Introduction
Skeletal muscle injury is a common injury in daily
life and/or during physical exercise. Skeletal muscle has the
remarkable ability to self-regenerate following injury. The
mechanism of skeletal muscle repair is one of the major issues
surrounding the field of sports medicine. In particular, skeletal
muscle contusion is a common form of injury. It is a contact injury
caused mainly by an acute, relatively large blunt trauma that is
characterized by intact skin and no external damage. The repair of
damaged skeletal muscle is a complex process which mainly consists
of the inflammatory response, myofiber regeneration, angiogenesis
and fibrosis (1). The first phase
occurs in the first few days after injury, characterized by muscle
fiber rupture, necrosis and infiltration of inflammatory cells. The
second phase entails myofiber regeneration, consisting of the
phagocytosis of necrotic muscle fibers and formation of new muscle
fibers (2). The last phase, namely
tissue remodeling, is characterized by the maturation of
regenerating myofibers and formation of scar tissue (3–5).
Effective repair of damaged skeletal muscle requires
the coordinated action of several cell types and a variety of
factors. For example, macrophages serve complex roles in damaged
skeletal muscle, and may be involved in all phases of skeletal
muscle regeneration mentioned above (6). Inflammatory factors including
transforming growth factor (TGF)-β1, interleukin (IL)-10, IL-6,
IL-1β, tumor necrosis factor (TNF)-α and interferon (IFN)-γ,
produced by macrophages and monocytes, also have the potential to
influence muscle repair and regeneration by modulating the
proliferation and differentiation of satellite cells in the injured
tissue (3). In addition, other
physiological processes involved in muscle regeneration, namely
myoblast proliferation, migration and subsequent fusion into
myotubes, are under the control of a number of regulatory factors
including growth factors and myogenic regulatory factors (MRFs),
which constitute the key determinants of the progression of
satellite cell activation during myogenesis and muscle regeneration
(7–9).
In previous years, the roles of long non-coding RNAs
(lncRNAs) have become the focus of research. lncRNAs, which can,
are non-coding RNAs with a transcript length of >200
nucleotides, which have emerged as an important class of regulators
of gene expression, and localize to the nucleus and the cytosol
(10,11). lncRNAs participate in various of
molecular regulatory processes including transcriptional and
post-transcriptional regulation, protein localization, telomere
replication and RNA interference (12). Accumulating evidence from myoblast
differentiation in vitro, cardiotoxin (CTX)-mediated injury
or mdx mice models suggested that certain lncRNAs, including
metastasis-associated lung adenocarcinoma transcript 1 (Malat1),
H19, long intergenic non-protein coding RNAs (linc)-muscle
differentiation 1 (linc-MD1), linc-yin yang 1 (linc-YY1), sirtuin
1-antisense (Sirt1 AS) lncRNA and myogenesis-associated lnc
(lnc-mg), can modulate myogenesis and muscle regeneration (13–16).
However, few studies have evaluated the roles of lncRNAs in
contused muscle (17). In addition,
the association between lncRNAs and macrophages, inflammatory
factors and angiogenic factors in the regeneration of contused
skeletal muscle remains unclear. Therefore, the aim of the present
study was to determine whether lncRNAs may be involved in the
repair of skeletal muscle following contusion injury.
Materials and methods
Animals
A total of 40, 8 week old C57BL/6 male mice weighing
18.2–22.9 g, purchased from JiesiJie-Lab Animal Research Center
(Shanghai JiesiJie Experimental Animal Co., Ltd.), were housed at
21±2°C and 50±5% humidity on a 12 h light/dark cycle, and received
water and food ad libitum. Following acclimatization to the local
environment for 7 days, the mice were randomly divided into two
groups: The uninjured control group (group C) and the muscle
contusion group (group M). Mice from group M were used for the
induction of contusion injury. All experimental protocols were
approved by the Ethics Review Committee for Animal Experimentation
of Shanghai University of Sports (approval no. 2016006).
Contusion injury model induction
A simple and reproducible muscle contusion model in
mice was applied as previously described with little modification
(5). Prior to contusion, mice were
anesthetized with 400 mg/kg chloral hydrate administered
intraperitoneally. The knee joints of the mice were placed in the
extension position at 0° while the ankle joints were placed in the
back-extension position at 90°. A 16.8 g stainless steel ball
(diameter, 1.59 cm) was dropped from a height of 125 cm through a
tube (interior diameter of the tube, 1.60 cm) onto an impactor
(surface, 28.26 mm2) resting on the middle of the
gastrocnemius muscle (GM) of the mice, resulting in an acute
skeletal muscle injury (5). The
muscle contusion created by this method was a high-energy blunt
trauma that resulted in the formation of a large hematoma, breakage
of muscle fibers, exudation of red blood cells and infiltration of
inflammatory cells. This was followed by acute inflammatory
reactions and extensive muscle regeneration (10), a healing process that is comparable
to that observed in humans (18).
All mice were sacrificed for GM isolation at days 3, 6, 12 and 24
following the induction of contusion injury.
Hematoxylin and eosin (H&E)
staining
At days 3, 6, 12 and 24 following muscle contusion,
the right GM was harvested, fixed in 4% paraformaldehyde at 4°C for
24 h and then embedded in paraffin (n=6 mice/group). Cross sections
cut at 4 µm were produced from the GM, which were subsequently
stained with H&E to evaluate the general morphology using a
method described previously (5).
Images were captured for each muscle section using a brightfield
microscope (magnification, ×200; Labophot-2 microscope; Nikon
Corporation).
Masson's trichrome staining
To visualize fibrosis in the muscle injury sites,
Total collagen staining was performed to detect fibrosis in injured
muscle via Masson's trichrome staining (total collagen staining;
Servicebio, Inc.). The procedure was as follows: GM tissue samples
were cut into 4-µm-thick sections and stained with hematoxylin for
5 min, 1% hydrochloric acid alcohol for 5 sec, Biebrich
scarlet-acid fuchsin for 8 min, Phosphomolybdic acid aqueous
solution for 4 min, Aniline blue solution for 5 min, and 1% glacial
acetic acid for 1 min. All staining was performed at room
temperature. Following Masson's trichrome staining, images were
captured for each muscle section viewed under a bright-field
microscope (magnification, ×400; Labophot-2; Nikon Corporation).
The ratio of the fibrotic area to the total cross-sectional area of
the muscle was calculated to estimate the extent of fibrosis
formation using Image Pro 6.0 (Media Cybernetics, Inc.). A total of
six different fields of view (magnification, ×400) were randomly
selected from each section.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA from the GM was extracted using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.),
and the concentration and purity were determined by measuring the
absorbance at 260 and 280 nm with a microplate reader (Model 550
Microplate Reader; Bio-Rad Laboratories, Inc.). Total RNA (2 µg)
was subsequently reverse transcribed into complementary cDNA (cDNA)
using the Revertaid First Strand cDNA Synthesis kit (Thermo Fisher
Scientific, Inc.). The temperature protocol for RT was as follows:
25°C for 5 min followed by 42°C for 60 min, termination at 70°C for
5 min and cooling at 4°C. The qPCR reaction system included SYBR
Green (Fermentas; Thermo Fisher Scientific, Inc.), nuclease-free
water, upstream and downstream primers (designed and synthesized by
Shanghai Shenggong Biology Engineering Technology Service, Ltd.;
primer sequences presented in Table
I) and 1 µl cDNA, made to a total volume of 20 µl/well. An
Applied Biosystems 7500 Real-Time PCR System (Thermo Fisher
Scientific, Inc.) was used for amplification by applying the
following parameters: Denaturation at 95°C for 10 min, 40 cycles of
priming at 95°C for 15 sec, and annealing/extension at 60°C for 1
min. Relative expression values were calculated using the
comparative quantification cycle (2−ΔΔCq) method and
GAPDH was used as the reference gene (19).
 | Table I.Primers used for reverse
transcription-quantitative PCR. |
Table I.
