Introduction
MicroRNAs (miRNAs or miRs) are a group of short
non-coding RNAs (19–25 nucleotides in length), which are abundant
in eukaryotic cells (1). miRNAs
regulate post-transcriptional gene expression by binding to the
3′untranslated region (UTR) of target mRNAs and inhibiting their
degradation or translation (1,2). It has
been demonstrated that miRNAs serve a key role in a variety of
biological processes, including cell proliferation, apoptosis and
differentiation (2–5). The abnormal expression of miRNA is
associated with the development and progression of a variety of
diseases including cancer, cardiovascular, metabolic, neurological
and bone associated diseases (6–11).
Bone regeneration is critical in many common bone
diseases, which include trauma and osteoporosis (12). Although progress has been made in the
treatment of these diseases, highly effective therapies are
insufficient (13). A study into the
molecular mechanisms of bone regeneration will therefore aid the
discovery of novel bone disease treatments. In previous studies, it
has been demonstrated that miRNAs serve an important role in the
regulation of osteoblast differentiation or bone formation
(14,15). Tang et al (16) revealed that miR-282 negatively
regulated osteoblastic differentiation by targeting special
AT-rich-sequence-binding protein 2. A study performed by Arfat
et al (17) demonstrated the
inhibitory effect of miR-208a-3p on osteoblast differentiation and
bone formation by targeting activin A receptor type 1. Furthermore,
Zhang et al (18) indicated
that miR-335-5p may promote bone formation and regeneration in
vivo. miR-342-3p was also demonstrated to increase osteogenic
differentiation by inhibiting bone formation or regeneration in
vitro (19). These previous
studies indicated that miRNAs may serve a role in the promotion or
inhibition of bone formation/regeneration. However, the role of
miRNAs in these processes remains largely unclear.
miRNA-98-5p has been reported to play important
roles in cancer cell proliferation, apoptosis, and metastasis in
various types of cancer, including ovarian cancer (20), pancreatic ductal adenocarcinoma
(21), non-small-cell lung cancer
and hepatocellular carcinoma (22,23). It
has also been demonstrated that miR-98-5p serves an important role
in the development of Alzheimer's disease (24). It has also been revealed that
miR-98-5p serves an important role in the differentiation of human
embryonic stem cells (25). The
results of the aforementioned studies indicate that miR-98-5p
serves a crucial role in the regulation of cell proliferation,
apoptosis and differentiation, which are involved in bone
formation/regeneration. However, to the best of our knowledge no
study has assessed the role of miR-98-5p in bone
formation/regeneration.
High mobility group AT-Hook 2 (HMGA2), also known as
HMGI-C, is a nuclear-binding protein, which serves a key role in
the regulation of cell proliferation and differentiation (26). A previous study revealed that HMGA2
knockdown significantly inhibited the proliferation of bone
marrow-derived mesenchymal stem cells (MSCs) (27). However, in recent years, the role of
HMGA2 in osteoblast differentiation has been controversial. Wei
et al (28) demonstrated that
miR let-7 promoted MSC osteogenesis in vitro and in
vivo by inhibiting HMGA2 expression (28). It has also been demonstrated that
miR-495 prevents new bone regeneration by targeting HMGA2 (29). However, the association between
miR-98-5p and HMGA2 has not yet been elucidated.
Therefore, the aim of the current study was to
investigate the potential role of miR-98-5p in bone regeneration
and to further determine the molecular mechanisms, which govern
this.
Materials and methods
Clinical samples
Bone marrow (5–7 ml) was obtained from five male
healthy voluntary donors (22–32 years old) at Department of
Orthopedics, Qinghai Provincial People's Hospital (Quinghai, China)
from January 2017 to March 2018. The use of human samples was
approved by the Ethical Committee of the Qinghai Provincial
People's Hospital and informed consent was obtained from each
patient.
Animals
A total of 10 male C57BL6J mice (4–6 weeks old;
18–20 g) were obtained for use in the current study (Charles River
Laboratories, Inc.). All mice were housed at 25±5°C with 40–50%
humidity and a 12 h dark/light cycle, and access to food and water
ad libitum. All animal experiments were performed following
the guidelines provided by the National Institutes of Health for
the Care and Use of Laboratory Animals. The current study was
approved by the Ethics Committee of Qinghai Provincial People's
Hospital (Quinghai, China).
Mouse (m)/human (h) bone
marrow-derived mesenchymal stem cells (BMMSCs) isolation and cell
culture
Primary mBMMSCs were isolated from C57BL/6 mice and
hBMMSCs were isolated from human bone marrow as previously
described (30). BMMSCs were
cultured in DMEM supplemented with 10% fetal bovine serum (FBS) and
antibiotics [100 units/ml penicillin G and 100 µg/ml streptomycin
(all, Gibco; Thermo Fisher Scientific, Inc.)]. Cells from passages
3–5 were used in the subsequent experiments.
The mouse preosteoblast cell line, MC3T3-E1, was
obtained from the American Type Culture Collection (cat. no.
CRL-2594). Cells were cultured in α-minimum essential medium
(α-MEM; Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS
and 1% penicillin-streptomycin mix. Cells were then incubated at
37°C with 5% CO2.
Osteogenic differentiation
To induce osteogenic differentiation, mBMMSC, hBMMSC
and MC3T3-E1 cells (4×104 cells) were seeded into
six-well plates. When 80% confluence was achieved, cells were
incubated in osteogenic-inducing medium [10% FBS; 5 mM
L-glycerophosphate; 100 nM dexamethasone (Sigma-Aldrich; Merck
KGaA) and 50 mg/ml ascorbic acid (Sigma-Aldrich; Merck KGaA)] at
37°C for 14 days. Control group cells were cultured in α-MEM
supplemented with 10% FBS at 37°C. To assess the osteoblast
phenotype, osteoblast marker genes were examined by RT-qPCR and
alkaline phosphatase (ALP) activity was determined.
