Introduction
Myocardial infarction (MI), also referred to as a
heart attack, is characterized by acute heart muscle damage caused
by a limited blood supply, which can result in cardiogenic shock
and cardiac arrest (1). Globally,
>15 million MI cases are reported each year and this incurs an
enormous burden on affected patients (2). A number of treatments have been
effectively used to treat patients experiencing MI, including
aspirin, nitroglycerin, opioids, oxygen supplementation and
angioplasty, but long-term medication and treatments are still
required (3). Previous
epidemiological and pathological studies have demonstrated that the
majority of MI cases are induced by coronary artery blockages,
which are closely associated with a number of conditions, including
high blood pressure, diabetes, high blood cholesterol and smoking
(4). Cardiomyocyte apoptosis
mediates the development of rapid myocardial damage following MI
(5), while autophagy exhibits
protection against cardiac damage following MI (6,7).
However, the cellular pathogenesis of MI is still poorly
understood, and novel cardiomyocyte apoptosis-inhibiting methods
require further elucidation.
Mesenchymal stem cells (MSCs) are multipotent cells
that can differentiate into a variety of cell types, including
myocytes, osteoblasts, chondrocytes and adipocytes (8). MSCs that are derived from bone marrow
have been demonstrated to mediate pleiotropic processes, including
immunomodulation and tissue damage healing (9). MSCs have also been identified as
potential targets for cellular treatments that can be used in a
variety of disorders, including autoimmune diseases, graft versus
host disease and sepsis (9). MSCs
have also been found to be effective in MI treatment. For example,
previous studies using experimental animal models demonstrated that
the injection of MSCs immediately after MI onset improved
myocardial compliance, promoted cardiac nerve sprouting and
improved ventricular function (10–12). The
effects of autologous MSCs on MI have also been validated in
patients exhibiting MI (13),
indicating the potential of these cells as a promising therapy for
use in MI treatment. Based on the results from previous
investigations, the beneficial effects of MSCs on MI have been
suggested to be mediated by immuno-inflammatory regulation;
functional gene expression modulation, including in the vascular
endothelial growth factor (VEGF) gene; and major cellular
signaling, including in neuregulin1-erythroblastic leukemia viral
oncogene homolog 4 signaling (14–16). It
has also been revealed that the VEGF, from bone marrow-derived
MSCs, significantly inhibits cardiomyocyte apoptosis during MI via
the suppression of microRNA (miRNA)-23a and miRNA-92a expression
(17). However, the molecular
mechanisms of MSC MI treatment are still to be fully
elucidated.
Exosomes are characterized as microRNA-containing
vesicles that are derived from a variety of cell types, including
stem cells, and are widespread in blood, urine and are often
detected in cell culture medium (18,19).
Increasing evidence has demonstrated that exosomes exhibit key
regulatory functions in a variety of biological processes,
including in coagulation and intercellular signaling, and this
indicates the potential for their application in clinical
diagnosis, treatment and prognosis prediction in numerous human
disorders (18–21). It has also been revealed that
exosomes are closely associated with cardiac repair and
regeneration, due to their anti-apoptotic, anti-fibrotic and
pro-angiogenic roles (19). For
example, exosomes that are secreted by mouse bone marrow-derived
dendritic cells have been indicated to greatly enhance the
expression of chemokines and inflammatory cytokines, and improve
the cardiac function of mice following MI, via the activation of
CD4+ T lymphocytes (22).
One recent report demonstrated that exosomes that are derived from
embryonic stem cells (ESCs) promoted the endogenous repair system
and strengthened cardiac function following MI (23). Furthermore, exosomes that are
secreted by MSCs in the bone marrow have also been previously
demonstrated to reduce myocardial ischemia/reperfusion (I/R) injury
(24). However, further validation
of these observations is required, and the underlying mechanisms
remain undetermined.
The aim of the current study was to investigate the
effects of exosomes derived from bone marrow-derived MSCs on MI,
using cellular and animal models, and to determine the underlying
mechanisms of this interaction, which may provide novel insights
into the therapeutic value of exosomes for the treatment of
cardiovascular disease.
Material and methods
Rat MSC isolation and culture
MSCs were isolated from the bone marrow tissue of 12
3-week-old male Sprague Dawley (SD) rats weighing 80–100 g, which
were supplied by the Animal Center of Southern Medical University
as previously described (25),
housed in cages at 22–24°C in a 12-h light/dark cycle and fed with
standard food pellets and tap water ad libitum. All surgical
procedures in the current study were performed following The
Guidelines for Laboratory Animal Care and Use by Sun Yat-sen
University. The present study was approved by the Animal Ethics
Committee of The Third Affiliated Hospital of Sun Yat-sen
University. Male rats aged three weeks upon purchase were
sacrificed for the collection of femora bone marrow tissues, which
were flushed with minimal essential medium (Thermo Fisher
Scientific, Inc.) supplemented with 15% FBS (Thermo Fisher
