Introduction
Hand, foot, and mouth disease (HFMD) is a
significant childhood illness. It is characterized by fever and
vesicular eruptions on the hands, feet and mouth. Also, certain
complications occur, although rarely, such as pneumonia,
meningitis, rhabdomyolisis and encephalitis. The most common
infective agents in HFMD are coxsackievirus A16 (CA16) and
enterovirus 71 (EV71) (1). A large
number of studies have focused on EV71-associated HFMD while less
attention has been paid to CA16 strains due to the mild and benign
symptoms following infection (1,2). Based
on phyletic evolution analysis, CVA16 is closely related to EV71;
however, the pathogenesis and symptoms may differ, which has not
been fully determined (1). Since
2007, large-scale outbreaks of CA16 infection in China have
occurred (3). Genetic diversities
and the evolution of CA16 strains have been identified in several
studies (2). A total of 42 CV16
strains have been reported from different locations in China,
including Inner Mongolia, Gansu, Qinghai and Shandong provinces
(2). With the exception of strain
identification, little information has been presented on the
pathogenetic mechanisms. A previous study demonstrated that CA16
plays only a subsidiary role in the pathogenesis (4). Nevertheless, understanding the
mechanism of the effects of CA16 is likely to aid therapeutic
strategies for HFMD.
CV16 and EV71 share the same receptor, scavenger
receptor class B member 2 (SCARB2) (5). By developing transgenic mice with
expression of human SCARB2, Chang et al investigated the
efficacy conferred by a neutralizing antibody, N3 (5). The results showed that accumulated
viral loads and the expression of pro-inflammatory cytokines were
positively correlated. The authors concluded that the neutralizing
antibody, N3, could be used as a new potential therapy for treating
severe HFMD. The immunological reactions following EV71 infection
have been deeply studied. Changes in the expression levels of
several inflammatory mediators following EV71 infection have been
investigated, including interferon (IFN)-γ, interleukin (IL)-13,
IL-6, IL-1β and IL-10, as well as chemokines IL-8 and IL-10
(6). The expression of these
mediators was found to correlate with disease severity and outcome.
In a mouse model, the mice with a lack of CD4+ or
CD8+ T cells had more severe disease (7). In addition, high expression levels of
IL-6 were found in newborn mice following EV71 infection and
resulted in tissue damage (8). Li
et al reported that Th1/Th2 and Th17/Treg imbalances were
observed following EV71 infection (9). These findings illustrate the importance
of cellular immunity in the pathogenesis of HFMD. However, at
present, the pathogenesis of CV16 remains unclear.
T cells, also named T lymphocytes, play a crucial
role in immune regulation. Dependent upon the cluster of
differentiation (CD) receptors that they express, these cells can
be classified into CD4+ and CD8+ T cells
(10). CD4+ T cells
include Th1, Th2, Th17 and Treg subsets whereas CD8+ T
cells include Tc1, Tc2 and Tc17 subsets. Type 1 cytokines, such as
IL-2, tumor necrosis factor (TNF)-α and IFN-γ, are secreted by Th1
and Tc1 cells while type 2 cytokines such as IL-4, IL-5 and IL-13
are secreted by Th2 and Tc2 cells (11). Th17 and Tc17 cells secrete IL-17,
while Treg cells are characterized by IL-10 and TGF-β expression
(12). This secretion specificity
and the different kinds of T cell subsets are in balance under
normal conditions; however, when organisms suffer from an
infectious disease, the system becomes imbalanced. For example,
following Staphylococcus aureus implant infection, the
Th1/Th17 response increased while Th2/Treg also responded to
chronic infection (13). In
EV71-infected patients with HFMD, the percentages of Th1, Tc1 and
Th17 cells were found to be increased, and the Th1/Th2 ratio and
IL-17A and IFN-γ levels were upregulated (9). These studies demonstrate the imbalance
of inflammatory mediators and T cells in the pathogenesis of HFMD
subsequent to EV71 infection. Also, they suggest that T-cell subset
imbalance has an important role in the pathogenesis of
infection.