Cell transfection
MC3T3-E1 cells were seeded into six-well plates at a
density of 1×106 cells per well and incubated at 37°C
for 24 h. Cells were then transfected with 100 nM miR-98-5p mimic
(sense 5′-UGAGGUAGUAAGUUGUAUUGUU-3′, antisense
5′-CAAUACAACUUACUACCUCAUU-3′), 100 nM mimic control (sense
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′; both Guangzhou RiboBio Co., Ltd.), 2
µg Control CRISPR activation plasmid (control-plasmid; cat. no.
sc-437275), 2 µg HMGI-C CRISPR Activation Plasmid (HMGA2-plasmid;
cat. no. sc-420880-ACT; Santa Cruz Biotechnology, Inc.) or 100 nM
miR-98-5p mimic with 2 µg HMGA2-plasmid using Lipofectamine
2000® (cat. no. 11668-027; Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Control
cells received no treatment. 48 h after treatment, transfection
efficiency was assessed using RT-qPCR.
ALP activity measurement
To detect extracellular ALP activity, the
spectrophotometric method (31) was
performed with an Alkaline Phosphatase Assay kit (cat. no. P0321;
Beyotime Institute of Biotechnology) was used according to the
manufacturer's protocol. Absorbance was detected at a wavelength of
at 405 nm using a microplate reader (BD Biosciences) following the
manufacturer's protocol.
Cell viability detection assay
Transfected MC3T3-E1 cells were seeded into 96-well
plates (5×103 cells per well). After transfection for 48
h with miR-98-5p mimics, mimic controls, or miR-98-5p mimics in
combination with the HMGA2-plasmid, 100 µl α-MEM medium mixed with
10 µl cell counting kit-8 (CCK-8) solution (cat. no. C0041;
Beyotime Institute of Biotechnology) was added to each well. Cells
were subsequently incubated at 37°C for 30 min. Cell viability was
then evaluated by detecting absorbance at 450 nm using a FLUOstar
Omega Microplate Reader (BMG Labtech GmbH). Cells without treatment
were used as controls. The aforementioned experiment was repeated
three times.
Cell apoptosis assay
To assess the effect of miR-98-5p on MC3T3-E1 cell
apoptosis, a FITC/propidium iodide (PI) apoptosis detection kit
(cat. no. 70-AP101-100; MultiSciences Biotech Co., Ltd.) was
utilized. Subsequent to 48 h transfection with miR-98-5p mimics,
mimic controls, or miR-98-5p mimics in combination with the
HMGA2-plasmid, MC3T3-E1 cells were digested with 0.25% Trypsin,
washed with PBS and subsequently stained with 5 µl Annexin V-FITC
and 5 µl PI for 30 min at room temperature. Finally, to analyze
cell apoptosis, the FACSCanto II flow cytometer (BD Biosciences)
was used according to the manufacturer's protocol, and data was
analyzed by Cell Quest software (version 5.1; BD Biosciences).
Cells without treatment were used as the control.
Western blot analysis
Proteins were extracted from MC3T3-E1 cells using
RIPA buffer (cat. no. P0013E; Beyotime Institute of Biotechnology)
following the manufacturer's protocol. A Bicinchoninic Acid Assay
kit (cat. no. BCA1-1KT; Sigma-Aldrich; Merck KGaA) was used to
quantify protein samples. Equal quantities of protein (30 µg per
lane) were separated on 10% SDS-PAGE and transferred onto PVDF
membranes. Membranes were then blocked with 5% non-fat milk at room
temperature for 1 h and incubated with the following primary
antibodies (all 1:1,000) overnight at 4°C: HMGA2 (cat. no. 8179;
Cell Signaling Technology, Inc.), ALP (cat. no. sc-365765; Santa
Cruz Biotechnology), runt related transcription factor 2 (Runx2;
cat. no. 12556; Cell Signaling Technology, Inc.), transcription
factor sp7 (Osterix; cat. no. sc-393060; Santa Cruz Biotechnology,
Inc.) and β-actin (cat. no. 4970; Cell Signaling Technology, Inc.).
Samples were then incubated with a horseradish
peroxidase-conjugated anti-rabbit immunoglobulin G secondary
antibody (cat. no. 7074; 1:1,000; Cell Signaling Technology Inc.)
at room temperature for 2 h. Corresponding protein bands were
subsequently visualized using Western Blotting Luminol reagent
(cat. no. sc-2048; Santa Cruz Biotechnology, Inc.) following the
manufacturers' protocol.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. Total RNA was then reverse transcribed
into cDNA using the TaqMan MicroRNA Reverse Transcription kit (cat.
no. 4366596; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Synthesized cDNA was subsequently analyzed
via qPCR using a SYBR Premix Ex TaqTM II (TliRNaseH Plus) kit (cat.
no. RR820a; Takara Bio, Inc.). Primer sequences were as following:
miR-98-5p forward, 5′-ACACTCCCUAUACAACUUAC-3′ and reverse,
5′-GGGAAAGUAGUGAGGCCTCAGA-3; U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3; GAPDH forward,
5′-CTTTGGTATCGTGGAAGGACTC-3′ and reverse,
5′-GTAGAGGCAGGGATGATGTTCT-3; RUNX2 forward,
5′-GATGATGACACTGCCACCTCT-3 and reverse, 5′-AGGGCCCAGTTCTGAAGC-3;
ALP forward, 5′-CCGAATTCATGTTGGCCTGTTCAACT-3′ and reverse,
5′-ATGTCGACTTAGTTATTTTCATAATACCAAATTCC-3; Osterix forward,
5′-AGAGATCTGAGCTGGGTAG-3; and reverse, 5′-AAGAGAGCCTGGCAAGAGG-3;
HMGA2 forward, 5′-TCCCTCTAAAGCAGCTCAAAA-3′ and reverse,
5′-ACTTGTTGTGGCCATTTCCT-3. The following thermocycling conditions
were used for the qPCR: Initial denaturation at 95°C for 10 min,
and 35 cycles of 95°C for 15 sec and 55°C for 40 sec. U6 or GAPDH
was used as an internal control for miRNA and mRNA expression,
respectively. The 2−ΔΔCq method was used to calculate
relative gene expression (32).