Scientific, Inc.), penicillin and streptomycin. After filtration
and centrifugation, the pellets containing bone marrow-derived MSCs
were cultured use in for subsequent assays. MSCs were cultured in
dishes for 7 days in a humidified chamber at 37°C, and supplied
with 5% CO2. Culture medium was replaced three times per
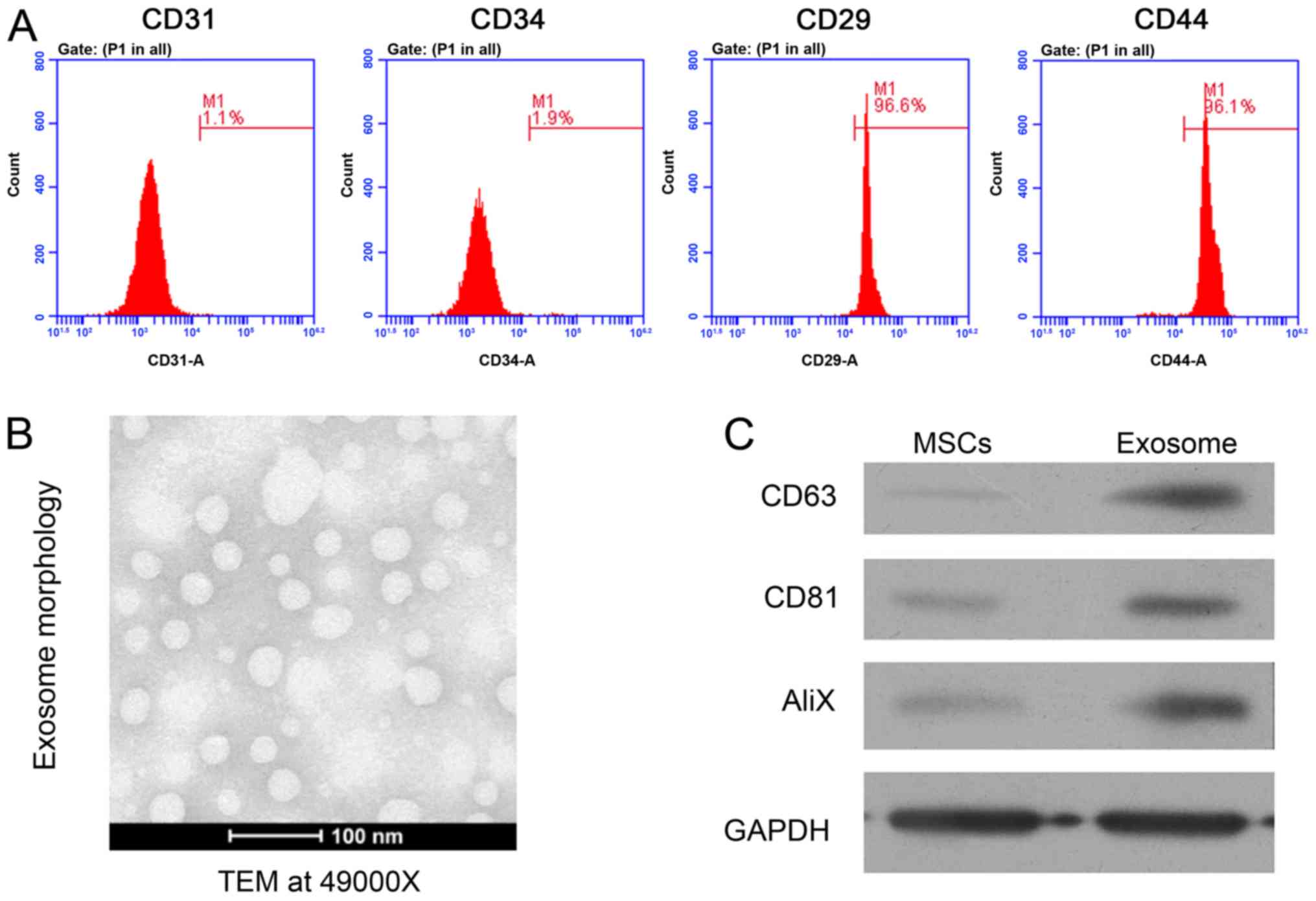
week. The identity of isolated MSCs was validated using flow
cytometry as previously described (26) and defined by the positive expression
of cell surface antigens CD29 and CD44, without CD31 and CD34
expression. Primary antibodies against CD29 (1:100; cat. no.
11-0291-82), CD44 (1:100; cat. no. MA5-16908), CD31 (1:100; cat.
no. MA5-16953) and CD34 (1:100; cat. no. PA5-85917) provided by
Thermo Fishier Scientific. Inc. were used for flow cytometry.
Exosome isolation and electron
microscopy
The isolation of exosomes from rat MSCs was
performed using a previously described method (23). Isolated MSCs were cultured for >40
h and exosomes in culture medium were collected using an
ultracentrifugation at 100,000 × g for 70 min at 4°C. Purified
exosomes from MSCs were then resuspended and preserved in saline
for subsequent experiments. For electron microscopy, exosomes
suspensions were first mixed with 4% paraformaldehyde at RT for 15
min, applied onto formvar-coated carbon grids which was pre-coated
with 0.01% Poly Lysine at RT for 1 min, stained with 1%
phosphotungstic acid at RT for 2 min, the morphological features of
isolated exosomes were finally observed using an FEI Tecnai Spirit
G2 transmission electron microscope (Thermo Fisher Scientific,
Inc.) with 120-kV voltage and ×49,000 magnification.
Rat myocardial I/R model and exosome
administration
The establishment of the myocardial I/R model in SD
rats was performed using a method previously described (23). A total of 18 male rats aged 7 weeks
were randomly divided into the surgery-only, sham and surgery +
exosomes groups. Rats in the surgery-only and surgery + exosomes
groups were subjected to surgery to ligate the left anterior
descending coronary artery. Sham surgery was also performed on 6
rats in the sham group, which were used as negative controls.
Subsequently, the exosomes (10 µg) isolated from rat MSCs were
intramyocardially injected into the left ventricular wall of 6 rats
in the surgery + exosomes group that underwent left anterior
descending coronary artery ligation surgery.
Histological analysis of rat
myocardial tissues
The hearts of experimental rats were collected,
after rats were sacrificed using pentobarbital sodium (135 mg/kg),
following the establishment of the myocardial I/R model and the
specified treatments. The pathogenic features of rat myocardial
tissues were evaluated using histological analysis. The heart
tissues were fixed in 4% paraformaldehyde at RT for 24 h,
dehydrated with 70% ethonal for 2 min, 90% ethanol for 2 min and
95% ethanol for 2 min, embedded in paraffin and sliced into 4 µm
thick serial sections. For hematoxylin-eosin (H&E) staining,
tissue slides were stained with hematoxylin at RT for 5 min and
stained with eosin at RT for 2 min, finally observed with
microscope (TE200, Nikon Corporation) with a ×400 magnification for
at least 6 fields. TUNEL staining was performed by incubation with
Protease K solution at 37°C for 30 min, incubation with TdT and
dUTP mixture (2:29) at 37°C for 2 h, blocking of endogenous
peroxidase with 3% hydrogen peroxide solution in methanol in the
dark for 15 min and incubation with converter-POD at 37°C for 30
min. mounting with Antifade Mounting Medium (cat. no. P0126;
Beyotime Institute of Biotechnology) and observed using an inverted
fluorescence microscope (TE2000-E; Nikon Corporation) with a ×400
magnification for at least 6 fields. 2,3,5-triphenyltetrazolium
chloride (TTC) staining was performed by immersion in 1% TTC at RT
for 15 min. For Masson staining, the tissue slides were stained
with Ponceau S acid fuchsin at RT for 5–10 min, followed by
incubation with a molybdophosphoric acid solution at RT for 3 min
and staining with aniline blue at RT for 5 min, and finally
observed with a camera (EOS 80D; Canon, Inc.).
Cellular hypoxia-reoxygenation (H/R)
model and treatment
Rat cardiac H9c2 cells were used as the cellular H/R
model in the present study. H9c2 cells were obtained from the
American Type Culture Collection and were cultured in DMEM (Thermo
Fishier Scientific, Inc.) containing high glucose supplemented with
10% FBS (Thermo Fishier Scientific, Inc.), penicillin and
streptomycin at 37°C with 5% CO2. As previously
described (27), the cellular H/R
model was established by culturing H9c2 cells in DMEM containing
low glucose with no supplementation of FBS and with 5%
CO2 in a hypoxic atmosphere for 4 h, followed by
culturing under a normal oxygen supply for 24 h. To analyze the
exosome effect, the H9c2 cells were then treated with 2 µg exosomes
at 37°C for 48 h. The H9c2 cells were pretreated with 10 mM
3-methyladenine (3-MA) at 37°C for 30 min to inhibit autophagy
progression.
Reverse transcription-quantitative
(RT-q)PCR assay
Total RNA samples were extracted from rat tissues
and cultured cells using TRIzol® solution (cat. no.
12183555; Thermo Fishier Scientific, Inc.) following the
manufacturer's protocol. The cDNA synthesis was then performed
using the Bestar qPCR RT kit (cat. no. 2220; DBI Bioscience) as
described by the manufacturer's protocol, with 3 µg RNA sample used
as a template and a setting of 37°C for 15 min and 98°C for 5 min.