Studies concerning different T-cell subsets in
CA16-induced HFMD are very limited compared with those on
EV71-induced HFMD. As there is a lack of information concerning
vaccines or drugs for the prevention of CA16 infection, an
increased understanding of CA16 infection is likely to contribute
to new therapeutic strategies. Thus, in the present study, to
elucidate the changes in the balance of T-cell subsets associated
with CA16 infection, the distribution of T-cell subsets as well as
the mediator expression in these cells were detected. These data
may provide new insights for future study and be of diagnostic and
therapeutic value for CA16-induced HFMD.
Materials and methods
Clinical samples
Clinical samples were collected from the Department
of Pediatrics, Shilong People's Hospital (Dongguan, China). CA16
infection was identified by reverse transcription quantitative
polymerase chain reaction (RT-qPCR) of nucleic acids in throat
swabs or stool specimens. In total, 100 CA16 infected children with
HFMD (aged 0.2–6.1 years old) were enrolled in this study. All the
samples were taken at the same time, which was 3 days from the
appearance of HFMD symptoms. In addition, 30 healthy children with
no symptoms of HFMD were enrolled as controls. The clinical
collection was approved by the Ethics Committee of the Department
of Pediatrics, Dongguan Shilong People's Hospital. In addition,
informed consent was obtained from the parents or guardians of all
the children that participated in the present study.
Sample preparation
Samples, including throat swabs and stool samples,
were collected from the patients or control children and then
stored at −80°C until analyzed. Also, blood samples were collected
and the lymphocytes were isolated according to the manufacturer's
instruction (Lymphoprep™; Axis-Shield, Oslo, Norway).
Peripheral blood mononuclear cells (PBMCs) were suspended at
density of 1×106 cells/ml in RPMI-1640 (Gibco, Grand
Island, NY, USA) with 10% fetal bovine serum (Gibco).
RT-qPCR
The RNAs were extracted from throat swabs and stool
specimens using a PureLink® Viral RNA/DNA Mini kit (Invitrogen Life
Technologies, Carlsbad, CA, USA) according to the manufacturer's
instructions. The detection of CA16 was conducted by RT-qPCR as
described in a previous study by Xiao et al (14). In brief, the RT-PCR thermal profile
consisted of: reverse transcription program (50°C for 30 min),
denaturation program (95°C for 3 min) and amplification program
repeated 40 times (95°C for 5 sec, 60°C for 40 sec) and FAM, VIC
and CY5 fluorescence measurement at 60°C. RT-qPCR was carried out
on a ABI 7500 Real-Time PCR system (Applied Biosystems®, Invitrogen
Life Technologies) The primers (purchased from Sangon Biotech,
Shanghai, China) were as follows: tumor necrosis factor (TNF)-γ
forward: 5′-TGGCTTAATTCTCTCGGAAACG-3′, reverse: 5′-TTT
TACATATGGGTCCTGGCAGTA-3′ and probe: 5′-ATCTTG
GCTTTTCAGCTCTGCATCGTTTT-3′; IL-4 forward:
5′-CCCTCTGTTCTTCCTGCTAGCA-3′, reverse: 5′-CCT
GTAAGGTGATATCGCACTTGT-3′ and probe: 5′-TGCCGG CAACTTTGTCCACGG-3′;
IL-17A forward: 5′-GGGCCT GGCTTCTGTCTGA-3′, reverse:
5′-AAGTTCGTTCTG CCCCATCA-3′ and probe: 5′-CAAGGCACCACACAACCCA
GAAAGGA-3′; transforming growth factor (TGF)-α forward
5′-GCCATTCTGGGTACGTTGGT-3′, reverse: 5′-TGATGG CCTGCTTCTTCTGA-3′
and probe: 5′-CACGCTGTGAGC ATGCGGACCTC-3′.
Enzyme-linked immunosorbent assay
(ELISA)
In the present study, ELISA was used to detect the
expression levels of IFN-γ, IL-4, IL-17A and TGF-α in plasma using
IFN-γ, IL-4, IL-17A and TGF-α ELISA kits provided by R&D
Systems (Minneapolis, MN, USA). The assays were conducted according
to the manufacturer's instructions. All samples were tested in
three replicated assays.