Dual luciferase reporter assay
miR-98-5p targets were predicted using TargetScan
bioinformatics software (www.targetscan.org/vert_71) and binding sites between
HMGA2 and miR-98-5p were observed. The wild type (WT-HMGA2) and
mutant (MUT-HMGA2) 3′-UTR of HMGA2 were cloned into a
pmiR-RB-ReportTM dual luciferase reporter gene plasmid vector
(Guangzhou RiboBio Co., Ltd.). MC3T3-E1 cells were co-transfected
with WT-HMGA2, MUT-HMGA2 and miR-98-5p mimics or mimic control
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Subsequent to 48 h of cell transfection, luciferase activity was
measured using the dual-luciferase assay system (Promega
Corporation) and normalized to Renilla luciferase
activity.
Statistical analysis
All experiments were performed three times. SPSS
software version 17.0 was used to analyze data (SPSS, Inc.).
Experimental data were expressed as the mean ± standard deviation.
Differences between multiple groups were identified using one-way
ANOVA followed by a Tukey's post-hoc. A Student's t-test was used
to compare the difference between two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Significant downregulation of
miR-98-5p during osteogenic differentiation
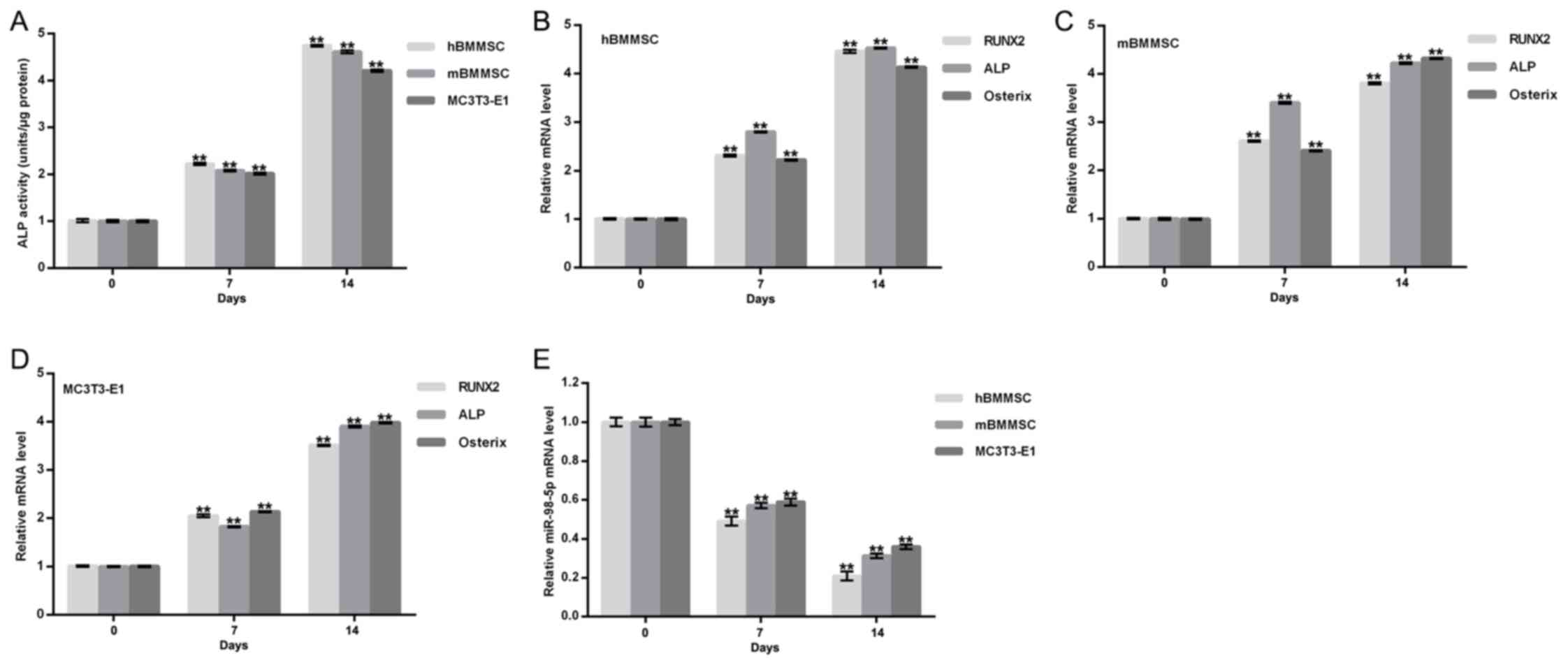
To assess whether miR-98-5p served a role in bone
formation, miR-98-5p expression during osteogenic differentiation
was detected. Three osteoblast cell models (primary hBMMSC, primary
mBMMSC and MC3T3-E1 cells) were used to achieve this. The
osteoblast phenotype was confirmed by increased extracellular ALP
activity (Fig. 1A) and by the
enhanced expression of osteoblast makers including ALP, Runx2 and
Osterix (Fig. 1B-D). Compared with
day 0, levels of miR-98-5p in hBMMSC, mBMMSC and MC3T3-E1 cells
were all significantly reduced at day 7 and day 14 after osteogenic
differentiation incubation (Fig.
1E). The results indicate that miR-98-5p may therefore be
involved in bone formation.
