Introduction
Pogostemonis Herba is the dried aerial part of
Pogostemon cablin (Blanco) Benth. (Labiatae),
commonly known as ‘Guang-Huo-Xiang’ in Chinese or Cablin Patchouli
in English. It has been traditionally used in Chinese medicine to
remove dampness, relieve summer heat and exterior syndrome, and as
an antiemetic and appetite stimulant (1). Pogostemonis Herba is a common Chinese
herb frequently used as a component in popular traditional
formulae, such as Baoji Pill and Houdan Pill, for the treatment of
inflammatory diseases (2,3). In addition, patchouli oil (essential
oil of Patchouli) has been widely used in the cosmetic and oral
hygiene industries to scent perfumes and flavor toothpaste. Recent
research has demonstrated that patchouli oil has a variety of
pharmacological activities, including antiemetic (4), anti-inflammatory (3,5),
anti-allergic (6),
immunomodulatory (7) and
antimicrobial actions (8).
Chemically, it has been reported that Pogostemon cablin
contains sesquiterpenes (9),
cytotoxic chalcones (10) and
antimutagenic flavones (11).
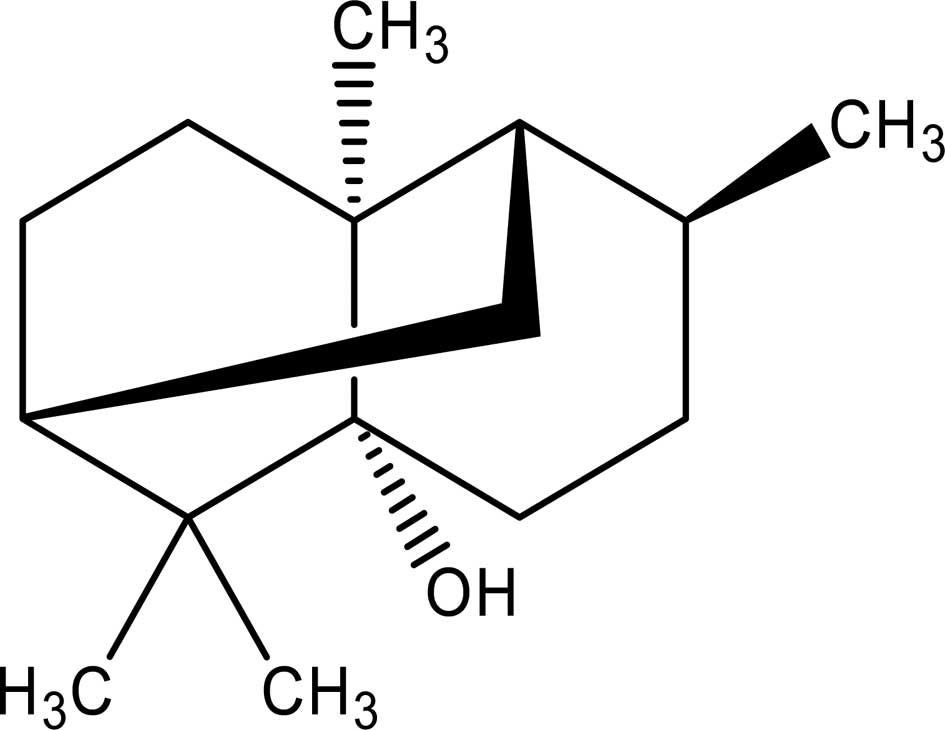
Patchouli alcohol (PA; chemical structure in Fig. 1), a tricyclic sesquiterpene, is a
major active ingredient of Pogostemonis Herba and is the most
odorintensive component of patchouli oil (12). Pogostemonis Herba has been reported
to contain 0.023–0.039% PA (12).
In recent years, PA has attracted significant research attention
due to its potential as a cognitive enhancing, learning impairment
attenuating and neuroprotective agent (13,14).
However, it remains to be determined whether PA is the active
constituent responsible for the anti-inflammatory effect of
Pogostemonis Herba.
Macrophages are the first line of host defense
against bacterial infection and cancer growth, and are believed to
play essential roles in the initiation, maintenance and resolution
of inflammation. When stimulated by bacterial endotoxin, such as
lipopolysaccharide (LPS), macrophages produce a number of
cytokines, such as tumor necrosis factor (TNF)-α, interleukin
(IL)-1β and IL-6; chemokines, such as prostaglandin E2
(PGE2) and nitric oxide (NO); inflammation-related
enzymes, such as cyclooxygenase (COX)-2 and inducible nitric oxide
synthase (iNOS) for the primary protection of the host (15,16).
These cytokines and chemokines are essential for the inflammatory
response to pathogenic germs or toxicants (17). However, overproduction of these
inflammatory mediators is associated with numerous diseases, such
as rheumatoid arthritis and atherosclerosis (18–21).
Thus, inhibition of the overproduction of these inflammatory
mediators may have a beneficial effect on these inflammatory
diseases.
RAW264.7, a mouse macrophage cell line, has been
widely used as an in vitro inflammatory model (15,22,23).
In the present study, we aimed to investigate whether PA has an
anti-inflammatory effect on LPS-stimulated RAW264.7 cells. We also
examined the molecular mechanisms underlying the anti-inflammatory
effect of PA by evaluating the mRNA expression of a number of
inflammatory mediators, including TNF-α, IL-1β, IL-6, iNOS and
COX-2.
Materials and methods
Plant materials and reagents
The aerial parts of Pogostemon cablin were
collected in June 2009 in Maoming, Guangdong, China. The plant was
authenticated by one of the authors (X.-P.L., experienced in
pharmacognosy) at the College of Chinese Medicines, Guangzhou
University of Chinese Medicine, where a voucher specimen (no.
090612) was deposited.
LPS, RPMI-1640 culture medium, sulfanilamide and
N-1-naphthylethylenediamine dihydrocholide were purchased from
Sigma-Aldrich (St. Louis, MO, USA). Fetal bovine serum (FBS),
penicillin and streptomycin were purchased from Gibco (Grand
Island, NY, USA). All other reagents and chemicals used in the
study were of analytical grade.