Flow cytometric analysis
For the analysis of different subsets of T cells,
the cell suspension was provoked by culture with 25 ng/ml phorbol
12-myristate 13-acetate (Sigma-Aldrich, St. Louis, MO, USA) and 1
µg/ml ionomycin (Sigma-Aldrich,) at 37°C for 4 h. Subsequently, the
cells were incubated with polyclonal PerCP-Cy5.5-labeled goat
anti-human and polyclonal FITC-labeled goat anti-human CD8
antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 20
min at 4°C. For Treg cells, the cell suspension was incubated with
a mixture of polyclonal FITC-labeled goat anti-human CD4 and
polyclonal PE-labeled goat anti-human CD25 (Santa Cruz
Biotechnology) for 20 min at 4°C. Subsequently, the cells were
incubated with polyclonal Alexa Fluor 647-labeled goat anti-human
Foxp3 (Santa Cruz Biotechnology). After fixing and permeabilizing,
the stained cells were assayed by an Epics XL-MCL flow cytometer
(Beckman Coulter, Miami, FL, USA). The ratios of the cells were
calculated by the antibody signals of the specific proteins.
Isotype controls were used to correct compensation and confirm
antibody specificity. All calculations were performed using EXPO32
ADC software (Beckman Coulter).
Statistic analysis
Statistical analysis was performed using SPSS
software, version 13.0 (SPSS, Inc., Chicago, IL, USA). The
significance of differences was determined by one-way analysis of
variance (ANOVA). Differences were considered significantly if
P≤0.05. If F ratios exceeded the critical value (P≤0.05), the
Newman-Keul's post-hoc test was performed to compare the
groups.
Results
Case overview
Of the 100 patients with HFMD, 49 and 51 patients
were diagnosed with mild and severe HFMD, respectively, according
to the diagnostic criteria described in a previous study (15). No significant differences in age and
gender were identified among the studied groups. The duration of
hospitalization and fever duration were significantly higher in the
severe group compared with the control and mild groups. Among all