Primers used for reverse
transcription-quantitative PCR.
| Target gene | Forward primer
sequence | Reverse primer
sequence |
|---|
| CD68 |
5′-CAAAGCTTCTGCTGTGGAAAT-3′ |
5′-GACTGGTCACGGTTGCAAG-3′ |
| CD163 |
5′-GCAAAAACTGGCAGTGGG-3′ |
5′-GTCAAAATCACAGACGGAGC-3′ |
| CD206 |
5′-GGATTGTGGAGCAGATGGAAG-3′ |
5′-CTTGAATGGAAATGCACAGAC-3′ |
| IFN-γ |
5′-GCTTTGCAGCTCTTCCTCAT-3′ |
5′-GTCACCATCCTTTTGCCAGT-3′ |
| TNF-α |
5′-CTTCTGTCTACTGAACTTCGGG-3′ |
5′-CACTTGGTGGTTTGCTACGAC-3′ |
| IL-1β |
5′-TGACGTTCCCATTAGACAACTG-3′ |
5′-CCGTCTTTCATTACACAGGACA-3′ |
| IL-6 |
5′-GAACAACGATGATGCACTTGC-3′ |
5′CTTCATGTACTCCAGGTAGCTATGGT-3′ |
| TGF-β1 |
5′-TGCGCTTGCAGAGATTAAAA-3′ |
5′-CGTCAAAAGACAGCCACTCA-3′ |
| IL-10 |
5′-CAAGGAGCATTTGAATTCCC-3′ |
5′-GGCCTTGTAGACACCTTGGTC-3′ |
| Myo D |
5′-GAGCGCATCTCCACAGACAG-3′ |
5′-AAATCGCATTGGGGTTTGAG-3′ |
| Myogenin |
5′-CCAGTACATTGAGCGCCTAC-3′ |
5′-ACCGAACTCCAGTGCATTGC-3′ |
| Myf5 |
5′-GGAATGCCATCCGCTACATT-3′ |
5′-CGTCAGAGCAGTTGGAGGTG-3′ |
| Myf6 |
5′-CCTCAGCCTCCAGCAGTCTT-3′ |
5′-TTCTCCACCACCTCCTCCAC-3′ |
| VEGF |
5′-TAACAGTGAAGCGGAGTG-3′ |
5′-TTTGACCCTTTCCCTTTCCTCG-3′ |
| HIF-1α |
5′-GGCGAGAACGAGAAGAAAAAGATGA-3′ |
5′-GCTCACATTGTGGGGAAGTGG-3′ |
| Angpt1 |
5′-AACCGGATTCAACATGGGCA-3′ |
5′-GAGCGTTGGTGTTGTACTGC-3′ |
| Malat1 |
5′-CACTTGTGGGGAGACCTTGT-3′ |
5′-TGTGGCAAGAATCAAGCAAG-3′ |
| H19 |
5′-TGACTTCATCATCTCCCTCCTGTC-3′ |
5′-GGGTAAATGGGGAAACAGAGTCAC-3′ |
| lnc-mg |
5′-CTGCATCACGGAAGGAGATA-3′ |
5′-AACAATCCATCCTCATTGGC-3′ |
| Sirt1 AS |
5′-AATCCAGTCATTAAACGGTCTACAA-3′ |
5′-TAGGACCATTACTGCCAGAGG-3′ |
| linc-MD1 |
5′-GCAAGAAAACCACAGAGGAGG-3′ |
5′-GTGAAGTCCTTGGAGTTTGAGCA-3′ |
| Linc-YY1 |
5′-AGTTACAGGGAAGTTTGGGCTAC-3′ |
5′-AGGCAAAGGACGGCTGTGAG-3′ |
| GAPDH |
5′-ACTCCACTCACGGCAAATTC-3′ |
5′-TCTCCATGGTGGTGAAGACA-3′ |
Statistical analysis
All data were analyzed using the SPSS 22.0 software
(IBM Corp.) and are presented as the mean ± standard deviation of
at least three experiments. Statistical analysis was carried out
using one-way analysis of variance, and post-hoc multiple
comparisons were performed using the Bonferroni test. Image Pro 6.0
software was used to assess fibrosis, which was compared using an
independent samples t-test. Correlations were calculated according
to Pearson's correlation coefficient. P<0.05 was considered to
indicate a statistically significant difference.
Results
Evaluation of skeletal muscle repair
following contusion injury by H&E staining
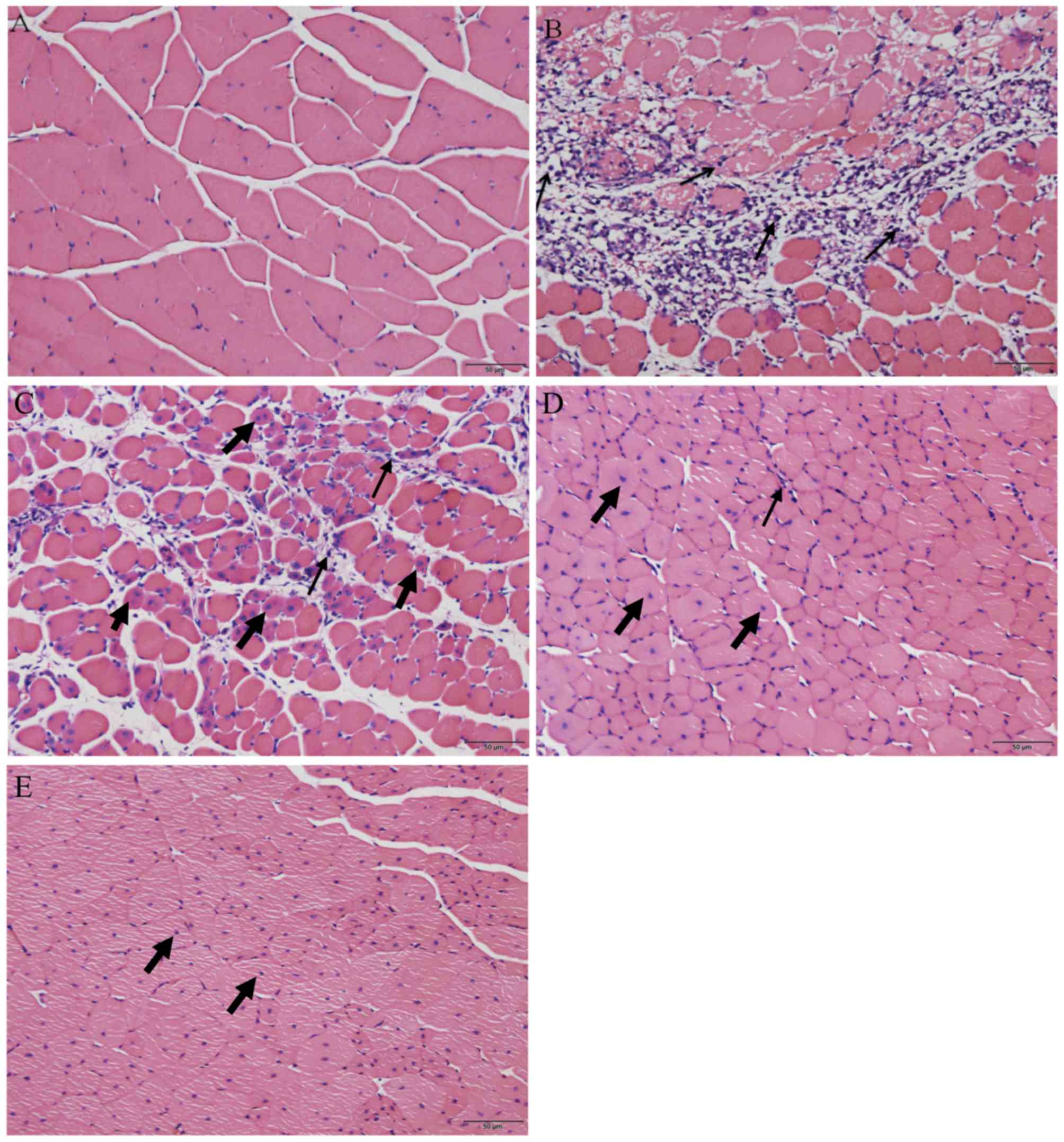
Following H&E staining, the histological
appearance of the skeletal muscle was compared between the
uninjured control group and the muscle contusion group. Skeletal
muscles that were not injured exhibited cells that were arranged
regularly with the nuclei, stained blue-black, located primarily in
the cell periphery (Fig. 1A). On day
3 following injury induction, a greater number of inflammatory
cells and necrotic muscle fibers were observed (Fig. 1B). However, 6 days after contusion
injury, the necrotic muscle fibers had been replaced mostly by
muscle fibers containing centrally localized nuclei or
polynucleated myoblasts/myotubes in the injured areas (Fig. 1C). In addition, inflammatory cells
gradually disappeared from the injury site from day 6 to day 24
(Fig. 1C-E). On day 12 following
injury induction, a small number of developing myofibers with
centrally localized nuclei could be observed in the injured area
(Fig. 1D). Finally, on day 24
post-injury, the regenerated fibers appeared to have matured with
their nuclei having migrated from the center of the cell to the
periphery (20) (Fig. 1E). This indicates that muscle
regeneration was on the verge of completion on day 24 following
injury.