Extraction and isolation of PA
The dried aerial parts of Pogostemon cablin
(5 kg) were refluxed with 95% aqueous EtOH (40 liters × 2, 60 min
each). The extract was evaporated under reduced pressure to obtain
a residue (100 g). The residue was dissolved in acetone and
subjected to silica gel column chromatography eluted with a
petroleum ether-ethyl acetate-0.1% formic acid (20:1:0.1, 9:1:0.1,
8:3:0.1 and 7:4:0.1) gradient elution system. The fraction eluted
with petroleum ether-ethyl acetate-0.1% formic acid (9:1:0.1) was
combined and further evaporated to dryness, and a yellowish oily
liquid was obtained. After crystallization from cyclohexane, white
crystals of PA (540 mg, yield 0.011%) were finally obtained. The
chemical structure of PA was identified by comparing its spectral
data (MS, 1H- and 13C-NMR) to those published
previously (24). The purity of PA
was found to be >98% based on gas chromatography (GC) analysis.
PA was dissolved in dimethyl sulfoxide (DMSO) and the solvent
concentration was <0.1% DMSO in all experiments.
Cell culture and drug treatment
The RAW264.7 cell line, derived from murine
macrophages, was obtained from the American Type Culture Collection
(ATCC; Rockville, MD, USA). The cells were maintained in RPMI-1640
medium supplemented with 2 mM glutamine, antibiotics (100 U/ml
penicillin and 100 U/ml streptomycin) and 10% heat-inactivated FBS
at 37˚C in a humidified atmosphere of 95% air and 5%
CO2. The cells were seeded onto 96-well culture plates
at 5×104 cells/well, unless otherwise specified. After
seeding for 24 h, the cells were cultured in serum-free medium and
incubated with different concentrations of PA (final
concentrations: 0, 10, 20 and 40 μM) for 2 h. LPS at a final
concentration of 100 ng/ml was then added for an additional 24
h.
Cell viability assay
Cell viability was measured by a CellTiter
96® AQueous One Solution Cell Proliferation assay
(Promega, Madison, WI, USA). In brief, the cells were washed with
D-Hank’s solution after drug treatment. Then, 100 μl of serum-free
medium and 20 μl of CellTiter 96 AQueous One Solution were added to
each well. The cells were further incubated at 37˚C for 1 h. The
quantity of formazan product, which is directly proportional to the
number of living cells, was measured with a FLUOstar Optima
microplate reader (BMG Labtech, Offenbury, Germany) at 490 nm. Cell
viability was expressed as a percentage of the non-treated
control.
Nitrite oxide assay
The nitrite concentration in the culture medium was
measured as an indicator of NO production according to the Griess
reaction method described elsewhere (15). Briefly, the supernatants were
collected at the end of the drug treatment. A total of 100 μl of
each supernatant was mixed with the same volume of Griess reagent
(50 μl 1% sulfanilamide in 5% phosphoric acid and 50 μl 0.1%
N-1-naphthylethylenediamine dihydrocholide in water). After
incubation for 10 min at room temperature, the absorbance was
measured at 540 nm using a microplate reader. The content of
nitrite was expressed as a percentage of the non-treated
control.
PGE2 assay
After PA treatment, the supernatants were collected
and used for the PGE2 assay. The PGE2
concentration in the supernatant was determined using a
commercially available PGE2 EIA kit (Cayman Chemical
Co., Ann Arbor, MI, USA) according to the manufacturer’s
instructions. Briefly, 50 μl of diluted standard/samples was added
to a 96-well plate pre-coated with goat polyclonal anti-mouse IgG
antibody. Aliquots of a PGE2 monoclonal antibody and
PGE2 acetylcholine esterase (AChE) conjugate were added
to each well and allowed to incubate at room temperature for 60 min
on an orbital shaker. After washing five times with wash buffer
containing 0.05% Tween-20, 200 μl of Ellman’s reagent comprising
acetylthiocholine and 5,5′-dithio-bis-(2-nitrobenzoic acid) was
added to the wells. The plates were then incubated for 60 min at
room temperature in the dark, then the absorbance was read at 405
nm using a microplate reader. The content of PGE2 was
expressed as a percentage of the non-treated control.
Cytokine determination
After PA treatment, the supernatants were collected
and used for TNF-α, IL-1β and IL-6 assays. The levels of TNF-α,
IL-1β and IL-6 in the supernatant were measured using commercially
available sandwich enzymelinked immunosorbent assay (ELISA) kits
(Invitrogen Co., Carlsbad, CA, USA) as per the manufacturer’s
instructions. Briefly, samples and biotinylated anti-TNF-α,
anti-IL-1β and anti-IL-6 antibodies (Biotin Conjugate) were
respectively added to the 96-well plates pre-coated with monoclonal
anti-mouse TNF-α, anti-mouse IL-1β and anti-mouse IL-6 antibodies,
and incubated for 90 min at room temperature, 90 min at 37˚C and 2
h at room temperature, respectively. After washing four times, a
streptavidin-HRP working solution was added and incubated for 30
min at room temperature. Then, tetramethylbenzidine (TMB) was added
and incubated for 30 min at room temperature in the dark after
washing. The reaction was stopped with stop solution and the
absorbance was immediately recorded at 450 nm. The levels of TNF-α,
IL-1β and IL-6 were expressed as a percentage of the non-treated
control.
Real-time PCR analysis
The RAW264.7 cells were seeded at 2×106
cells/well in 6-well plates. The cells were washed twice with
D-Hank’s solution after PA treatment. Total RNA was isolated from
the cells with TRIzol reagent (Gibco). The concentration of
extracted RNA was measured spectrophotometrically at 260 nm. The
quality of RNA was assessed by the ratio of absorbance at 260 and
280 nm. The values of A260/A280 from 1.9 to 2.1 were considered
acceptable. Total RNA (1.5 μg) was used to synthesize cDNA using
the High capacity cDNA reverse transcription kit (Applied
Biosystems, Foster City, CA, USA) following the manufacturer’s
instructions. Real-time PCR was performed with a Taq Man
fast universal PCR master mix kit (2X) (Applied Biosystems) and
mouse Taq Man gene expression assays were conducted (Applied
Biosystems; assay ID: TNF-α, Mm00443258_m1; IL-6, Mm99999064_m1;
IL-1β, Mm00434228_m1; COX-2, Mm01307330_g1; iNOS, Mm00440488_m1 and
β-actin, Mn02619580_g1). The reactions were run at 50˚C for 2 min
and 95˚C for 10 min, followed by 60 cycles at 95˚C for 15 sec and
60˚C for 1 min on the Applied Biosystems Step-One Fast Real-Time
PCR system. Sequence Detection Software 2.0 (Applied Biosystems)
was used for data analysis. The relative expression of TNF-α,
IL-1β, IL-6, COX-2 and iNOS mRNA was normalized to the amount of
β-actin in the same cDNA using the relative quantification
2−ΔΔCt method (25). The fold change in target gene cDNA
relative to the β-actin internal control was determined using the
following formula: Fold change = 2−ΔΔCt,
where ΔΔCt = (Cttarget gene − Ctβ-actin) −
(Ctcontrol − Ctβ-actin).