The relative expression of genes were finally determined by RT-q
PCR using the Bestar™ qPCR MasterMix (cat. no. 2043; DBI
Bioscience) according to the manufacturer's protocol, on an Agilent
Stratagene PCR system (Mx3000P; Agilent Technologies, Inc.). The
following conditions were used: Pre-denaturation at 95°C for 2 min
followed by 40 cycles of denaturation at 94°C for 20 sec, annealing
at 58°C for 20 sec and extension at 72°C for 20 sec. The relative
expression of genes was determined using the 2−ΔΔCq
method (28). Primers used for
quantification were as follows: Apoptotic protease activating
factor-1 (Apaf1) forward, 5′-TCAAAAGTCAGGCCACTCAA-3′ and reverse,
5′-TGAAGTGTTTCCACCGTCTG-3′; elevated autophagy-related protein 13
(ATG13) forward, 5′-TCCACCAGGCAATTTGAGAG-3′ and reverse,
5′-TCAAAAGTCAGGCCACTCAA-3′; and GAPDH forward,
5′-CCTCGTCTCATAGACAAGATGGT-3′ and reverse,
5′-GGGTAGAGTCATACTGGAACATG-3′.
Western blot analysis
Total proteins from exosomes, rat tissues and
cultured cells were separately extracted using RIPA lysis and
extraction buffer (Thermo Fisher Scientific, Inc.) for western blot
analysis. The protein concentration was determined using the Thermo
Bicinchoninic Acid Protein Assay kit (cat. no. 23227; Thermo Fisher
Scientific, Inc.). Total proteins (20 µg) were loaded onto and
separated using 10% SDS-PAGE gels and blotted onto a PVDF membrane.
After being blocked with 5% lipid-free milk solution for 2 h at
room temperature, the PVDF containing targeted proteins were probed
with primary (RT; 2 h) and secondary antibodies (RT; 2 h)
successively. Blots were developed using enhanced chemiluminescence
detection reagents (cat. no. WB KLS0500; EMD Millipore). Band
intensity was analyzed using the Image-Pro Plus 6.0 software (Media
Cybernetics, Inc.). GAPDH was applied as the internal standard for
relative protein abundance determination. The following antibodies
were used in the current study: Anti-CD63 (1:1,000; cat. no.
ab108950; Abcam), anti-CD81 (1:1,000; cat. no. ab232390; Abcam),
anti-ALG-2-interacting protein X (AliX; 1:1,000; cat. no. ab117600;
Abcam), anti-ATG13 (1:1,000; cat. no. ab201467; Abcam), anti-Apaf1
(1:2,000; cat. no. ab2001; Abcam), anti-Bcl2 (1:1,000; cat. no.
ab196495; Abcam), anti-Beclin-1 (1:1,000; cat. no. ab207612;
Abcam), anti-Bax (1:1,000; cat. no. ab32503; Abcam), anti-GAPDH
(1:10,000; cat. no. ab8245; Abcam), horseradish peroxidase (HRP)
goat anti-Rabbit IgG (1:20,000; cat. no. BA1054; Wuhan Boster
Biological Technology) and HRP goat anti-mouse IgG (1:20,000; cat.
no. BA1051; Wuhan Boster Biological Technology).
Cell proliferation, apoptosis and
migration tests
H9c2 cell proliferation was evaluated using the
5-ethynyl-2′-deoxyuridine (EdU) method. Cultured H9c2 cells
(1×105/ml) were stained at 37°C with 1 µM EdU solution
(Thermo Fisher Scientific, Inc.) for 30 min, incubated with 1X
Hoechst 33342 solution (Beyotime Institute of Biotechnology) for 10
min at room temperature, and observed using a fluorescence
microscope (magnification, ×200). The apoptosis of H9c2 cells was
analyzed by staining cells with 5 µg/ml Hoechst 33258 for 20 min at
room temperature, and observed with a confocal microscope
(magnification, ×120). H9c2 cell apoptosis analysis was performed
using flow cytometry using the Annexin V-FITC Apoptosis
Staining/Detection kit (cat. no. ab14085; Abcam), and cells were
incubated with 5 µl Annexin V-FITC and 5 µl propidium iodide at
room temperature for 5 min in the dark, and analyzed using WinMDI
software (Version 2.8, J.; Trotter, Scripps Research Institute).
H9c2 cell migration was evaluated using the Transwell system-based
assay. H9c2 cells (5×105/ml) resuspended in serum-free
DMEM were seeded and cultured in the upper chamber for 24 h. FBS
(10%) in DMEM was used as a chemoattractant. The migrated cells
were finally observed using a light microscope (TE200, Nikon
Corporation; magnification, ×200) by staining with 0.1% crystal
violet at RT for 15 min.
Immunofluorescence
The Apaf1 and autophagy-related protein 13 (ATG13)
protein expression in cultured H9c2 cells was performed using an
immunofluorescence assay. H9c2 cells seeded on cover slides were
fixed using 2% paraformaldehyde at RT for 15 min, blocked with 5%
BSA at RT for 70 min and incubated with the Apaf1 or ATG13
antibodies for 30 min at RT and washed three times with PBS for 10
min. After incubation with secondary antibodies at RT for 1 h, the
slides were mounted with DAPI-containing SlowFade Antifade Reagents
(cat. no. S36939; Thermo Fisher Scientific, Inc.), and finally
observed using a confocal microscope (magnification, ×200).
Statistical analysis
Statistical analyses were performed using the SPSS
18.0 software (IBM Corp.), with data presented as the mean ±
standard deviation of three replicates. Statistical analyses were
performed using an ANOVA statistical test and the Tukey post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Isolation of exosomes secreted by rat
MSCs
To investigate the potential roles of rat
MSC-derived exosomes in MI, primary MSCs were isolated from the
bone marrow tissues of SD rats. Flow cytometry was performed to
confirm the identity of the collected MSCs. The positive expression
of cell surface genes CD29 and CD44, combined with lower expression
of CD31 and CD34 were used as biomarkers to confirm the isolation
of MSCs (Fig. 1A). Subsequently,
exosomes were extracted from the culture medium of isolated rat
MSCs. Using a transmission electron microscope, the regular shape
of isolated MSC-derived exosomes was observed (Fig. 1B). The expression of three exosome
marker genes, CD63, CD81 and AliX, were further analyzed using
western blot analysis, which demonstrated the significantly higher
expression of these proteins in exosomes compared with MSCs
(Fig. 1C). Collectively, these
results confirmed the successful isolation of MSCs from rat bone
marrow tissues and enriched MSC-derived exosomes, which were used
for the subsequent assays.
Inhibition of myocardial damage by
MSC-derived exosomes
To analyze the protective effects of exosomes
isolated from rat MSCs on MI, the myocardial I/R model rats were
established using a surgical procedure and treated with exosomes.
TTC staining revealed that the I/R model rats showed considerable
myocardial damage when compared with the sham surgery group.
