AR.
AR is one of the most common types of rhinitis.
Previous epidemiological studies indicate that the average
incidence rate of AR is 10–20% and the annual incidence rate is
increasing (29,30). In the past few years, the incidence
and severity of AR in developed countries has gradually increased
and AR has become a serious public health problem (4,31).
Although some progress has been made in the treatment of AR, the
therapeutic effect is still unsatisfactory (5–8).
Therefore, finding new and effective methods and targets for the
treatment of AR is essential.
TLR4, a well-characterized receptor for
gram-negative bacterial endotoxins or LPS (32,33),
participates in inflammatory signaling responses. TLR4 recognizes
ligands including LPS, lipooligosaccharides, bacterial endotoxins
and substances secreted by viruses, fungi and mycobacteria by
pathogenassocial-edmolecular patterns (34–36). In
addition, TLR4 recognizes endogenous ligands including high
mobility group box protein 1 and β-defensin, among the
dangerous/damage-associated molecular patterns (36). TLR4 induces a pro-inflammatory immune
response by recognizing pathogens and endogenous ligands, but
abnormal expression can also promote the occurrence of various
diseases (37). Cui et al
(25) reported that TLR4 is highly
expressed in the nasal mucosa of patients with AR. Various studies
have also alluded to the critical roles of TLR4 in the development
of AR (38,39). However, the precise role of TLR4 in
AR progression remains largely unclear. Therefore, the current
study was performed.
In the current study, an OVA-induced AR mouse model
was established and treated with TLR4-shRNA. Consistent with a
previous study (25), the results
demonstrate that mRNA and protein levels of TLR4 in the nasal
mucosa of AR mice were significantly upregulated in comparison with
control mice. TLR4-shRNA administration markedly reduced the
frequency of sneezing and nose friction in AR mice, suggesting that
symptoms were alleviated. During the pathogenesis of AR, an
allergen triggers a Th2-predominant immune response that produces
antigen-specific IgE which bind to mast cells (40). Repeated exposure to the same allergen
activates IgE-binding mast cells, which release inflammatory
mediators including histamine and leukotriene C4 (41). These promote the initial irritation
and sneeze reflex of mice (42). ECP
is the most widely used marker for monitoring diseases involving
eosinophils (43). PGD2 is critical
in the interaction between different immune cells including mast
cells, Th2 cells, eosinophils and dendritic cells (44). Consistent with a previous study,
levels of OVA-specific IgE, ECP, LTC4 and PGD2 in AR mice
significantly increased (27). The
present study indicated that TLR4-shRNA administration markedly
decreased the levels of OVA-specific IgE, ECP, LTC4 and PGD2 in AR
mice. Consistent with these results, the results of the current
study also revealed that the number of upregulated leucocytes,
eosinophils, neutrophils and lymphocytes, induced by OVA induction,
was markedly reduced by the TLR4-shRNA treatment. Emerging evidence
has demonstrated that inflammatory mechanisms serve an important
role in AR development (45).
Further analysis indicates that the serum levels of IL-4, IL-5,
IL-13, IL-17 and TNF-α, which are major mediators produced by Th2
cells, were reduced following TLR4-shRNA treatment. Levels of IFN-γ
and IL-2, which are primarily produced by Th1 cells, were
upregulated by TLR4-shRNA. TLR4-shRNA may therefore exhibit a
therapeutic effect on AR mice by downregulating pro-inflammatory
cytokines produced by Th2 cells. As the major transmembrane
receptor for allergens, TLR4 transcribes NF-κB in the nucleus
through a series of complex intracellular signal transduction
pathways mediated by the adaptor protein myeloid differentiation
primary response protein-88, thereby inducing the release of a
large number of inflammatory factors including TNF-α, IL-1 and
IL-6, which in turn mediate and aggravate airway inflammation
(46). The current study also
elucidated as to whether NF-κB pathway was involved in the effects
of TLR4 on AR mice and the data indicated that OVA-induced
activation of the NF-κB pathway was inhibited by TLR4-shRNA
administration.
In summary, the expression of TLR4 was upregulated
in the OVA-induced AR mouse model. Administration of TLR4-shRNA
alleviated the allergic symptoms of AR mice by regulating the
production of pro-inflammatory mediators. The present study
revealed that TLR4-shRNA may effectively alleviate allergic
symptoms in AR mice and is therefore a promising therapeutic agent
for AR treatment. The current study is, however, preliminary and
has limitations. The NF-κB inhibitor should be used to confirm the
effect of TLR4-shRNA on the NF-κB pathway. Percentages of the
different cell types (leucocytes, eosinophils, neutrophils and
lymphocytes) were not analyzed in this study. In addition, the
relevant TLR4 ligand(s) in OVA-induced AR, whether LPS presented in
the environment or possibly in the OVA and whether OVA was
endotoxin-free were also not determined in the current study. These
issues should be further investigated to fully elucidate the role
of TLR4 in AR development.
Acknowledgements
Not applicable.
Funding
The present study was supported by Taizhou Science
and Technology Department Foundation of Zhejiang Province (grant
no. 2016A33631).