Statistical analysis
Data were expressed as the mean ± SEM. Multiple
group comparisons were performed using one-way analysis of variance
(ANOVA) followed by Dunnett’s test to detect differences between
different treatment groups and the control. Differences were
considered statistically significant at p<0.05.
Results
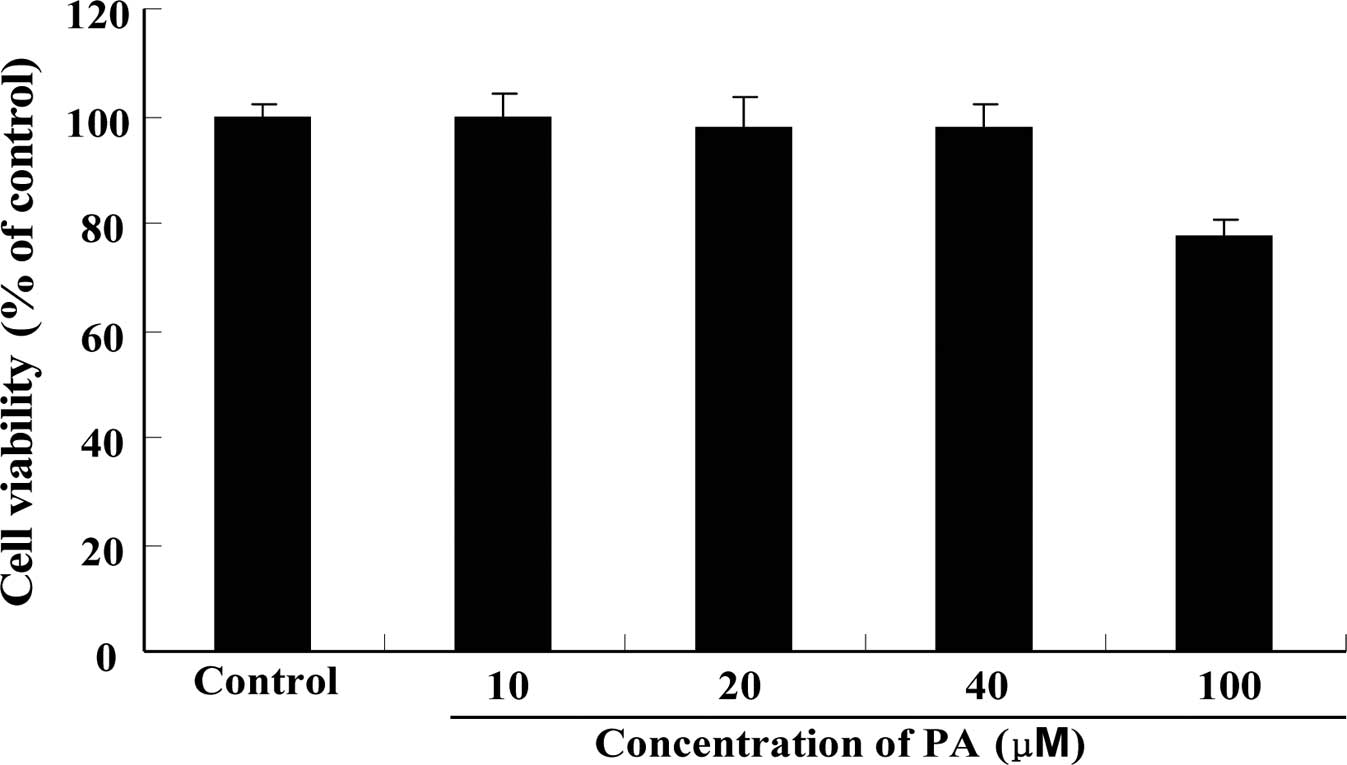
Cytotoxicity of PA on RAW264.7 cells
As shown in Fig. 2,
PA at concentrations up to 40 μM did not show significant
cytotoxicity to RAW264.7 cells when incubated for 24 h.
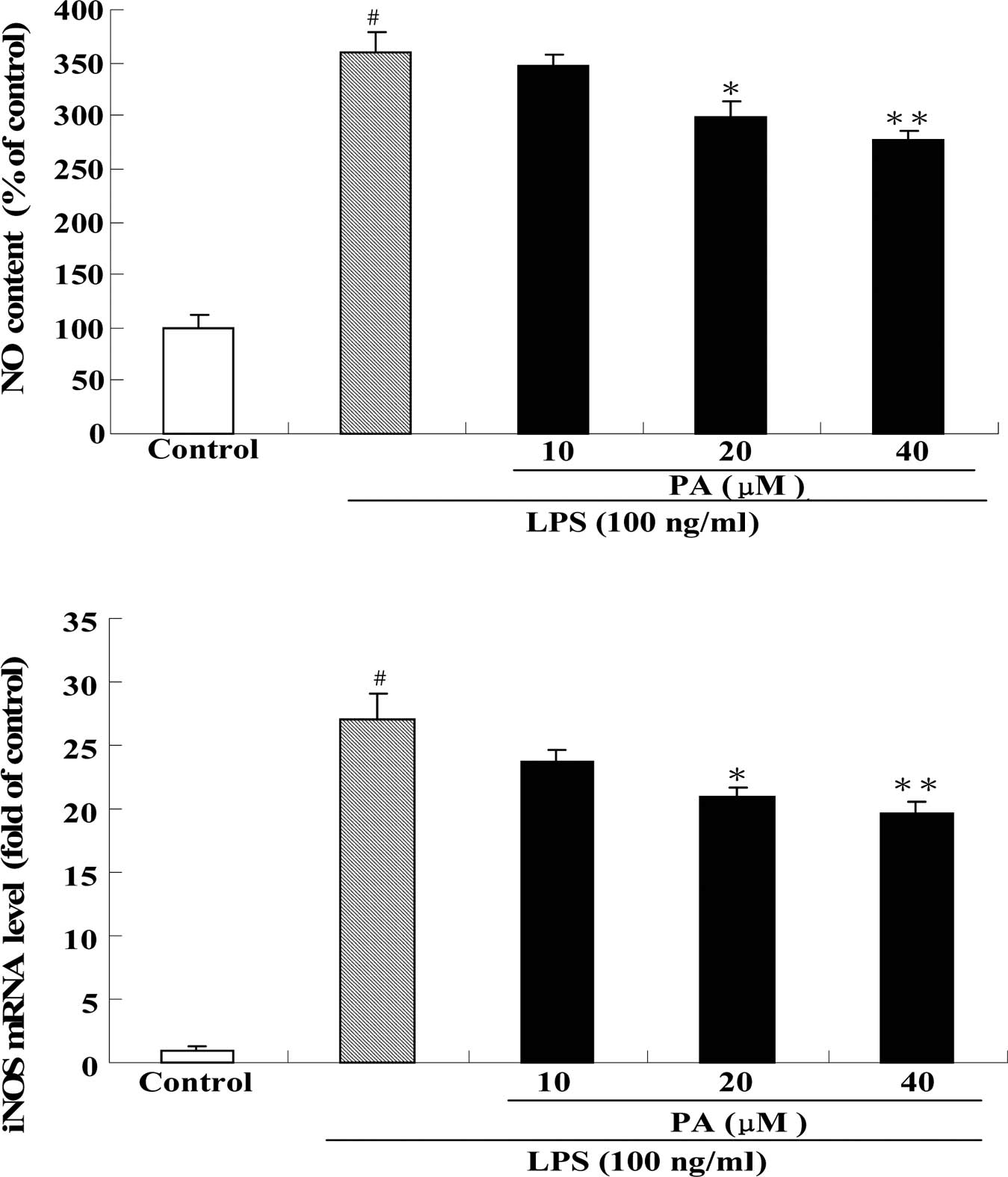
Effect of PA on NO production and iNOS
mRNA expression in LPS-stimulated RAW264.7 cells
Fig. 3 shows the
effect of PA on NO production (A) and iNOS mRNA expression (B) in
LPS-stimulated RAW264.7 cells. After exposure of RAW264.7 cells to
100 ng/ml LPS for 24 h, the production of NO and the mRNA
expression of iNOS were significantly increased by 3.6- and
27-fold, respectively, as compared to the control group.
Pre-treatment with 20 and 40 μM PA significantly decreased the