However, the application of exosomes from the rat MSCs inhibited
the progression of myocardial damage that was induced by MI,
although this result was not exhibited across all samples (Fig. 2A). The histological features of rat
myocardial tissues were further analyzed using H&E and Masson
staining, which revealed an irregular alteration of cell shape and
lining in the rat myocardium after the induction of MI. The
injection of MSC-derived exosomes mitigated the abnormal cellular
alterations (Fig. 2B). Furthermore,
TUNEL staining was performed to evaluate cell apoptosis in rat
myocardial tissues. A marked increase in apoptotic cells was
indicated in the I/R rats. Cardiomyocyte apoptosis was suppressed
by exosomes treatment in rats that underwent left anterior
descending coronary artery ligation (Fig. 2B).
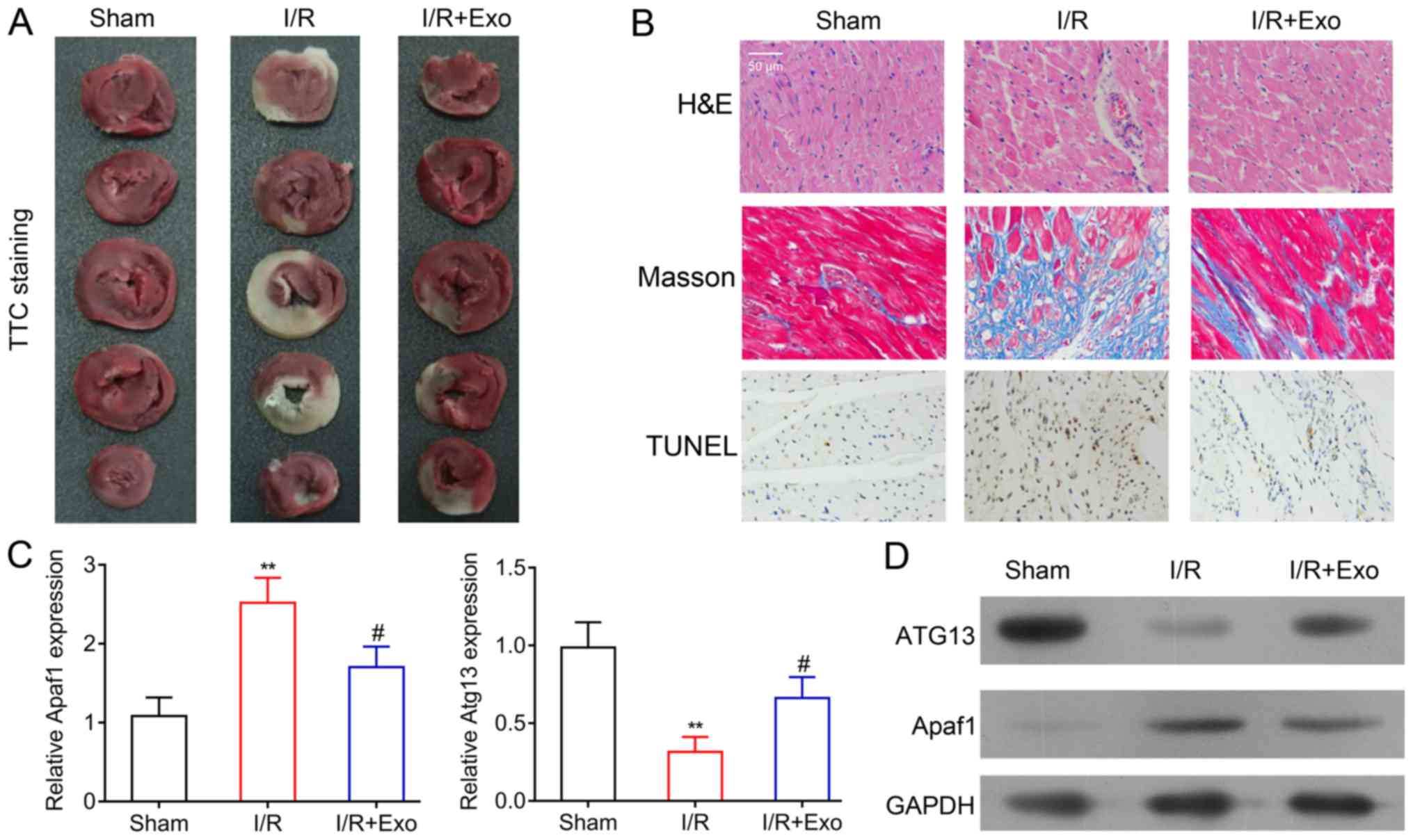 | Figure 2.Inhibition of rat myocardial damages
by MSC-derived exosomes. (A) TTC staining and (B) H&E, Masson
and TUNEL staining of the Sham, I/R and I/R+Exo groups
(magnification, ×400). (C) Reverse transcription-quantitative PCR
and (D) western blot analysis of Apaf1 and ATG13 mRNA and protein
levels in the Sham, I/R and I/R+Exo groups. GAPDH was applied as
the internal standard. **P<0.01 vs. Sham group; #P<0.05 vs.
I/R group. MSC, mesenchymal stem cell; TTC,
2,3,5-triphenyltetrazolium chloride; H&E, hematoxylin and
eosin; I/R, ischemia reperfusion; Exo, exosomes; Apaf1, apoptotic
protease activating factor-1; ATG13, autophagy-related protein
13. |
Furthermore, Apaf1 mRNA expression was increased in
the myocardial tissues of I/R rats, compared with the sham group,
while the expression of ATG13 expression was significantly
decreased in the myocardial tissues of I/R rats (Fig. 2C). Consistent with the histological
alteration of rat heart tissues, the application of exosomes from
rat MSCs recovered Apaf1 and ATG13 mRNA expression in rat
myocardial tissues (Fig. 2C).
Additionally, the changes in Apaf1 and ATG13 expression in the
sham, I/R and exosome treatment groups of rat hearts were further
validated using western blot analysis (Fig. 2D). Overall, the histological and
molecular findings suggested that exosomes from rat MSCs, could
effectively suppress myocardial tissue damage induced by MI.
Exosomes mitigate H/R injuries in H9c2
cells
To further investigate the roles of mesenchymal stem
cell-secreted exosomes in MI therapy, a cellar H/R model was
applied using rat cardiac H9c2 cells. EdU and Hoechst 33342
staining revealed that MSC-derived exosomes induced no significant
changes in the rate of H9c2 cell proliferation compared with normal
H9c2 cells (Fig. 3A). However, H9c2
cell proliferation was significantly suppressed by H/R but
significantly recovered by exosome treatment (Fig. 3A). The results of Hoechst 33258
staining also demonstrated that H/R induced apoptosis in H9c2
cells, while the mesenchymal stem cell-derived exosome treatment
significantly reduced H9c2 apoptosis (Fig. 3B). The inhibitory effects of
MSC-derived exosomes on H9c2 cell apoptosis during H/R was further
validated using flow cytometry (Fig.
3C). Additionally, the migration capability of H9c2 cells was
detected using a Transwell system, and H/R significantly decreased
H9c2 migration capacity (Fig. 3D).