HMGA2 is a target gene of
miR-98-5p
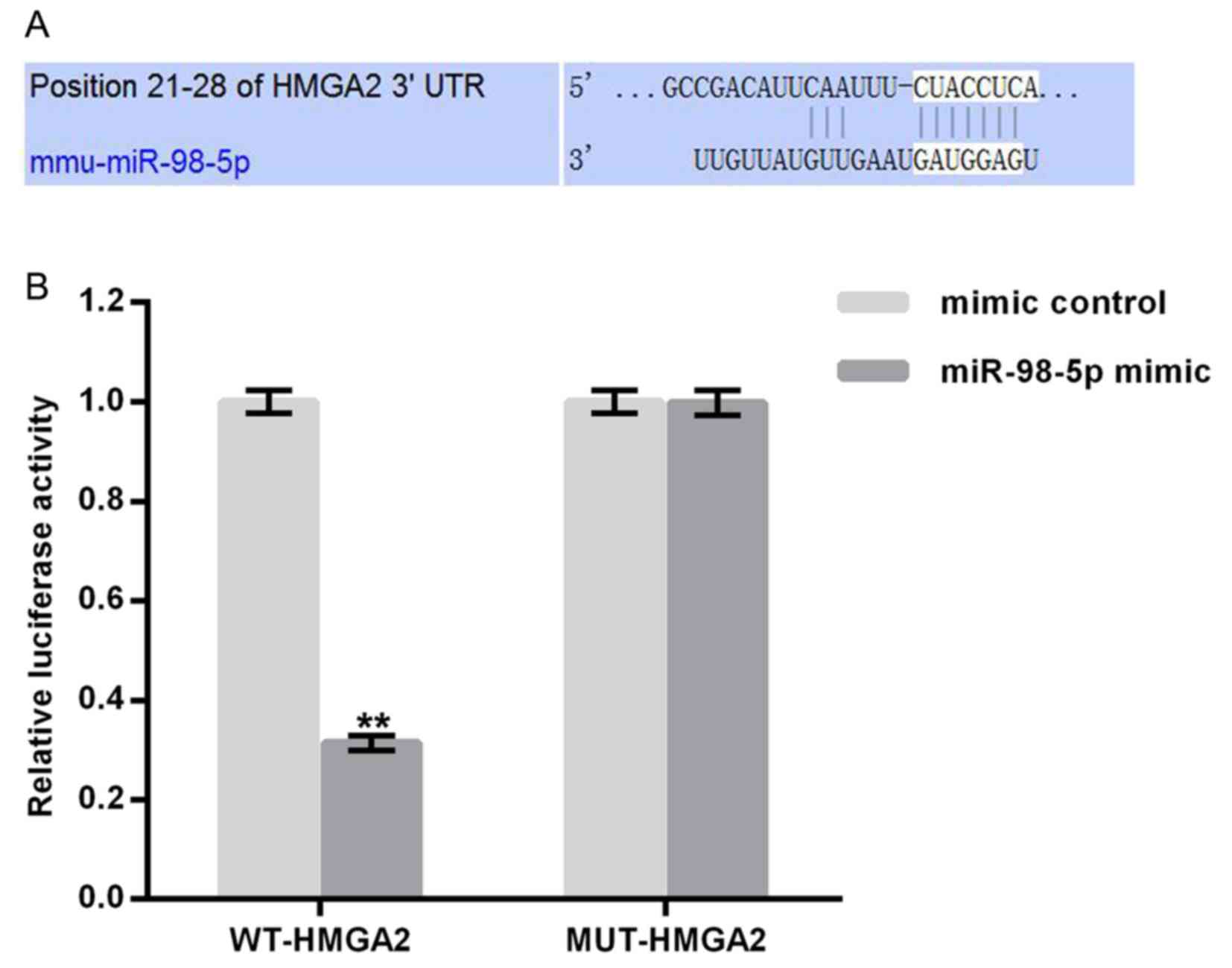
TargetScan (http://www.targetscan.org) was utilized to predict
potential targets of miR-98-5p. Binding sites between HMGA2 and
miR-98-5p were revealed (Fig. 2A).
To confirm these, potential binding sites a luciferase reporter
assay was performed. As presented in Fig. 2B, compared with MUT-HMGA2 and
miR-98-5p mimic transfected cells, luciferase activity was
significantly reduced following WT-HMGA2 and miR-98-5p mimic
transfection. The results demonstrate that miR-98-5p may directly
target HMGA2.