production of NO by 17 and 23%, respectively, as compared to the
LPS group. Pre-treatment with 20 and 40 μM PA also significantly
suppressed the mRNA expression of iNOS by 23 and 28%, respectively,
as compared to the LPS group.
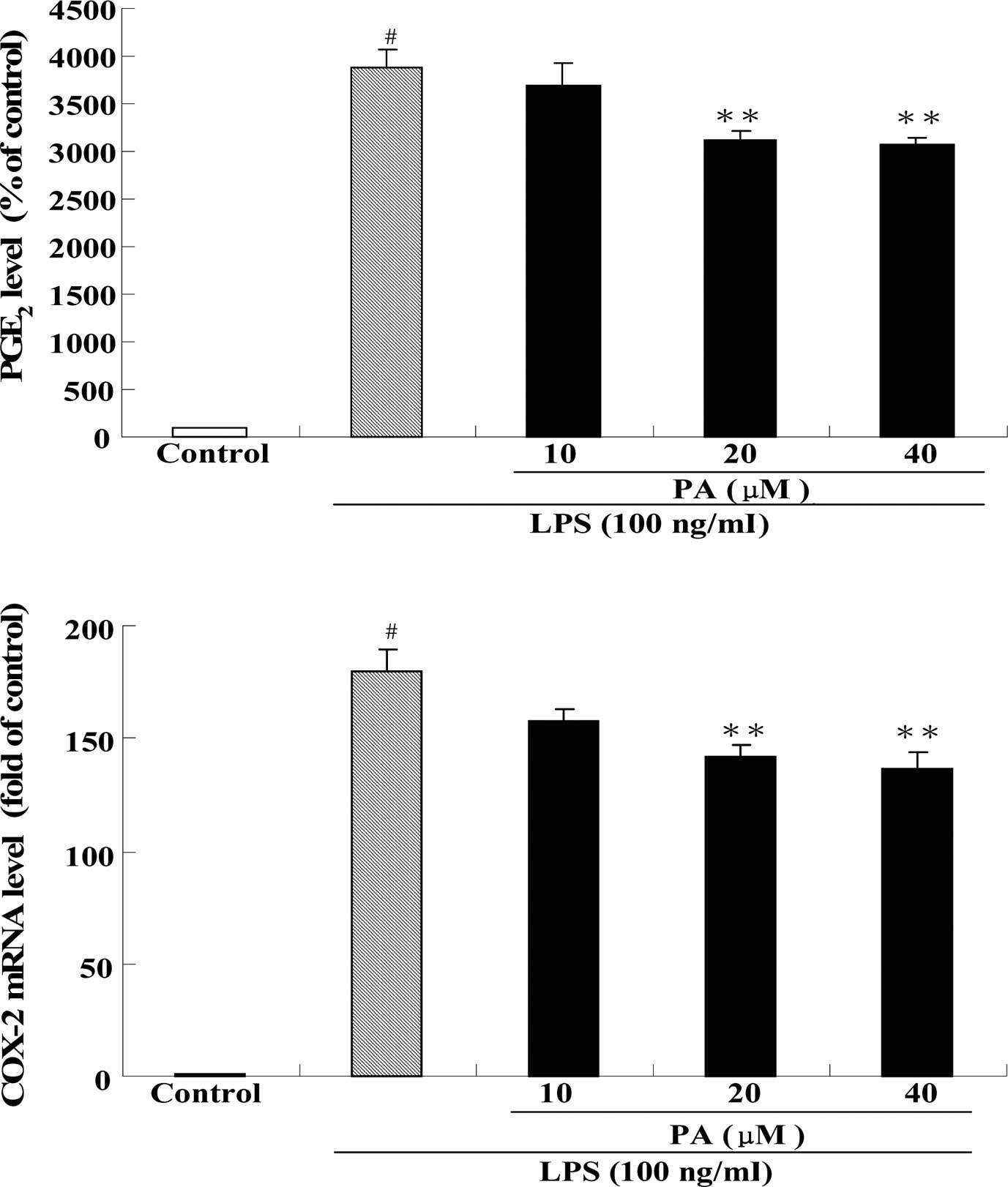
Effect of PA on PGE2
production and COX-2 mRNA expression in LPS-stimulated RAW264.7
cells
Fig. 4 shows the
effect of PA treatment on PGE2 production (A) and COX-2
mRNA expression (B) in LPS-stimulated RAW264.7 cells. After
exposure of RAW264.7 cells to 100 ng/ml LPS for 24 h, the
production of PGE2 and the mRNA expression of COX-2 were
significantly increased by 39- and 180-fold, respectively, as
compared to the control group. Pre-treatment with 20 and 40 μM PA
significantly decreased the production of PGE2 by 19 and
21%, respectively, as compared to the LPS group. In addition,
pre-treatment with 20 and 40 μM PA also significantly decreased the
mRNA expression of COX-2 by 21 and 24%, respectively, as compared
to the LPS group.