Fibrosis during damaged skeletal
muscle repair
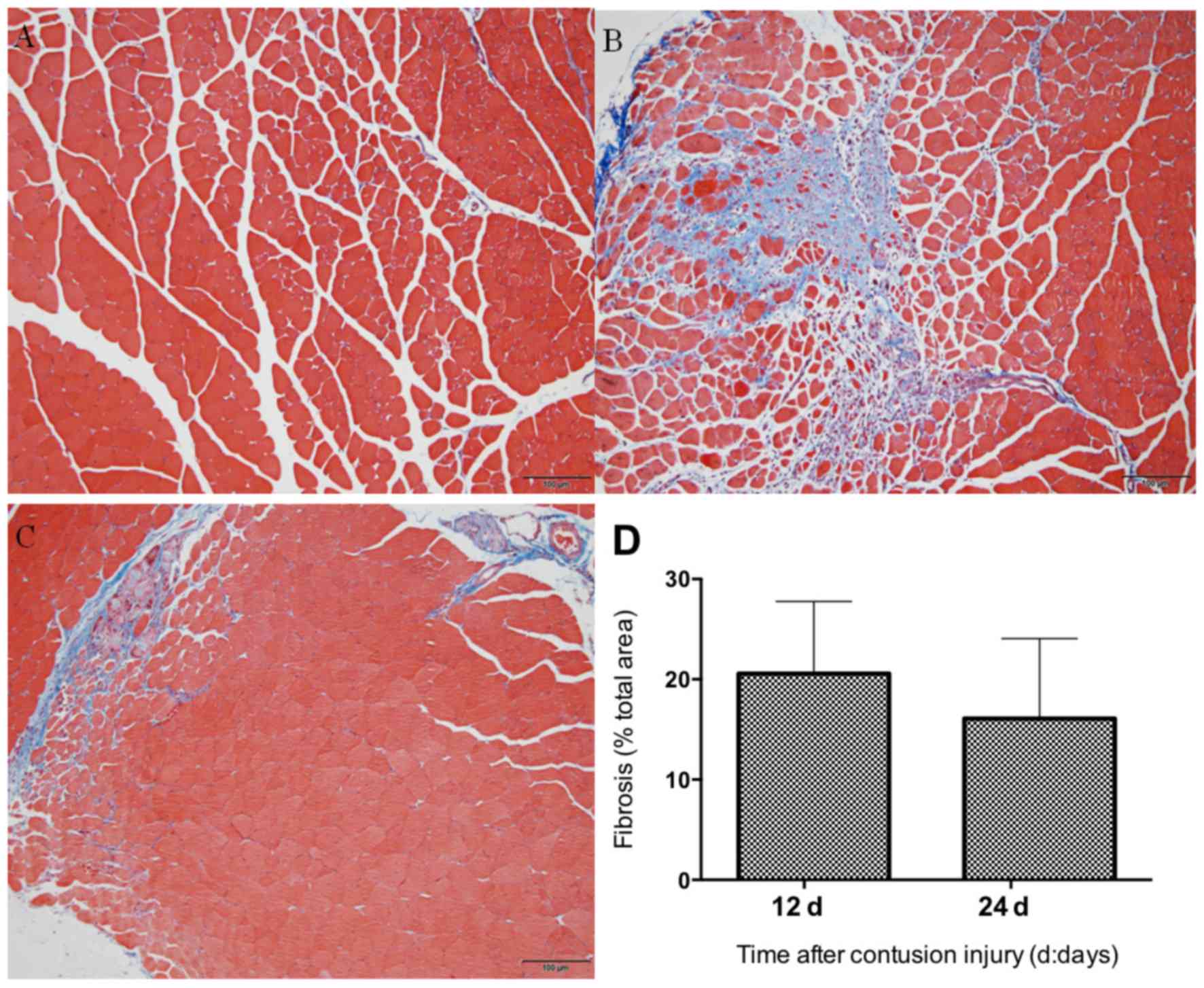
Following Masson's trichrome staining, the tissue in
the injured area of the GM was assessed. Fibrotic scar tissues, in
the form of collagen, were stained in blue, whereas skeletal muscle
cells were stained in red (Fig. 2).
Little or no blue collagen fibers were observed in the uninjured
muscle (Fig. 2A), whereas intense
deposition of blue collagen fibers were noted surrounding the
regenerating myofibers 12 days after the induction of contusion
injury (Fig. 2B). The fibrotic area
in the muscle contusion group on day 24 post-injury was slightly
smaller compared with that in day 12 post-injury (Fig. 2C), but the difference was not
statistically significant (P>0.05; Fig. 2D).
Expression of macrophage-specific
markers following skeletal muscle injury
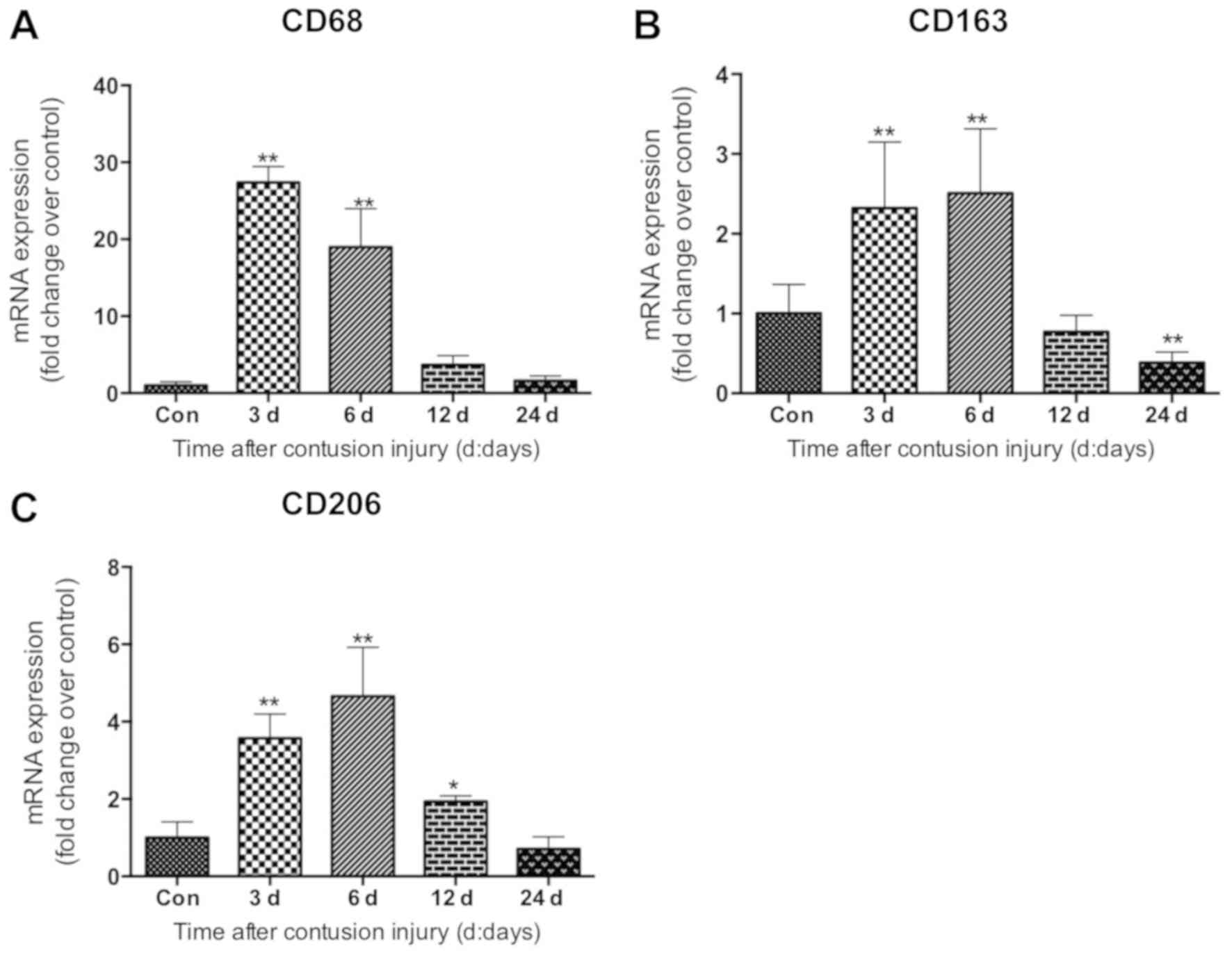
The mRNA levels of specific markers of macrophages
in muscle were evaluated. Compared with the uninjured control
group, the mRNA levels of CD68, which is a specific marker of M1
macrophages (21), increased
significantly in the muscle samples on days 3 and 6 following
injury induction (both P<0.01), peaking at 3 days post-injury
(Fig. 3A). The data also revealed
that the mRNA levels of CD163, a molecular marker of M2 macrophages
(M2c) (21), increased significantly
on days 3 and 6 (both P<0.01) after injury, peaking on day 6
post-injury (Fig. 3B). Similarly,
the mRNA levels of CD206, another marker of the M2 macrophage (M2a
and M2c) subset (22), increased
significantly on days 3, 6 and 12 (all P<0.05) after injury
induction compared with the uninjured control group (Fig. 3C).