miR-98-5p expression prevents the
osteogenic differentiation of MC3T3-E1 cells
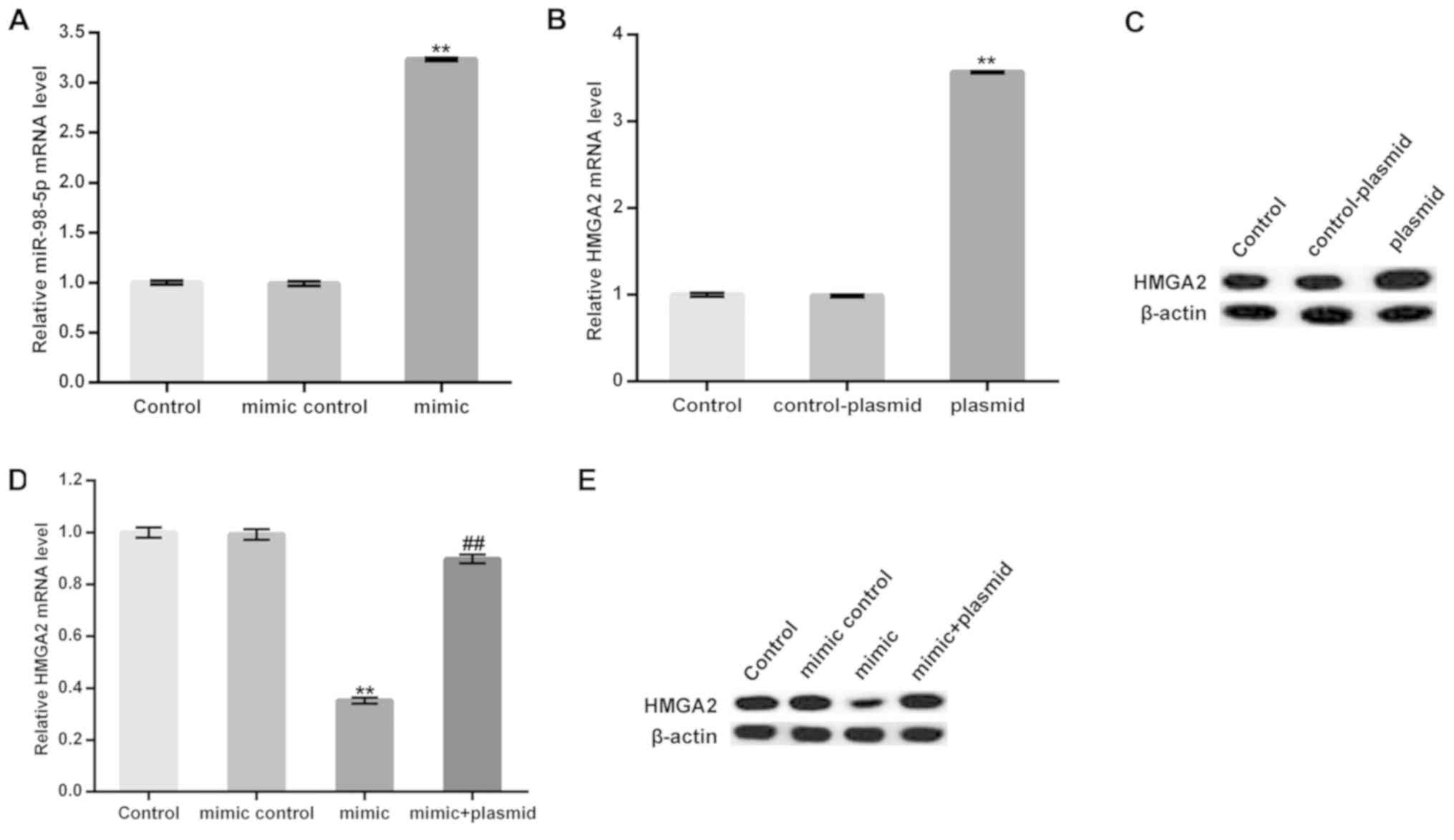
MC3T3-E1 cells were transfected with a miR-98-5p
mimic, a mimic control, a control-plasmid, a HMGA2-plasmid, or a
miR-98-5p mimic in combination with a HMGA2-plasmid for 48 h and
subsequently subjected to osteogenic differentiation induction.
Transfection efficiency was measured using RT-qPCR. As presented in
Fig. 3A-C, the miR-98-5p mimic
significantly upregulated miR-98-5p in MC3T3-E1 cells. Furthermore,
protein and mRNA levels of HMGA2 in MC3T3-E1 cells were enhanced
following treatment with the HMGA2-plasmid. The data demonstrated
that the miR-98-5p mimic significantly reduced the mRNA level of
HMGA2. The protein level of HMGA2 was markedly decreased by
miR-98-5p mimic. However, these decreases were reversed by
HMGA2-plasmid co-transfection (Fig. 3D
and E).
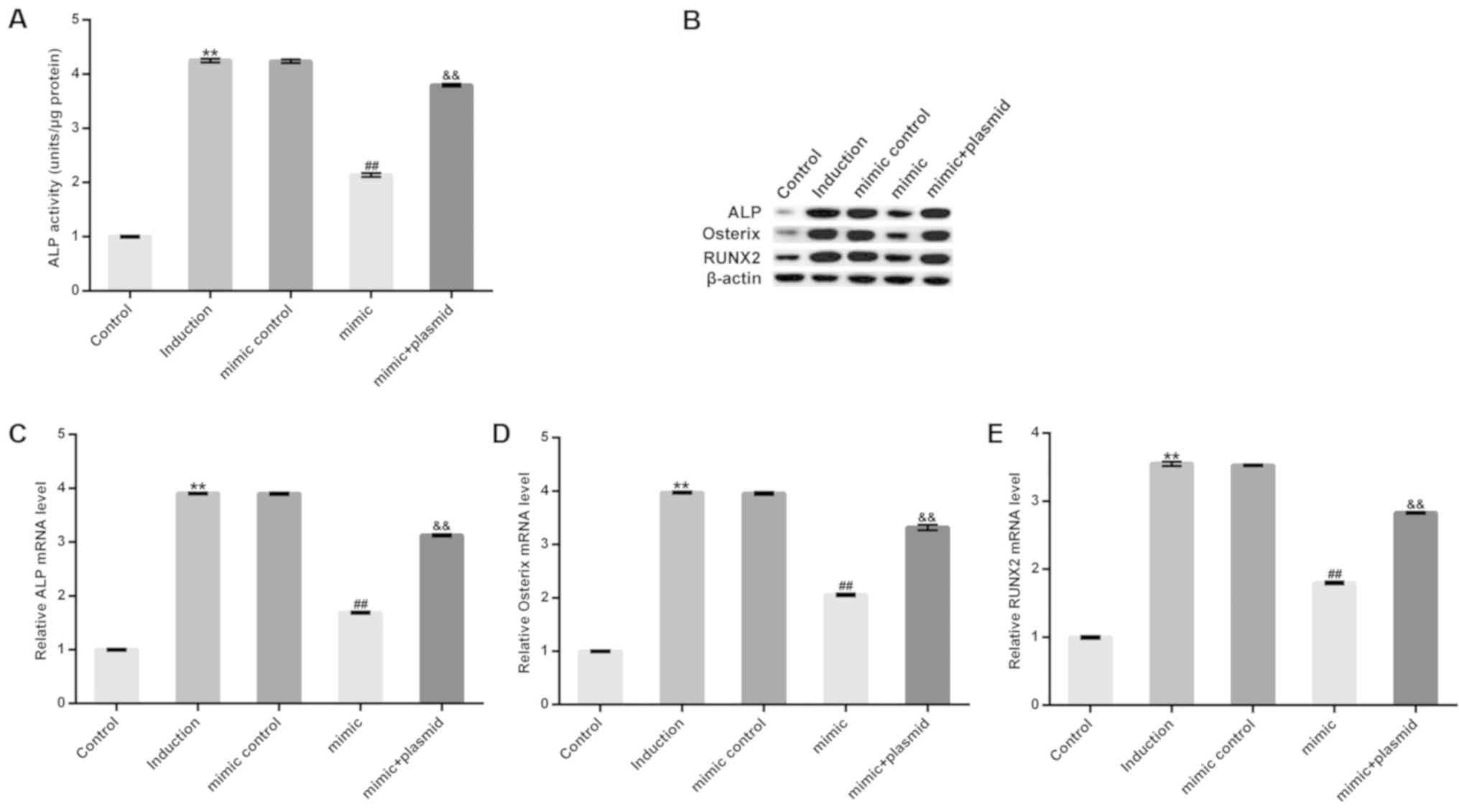
On day 14 after induction, extracellular ALP
activity increased and was subsequently inhibited by treatment with
the miR-98-5p mimic. However, this inhibition was eliminated
following HMGA2-plasmid treatment (Fig.