However, exosome treatment increased H9c2 cell proliferation after
H/R (Fig. 3D). Collectively, these
cellular biology assays demonstrated that exosomes from rat MSCs
could effectively ameliorate the H/R injury induced in rat H9c2
cells. These results further validated the use of exosomes isolated
from the rat MSCs in protection against MI.
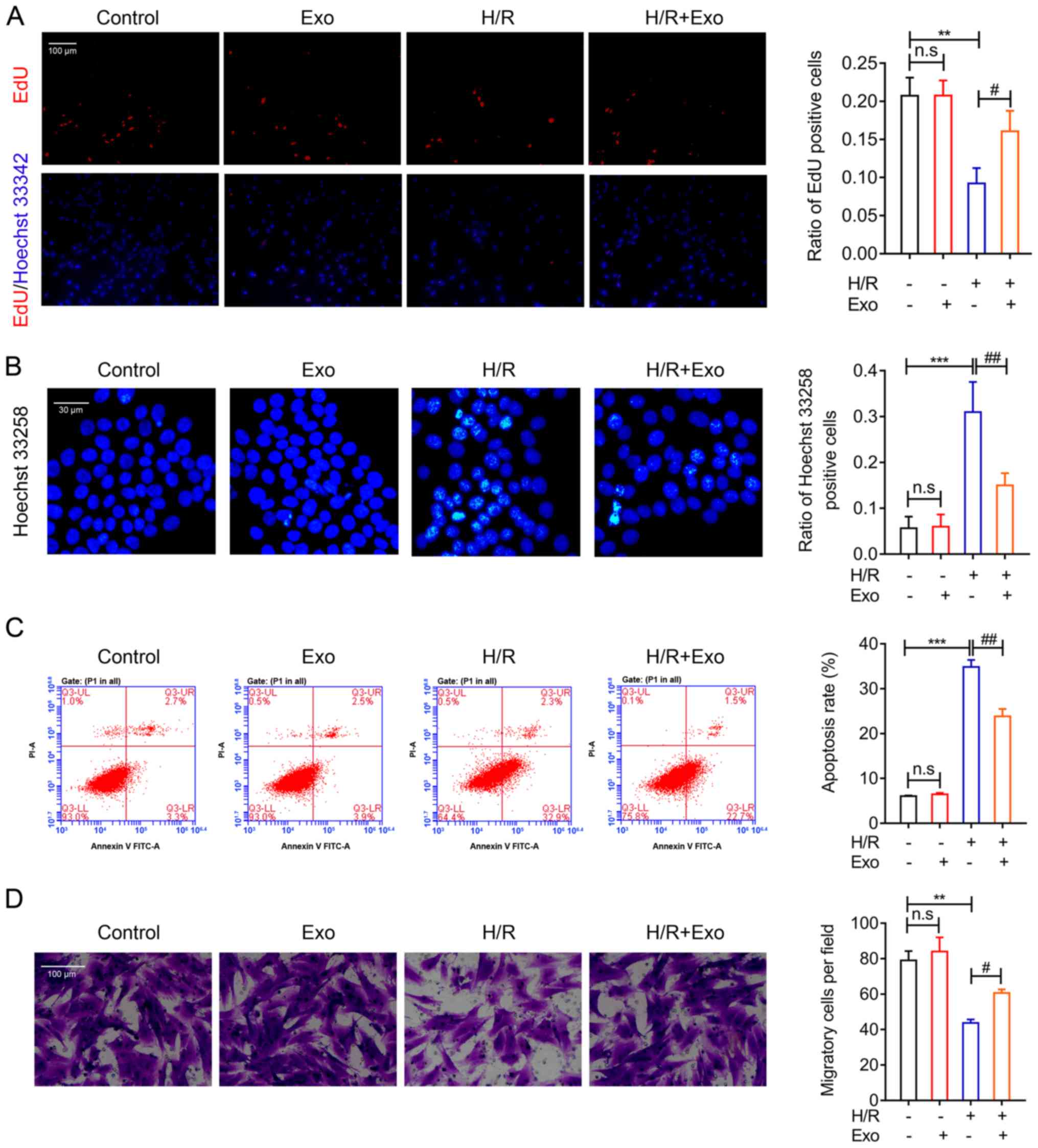 | Figure 3.Inhibition of H9c2 cell H/R injuries
by MSC exosomes. (A) EdU and Hoechst 33342 staining of the
proliferation capability of rat H9c2 cells after H/R and exosome
treatment (magnification, ×100). (B) Hoechst 33258 and (C) flow
cytometry analysis of apoptosis in rat H9c2 cells after H/R and
exosome treatment. (D) Migration capability of H9c2 cells after H/R
and MSC exosome treatment (magnification, ×200). **P<0.01,
***P<0.001; #P<0.05, ##P<0.01. EdU,
5-ethynyl-2′-deoxyuridine; MSC, mesenchymal stem cell; H/R,
hypoxia-reoxygenation; Exo, exosomes; ns, not significant; PI,
propidium iodide. |
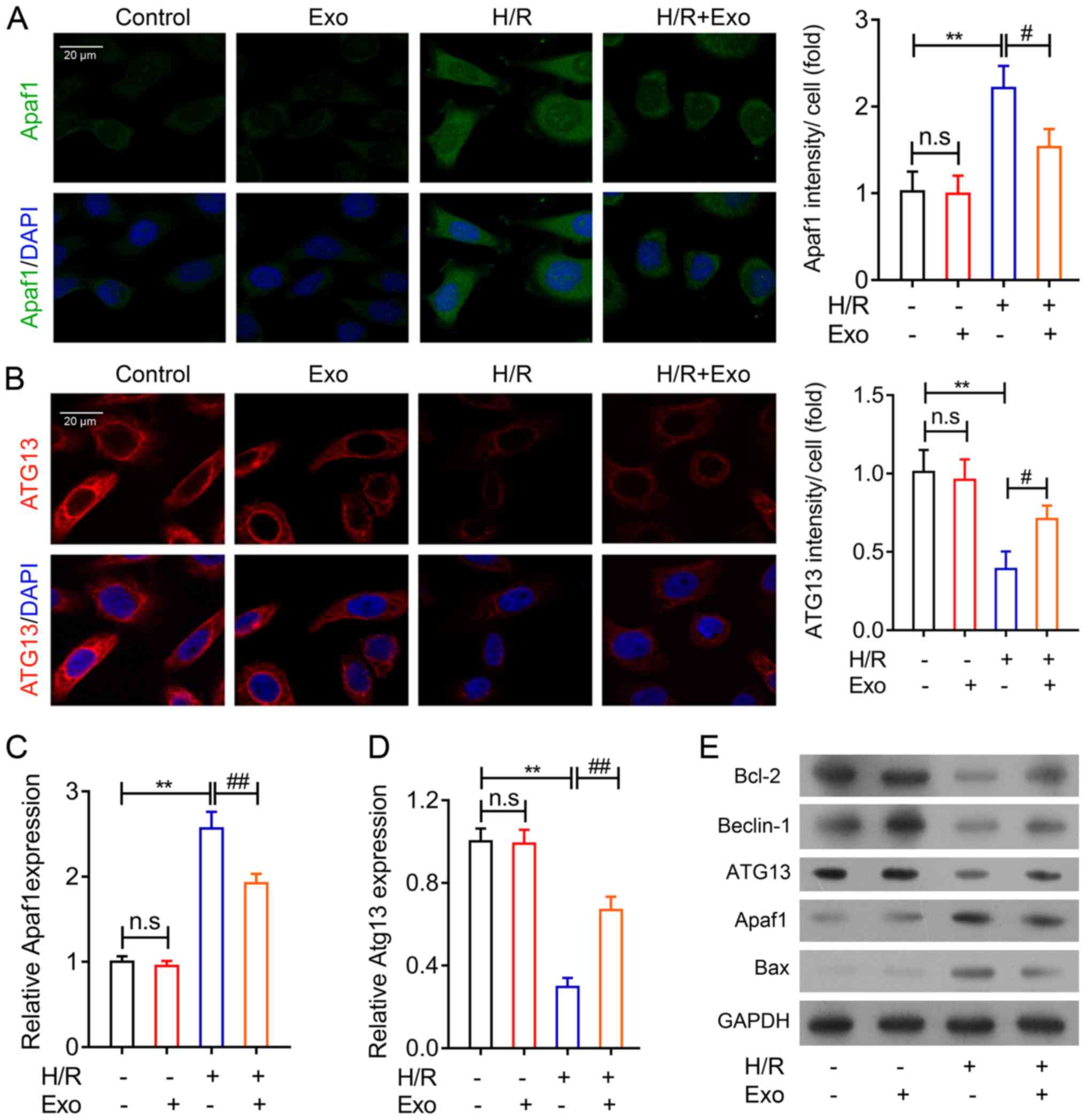
Apoptosis-related and
autophagy-related gene expression in H9c2 cells regulated by
exosomes
To further confirm the inhibitory effects of rat
mesenchymal stem cell-derived exosomes on MI, two major functional
genes associated with cell apoptosis and autophagy were analyzed in
H9c2 cells following H/R. An immunofluorescence assay was used, and
the results demonstrated that Apaf1 expression in H9c2 cells was
increased during H/R but was significantly suppressed by exosome
treatment (Fig. 4A). The results
also indicated that ATG13 expression was repressed by H/R in H9c2
cells, but MSC exosomes enhanced ATG13 expression in H9c2 cells
after H/R (Fig. 4B). The alteration
of Apaf1 and ATG13 gene and protein expression in H9c2 cells
accompanying H/R injury was confirmed using RT-qPCR (Fig. 4C and D) and western blotting
(Fig. 4E). Additionally, Bcl-2 and
Bax expression were not influenced by exosome treatment in H9c2
cells under normal conditions. However, Bcl-2 gene expression was
markedly increased following MSC-derived exosome treatment