Effect of PA on pro-inflammatory cytokine
production and mRNA expression in LPS-stimulated RAW264.7
cells
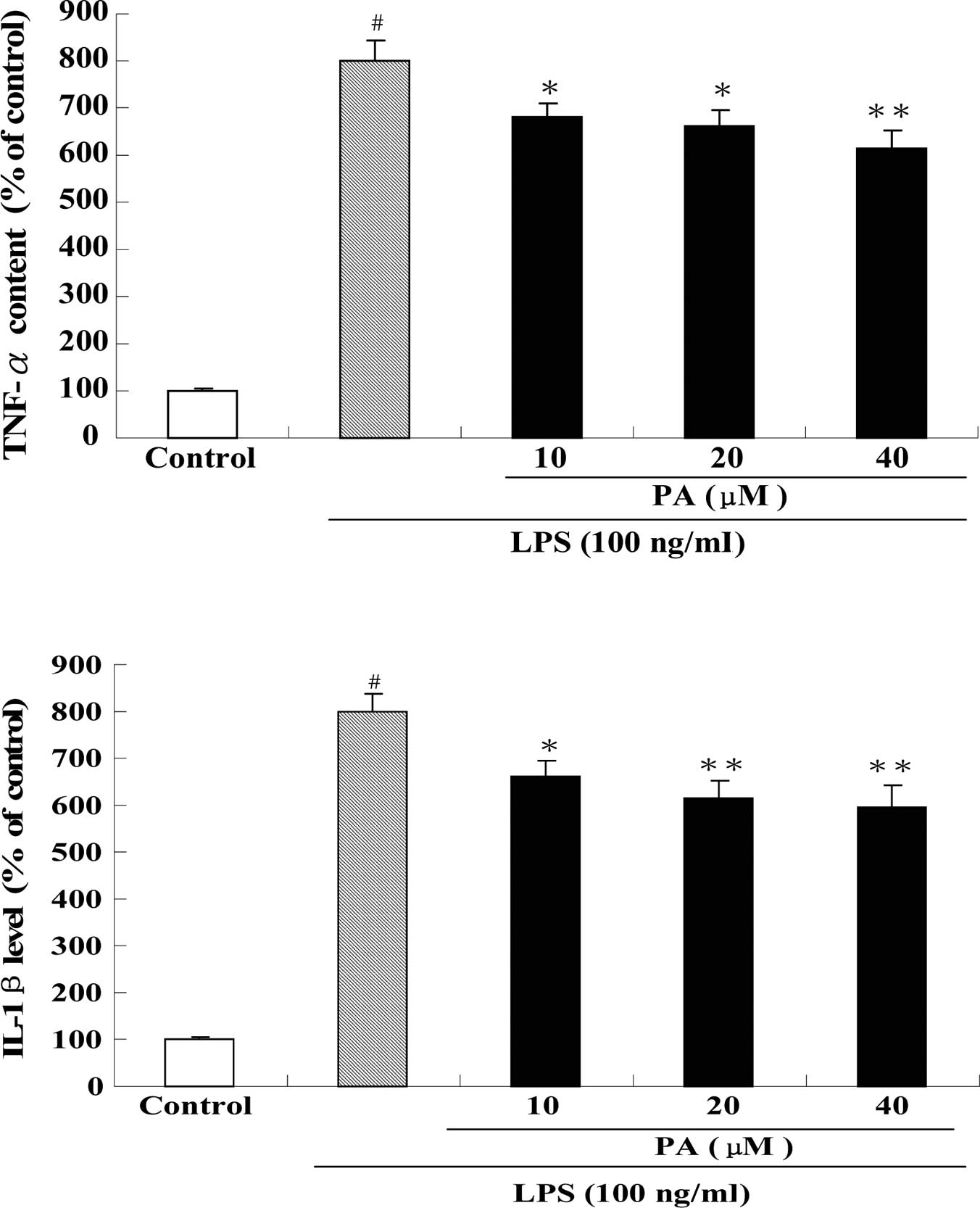
Fig. 5 shows the
effect of PA treatment on the protein levels of TNF-α (A), IL-1β
(B) and IL-6 (C) in LPS-stimulated RAW264.7 cells. After exposure
of RAW264.7 cells to 100 ng/ml LPS for 24 h, the protein levels of
TNF-α, IL-1β and IL-6 were significantly increased by 8-, 8- and
20-fold, respectively, as compared to the control group.