4A). Furthermore, treatment with the miR-98-5p mimic markedly
decreased the protein levels, and significantly decreased the mRNA
levels of osteoblast makers including ALP, Runx2 and Osterix.
However, these decreases were reversed following HMGA2-plasmid
transfection (Fig. 4B-E). The
results demonstrate that miR-98-5p may prevent osteogenic
differentiation.
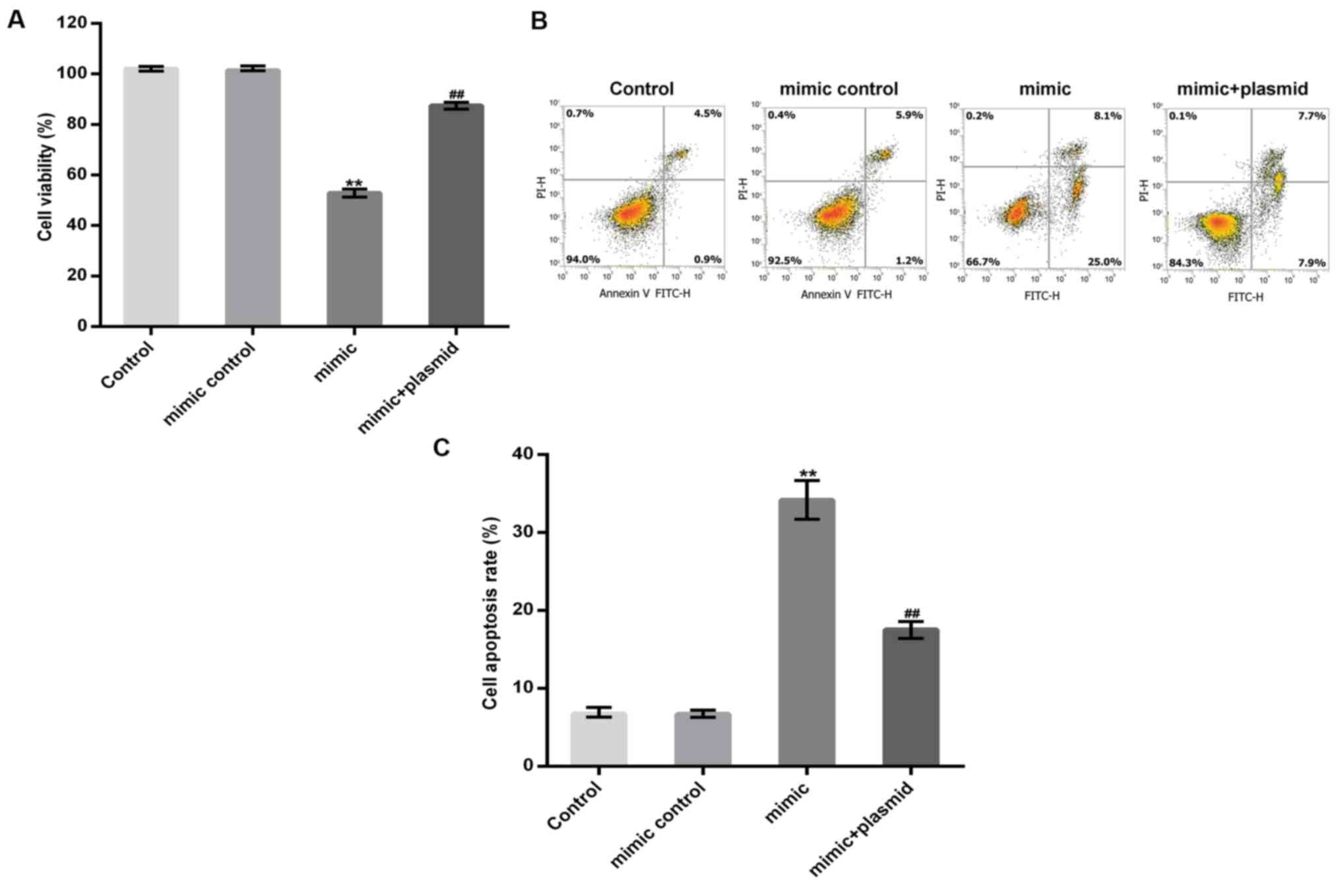
miR-98-5p overexpression inhibits
osteoblast viability and induces apoptosis
MC3T3-E1 cells were transfected with a miR-98-5p
mimic, a mimic control, or a miR-98-5p mimic in combination with a
HMGA2-plasmid for 48 h. A CCK-8 assay and flow cytometry were
subsequently performed to detect MC3T3-E1 cell viability and
apoptosis, respectively. The results revealed that the miR-98-5p
mimic significantly reduced MC3T3-E1 cell viability and induced
MC3T3-E1 cell apoptosis and these changes were reversed by HMGA2
upregulation (Fig. 5A-C).