following H/R, while Bax gene expression was suppressed by MSC
exosome treatment in H9c2 cells after H/R treatment (Fig. 4E). Beclin-1 gene expression, an
additional autophagy-associated gene, was also suppressed by H/R in
H9c2, which was mitigated by exosome treatment (Fig. 4E). These results further indicated
that exosomes isolated from rat MSCs could regulate apoptosis and
autophagy in rat myocardial cells.
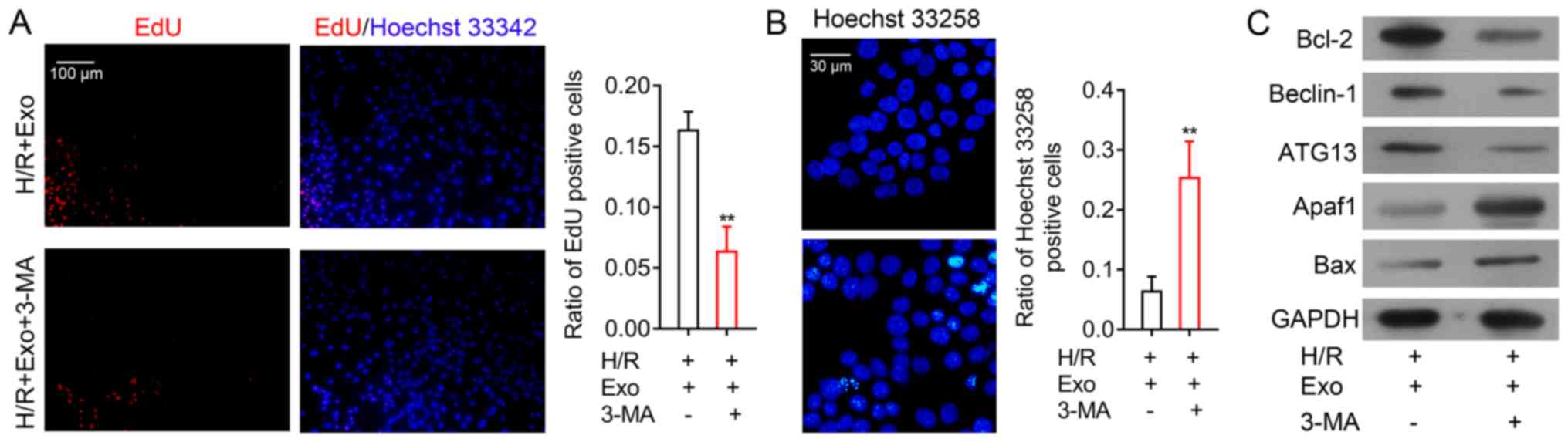
Role of autophagy in exosome-induced
H/R injury inhibition
To analyze the role of autophagy in the inhibition
of H9c2 cell H/R injury by MSC-derived exosomes, H9c2 cells after
H/R and exosome treatment were treated with 3-MA, a known inhibitor
of cell autophagy. The results demonstrated that H9c2 cell
proliferation after exosome treatment and following H/R was
significantly suppressed by a subsequent 3-MA treatment (Fig. 5A). In contrast, H9c2 cell apoptosis
after H/R and exosome treatment was greatly increased following
3-MA treatment (Fig. 5B). Beclin-1
and ATG13 expression in H9c2 cells were suppressed by 3-MA
(Fig. 5C). Moreover, while the
expression of the other apoptosis-associated proteins Bcl-2 was
markedly decreased while Apaf1 and Bax were consistently markedly
increased by autophagy inhibition in H9c2 cells (Fig. 5C). The cellular and molecular
alterations induced by 3-MA in H9c2 cells demonstrated that the
inhibition of H/R injury in H9c2 cells by MSC exosomes was mediated
by the promotion of autophagy.
Discussion
Exosomes were first characterized in mammalian
reticulocytes as being important in plasma membrane protein removal
during erythrocyte formation (29).
The major constituents of exosomes are proteins, numerous mRNAs and
miRNAs, and exosomes are widely associated with multiple biological
processes, including immune response regulation, due to their
capability for membrane vesicle trafficking (19,20,29).
Exosomes have been recognized for their therapeutic potential in
injury repair and a variety of human disorders, especially exosomes
derived from multipotent stem cells (19,20).