Pre-treatment with 10, 20 and 40 μM PA dose-dependently suppressed
the protein level of TNF-α by 15, 17 and 23%, respectively, as
compared to the LPS group. In a similar fashion, pre-treatment with
10, 20 and 40 μM PA dose-dependently decreased the protein level of
IL-1β by 17, 23 and 25%, respectively, as compared to the LPS
group. In addition, pre-treatment with 20 and 40 μM PA also
significantly reduced the protein level of IL-6 by 17 and 35%,
respectively, as compared to the LPS group. Consistent with the
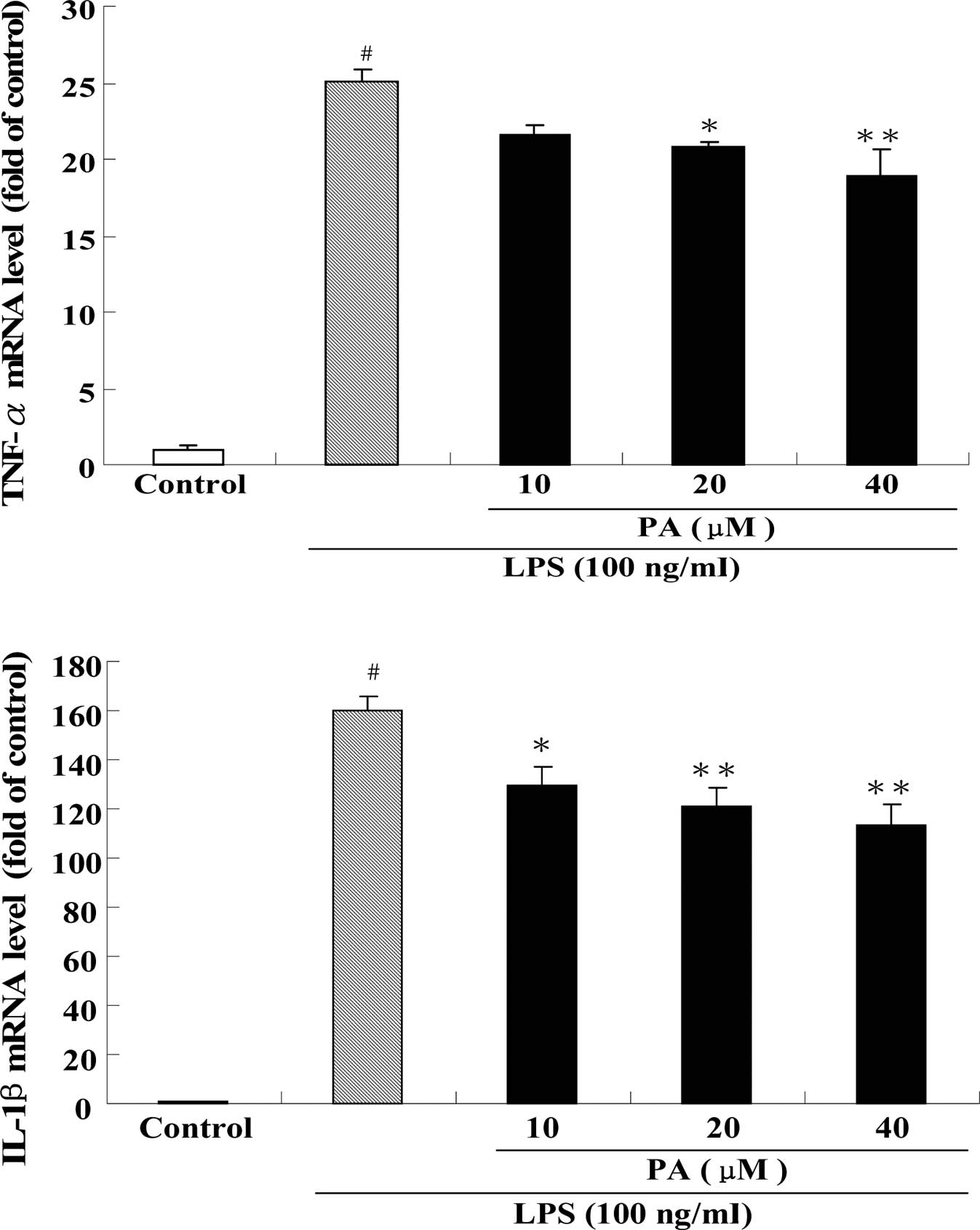
results of the protein levels, real-time PCR revealed that
pre-treatment with PA also significantly inhibited the mRNA
expression of TNF-α, IL-1β and IL-6 in LPS-stimulated RAW264.7
cells (Fig. 6).
Discussion
The present study clearly demonstrated that
treatment of RAW264.7 cells with LPS caused a significant increase
in the protein and mRNA levels of iNOS, COX-2, TNF-α, IL-6 and
IL-1β. However, pre-treatment with PA at concentrations of 10, 20
and 40 μM dose-dependently decreased the protein and mRNA levels of
NO, PGE2, TNF-α and IL-1β in LPS-stimulated RAW264.7
cells.
NO, an important messenger molecule, is intimately
involved in the inflammatory response (26,27).
NO is generated by a family of enzymes called NO synthase (NOS)
(28). It is known that NO plays a
beneficial role in anti-tumor and anti-virus replication, and in
anti-inflammatory processes (29,30).
However, overproduction of NO by macrophages has been implicated in
bacterial septic shock (31), and
in certain inflammatory and autoimmune diseases (32,33).
For this reason, down-regulation of the overproduction of NO has
become a new therapeutic strategy for the treatment of chronic
inflammatory diseases (34,35).
In the present study, we found that exposure of RAW264.7 cells to
LPS caused a significant increase in NO production and iNOS mRNA
expression. This observation is consistent with the findings
previously reported by other researchers (15,36).
Pre-treatment with PA markedly reduced NO production and iNOS mRNA
expression in LPS-stimulated RAW264.7 cells, suggesting that PA is
capable of exerting anti-inflammatory action by inhibiting iNOS
mRNA expression.
PGE2 is considered to be one of the most
potent inflammatory mediators in the inflammatory response and
plays a major role in the pathogenesis of various inflammatory
diseases, edema, angiogenesis, tumor growth and invasion (15,37).
PGE2 is transformed from arachidonic acid via a COX-2
catalytic reaction (38). COX-2
has been demonstrated to be a critical pro-inflammatory enzyme
which contributes to the development of many chronic inflammatory
diseases, such as vascular atherosclerosis and rheumatoid arthritis
(39). Recent years have seen an
increasing interest in the use of COX-2 inhibitors as novel
anti-inflammatory agents. Indeed, non-steroidal anti-inflammatory
drugs (NSAIDs) exert their antipyretic, anti-inflammatory and
analgesic effects through the inhibition of COX activity and the
reduction of the production of inflammatory mediators, such as
PGE2 (15). Our present
study demonstrated that PGE2 production and COX-2 mRNA
expression were markedly increased in LPS-stimulated RAW264.7
cells. However, pre-treatment with PA significantly mitigated the
augmented PGE2 production and COX-2 mRNA expression in
these cells. These results suggest that PA exhibits a selective
inhibitory effect on COX-2 similar to traditional NSAIDs. Thus, PA
is worthy of further development as a new anti-inflammatory
agent.
Cytokines play important roles in the regulation of
inflammation. TNF-α, IL-1β and IL-6 are multifunctional
proinflammatory cytokines and exhibit various pro- inflammatory
effects in chronic inflammatory diseases, such as rheumatoid
arthritis and atherosclerosis (40–42).
TNF-α has long been considered as a key mediator for the induction
of apoptosis and the development of the humoral immune response. At
a high concentration, however, it elicits detrimental effects, such
as causing tissue injury and potentiating septic shock (43,44).