Expression of inflammatory cytokines
following skeletal muscle injury
The present study evaluated the expression of
inflammatory cytokines (IL-1β, IL-6, TNF-α, INF-γ, IL-10 and
TGF-β1) in isolated GM samples. The mRNA levels of proinflammatory
cytokines IL-1β, TNF-α, and IFN-γ increased significantly in
skeletal muscle samples on days 3 and 6 following contusion
compared with control (all P<0.01; Fig. 4A, C and D). The mRNA levels of IL-6
also increased significantly on days 3 and 6 after injury compared
with the control (both P<0.01; Fig.
4B), and were higher compared with those in the uninjured
control group at 24 days post-injury (P<0.05; Fig. 4B). The levels of TGF-β1 mRNA were
significantly higher in the GM muscle samples at 3, 6 and 12 days
after injury compared with those in the uninjured control group
(all P<0.01; Fig. 4F). Lastly,
the mRNA levels of the anti-inflammatory factor IL-10 increased
significantly at 3 and 6 days after injury compared with the
uninjured control group, and returned to a level comparable to that
of the control by 24 days (all P<0.01; Fig. 4E).
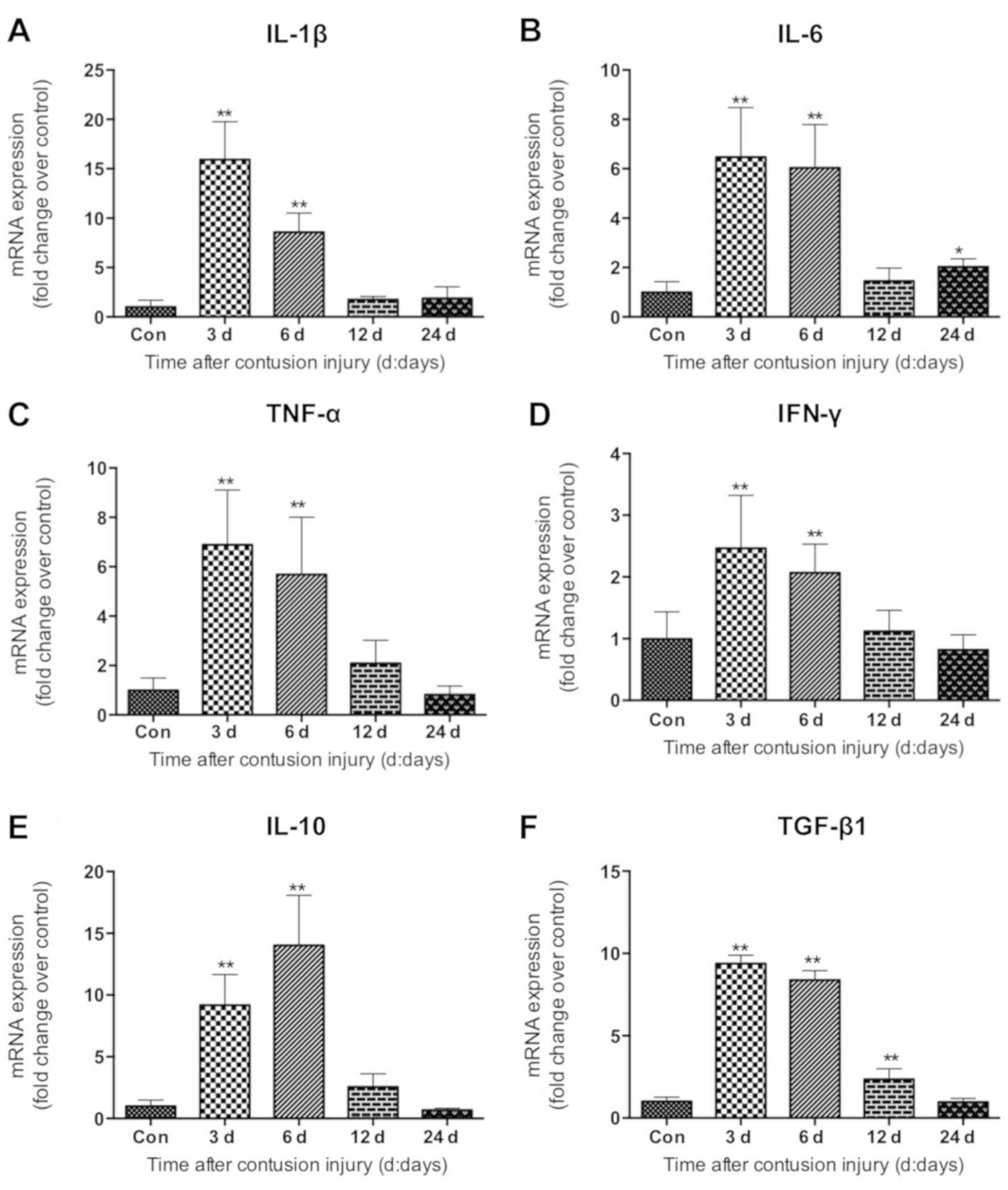 | Figure 4.Expression of inflammatory factors in
gastrocnemius muscle samples following muscle contusion. mRNA
expression levels of (A) IL-1β, (B) IL-6, (C) TNF-α, (D) IFN-γ, (E)
IL-10 and (F) TGF-β1. Data are presented as the mean ± standard
deviation (n=8). *P<0.05, **P<0.01 vs. con. Con, control;
IL-1β, interleukin-1β; IL-6, interleukin-6; TNF-α, tumor necrosis
factor-α; IFN-γ, interferon-γ; IL-10, interleukin-10; TGF-β1,
transforming growth factor-β1. |
Expression of myogenic regulatory
factors following skeletal muscle injury
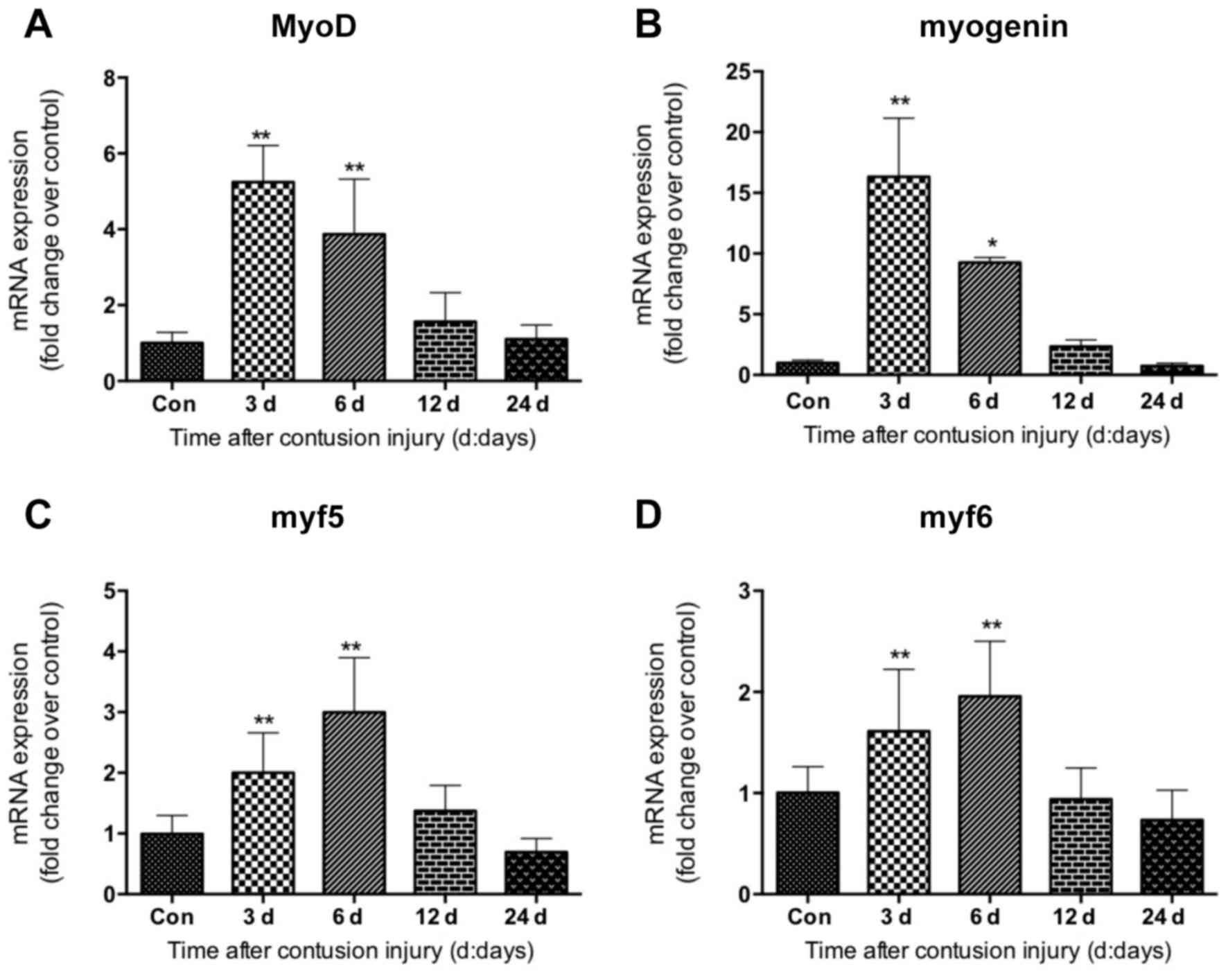
The expression of myogenic regulatory factors
including MyoD, myogenin, myf5 and myf6 was investigated in GM
samples following contusion injury. The data revealed that MyoD,
myogenin, myf5 and myf6 displayed similar gene expression patterns.