100 patients with HFMD, all cases were confirmed by RT-qPCR to be
infected with CA16, and CA16 nucleic acid was detected in both
throat swabs and stool specimens. The results showed that the
severe group had the higher number of copies of CA16 in both throat
swabs and stool species (Table
I).
 | Table I.Basic information of studied
samples. |
Table I.
Basic information of studied
samples.
| Variables | Control | Mild | Severe |
|---|
| Number of
samples | 30 | 49 | 51 |
| Age (years) | 2.05±1.62 | 2.31±1.54 | 1.97±1.28 |
| Gender
(male/female) | 15/15 | 24/25 | 27/24 |
| Length of
hospitalization (days) | 0 | 6.00 (4.00–7.00) | 9.00
(7.00–11.00)a |
| Highest body
temperature (°C) | - | 38.42±0.88 | 38.78±0.48 |
| Fever duration
(days) | - | 2.13±0.53 |
4.53±0.62a |
| WBC
(×109/l) | 7.03 (5.47–8.11) | 11.53
(9.39–13.02) | 12.32
(10.83–14.02)a |
| Neutrophil (%) | 59.35±8.63 | 57.56±7.26 | 60.23±4.69 |
| CRP (mg/l) | - | 7.00 (5.00–9.53) | 7.51
(4.53–10.04) |
| CA16 (copies/ml)
hroat swabs | 0 |
5×106-4×107 |
5×108-4×1010a |
| CA16 (copies/ml)
stool | 0 |
3×104-4×105 |
2×105-7×108a |
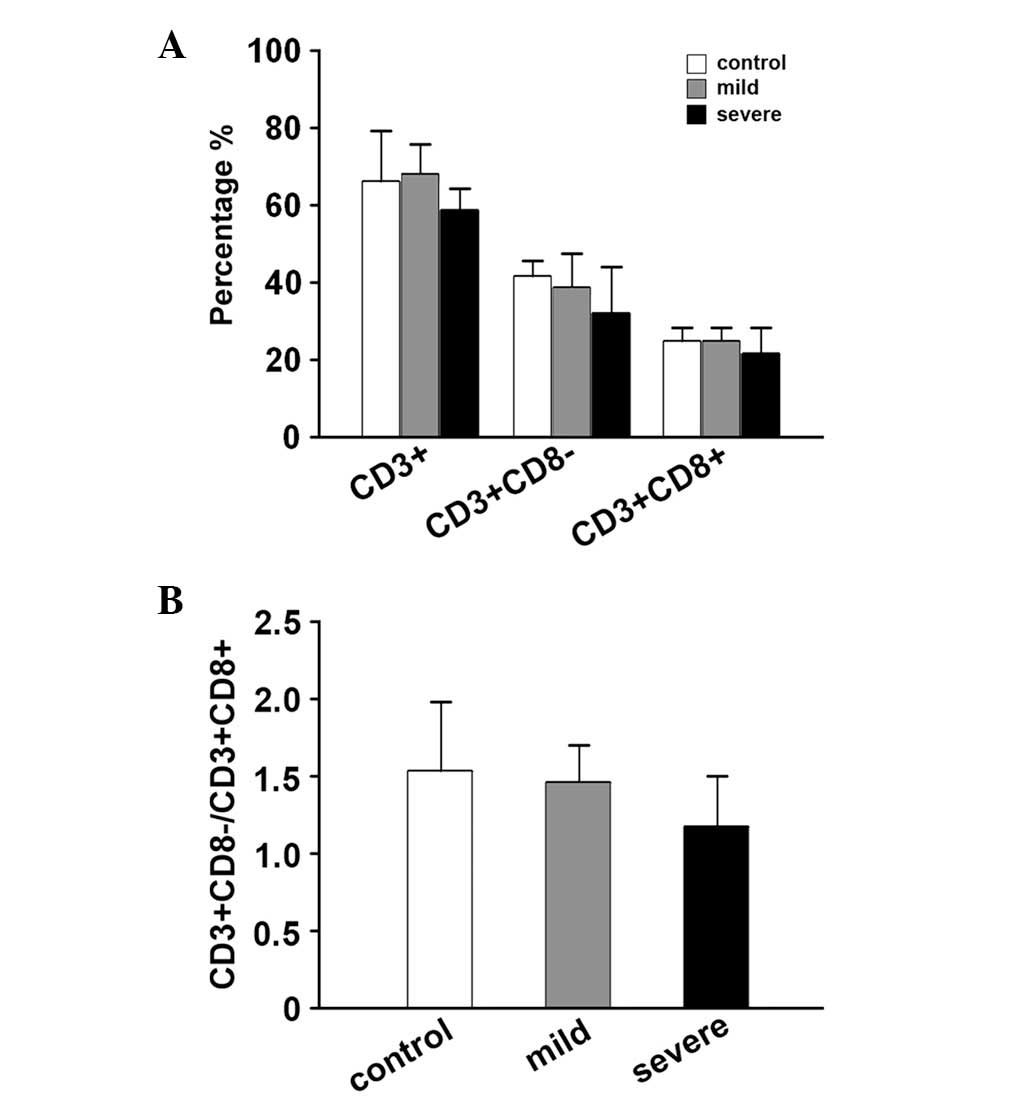
The percentages of different subsets of T cells in
the control, mild HFMD and severe HFMD groups were assayed. The
percentages of CD3+ T cells in the control, mild HFMD
and severe HFMD groups were 67.32±12.52, 70.03±7.37 and
58.92±8.74%, respectively. The percentages of
CD3+CD8− T cells in the control, mild HFMD
and severe HFMD groups were 43.44±4.85, 40.05±7.55 and
37.22±13.24%, respectively. The percentages of
CD3+CD8+ T cells in the control, mild HFMD
and severe HFMD groups were 26.23±2.03, 26.18±2.23 and 19.63±4.03%,
respectively. Statistically, the percentages of these subsets of T
cells were not significantly different among all the tested groups
(Fig. 1A). The values of
CD3+CD8− and CD3+CD8+
were used to calculate the
CD3+CD8−/CD3+CD8+
ratio. This ratio also was not identified to differ significantly
among the groups (Fig. 1B).