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HX designed the current study, collected and
analyzed the data, performed statistical analysis, searched the
literature, and prepared the manuscript. HS, JZ and JS collected
the data, performed the statistical analyses and interpreted the
data.
Ethics approval and consent to
participate
All animal experiments were performed in accordance
with the protocol approved by the Care and Use of Laboratory
Animals Committee. The current study was approved by the Committee
on the Use and Care of Animals of Taizhou Central Hospital (Taizhou
University Hospital, Taizhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bousquet J, Khaltaev N, Cruz AA, Denburg
J, Fokkens WJ, Togias A, Zuberbier T, Baena-Cagnani CE, Canonica
GW, van Weel C, et al: Allergic Rhinitis and its Impact on Asthma
(ARIA) 2008 update (in collaboration with the World Health
Organization, GA(2)LEN and AllerGen). Allergy. 63:8–160. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang Y, Lin L and Zheng C: Downregulation
of Orai1 expression in the airway alleviates murine allergic
rhinitis. Exp Mol Med. 44:177–190. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rosati MG and Peters AT: Relationships
among allergic rhinitis, asthma and chronic rhinosinusitis. Am J
Rhinol Allergy. 30:44–47. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Settipane RA and Schwindt C: Chapter 15:
Allergic rhinitis. Am J Rhinol Allergy. 27 (Suppl 1):S52–S55. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sur DK and Plesa ML: Treatment of allergic
rhinitis. Am Fam Physician. 92:985–992. 2015.PubMed/NCBI
|
|
6
|
Bernstein DI, Schwartz G and Bernstein JA:
Allergic rhinitis: Mechanisms and treatment. Immunol Allergy Clin
North Am. 36:261–278. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Andersson M, Greiff L, Ojeda P and Wollmer
P: Barrier-enforcing measures as treatment principle in allergic
rhinitis: a systematic review. Curr Med Res Opin. 30:1131–1137.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Steelant B, Farré R, Wawrzyniak P, Belmans
J, Dekimpe E, Vanheel H, Van Gerven L, Kortekaas Krohn I, Bullens
DMA, Ceuppens JL, et al: Impaired barrier function inpatients with
house dust mite-induced allergic rhinitis is accompanied
bydecreased occludin and zonula occludens-1 expression. J. Allergy
Clin.Immunol. 137:1043–1053. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang DY: Risk factors of allergic
rhinitis: Genetic or environmental? Ther Clin Risk Manag.
1:115–123. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ciprandi G, Marseglia GL, Castagnoli R,
Valsecchi C, Tagliacarne C, Caimmi S and Licari A: From IgE to
clinical trials of allergic rhinitis. Expert Rev Clin Immunol.
11:1321–1333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hashimoto C Hudson KL and Anderson KV: The
toll gene of Drosophila, required for dorsal-ventral embryonic
polarity, appears to encode a transmembrane protein. Cell.
52:269–279. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lemaitre B Nicolas E, Michaut L, Reichhart
JM and Hoffmann JA: The dorsoventral regulatory gene cassette
spätzle/Toll/cactus controls the potent antifungal response in
Drosophila adults. Cell. 86:973–983. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Medzhitov R, Preston-Hurlburt P and
Janeway CA Jr: A human homologue of the Drosophila Toll protein
signals activation of adaptive immunity. Nature. 388:394–397. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Iwasaki A and Medzhitov R: Regulation of
adaptive immunity by the immune system. Science. 327:291–295. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Akira S and Takeda K: Toll-like receptor
signalling. Nat Rev Immunol. 4:499–511. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Takeda K, Kaisho T and Akira S: Toll-like
receptors. Annu Rev Immuno1. 21:335–376. 2003. View Article : Google Scholar
|
|
17
|
Molteni M, Gemma S and Rossetti C: The
role of toll-like receptor 4 in infectious and noninfectious
inflammation. Mediators Inflamm. 2016:69789362016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Beutler B: TLR4 as the mammalian endotoxin
sensor. Curr Top Microbiol Immunol. 270:109–120. 2002.PubMed/NCBI
|
|
19
|
Beutler B, Du X and Poltorak A:
Identification of toll-like receptor 4 (TLR4) as the sole conduit
for LPS signal transduction: Genetic and evolutionary studies. J
Endotoxin Res. 7:277–280. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Garibotto G, Carta A, Picciotto D, Viazzi
F and Verzola D: Toll-like receptor-4 signaling mediates
inflammation and tissue injury in diabetic nephropathy. J Nephrol.
30:719–727. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Maglione PJ, Simchoni N and
Cunningham-Rundles C: Toll-like receptor signaling in primary
immune deficiencies. Ann N Y Acad Sci. 1356:1–21. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dong L, Li H, Wang S and Li Y: Different
doses of lipopolysaccharides regulate the lung inflammation of
asthmatic mice via TLR4 pathway in alveolar macrophages. J Asthma.
46:229–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peters M, Dudziak K, Stiehm M and Bufe A:
T-cell polarization depends on concentration of the danger signal
used to activate dendritic cells. Immunol Cell Biol. 88:537–544.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takabayashi K, Libet L, Chisholm D,
Zubeldia J and Horner AA: Intranasal immunothcrapy is more
effective than intradcrmal immunotherapy for the induction of
air-way allergen tolerance in Th2-sensitized mice. J Immunol.