Their mRNA levels were elevated significantly at 3 and 6 days after
injury compared with uninjured control, which returned to normal 24
days after injury (Fig. 5). The
expression of MyoD and myogenin peaked at 3 days following injury
induction (Fig. 5A and B); whereas
the levels of myf5 and myf6 peaked on day 6 following injury
(Fig. 5C and D).
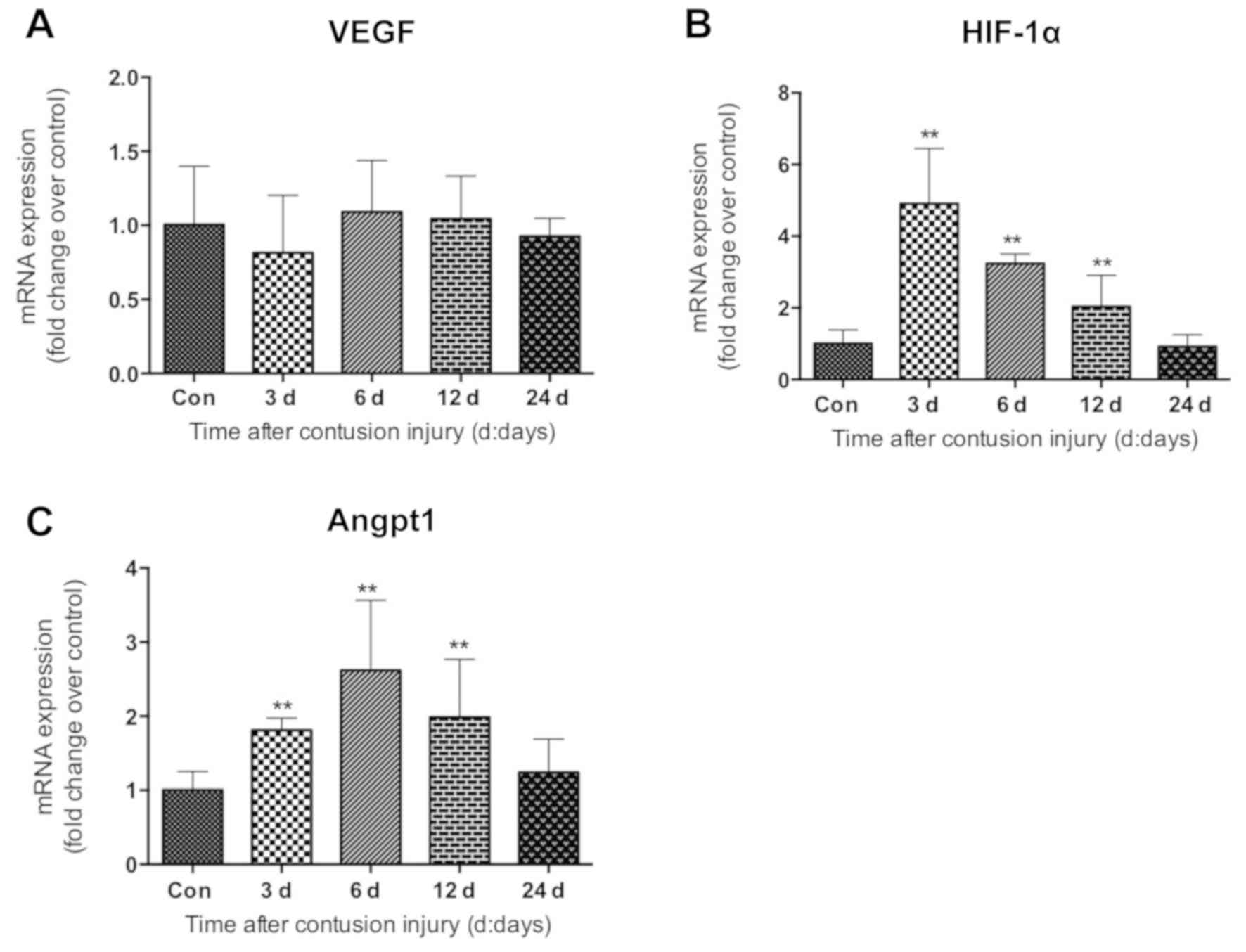
Expression of angiogenic factors
following skeletal muscle injury
Next, the expression of angiogenic factors was
evaluated in the skeletal muscle tissues isolated following muscle
contusion injury. Vascular endothelial growth factor (VEGF),
hypoxia-inducible factor-1α (HIF-1α) and angiopoietin-1 (Angpt-1)
exhibited differential expression patterns. The mRNA levels of VEGF
did not appear to be significantly altered during the healing
process following muscle injury (Fig.
6A). However, compared with the uninjured control group, HIF-1α
mRNA levels were revealed to be significantly increased at 3, 6 and
12 days after injury induction (all P<0.01), peaking on day 3
(Fig. 6B). Increased expression was
observed for Angpt-1 at days 3, 6 and 12 after injury, which was
significantly higher compared with that in the uninjured control
group (all P<0.01; Fig. 6C).
Expression of lncRNAs following
skeletal muscle injury
The expression levels of lncRNAs (Malat1, H19,
lnc-mg, Sirt1 AS, linc-MD1 and linc-YY1) during GM regeneration
were subsequently determined using RT-qPCR. The expression levels
of linc-MD1 and Sirt1 AS were significantly increased compared with
the uninjured control group at 3, 6 and 12 days following injury
(all P<0.01), and returned to normal levels 24 days after injury
(Fig. 7D and E). Compared with the
uninjured control group, Malat1 expression in the skeletal muscle
of the muscle contusion group also increased on day 3 following
injury induction (P<0.01), and remained elevated on day 6
(P<0.01; Fig. 7A). linc-YY1 and
H19 exhibited similar gene expression patterns, as both were
elevated significantly 6 days post-injury compared with the control
(both P<0.01; Fig. 7B and F).
However, their gene expression levels did not change significantly
3, 12 or 24 days post-injury in the muscle contusion group compared
with the uninjured control group. It was additionally demonstrated
that lnc-mg mRNA levels did not appear to be significantly altered
during the process of regeneration following muscle contusion
injury, although the levels were observed to be decreased 12 days
following injury compared with the control group (P<0.01;
Fig. 7C).