Exosomes from MSCs have been reported to promote would healing by
activating numerous cellular signaling pathways (30). However, the effects of MSC-derived
exosomes on injuries associated with MI have not been fully
determined, despite reports demonstrating the significant roles of
exosomes isolated from stem and dendritic cells in this process
(22–24). In the present study, MSCs were
isolated from rat bone marrow tissues using a widely applied
method. The identity of MSCs was validated using flow cytometry to
detect CD29 and CD44 expression on the cell surface, which are
previously identified specific biomarkers for MSC-derived exosomes
(31). CD31 and CD34 were also
detected as the negative control. The identification of additional
biomarkers, including CD90 and CD45, would further increase the
characterization specificity. Exosomes isolated from the culture
medium of rat MSCs were further verified using TEM and the
confirmed expression of several exosome-specific proteins. The
isolation and validation of rat MSCs and secreted exosomes provided
a basis for the subsequent analyses.
The establishment of an I/R model by left anterior
descending coronary artery ligation has been successfully applied
in pathogenesis and pharmaceutical studies of MI, due to its ease
of use, low cost and availability of adequate tissue samples for
mechanistic investigation (23). To
explore the effects of rat mesenchymal stem cell-derived exosomes
on MI, rat myocardial I/R was performed and cells were then treated
with exosomes that were isolated from rat MSCs. Using multiple
histological and cellular methods, the results demonstrated that
exosomes from MSCs suppressed the myocardial injury and cell
apoptosis induced by I/R. Apaf1 is a key regulator of the
mitochondrial pathways in cell apoptosis (32), and in the current study it was
indicated that its expression was significantly repressed by
MSC-derived exosomes. Furthermore, ATG13 expression, which promotes
autophagy machinery through the mechanistic target of the mTOR
signaling pathway (33), was
increased by MSC exosome treatment in rats undergoing I/R. These
in vivo assays using I/R rats demonstrated that exosomes
secreted by MSCs could inhibit the myocardial injuries caused by
MI, possibly by regulating autophagy.
The rat myoblast cell line H9c2 is a beneficial
model used to study myocardial physiology and pathogenic processes,
including MI (27,34). To provide evidence of MSC exosome
inhibition of MI-related cell damage, H9c2 cells were treated with
H/R. Using this cellular model, it was demonstrated that MSC
exosomes inhibited the suppression of cell proliferation and
migration during H/R, and also suppressed the myocardiocyte
apoptosis that was induced by H/R. Furthermore Apaf1 and ATG13 gene
expression, as well as a number of apoptosis-associated and
autophagy-associated functional proteins in H9c2 cells treated with
MSC exosomes following H/R, indicated alterations that were
consistent with the rat I/R models. These results further validated
the significant regulatory effects of MSC exosomes on myocardial
injuries that are associated with infarctions. Together with other
evidence obtained using exosomes from embryonic stem cells and
dendritic cells (22,23), it is reasonable to conclude that
exosomes secreted by certain cell types might be useful as
therapeutic agents for use in MI treatment.
The autophagy process is associated with the
pathogenic progression during MI (35,36). The
results of the current study revealed that the inhibition of
autophagy using 3-MA mitigated the injury-suppressing effects of
MSC exosomes on H9c2 cells, demonstrating that the autophagy
process serves a critical role in the pathogenesis of MI, as well
as in exosome-mediated infarction-induced tissue damage. It was
also suggested that modulation of autophagy progression might be an
effective strategy for the treatment of MI. The evidence of the
current study, of autophagy mediating infarction-induced tissue
damage, is limited by a lack of in vivo investigation. The
roles of autophagy, in MI inhibition, by MSC-derived exosomes need
further determination using animal models treated with a
combination of exosomes and autophagy inhibitors.
In summary, animal and cellular models were used to
validate exosome secretion by rat MSCs and if these exosomes could
exert inhibitory effects on myocardiocyte apoptosis associated with
MI, which is mediated by the regulation of autophagy machinery.
Conclusions of the present study provided a basis for the
development of a novel exosome-based therapy for MI.
Acknowledgements
Not applicable.
Funding
The current study was supported by the National
Natural Science Foundation of China (grant no. 81370214).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LZ, XM and CP conceived and supervised the study.
LZ, XM and SL designed and performed experiments. SL, BW and YC
analyzed the data. LZ, BW, YC and CP drafted the manuscript and
made revisions. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal Ethics
Committee of The Third Affiliated Hospital of Sun Yat-sen
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Thygesen K, Alpert JS and White HD; Joint
ESC/ACCF/AHA/WHF Task Force for the Redefinition of Myocardial
Infarction, : Universal definition of myocardial infarction. Eur
Heart J. 28:25252008.
|
|
2
|
GBD 2016 Disease, Injury Incidence and
Prevalence Collaborators: Global, regional, and national incidence,
prevalence, and years lived with disability for 328 diseases and
injuries for 195 countries, 1990–2016: A systematic analysis for
the Global Burden of Disease Study 2016. Lancet. 390:1211–1259.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Levine GN, Dai X, Henry TD, Calfon Press
M, Denktas AE, Garberich RF, Jacobs AK, Jaski BE, Kaul P, Kontos
MC, et al: In-hospital ST-segment elevation myocardial infarction:
Improving diagnosis, triage, and treatment. JAMA Cardiol.
3:527–531. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Varghese T, Hayek SS, Shekiladze N,
Schultz WM and Wenger NK: Psychosocial risk factors related to
ischemic heart disease in women. Curr Pharm Des. 22:3853–3870.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Abbate A, Salloum FN, Vecile E, Das A,
Hoke NN, Straino S, Biondi-Zoccai GG, Houser JE, Qureshi IZ, Ownby
ED, et al: Anakinra, a recombinant human interleukin-1 receptor
antagonist, inhibits apoptosis in experimental acute myocardial
infarction. Circulation. 117:2670–2683. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Riquelme JA, Chavez MN, Mondaca-Ruff D,
Bustamante M, Vicencio JM, Quest AF and Lavandero S: Therapeutic
targeting of autophagy in myocardial infarction and heart failure.
Expert Rev Cardiovasc Ther. 14:1007–1019. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bravo-San Pedro JM, Kroemer G and Galluzzi
L: Autophagy and mitophagy in cardiovascular disease. Circ Res.
120:1812–1824. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ankrum JA, Ong JF and Karp JM: Mesenchymal
stem cells: Immune evasive, not immune privileged. Nat Biotechnol.
32:252–260. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Heirani-Tabasi A, Hassanzadeh M,
Hemmati-Sadeghi S, Shahriyari M and Raeesolmohaddeseen M:
Mesenchymal stem cells; Defining the future of regenerative
medicine. J Genes Cells. 1:34–39. 2015. View Article : Google Scholar
|
|
10
|
Berry MF, Engler AJ, Woo YJ, Pirolli TJ,
Bish LT, Jayasankar V, Morine KJ, Gardner TJ, Discher DE and
Sweeney HL: Mesenchymal stem cell injection after myocardial
infarction improves myocardial compliance. Am J Physiol Heart Circ
Physiol. 290:2196–2203. 2006. View Article : Google Scholar
|
|
11
|
Pak HN, Qayyum M, Kim DT, Hamabe A,
Miyauchi Y, Lill MC, Frantzen M, Takizawa K, Chen LS, Fishbein MC,
et al: Mesenchymal stem cell injection induces cardiac nerve
sprouting and increased tenascin expression in a Swine model of
myocardial infarction. J Cardiovasc Electrophysiol. 14:841–848.