It has been reported that TNF-α elicits downstream pro-inflammatory
events, such as the release of IL-6, another inflammatory cytokine
(45). IL-6 plays essential roles
in host defense, acute phase reactions, immune responses and nerve
cell functions (46–49). A high level of IL-6 has been
observed in various pathological conditions, including bacterial
and viral infections, trauma, autoimmune diseases and inflammations
(47,48). IL-1β is mainly produced by
macrophages, monocytes and T cells and is also involved in immune
defense against infection. In the present study, we clearly
demonstrated that the protein and mRNA levels of TNF-α, IL-1β and
IL-6 were significantly increased in LPS-stimulated RAW264.7 cells,
and pre-treatment with PA dose-dependently reduced the protein and
mRNA levels of TNF-α, IL-1β and IL-6 in these cells. These findings
indicate that the anti-inflammatory effect of PA is associated with
the down-regulation of the mRNA expression of TNF-α, IL-1β and
IL-6.
In conclusion, the present experimental results
demonstrated that PA exerts an anti-inflammatory effect on
LPS-stimulated RAW264.7 cells. The observed anti-inflammatory
action of PA may be mediated, at least in part, by down-regulation
of the mRNA expression of TNF-α, IL-1β, IL-6, iNOS and COX-2 in
LPS-stimulated RAW264.7 cells. These promising experimental
findings not only justify the use of Pogostemonis Herba in Chinese
medicine for inflammatory conditions, but also suggest that PA, an
essential active constituent of Pogostemonis Herba, to be a
potential chemical agent for the treatment of inflammatory
diseases. Further investigation to elucidate the underlying
mechanism of action associated with the anti-inflammatory effect of
PA is currently in progress at our laboratory.
Acknowledgements
This study was supported by a grant
from the National Natural Science Foundation of China (project no.
u0732004).
References
|
1.
|
Chinese Pharmacopoeia Committee:
Pharmacopoeia of China. Chinese Medical Science and Technology
Press; Beijing: pp. 2010
|
|
2.
|
Zhang D, Xiao LY, Cheng YW, Li HL, Feng
ZM, Lin PY, Wu WY and Huang KR: Pharmacological action of Baoji
Pill. Tradit Chin Drug Res Clin Pharmacol. 9:212–214. 1998.
|
|
3.
|
Xian YF, Suo J, Huang XD, Hou SZ, Chen JN,
Ye MR and Su ZR: A pharmacological study on anti-inflammatory
effects of refined Huodan recipe. Chin J Exp Tradit Med Formul.
13:54–56. 2007.
|
|
4.
|
Yang Y, Kinoshita K, Koyama K, Takahashi
K, Tai T, Nunoura Y and Watanabe K: Anti-emetic principles of
Pogostemon cablin (Blanco) Benth. Phytomedicine. 6:89–93.
1999. View Article : Google Scholar
|
|
5.
|
Zhao SC, Jia Q and Liao FL: The
anti-inflammatory and analgesic pharmacological study of Patchouli
extract. Chin Tradit Pat Med. 29:285–287. 2007.
|
|
6.
|
Suo J, Xian YF, Huang XD, Hou SZ, Chen JN,
Ye MR and Su ZR: A pharmacological study on the anti-allergy
effects of refined Houdan recipe. Chin J Exp Tradit Med Formul.
13:47–49. 2007.
|
|
7.
|
Qi SS, Hu LP, Chen WN, Sun HB and Ma XD:
Immunological regulation effects of essential oil in leaves of
Cablin Patchouli herbal on mice. Chin Arch Tradit Chin Med.
27:774–776. 2009.
|
|
8.
|
Liu XR, Fan R, Zhang YY and Zhu MJ: Study
on antimicrobial activities of extracts from Pogestemon
cablin (Blanco) Benth. Food Sci Technol. 24:220–227. 2009.
|
|
9.
|
Yang D, Chaumont JP and Millet J:
Antifungal activity of the essential oils from Agastache
rugosa and Pogostemon cablin against dermatophytes and
opportunistic fungi. Zhongguo Yao Xue Za Zhi. 35:9–11. 2000.
|
|
10.
|
Park EJ, Park HR, Lee JS and Kim J:
Licochalcone A: an inducer of cell differentiation and cytotoxic
agent from Pogostemon cablin. Planta Med. 64:464–466. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Miyazawa M, Okuno Y, Nakamura S and Kosaka
H: Suppression of the furylfuramide-induced SOS response by
monoterpenoids with a p-menthane skeleton using the Salmonella
typhimurium TA1535/pSK1002 Umu test. J Agric Food Chem.
48:642–647. 2000.PubMed/NCBI
|
|
12.
|
Zhao ZZ, Lu J, Leung K, Chan CL and Jiang
ZH: Determination of patchoulic alcohol in herba
Pogostemonis by GC-MS-MS. Chem Pharm Bull. 53:856–860. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Huang XW, Bai L, Xu FH and Wu YJ:
Inhibitory activities of patchouli alcohol on neurotoxicity of
β-amyloid peptide. Jie Fang Jun Yi Xue Za Zhi. 24:338–340.
2008.
|
|
14.
|
Huang XW, Liu RT and Lü QJ: Patchouli
alcohol on memory impairment induced by scopolamine learning and
memory function in mice. Zhong Yao Cai. 40:1431–1433. 2009.
|
|
15.
|
Hu XD, Yang Y, Zhong XG, Zhang XH, Zhang
YN, Zheng ZP, Zhou Y, Tang W, Wang YF, Hu LH and Zuo JP:
Anti-inflammatory effects of Z23 on LPS-induced inflammatory
responses in RAW264.7 macrophages. J Ethnopharmacol. 120:447–451.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Tao JY, Zheng GH, Zhao L, Wu JG, Zhang XY,
Zhang SL, Huang ZJ, Xiong FL and Li CM: Anti-inflammatory effects
of ethyl acetate fraction from Melilotus suaveolens Ledeb on
LPS-stimulated RAW264.7 cells. J Ethnopharmacol. 123:97–105. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Liew FY: The role of innate cytokines in
inflammatory response. Immunol Lett. 85:131–134. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Bertolini A, Ottani A and Sandrini M:
Selective COX-2 inhibitors and dual acting anti-inflammatory drugs:
critical remarks. Curr Med Chem. 9:1033–1043. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Isomaki P and Punnone J: Pro and
anti-inflammatory cytokines in rheumatoid arthritis. Ann Med.