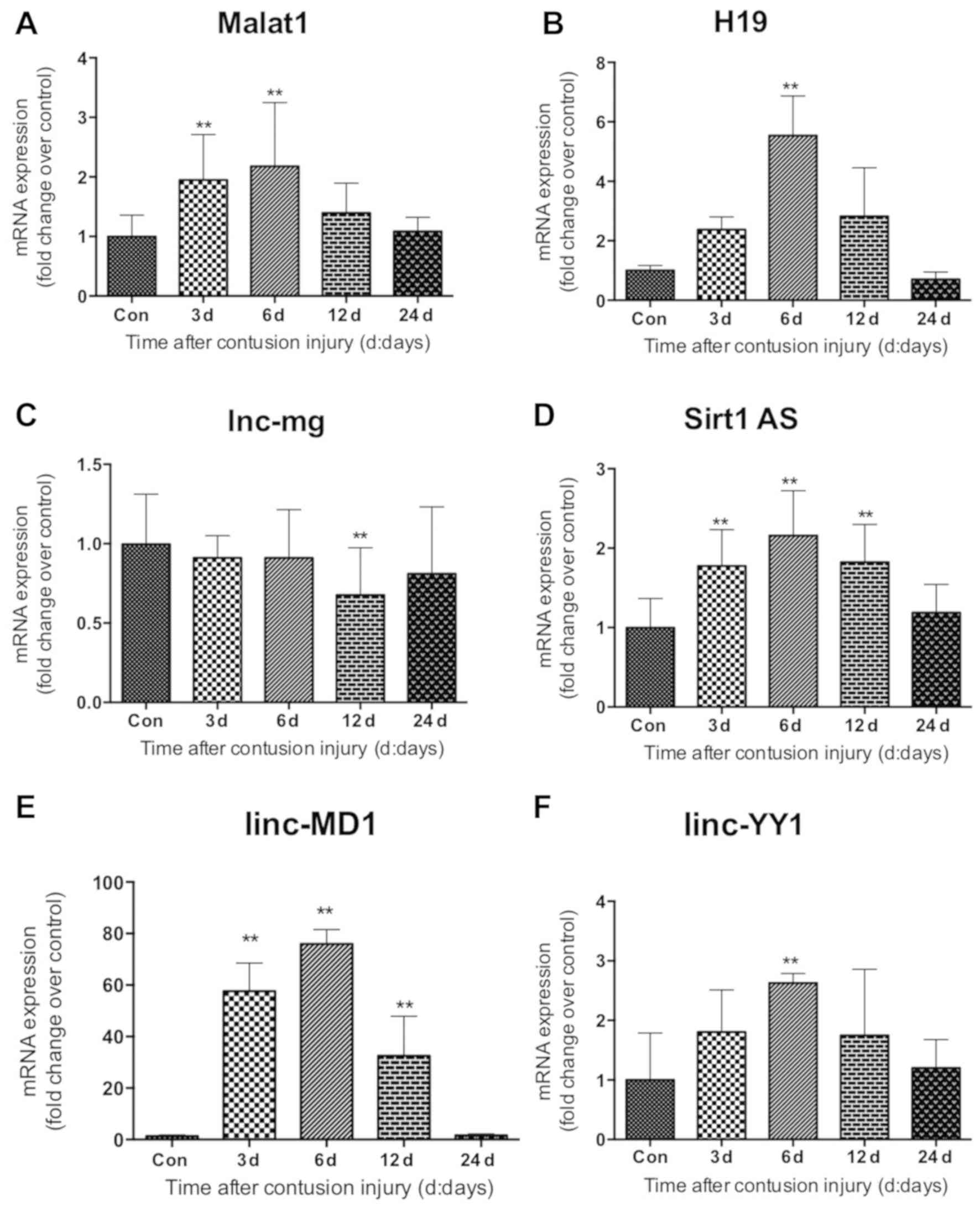 | Figure 7.Expression of long non-coding RNAs in
gastrocnemius muscle samples following muscle contusion. mRNA
expression levels of (A) Malat1, (B) H19, (C) lnc-mg, (D) Sirt1 AS,
(E) linc-MD1 and (F) linc-YY1. Data are presented as the mean ±
standard deviation (n=8). **P<0.01 vs. Con. Con, control;
Malat1, metastasis associated lung adenocarcinoma transcript 1;
lncRNA, long non-coding RNA; lnc-mg, myogenesis-associated long
non-coding RNA; Sirt1 AS, sirtuin 1-antisense; linc-MD1, long
intergenic non-protein coding RNAs-muscle differentiation 1;
linc-YY1, long intergenic non-protein coding RNA-yin yang 1. |
Correlation between the lncRNAs and
the specific markers of macrophages, inflammatory cytokines,
myogenic regulatory factors and angiogenic factors
To assess the association between lncRNAs and
specific markers of macrophages, inflammatory cytokines, myogenic
regulatory factors and angiogenic factors, Pearson's correlations
analysis was performed. The results of this analysis are summarized
in Table II. Positive correlations
were revealed between Malat1 and inflammatory cytokines (TGF-β1,
IL-10, IL-6 and TNF-α), myogenic regulatory factors (MyoD and
myogenin) and angiogenic factors (HIF-1α and Angpt1) (Table II). Although both H19 and Sirt1 AS
were demonstrated to correlate positively with myogenic regulatory
factors (myogenin, myf5 and myf6), only H19 correlated with
angiogenic factors (HIF-1α and Angpt1), whereas only Sirt1 AS was
correlated with MyoD (Table II).
Significant positive correlations were revealed between linc-MD1
and macrophage markers (CD163 and CD206), inflammatory cytokines
(TGF-β1, IL-10, IL-6, TNF-α and IFN-γ) and myogenic regulatory
factors (MyoD, myogenin, myf5 and myf6). A moderate correlation was
observed between linc-YY1 and the myogenic regulatory factors (MyoD
and myogenin). No correlation was found between lnc-mg and any of
the genes tested (Table II).
| Table II.Correlation between the lncRNAs and
the specific markers of macrophages, inflammatory cytokines,
myogenic regulatory factors and angiogenic factors. |
Table II.
Correlation between the lncRNAs and
the specific markers of macrophages, inflammatory cytokines,
myogenic regulatory factors and angiogenic factors.
|
|
lncRNA |
|---|
|
|
|
|---|
|
| Malat1 | H19 | lnc-mg | Sirt1 AS | linc-MD1 | linc-YY1 |
|---|
|
|
|
|
|
|
|
|
|---|
| Gene | r | P-value | r | P-value | r | P-value | r | P-value | r | P-value | r | P-value |
|---|
| CD68 | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. |
| CD163 | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 0.793 | 0.015 | N.S. | N.S. |
| CD206 | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 0.862 | 0.002 | N.S. | N.S. |
| TGF-β1 | 0.916 | 0.029 | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 0.912 | <0.001 | N.S. | N.S. |
| IL-10 | 0.986 | <0.001 | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 0.896 | <0.001 | N.S. | N.S. |
| IL-6 | 0.598 | 0.005 | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 0.850 | 0.024 | N.S. | N.S. |
| IL-1β | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. |
| TNF-α | 0.886 | 0.046 | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 0.906 | 0.034 | N.S. | N.S. |
| IFN-γ | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 0.884 | 0.037 | N.S. | N.S. |
| MyoD | 0.558 | 0.003 | N.S. | N.S. | N.S. | N.S. | 0.563 | <0.001 | 0.825 | <0.001 | 0.474 | 0.003 |
| myogenin | 0.600 | 0.012 | 0.470 | 0.003 | N.S. | N.S. | 0.535 | <0.001 | 0.773 | <0.001 | 0.423 | 0.007 |
| Myf5 | N.S. | N.S. | 0.797 | 0.001 | N.S. | N.S. | 0.703 | <0.001 | 0.782 | <0.001 | N.S. | N.S. |
| Myf6 | N.S. | N.S. | 0.674 | 0.007 | N.S. | N.S. | 0.620 | 0.001 | 0.897 | 0.039 | N.S. | N.S. |
| VEGF | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. |
| HIF-1α | 0.785 | 0.016 | 0.504 | 0.001 | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. |
| Angpt1 | 0.653 | 0.040 | 0.593 | <0.001 | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. |
Discussion
Skeletal muscle retains the ability to regenerate
following damage. The present study employed a mouse skeletal
muscle contusion injury model which can induce inflammatory
responses with macrophage infiltration as one of the signatures,
followed by regeneration. Histologically, a large number of
inflammatory cells and factors infiltrated the injured area in the
early stages of skeletal muscle injury. The infiltration patterns
of inflammatory cells were consistent with a previous study
(23). In addition, the necrotic
muscle fibers were replaced mostly by centrally-nucleated
developing muscle fibers or myotubes at day 6 following injury.
This indicated that satellite cells were committed into the
myoblast pathway to differentiate into myotubes following contusion
injury.
Fibrosis may occur when skeletal muscles experience
severe injury, which is characterized by the accumulation of
fibroblasts and myofibroblasts, and high levels of extracellular
matrix deposition (24). According
to the collagen staining performed in the present study, scar
tissues could be observed in the contused muscle 12 and 24 days
after injury; with more fibrotic scars recorded on day 12 than on
day 24. Total collagen staining was performed to detect fibrosis in
the injured muscle as previously described (25). These observations are concordant with
those reported by Ghaly et al (26). Fibrosis of skeletal muscle is a
characteristic feature of skeletal muscle repair, usually beginning
between the 2nd and 3rd week after injury. Resultant scar tissue
continues to develop and mature over time (27). The results of the present study
revealed that fibrosis occurred in the late stages of skeletal
muscle repair (12 and 24 days after injury). This indicates that
the skeletal muscle contusion injury was successfully induced in
the present study, which was followed by muscle repair.
Despite the rapidly increasing number of studies
investigating the functions of lncRNAs, their specific roles in
myogenesis remain poorly defined. The findings presented in this
study provided a comprehensive analysis of lncRNA (Malat1, H19,
lnc-mg, Sirt1 AS, linc-MD1 and linc-YY1) expression during skeletal
muscle regeneration following contusion injury. In addition, their
association with the expression levels of lincRNAs and macrophage
markers, inflammatory cytokines, myogenic factors and angiogenic
factors was elucidated. Malat1 and Sirt1 AS are lincRNAs that are
expressed in high abundance in proliferating and differentiating
myoblasts (15,28). The present study revealed that the
expression of Malat1 and Sirt1 AS were significantly upregulated
throughout the skeletal muscle regeneration process. It has been
suggested previously that Malat1 and Sirt1 AS can promote myoblast
proliferation and inhibit myoblast differentiation (15,16,29).