2010. View Article : Google Scholar
|
|
12
|
Price MJ, Chou CC, Frantzen M, Miyamoto T,
Kar S, Lee S, Shah PK, Martin BJ, Lill M, Forrester JS, et al:
Intravenous mesenchymal stem cell therapy early after reperfused
acute myocardial infarction improves left ventricular function and
alters electrophysiologic properties. Int J Cardiol. 111:231–239.
2004. View Article : Google Scholar
|
|
13
|
Mohyeddin-Bonab M, Mohamad-Hassani MR,
Alimoghaddam K, Sanatkar M, Gasemi M, Mirkhani H, Radmehr H, Salehi
M, Eslami M, Farhig-Parsa A, et al: Autologous in vitro expanded
mesenchymal stem cell therapy for human old myocardial infarction.
Arch Iran Med. 10:467–473. 2007.PubMed/NCBI
|
|
14
|
Du YY, Zhou SH, Zhou T, Su H, Pan HW, Du
WH, Liu B and Liu QM: Immuno-inflammatory regulation effect of
mesenchymal stem cell transplantation in a rat model of myocardial
infarction. Cytotherapy. 10:469–478. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deuse T, Peter C, Fedak PW, Doyle T,
Reichenspurner H, Zimmermann WH, Eschenhagen T, Stein W, Wu JC,
Robbins RC and Schrepfer S: Hepatocyte growth factor or vascular
endothelial growth factor gene transfer maximizes mesenchymal stem
cell-based myocardial salvage after acute myocardial infarction.
Circulation 120 (11 Suppl). S247–S254. 2009. View Article : Google Scholar
|
|
16
|
Liang X, Ding Y, Zhang Y, Chai YH, He J,
Chiu SM, Gao F, Tse HF and Lian Q: Activation of NRG1-ERBB4
signaling potentiates mesenchymal stem cell-mediated myocardial
repairs following myocardial infarction. Cell Death Dis.
6:e17652015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Song YS, Joo HW, Park IH, Shen GY, Lee Y,
Shin JH, Kim H and Kim KS: Bone marrow mesenchymal stem
cell-derived vascular endothelial growth factor attenuates cardiac
apoptosis via regulation of cardiac miRNA-23a and miRNA-92a in a
rat model of myocardial infarction. PLoS One. 12:e01799722017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Min PK and Chan SY: The biology of
circulating microRNAs in cardiovascular disease. Eur J Clin Invest.
45:860–874. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Singla DK: Stem cells and exosomes in
cardiac repair. Curr Opin Pharmacol. 27:19–23. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J,
Chen D, Gu J, He X and Huang S: Circular RNA is enriched and stable
in exosomes: A promising biomarker for cancer diagnosis. Cell Res.
25:981–984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Taylor DD and Gerceltaylor C: MicroRNA
signatures of tumor-derived exosomes as diagnostic biomarkers of
ovarian cancer. Gynecol Oncol. 110:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu H, Gao W, Yuan J, Wu C, Yao K, Zhang
L, Ma L, Zhu J, Zou Y and Ge J: Exosomes derived from dendritic
cells improve cardiac function via activation of CD4(+) T
lymphocytes after myocardial infarction. J Mol Cell Cardiol.
91:123–133. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Khan M, Nickoloff E, Abramova T, Johnson
J, Verma SK, Krishnamurthy P, Mackie AR, Vaughan E, Garikipati VN,
Benedict C, et al: Embryonic stem cell-derived exosomes promote
endogenous repair mechanisms and enhance cardiac function following
myocardial infarction. Circ Res. 117:52–64. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lai RC, Arslan F, Lee MM, Sze NS, Choo A,
Chen TS, Salto-Tellez M, Timmers L, Lee CN, El Oakley RM, et al:
Exosome secreted by MSC reduces myocardial ischemia/reperfusion
injury. Stem Cell Res. 4:214–222. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang L and Chan C: Isolation and
enrichment of rat mesenchymal stem cells (MSCs) and separation of
single-colony derived MSCs. J Vis Exp. 37:e1852010.
|
|
26
|
Boxall S and Jones E: The use of
multiparameter flow cytometry and cell sorting to characterize
native human bone marrow mesenchymal stem cells (MSC). Methods Mol
Biol 1235. 121–130. 2015. View Article : Google Scholar
|
|
27
|
Jia X, Zhang L and Mao X: S-propranolol
protected H9C2 cells from ischemia/reperfusion-induced apoptosis
via downregultion of RACK1 Gene. Int J Clin Exp Pathol.
8:10335–10344. 2015.PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Van NG, Porto-Carreiro I, Simoes S and
Raposo G: Exosomes: A common pathway for a specialized function. J
Biochem. 140:13–21. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Silva AM, Teixeira JH, Almeida MI,
Gonçalves RM, Barbosa MA and Santos SG: Extracellular Vesicles:
Immunomodulatory messengers in the context of tissue
repair/regeneration. Eur J Pharm Sci. 98:86–95. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liao W, Yang S, Song C, Li X, Li Y and
Xiong Y: Construction of ureteral grafts by seeding bone marrow
mesenchymal stem cells and smooth muscle cells into bladder
acellular matrix. Transplantat Proc. 45:730–734. 2013. View Article : Google Scholar
|
|
32
|
Yoshida H, Kong YY, Yoshida R, Elia AJ,
Hakem A, Hakem R, Penninger JM and Mak TW: Apaf1 is required for
mitochondrial pathways of apoptosis and brain development. Cell.
94:739–750. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jung CH, Jun CB, Ro SH, Kim YM, Otto NM,
Cao J, Kundu M and Kim DH: ULK-Atg13-FIP200 complexes mediate mTOR
signaling to the autophagy machinery. Mol Biol Cell. 20:1992–2003.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jana S, Patel D, Patel S, Upadhyay K,
Thadani J, Mandal R, Das S and Devkar R: Anthocyanin rich extract
of Brassica oleracea L. alleviates experimentally induced
myocardial infarction. PLoS One. 12:e01821372017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liao LZ, Chen YL, Lu LH, Zhao YH, Guo HL
and Wu WK: Polysaccharide from Fuzi likely protects against
starvation-induced cytotoxicity in H9c2 cells by increasing
autophagy through activation of the AMPK/mTOR pathway. Am J Chin
Med. 41:353–367. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lu L, Wu W, Yan J, Li X, Yu H and Yu X:
Adriamycin-induced autophagic cardiomyocyte death plays a
pathogenic role in a rat model of heart failure. Int J Cardiol.
134:92009. View Article : Google Scholar : PubMed/NCBI
|