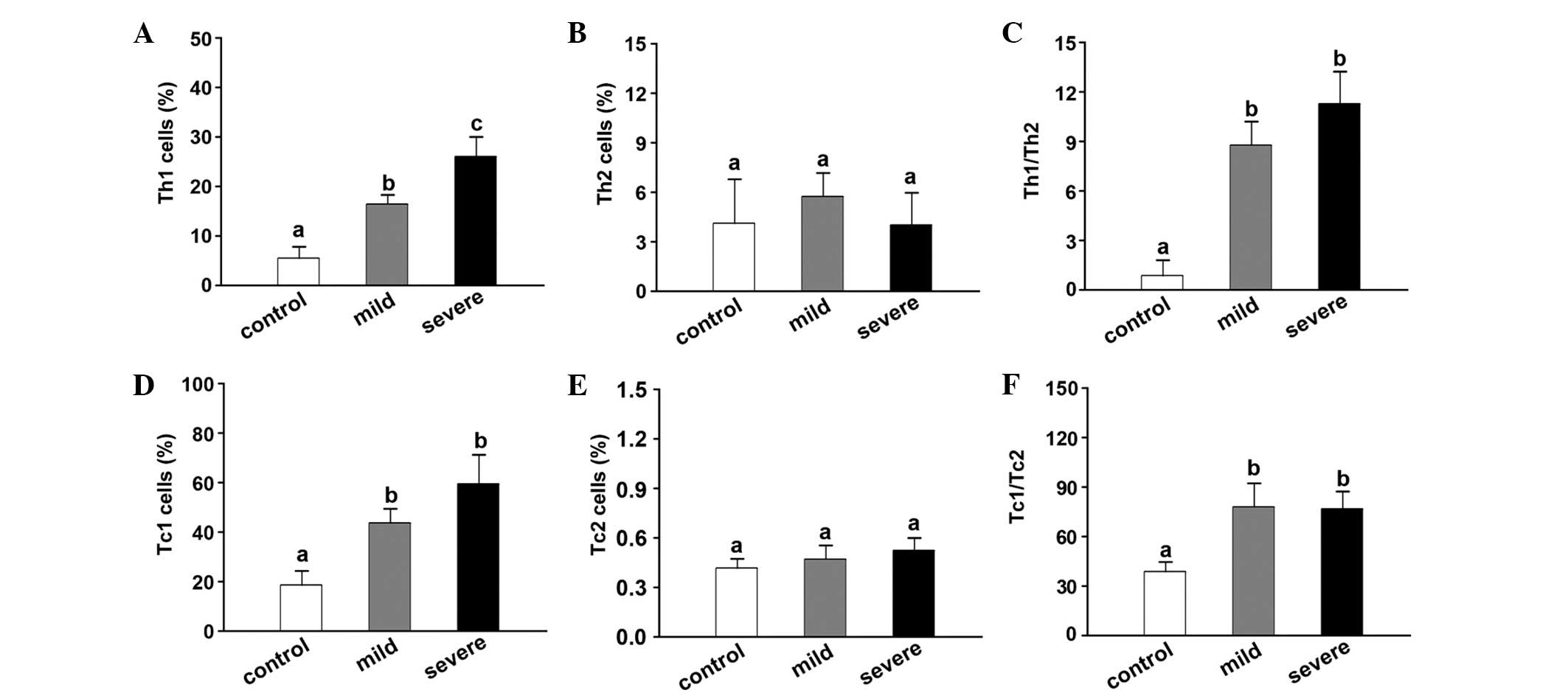
Imbalance of Th1, Th2, Tc1 and Tc2 in
CA16-infected patients
The percentages of Th1 cells were 6.12±2.05,
18.52±1.98 and 26.53±5.69% in the control, mild HFMD and severe
HFMD groups, respectively. The Th1 percentage in each group was
found to be significantly different compared with that in the other
two groups (Fig. 2A). By contrast,
no such difference was identified in the Th2 cell percentage
(Fig. 2B). The Th1/Th2 ratio was
significantly increased in the mild (8.82±1.98) and severe groups
(11.52±28.92) compared with that in the control group (0.93±1.02;
Fig. 2C).
The Tc1 cell percentage was significantly higher in
the mild (42.35±4.52%) and severe (58.68±11.37%) HFMD groups
compared with that in the control group (18.52±4.37%; Fig. 2D). By contrast, the Tc2 cell
percentage did not significantly differ among the groups (Fig. 2E). In a similar manner to the Th1/Th2
ratio, the Tc1/Tc2 ratio was higher in the mild (72.23±14.92) and
severe HFMD groups (70.79±17.54) compared with that in the control
group (41.13±5.89; Fig. 2F).