Therefore, it would be feasible that Malat1 and Sirt1 AS may serve
important functions in the regeneration of contused skeletal
muscle. In addition, Malat1 and Sirt1 AS were revealed to
positively correlate with myogenic transcription factors myogenin
and MyoD. Wang et al (15)
demonstrated that Sirt1 AS lncRNA overexpression downregulated the
expression of MyoD and myogenin; furthermore, Malat1 has been
previously postulated to modulate the expression of myogenin and
the activity of MyoD (16).
Consequently, it may be hypothesized that Malat1 and Sirt1 AS
lncRNA serve key roles in the regeneration of contused muscle,
possibly by regulating the expression of MyoD and myogenin.
Additionally, lnc-mg has also been suggested to be a
skeletal muscle-enriched lncRNA (30). A main purpose of the present study
was to investigate the role of lnc-mg in the regeneration of
skeletal muscle following contusion. The expression of lnc-mg was
significantly decreased on day 12 following contusion. This
suggests that lnc-mg may act as a negative regulator in the
regeneration of skeletal muscle following contusion. This result is
consistent with an earlier finding that the knockdown of lnc-mg
resulted in marked inhibition of muscle satellite cell
differentiation by downregulating MyoD and myogenin expression
(30). This observation supported
the hypothesis that lnc-mg can regulate the expression of MyoD and
myogenin in the present study. However, no correlation was found
between lnc-mg and MyoD or myogenin expression. This discrepancy
could be due to differences in the animal models used in the two
studies. Therefore, further research is necessary to elucidate
whether lnc-mg is involved in skeletal muscle regeneration
following contusion.
Although several lncRNAs have been demonstrated to
serve a number of roles in skeletal muscle cell differentiation and
myogenesis in vitro (13,28,29),
little is known about their function during the regeneration of
skeletal muscle following contusion. Therefore, the present study
also investigated the roles of H19, linc-MD1 and lncYY1 during
skeletal muscle regeneration. The present study revealed that the
expression of linc-MD1 was significantly increased throughout the
skeletal muscle regeneration process, whereas the upregulation of
H19 and lncYY1 was only observed on day 6 after muscle contusion.
This suggests that H19 and lncYY1 may perform important functions
during the early repair phase of contused skeletal muscle. Indeed,
a number of studies have demonstrated that skeletal muscle
deficient in lncRNAs H19, linc-MD1 and/or lncYY1 displays aberrant
skeletal muscle regeneration post-injury due to the downregulation
of MyoD and myogenin (14,31,32).
Data from the present study revealed that H19, linc-MD1 and lncYY1
all correlated positively with MyoD and/or myogenin. Therefore, it
may be hypothesized that H19, linc-MD1 and lnc-YY1 promote the
regeneration of contused skeletal muscle, possibly by modulating
MyoD and myogenin expression.
Myogenic factors Myf5 and Myf6 are essential for
muscle regeneration and can promote myoblast differentiation
(33). However, investigations into
the role of lncRNAs in regulating myf5 and myf6 expression during
contused muscle regeneration are lacking. In the present study, a
positive correlation was revealed between the expression of lncRNAs
(linc-MD1, Sirt AS and H19) and myf5/myf6; however, it remains
unclear how this relationship can impact contused muscle
regeneration. Further studies are required to investigate this
underlying mechanism.
During the skeletal muscle repair process, lncRNAs
have also been reported to be involved in the regulation of the
skeletal muscle inflammatory response, angiogenesis and fibrosis
(34–36). The inflammatory response is an
integral part of the reaction to muscle injury and serves a pivotal
role in subsequent muscle regeneration (37). Macrophages either induce inflammation
or repair damaged tissues by secreting a large quantity of
inflammatory cytokines. M1 macrophages produce TNF-α, IL-1β and
IL-6, while M2 macrophages produce IL-10 and TGF-β1 (21). A number of studies performed
previously have illustrated that Malat1 can regulate the
inflammatory response in skeletal muscle, the knockdown of which
increases the lipopolysaccharide-induced expression of TNF-α and
IL-6 (34,35,38).
These findings suggest that Malat1 may function as a regulator of
the inflammatory response in this organ. The present study revealed
that Malat1 correlated positively with the expression of a number
of inflammatory cytokines (TGF-β1, TNF-α, IL-6 and IL-10) in
contused muscle. As a result, this suggests that Malat1 can
modulate the inflammatory response during the regeneration of
contused skeletal muscle.
Vascular regeneration is part of the complete
regeneration of damaged skeletal muscle. In the present study, the
expression of HIF-1α and Angpt1 was markedly increased following
muscle contusion, which correlated positively with Malat1. Michalik
et al (39) observed that
Malat1-deficient mice displayed a reduction in blood vessel
density, suggesting that Malat1 may be involved in angiogenesis.
Therefore, Malat1 may contribute to angiogenesis in regeneration
after skeletal muscle injury, possibly by modulating the expression
of HIF-1α and Angpt1. However, further research is necessary to
investigate the underlying mechanism.
Recently, a growing body of evidence suggested that
lncRNAs are also involved in tissue fibrosis in several organs,
including the lungs, liver and heart (36,40).
However, the role of lncRNAs in injury-induced skeletal muscle
fibrosis remains unclear. Results from the present study revealed
that linc-MD1 and Malat1 significantly correlated with the
profibrotic factor TGF-β1. Therefore, lncRNAs may be involved in
the fibrosis of contused skeletal muscle by interacting with
TGF-β1. However, further research is necessary to investigate the
underlying mechanism.
lncRNAs such as Malat1 serve important roles in the
inflammatory response and angiogenesis of injured skeletal muscle.
To the best of our knowledge, only a small number of studies have
evaluated the role of lncRNAs in the inflammatory response and
angiogenesis following skeletal muscle injury (34,35). The
present study revealed that a number of lncRNAs are associated with
inflammatory and angiogenic factors. This suggests that the
inflammatory response and angiogenesis during skeletal muscle
regeneration are regulated by lncRNAs.
This present investigation was the first to
demonstrate that lncRNAs are associated with the regeneration of
contused skeletal muscle. The changes in the expression of a number
of candidate lncRNAs at multiple timepoints following skeletal
muscle contusion, as well as their association with other
physiological factors, were assessed. Results illustrated in the
present study support the hypothesis that lncRNAs may play
important roles in the regeneration of contused skeletal muscle,
but further research is needed to elucidate the underlying
mechanism. However, there are several limitations to the study; for
example, knockdown or overexpression experiments on the lncRNAs
were not performed. Although Pearson's correlation analysis
indicated correlations between lncRNAs and macrophage infiltration,
inflammation and angiogenesis, this did not reveal the mechanism
underlying the role of lncRNAs in contused muscle regeneration.
Nevertheless, this present investigation do lay the foundation for
further research into the functional role of lncRNAs in skeletal
muscle regeneration.
In conclusion, the expression of inflammatory
cytokines, myogenic regulatory factors and angiogenic factors were
demonstrated to be significantly increased following the induction
of skeletal muscle contusion, along with lncRNAs including Malat1,
H19, lnc-mg, linc-MD1, linc-YY1 and Sirt1 AS. There was a
correlation between lncRNAs and a variety of established regulatory
factors (TGF-β1, MyoD, myogenin, myf5, myf6, HIF-1α and Angpt1)
during the skeletal muscle regeneration process. These results
suggest that lncRNAs may serve important roles in the regeneration
of damaged skeletal muscle. Effective muscle regeneration is
essential for the treatment of muscle diseases including muscle
atrophy, muscular dystrophy and sporting injuries. Therefore, these
findings serve as a basis for the effective treatment of muscle
atrophy and muscular dystrophy.
Acknowledgements
The present study was previously presented at a
conference (https://ojs.uclouvain.be/index.php/EBR/article/view/8693).
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 31271273 and
31300975), Shanghai Natural Science Fund Project (grant no.
18ZR1437100) and Shanghai Key Laboratory of Human Movement
Development and Protection (Shanghai University of Sport; grant no.
11DZ2261100).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LZ analyzed the results and drafted the manuscript.
LZ performed histological staining and PCR. XL assisted with PCR.
PC and WX designed the current study and provided funds. All
authors reviewed and critiqued the manuscript and agreed to the
final submission of the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics Review
Committee for Animal Experimentation of Shanghai University of
Sports (approval no. 2016006).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lu H, Huang D, Saederup N, Charo IF,
Ransohoff RM and Zhou L: Macrophages recruited via CCR2 produce
insulin-like growth factor-1 to repair acute skeletal muscle
injury. FASEB J. 25:358–369. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carlson BM and Faulkner JA: The
regeneration of skeletal muscle fibers following injury: A review.
Medicine Sci Sports Exerc. 15:187–198. 1983. View Article : Google Scholar
|
|
3
|
Tidball JG: Mechanisms of muscle injury,
repair and regeneration. Compr Physiol. 1:2029–2062.