Discussion
The results of the present study demonstrated that
miR-98-5p was significantly downregulated during osteogenic
differentiation in vitro and that HMGA2 may be a direct
target of miR-98-5p. miR-98-5p upregulation inhibited ALP activity
and the expression of osteoblast markers, including ALP, Runx2 and
Osterix. This inhibition may have been due to miR-98-5p targeting
HMGA2, demonstrating the inhibitory effect of miR-98-5p on
osteogenic differentiation. In addition, the results of the current
study demonstrated that miR-98-5p upregulation inhibited cell
viability and induced cell apoptosis in the mouse preosteoblast
cell line, MC3T3-E1. miR-98-5p may have also inhibited bone
regeneration by targeting HMGA2 and therefore may be an encouraging
therapeutic target for the enhancement of bone regeneration.
New bone regeneration is essential in many common
bone diseases, including trauma and osteoporosis (12). Although previous studies have
identified a variety of therapies to treat these diseases, the
efficacy of these treatments remains unsatisfactory (13). Evidence has revealed that miRNAs
serve a regulatory role in cell proliferation, differentiation and
apoptosis (2–5). Increasing evidence has also revealed
that miRNAs are involved in the regulation of osteoblasts
differentiation, bone metabolism and bone formation (14–19).
miR-467g has been demonstrated to prevent new bone regeneration by
regulating the indian hedgehog protein/Runx2 signaling pathway
(33). miR-221 was revealed to
suppress osteoblast differentiation and bone formation by
regulating Runx2 expression (34).
miR-214 has also been identified to inhibit osteogenesis by
targeting baculoviral IAP repeat-containing protein 7 (35). Therefore, miRNAs may serve as novel
therapeutic targets for the treatment of bone degenerative
disorders and other abnormal bone formation disorders.
In the present study, three osteoblast cell models
(primary hBMMSC, primary mBMMSC and MC3T3-E1 cells) were used to
investigate the role of miR-98-5p in bone regeneration. miR-98-5p
significantly decreased during osteogenic differentiation in
vitro. Furthermore, HMGA2 was revealed to be a direct target of
miR-98-5p. HMGA2 is a nuclear-binding protein that serves an
important role in the regulation of cell proliferation and
differentiation (26). A previous
study has demonstrated that HMGA2 knockdown results in the
inhibition of MSC proliferation (27). The effect of miR-98-5p on the
osteogenic differentiation of the mouse preosteoblast cell line,
MC3T3-E1, was subsequently investigated in the current study.
Consistent with previous studies (27,29), the
results of the present study demonstrated that miR-98-5p
significantly inhibited MC3T3-E1 cell osteogenic differentiation by
repressing the expression of HMGA2. In addition, miR-98-5p
upregulation significantly inhibited cell viability and induced
apoptosis in MC3T3-E1 cells and these effects were eliminated by
HMGA2 overexpression.
In summary, the results obtained in the present
study indicated that miR-98-5p was downregulated during osteogenic
differentiation in vitro and its overexpression could
inhibit osteogenic differentiation and suppress the growth of
osteoblast cells by targeting HMGA2. Therefore, miR-98-5p may be a
promising therapeutic target for the enhancement of new bone
regeneration.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FZ was responsible for experimental design and
manuscript preparation. FW performed most of the experiments and ZX
analyzed the data.
Ethics approval and consent to
participate
The use of human samples was approved by the Ethical
Committee of the Qinghai Provincial People's Hospital (Quinghai,
China) and informed consent was obtained from each patient.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hammond SM: An overview of microRNAs. Adv
Drug Deliv Rev. 87:3–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Soifer HS, Rossi JJ and Saetrom P:
MicroRNAs in disease and potential therapeutic applications. Mol
Ther. 15:2070–2079. 2017. View Article : Google Scholar
|
|
4
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
O'Connell RM, Rao DS, Chaudhuri AA and
Baltimore D: Physiological and pathological roles for microRNAs in
the immune system. Nat Rev Immunol. 10:111–122. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tutar Y: miRNA and cancer; computational
and experimental approaches. Curr Pharm Biotechnol. 15:4292014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhong Z, Hou J, Zhang Q, Zhong W, Li B, Li
C, Liu Z, Yang M and Zhao P: Circulating microRNA expression
profiling and bioinformatics analysis of dysregulated microRNAs of
patients with coronary artery disease. Medicine (Baltimore).
97:e114282018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang S, Zhang R, Wu F and Li X:
MicroRNA-208a regulates H9c2 cells simulated ischemia-reperfusion
myocardial injury via targeting CHD9 through Notch/NF-kappa B
signal pathways. Int Heart J. 59:580–588. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Price NL, Ramírez CM and
Fernández-Hernando C: Relevance of microRNA in metabolic diseases.