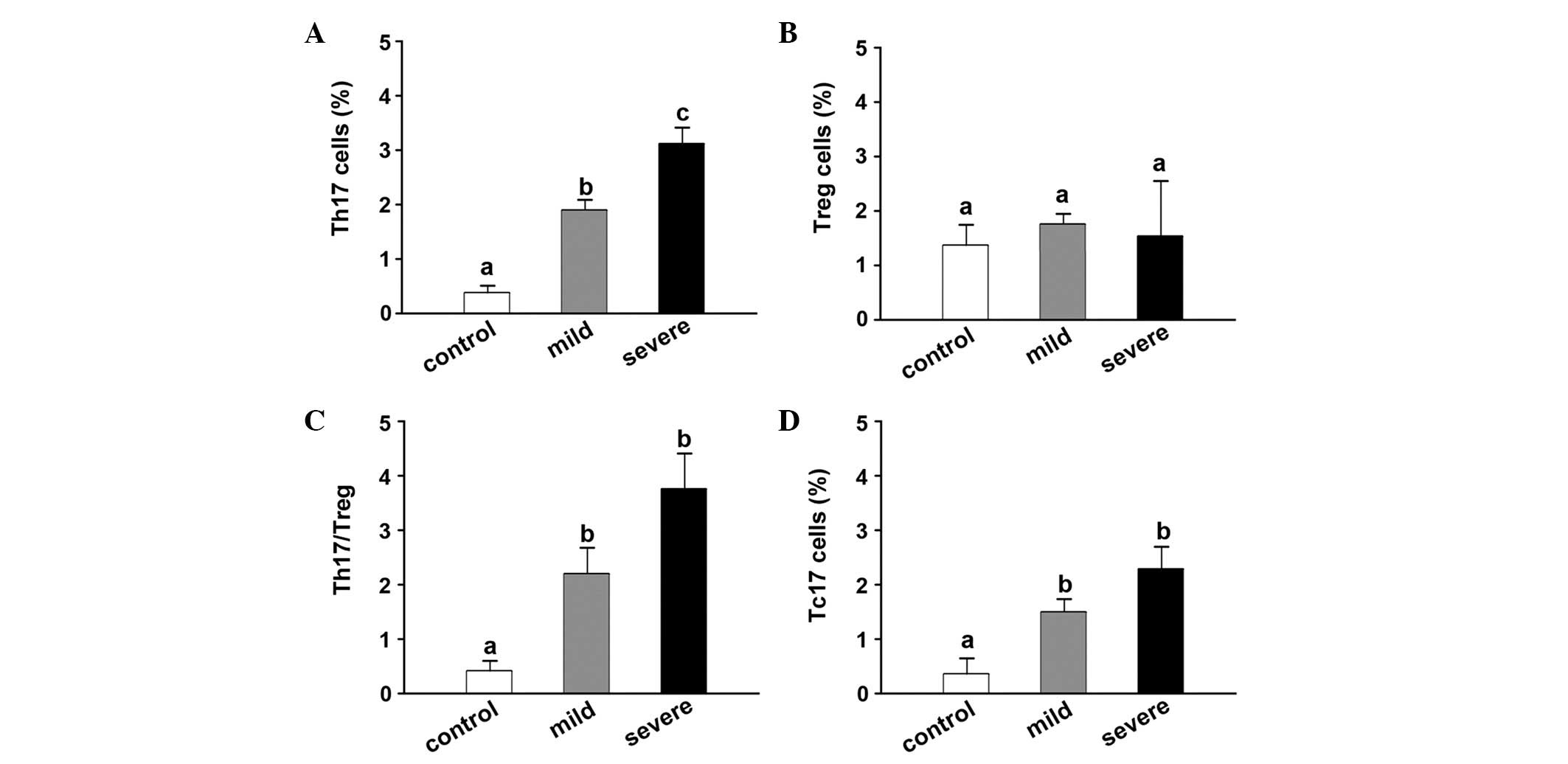
Disrupted balance of Th17/Treg in
CA16-infected patients
The cell frequencies of Th17 were significant higher
in the mild (1.83±0.17%) and severe HFMD groups (3.21±0.39%) than
in the control group (0.41±0.14%; Fig.