2011.PubMed/NCBI
|
|
4
|
Baggiolini M: Chemokines in pathology and
medicine. J Intern Med. 250:91–104. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xiao W, Liu Y and Chen P: Macrophage
depletion impairs skeletal muscle regeneration: The roles of
pro-fibrotic factors, inflammation and oxidative stress.
Inflammation. 39:2016–2028. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Angelini G, Salinari S, Bertuzzi A,
Iaconelli A and Mingrone G: Metabolic surgery improves insulin
resistance through the reduction of gut-secreted heat shock
proteins. Commun Biol. 1:692018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zammit PS: Function of the myogenic
regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal
muscle, satellite cells and regenerative myogenesis. Semin Cell Dev
Biol. 72:19–32. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kassar-Duchossoy L, Gayraud-Morel B, Gomès
D, Rocancourt D, Buckingham M, Shinin V and Tajbakhsh S: Mrf4
determines skeletal muscle identity in Myf5: Myod double-mutant
mice. Nature. 431:466–471. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zanou N and Gailly P: Skeletal muscle
hypertrophy and regeneration: Interplay between the myogenic
regulatory factors (MRFs) and insulin-like growth factors (IGFs)
pathways. Cell Mol Life Sci. 70:4117–4130. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
van Heesch S, van Iterson M, Jacobi J,
Boymans S, Essers PB, de Bruijn E, Hao W, MacInnes AW, Cuppen E and
Simonis M: Extensive localization of long noncoding RNAs to the
cytosol and mono- and polyribosomal complexes. Genome Biol.
15:R62014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Batista PJ and Chang HY: Long noncoding
RNAs: Cellular address codes in development and disease. Cell.
152:1298–1307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang J, Shao N, Ding X, Tan B, Song Q,
Wang N, Jia Y, Ling H and Cheng Y: Crosstalk between transforming
growth factor-beta signaling pathway and long non-coding RNAs in
cancer. Cancer Lett. 370:296–301. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xu X, Ji S, Li W, Yi B, Li H, Zhang H and
Ma W: LncRNA H19 promotes the differentiation of bovine skeletal
muscle satellite cells by suppressing Sirt1/FoxO1. Cell Mol Biol
Lett. 22:102017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dey BK, Pfeifer K and Dutta A: The H19
long noncoding RNA gives rise to microRNAs miR-675-3p and
miR-675-5p to promote skeletal muscle differentiation and
regeneration. Genes Dev. 28:491–501. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang GQ, Wang Y, Xiong Y, Chen XC, Ma ML,
Cai R, Gao Y, Sun YM, Yang GS and Pang WJ: Sirt1 AS lncRNA
interacts with its mRNA to inhibit muscle formation by attenuating
function of miR-34a. Sci Rep. 6:218652016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen X, He L, Zhao Y, Li Y, Zhang S, Sun
K, So K, Chen F, Zhou L, Lu L, et al: Malat1 regulates myogenic
differentiation and muscle regeneration through modulating MyoD
transcriptional activity. Cell Discov. 3:170022017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun Y, Sun X, Liu S, Liu L and Chen J: The
overlap between regeneration and fibrosis in injured skeletal
muscle is regulated by phosphatidylinositol 3-kinase/Akt signaling
pathway-A bioinformatic analysis based on lncRNA microarray. Gene.
672:79–87. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Diaz JA, Fischer DA, Rettig AC, Davis TJ
and Shelbourne KD: Severe quadriceps muscle contusions in athletes.
A report of three cases. Am J Sports Med. 31:289–293. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ota S, Uehara K, Nozaki M, Kobayashi T,
Terada S, Tobita K, Fu FH and Huard J: Intramuscular
transplantation of muscle-derived stem cells accelerates skeletal
muscle healing after contusion injury via enhancement of
angiogenesis. Am J Sports Med. 39:1912–1922. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Manferdini C, Paolella F, Gabusi E,
Gambari L, Piacentini A, Filardo G, Fleury-Cappellesso S, Barbero
A, Murphy M and Lisignoli G: Adipose stromal cells mediated
switching of the pro-inflammatory profile of M1-like macrophages is
facilitated by PGE2: In vitro evaluation. Osteoarthritis Cartilage.
25:1161–1171. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tidball JG and Villalta SA: Regulatory
interactions between muscle and the immune system during muscle
regeneration. Am J Physiol Regul Integr Comp Physiol.
298:R1173–R1187. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tonkin J, Temmerman L, Sampson RD,
Gallego-Colon E, Barberi L, Bilbao D, Schneider MD, Musarò A and
Rosenthal N: Monocyte/macrophage-derived IGF-1 orchestrates murine
skeletal muscle regeneration and modulates autocrine polarization.
Mol Ther. 23:1189–1200. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wynn TA and Ramalingam TR: Mechanisms of
fibrosis: Therapeutic translation for fibrotic disease. Nat Med.
18:1028–1040. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shen W, Li Y, Zhu J, Schwendener R and
Huard J: Interaction between macrophages, TGF-beta1 and the COX-2
pathway during the inflammatory phase of skeletal muscle healing
after injury. J Cell Physiol. 214:405–412. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ghaly A and Marsh DR:
Ischaemia-reperfusion modulates inflammation and fibrosis of
skeletal muscle after contusion injury. Int J Exp Pathol.
91:244–255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li Y, Cummins J and Huard J: Muscle injury
and repair. Curr Opin Orthop. 12:409–415. 2001. View Article : Google Scholar
|
|
28
|
Watts R, Johnsen VL, Shearer J and Hittel
DS: Myostatin-induced inhibition of the long noncoding RNA Malat1
is associated with decreased myogenesis. Am J Physiol Cell Physiol.
304:C995–C1001. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Han X, Yang F, Cao H and Liang Z: Malat1
regulates serum response factor through miR-133 as a competing
endogenous RNA in myogenesis. FASEB J. 29:3054–3064. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhu M, Liu J, Xiao J, Yang L, Cai M, Shen
H, Chen X, Ma Y, Hu S, Wang Z, et al: Lnc-mg is a long non-coding
RNA that promotes myogenesis. Nat Commun. 8:147182017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cesana M, Cacchiarelli D, Legnini I,
Santini T, Sthandier O, Chinappi M, Tramontano A and Bozzoni I: A
long noncoding RNA controls muscle differentiation by functioning
as a competing endogenous RNA. Cell. 147:358–369. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou L, Sun K, Zhao Y, Zhang S, Wang X, Li
Y, Lu L, Chen X, Chen F, Bao X, et al: Linc-YY1 promotes myogenic
differentiation and muscle regeneration through an interaction with
the transcription factor YY1. Nat Commun. 6:100262015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Petersson SJ, Jørgensen LH, Andersen DC,
Nørgaard RC, Jensen CH and Schrøder HD: SPARC is up-regulated
during skeletal muscle regeneration and inhibits myoblast
differentiation. Histol Histopathol. 28:1451–1460. 2013.PubMed/NCBI
|
|
34
|
Huang JK, Ma L, Song WH, Lu BY, Huang YB,
Dong HM, Ma XK, Zhu ZZ and Zhou R: LncRNA-MALAT1 promotes
angiogenesis of thyroid cancer by modulating tumor-associated
macrophage FGF2 protein secretion. J Cell Biochem. 118:4821–4830.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Marques-Rocha JL, Samblas M, Milagro FI,
Bressan J, Martinez JA and Marti A: Noncoding RNAs, cytokines and
inflammation-related diseases. FASEB J. 29:3595–3611. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Duru N, Wolfson B and Zhou Q: Mechanisms
of the alternative activation of macrophages and non-coding RNAs in
the development of radiation-induced lung fibrosis. World J Biol
Chem. 7:231–239. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang C, Li Y, Wu Y, Wang L, Wang X and Du
J: Interleukin-6/signal transducer and activator of transcription 3
(STAT3) pathway is essential for macrophage infiltration and
myoblast proliferation during muscle regeneration. J Biol Chem.
288:1489–1499. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhao G, Su Z, Song D, Mao Y and Mao X: The
long noncoding RNA MALAT1 regulates the lipopolysaccharide-induced
inflammatory response through its interaction with NF-kappaB. FEBS
Lett. 590:2884–2895. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Michalik KM, You X, Manavski Y,
Doddaballapur A, Zörnig M, Braun T, John D, Ponomareva Y, Chen W,
Uchida S, et al: Long noncoding RNA MALAT1 regulates endothelial
cell function and vessel growth. Circ Res. 114:1389–1397. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Piccoli MT, Gupta SK, Viereck J,
Foinquinos A, Samolovac S, Kramer FL, Garg A, Remke J, Zimmer K,
Batkai S and Thum T: Inhibition of the cardiac fibroblast-enriched
lncRNA Meg3 prevents cardiac fibrosis and diastolic dysfunction.
Circ Res. 121:575–583. 2017. View Article : Google Scholar : PubMed/NCBI
|