29:499–507. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Korhonen R, Lathi A, Kankaanranta H and
Moilanen E: Nitric oxide production and signaling in inflammation.
Curr Drug Targets Inflamm Allergy. 4:471–479. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Libby P, Aikawa M and Schönbeck U:
Cholesterol and atherosclerosis. Biochim Biophys Acta.
1529:299–309. 2000. View Article : Google Scholar
|
|
22.
|
Shin EM, Zhou HY, Guo LY, Kim JA, Lee SH,
Merfort I, Kang SS, Kim HS, Kim S and Kim YS: Anti-inflammatory
effects of glycyrol isolated from Glycyrrhiza uralensis in
LPS-stimulated RAW264.7 macrophages. Int Immunopharmacol.
8:1524–1532. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Jeong GS, Lee DS and Kim YC: Cudratricus
xanthone A from Cudrania tricuspidata suppresses
pro-inflammatory mediators through expression of anti-inflammatory
heme oxygenase-1 in RAW264.7 macrophages. Int Immunopharmacol.
9:241–246. 2009.PubMed/NCBI
|
|
24.
|
Guan L, Quan LH, Xu LZ and Cong PZ:
Chemical constituents of Pogostemon cablin (Blanco) Benth.
Zhongguo Zhong Yao Za Zhi. 19:355–356. 1994.
|
|
25.
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Zhang XJ, Li Y, Tai GX, Xu GY, Zhang PY,
Yang Y, Lao FX and Liu ZH: Effects of activin A on the activities
of mouse peritoneal macrophages. Cell Mol Immunol. 2:63–67.
2005.PubMed/NCBI
|
|
27.
|
Gayathri B, Manjula N, Vinaykumar KS,
Lakshmi BS and Balakrishnan A: Pure compound from Boswellia
serrata extract exhibits anti-inflammatory property in human
PBMCs and mouse macrophages through inhibition of TNFα, IL-1β, NO
and MAP kinases. Int Immunopharmacol. 7:473–482. 2007.
|
|
28.
|
Palmer RM, Rees DD, Ashton DS and Moncada
S: L-arginine is the physiological precursor for the formation of
nitric oxide in endothelium-dependent relaxation. Biochem Biophys
Res Commun. 153:1251–1256. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Schmidt HH and Walter U: NO at work. Cell.
78:919–925. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
MacMicking J, Xie QW and Nathan C: Nitric
oxide and macrophage function. Annu Rev Immunol. 15:323–350. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Petros A, Bennett D and Vallance P: Effect
of nitric oxide synthase inhibitors on hypotension in patients with
septic shock. Lancet. 338:1557–1558. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
McCartney-Francis N, Allen JB, Mizel DE,
Albina JE, Xie QW, Nathan CF and Wahl SM: Suppression of arthritis
by an inhibitor of nitric oxide synthase. J Exp Med. 178:749–754.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Kleemann R, Rothe H, Kolb-Bachofen V, Xie
QW, Nathan C, Martin S and Kolb H: Transcription and translation of
inducible nitric oxide synthase in the pancreas of prediabetic BB
rats. FEBS Lett. 328:9–12. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Hobbs AJ, Higgs A and Moncada S:
Inhibition of nitric oxide synthase as a potential therapeutic
target. Annu Rev Pharmacol Toxicol. 39:191–220. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Pacher P, Joseph S, Beckman JS and Liaudet
L: Nitric oxide and peroxynitrite in health and disease. Physiol
Rev. 87:315–424. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Lin QY, Jin LJ, Cao ZH and Xu YP:
Inhibition of inducible nitric oxide synthase by Acanthopanax
senticosus extract in RAW264.7 macrophages. J Ethnopharmacol.
118:231–236. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Claria J: Cyclooxygenase-2 biology. Curr
Pharma Des. 9:2177–2190. 2003. View Article : Google Scholar
|
|
38.
|
Rocca B and FitzGerald GA: Cyclooxygenases
and prostaglandins: shaping up the immune response. Int
Immunopharmacol. 2:603–630. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Seibert K, Zhang Y, Leahy K, Hauser S,
Masferrer J, Perkins W, Lee L and Isakson P: Pharmacological and
biochemical demonstration of the role of cyclooxygenase 2 in
inflammation and pain. Proc Natl Acad Sci USA. 91:12013–12017.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Bondeson J: The mechanisms of action of
disease-modifying antirheumatic drugs: a review with emphasis on
macrophage signal transduction and the induction of proinflammatory
cytokines. Gen Pharmacol. 29:127–150. 1997. View Article : Google Scholar
|
|
41.
|
Andreakos E, Foxwell B and Feldmann M: Is
targeting Toll-like receptors and their signaling pathway a useful
therapeutic approach to modulating cytokine-driven inflammation?
Immunol Rev. 202:250–265. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Dayer JM: The process of identifying and
understanding cytokines: from basic studies to treating rheumatic
diseases. Best Pract Res Clin Rheumatol. 18:31–45. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Shohami E, Ginis I and Hallenbeck JM: Dual
role of tumor necrosis factor alpha in brain injury. Cytokine
Growth Factor Rev. 10:119–130. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Yoon HJ, Moon ME, Park HS, Im SY and Kim
YH: Chitosan oligosaccharide (COS) inhibits LPS-induced
inflammatory effects in RAW 264.7 macrophage cells. Biochem Biophys
Res Commun. 358:954–959. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Straub RH, Linde DN, Mannel J, Scholmerich
W and Falk A: A bacteria-induced switch of sympathetic effect or
mechanisms augments local inhibition of TNF-α and IL-6 secretion in
the spleen. FASEB J. 14:1380–1388. 2000.PubMed/NCBI
|
|
46.
|
Hibi M, Nakajima K and Hirano T: IL-6
cytokine family and signal transduction: a model of the cytokine
system. J Mol Med. 74:1–12. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
47.
|
Hirano T, Matsuda T and Nakajima K: Signal
transduction through gp130 that is shared among the receptors for
the interleukin 6 related cytokine subfamily. Stem Cells.
12:262–277. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
48.
|
Van Snick J: Interleukin-6: an overview.
Annu Rev Immunol. 8:253–278. 1990.
|
|
49.
|
Taga T and Kishimoto T: Gp130 and the
interleukin-6 family of cytokines. Annu Rev Immunol. 15:797–819.
1997. View Article : Google Scholar : PubMed/NCBI
|