170:3898–3905. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cui XY, Chen X, Yu CJ, Yang J, Lin ZP, Yin
M and Cheng L: Increased expression of toll-like receptors 2 and 4
and related cytokines in persistent allergic rhinitis. Otolaryngol
Head Neck Surg. 152:233–238. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xiao L, Jiang L, Hu Q and Li Y:
MicroRNA-133b ameliorates allergic inflammation and symptom in
murine model of allergic rhinitis by targeting Nlrp3. Cell Physiol
Biochem. 42:901–912. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yuan Y, Liu Q, Zhao J, Tang H and Sun J:
SIRT1 attenuates murine allergic rhinitis by downregulated HMGB
1/TLR4 pathway. Scand J Immunol. 87:e126672018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Casale TB and Dykewicz MS: Clinical
implications of the allergic rhinitis-asthma link. Am J Med Sci.
327:127–138. 2014. View Article : Google Scholar
|
|
30
|
Brożek JL, Bousquet J, Agache I, Agarwal
A, Bachert C, Bosnic-Anticevich S, Brignardello-Petersen R,
Canonica GW, Casale T, Chavannes NH, et al: Allergic Rhinitis and
its Impact on Asthma (ARIA) guidelines:evision. J Allergy Clin
Immunol. 140:950–958. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Izquierdo-Domínguez A, Valero AL and
Mullol J: Comparative analysis of allergic rhinitis in children and
adults. Curr Allergy Asthma Rep. 13:142–151. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shimazu R, Akashi S, Ogata H, Nagai Y,
Fukudome K, Miyake K and Kimoto M: MD-2, a molecule that confers
lipopolysaccharide responsiveness on Toll-like receptor 4. J Exp
Med. 189:1777–1782. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rifkin IR, Leadbetter EA, Busconi L,
Viglianti G and Marshak-Rothstein A: Toll-like receptors,
endogenous ligands and systemic autoimmune disease. Immunol Rev.
204:27–42. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kang JY and Lee JO: Structural biology of
the Toll-like receptor family. Annu Rev Biochem. 80:917–941. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Peri F and Calabrese V: Toll-like receptor
4 (TLR4) modulation by synthetic and natural compounds an update. J
Med Chem. 57:3612–3622. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhong JJ, Wan YY, Diao YW, et al: The
research development of Toll-like receptors targeted drugs. Chin
Bull Life Dci. 27:439–444. 2015.(In Chinese).
|
|
37
|
Sasai M and Yamamoto M: Pathogen
recognition receptors: Ligands and signaling pathways by Toll-like
receptors. Int Rev Immunol. 32:116–133. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fuertes E, Brauer M, MacIntyre E, Bauer M,
Bellander T, von Berg A, Berdel D, Brunekreef B, Chan-Yeung M,
Gehring U, et al: Childhood allergic rhinitis, traffic-related air
pollution and variability in the GSTP1, TNF, TLR2 and TLR4 genes:
Results from the TAG study. J Allergy Clin Immunol. 132:342–352.e2.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Radman M, Golshiri A, Shamsizadeh A,
Zainodini N, Bagheri V, Arababadi MK and Kennedy D: Toll-like
receptor 4 plays significant roles during allergic rhinitis.
Allergol Immunopathol (Madr). 43:416–420. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhong H, Fan XL, Yu QN, Qin ZL, Chen D, Xu
R, Chen DH, Lin ZB, Wen W and Fu QL: Increased innate type 2 immune
response in house dust mite-allergic patients with
allergicrhinitis. Clin Immunol. 183:293–299. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Madore AM and Laprise C: Immunological and
genetic aspects of asthma and allergy. J Asthma Allergy. 3:107–121.
2010.PubMed/NCBI
|
|
42
|
Galli SJ and Tsai M: IgE and mast cells in
allergic disease. Nat Med. 18:693–704. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Metcalfe DD, Pawankar R, Ackerman SJ, Akin
C, Clayton F, Falcone FH, Gleich GJ, Irani AM, Johansson MW, Klion
AD, et al: Biomarkers of the involvement of mast cells, basophils
and eosinophils in asthma and allergic diseases. World Allergy
Organ J. 9:72016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fujitani Y, Kanaoka Y, Aritake K, Uodome
N, Okazaki-Hatake K and Urade Y: Pronounced eosinophilic lung
inflammation and Th2 cytokine release in human lipocalin-type
prostaglandin D synthase transgenic mice. J Immunol. 168:443–449.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bae JS, Kim JH, Kim EH and Mo JH: The role
of IL-17 in a lipopolysaccharide-induced rhinitis model. Allergy
Asthma Immunol Res. 9:169–176. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Eswarappa SM, Basu N, Joy O and
Chakravortty D: Folimycin (concanamycin A) inhibits LPS-induced
nitric oxide production and reduces surface localization of TLR4 in
murine macrophages. Innate Immun. 14:13–24. 2008. View Article : Google Scholar : PubMed/NCBI
|