Crit Rev Clin Lab Sci. 51:305–320. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qiu L, Tan EK and Zeng L: microRNAs and
neurodegenerative diseases. Adv Exp Med Biol. 888:85–105. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rossi M, Tagliaferri P and Tassone P:
MicroRNAs in multiple myeloma and related bone disease. Ann Transl
Med. 3:3342015.PubMed/NCBI
|
|
12
|
Majidinia M, Sadeghpour A and Yousefi B:
The roles of signaling pathways in bone repair and regeneration. J
Cell Physiol. 233:2937–2948. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hu C, Zhang T, Ren B, Deng Z, Cai L, Lei J
and Ping A: Effect of vacuum-assisted closure combined with open
bone grafting to promote rabbit bone graft vascularization. Med Sci
Monit. 21:1200–1206. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lian JB, Stein GS, van Wijnen AJ, Stein
JL, Hassan MQ, Gaur T and Zhang Y: MicroRNA control of bone
formation and homeostasis. Nat Rev Endocrinol. 8:212–227. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Valenti MT, Dalle Carbonare L and Mottes
M: Role of microRNAs in progenitor cell commitment and osteogenic
differentiation in health and disease (Review). Int J Mol Med.
41:2441–2449. 2018.PubMed/NCBI
|
|
16
|
Tang J, Zhang Z, Jin X and Shi H: miR-383
negatively regulates osteoblastic differentiation of bone marrow
mesenchymal stem cells in rats by targeting Satb2. Bone.
114:137–143. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Arfat Y, Basra MAR, Shahzad M, Majeed K,
Mahmood N and Munir H: miR-208a-3p suppresses osteoblast
differentiation and inhibits bone formation by targeting ACVR1. Mol
Ther Nucleic Acids. 11:323–336. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang L, Tang Y, Zhu X, Tu T, Sui L, Han
Q, Yu L, Meng S, Zheng L, Valverde P, et al: Overexpression of
MiR-335-5p promotes bone formation and regeneration in mice. J Bone
Miner Res. 32:2466–2475. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang M, Qing Y, Shi Q, Cao Y and Song K:
miR-342-3p elevates osteogenic differentiation of umbilical cord
mesenchymal stem cells via inhibiting Sufu in vitro. Biochem
Biophys Res Commun. 491:571–577. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Bao W, Liu Y, Wang S, Xu S, Li X,
Li Y and Wu S: miR-98-5p contributes to cisplatin resistance in
epithelial ovarian cancer by suppressing miR-152 biogenesis via
targeting Dicer1. Cell Death Dis. 9:4472018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fu Y, Liu X, Chen Q, Liu T, Lu C, Yu J,
Miao Y and Wei J: Downregulated miR-98-5p promotes PDAC
proliferation and metastasis by reversely regulating MAP4K4. J Exp
Clin Cancer Res. 37:1302018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu F, Mo Q, Wan X, Dan J and Hu H:
NEAT1/hsa-mir-98- 5p/MAPK6 axis is involved in non-small-cell lung
cancer development. J Cell Biochem. 120:2836–2846. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang T, Li M, Li Q, Guo Z, Sun X, Zhang
X, Liu Y, Yao W and Xiao P: MicroRNA-98-5p inhibits cell
proliferation and induces cell apoptosis in hepatocellular
carcinoma via targeting IGF2BP1. Oncol Res. 25:1117–1127. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li Q, Li X, Wang L, Zhang Y and Chen L:
miR-98-5p acts as a target for Alzheimer's disease by regulating Aβ
production through modulating SNX6 expression. J Mol Neurosci.
60:413–420. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sahu M and Mallick B: Deciphering
synergistic regulatory networks of microRNAs in hESCs and
fibroblasts. Int J Biol Macromol. 113:1279–1286. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Park SM, Shell S, Radjabi AR, Schickel R,
Feig C, Boyerinas B, Dinulescu DM, Lengyel E and Peter ME: Let-7
prevents early cancer progression by suppressing expression of the
embryonic gene HMGA2. Cell Cycle. 6:2585–2590. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kalomoiris S, Cicchetto AC, Lakatos K,
Nolta JA and Fierro FA: Fibroblast growth factor 2 regulates high
mobility group A2 expression in human bone marrow-derived
mesenchymal stem cells. J Cell Biochem. 117:2128–2137. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei J, Li H, Wang S, Li T, Fan J, Liang X,
Li J, Han Q, Zhu L, Fan L and Zhao RC: let-7 enhances osteogenesis
and bone formation while repressing adipogenesis of human
stromal/mesenchymal stem cells by regulating HMGA2. Stem Cells Dev.
23:1452–1463. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tian Z, Zhou H, Xu Y and Bai J:
MicroRNA-495 inhibits new bone regeneration via targeting high
mobility group AT-Hook 2 (HMGA2). Med Sci Monit. 23:4689–4698.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Matsubara T, Suardita K, Ishii M, Sugiyama
M, Igarashi A, Oda R, Nishimura M, Saito M, Nakagawa K, Yamanaka K,
et al: Alveolar bone marrow as a cell source for regenerative
medicine: Differences between alveolar and iliac bone marrow
stromal cells. J Bone Miner Res. 20:399–409. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ni P, Xie J, Chen C, Jiang Y, Zhao Z,
Zhang Y, Lu Y and Yu J: Spectrophotometric determination of the
activity of alkaline phosphatase and detection of its inhibitors by
exploiting the pyrophosphate-accelerated oxidase-like activity of
nanoceria. Mikrochim Acta. 186:3202019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kureel J, John AA, Dixit M and Singh D:
MicroRNA-467g inhibits new bone regeneration by targeting
Ihh/Runx-2 signaling. Int J Biochem Cell Biol. 85:35–43. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang Y, Gao Y, Cai L, Li F, Lou Y, Xu N,
Kang Y and Yang H: MicroRNA-221 is involved in the regulation of
osteoporosis through regulates RUNX2 protein expression and
osteoblast differentiation. Am J Transl Res. 9:126–135.
2017.PubMed/NCBI
|
|
35
|
Liu J, Li Y, Luo M, Yuan Z and Liu J:
MicroRNA-214 inhibits the osteogenic differentiation of human
osteoblasts through the direct regulation of baculoviral IAP
repeat-containing 7. Exp Cell Res. 351:157–162. 2017. View Article : Google Scholar : PubMed/NCBI
|