3A), while the percentage of Treg cells was not identified to
be significantly different among the groups (Fig. 3B). The Th17/Treg ratio was also found
to be higher in the mild (2.23±0.42) and severe HFMD groups
(3.76±0.72) compared with that in the control group (0.43±0.17;
Fig. 3C). Furthermore, the
percentage of Tc17 cells was also significantly higher in the mild
(1.73±0.14%) and severe HFMD groups (2.35±0.54%) than in the
control group (0.41±0.17%; Fig.
3D).
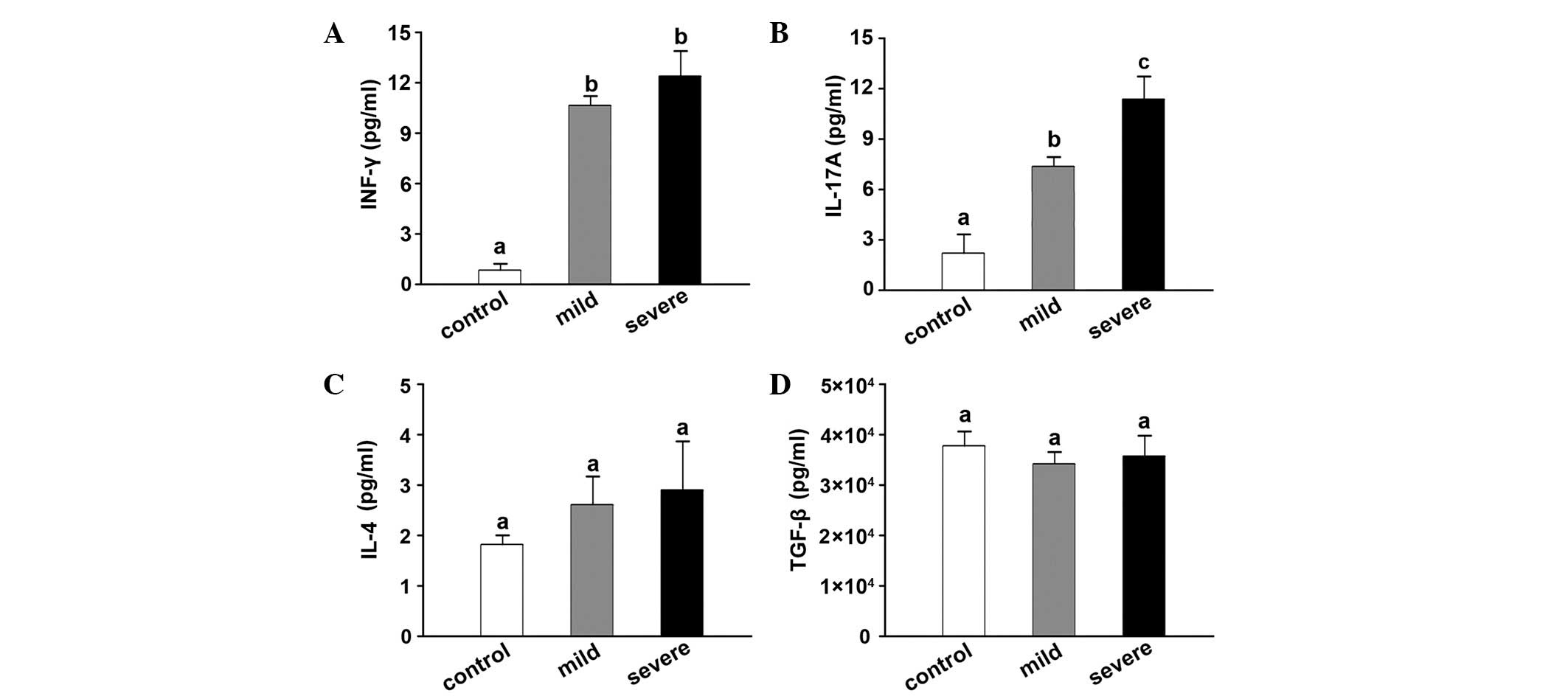
CA16 infection changes cytokine levels
in plasma
The expression levels of immune mediators,
specifically IFN-γ, IL-17A, IL-4 and TGF-β in plasma were
determined by ELISA. The levels of IFN-γ were significant higher in
the mild (11.03±0.75 pg/ml) and severe (12.14±1.49 pg/ml) HFMD
groups than in the control group (1.06±0.25 pg/ml; Fig. 4A). Similarly, IL-17A exhibited higher
levels in the mild (2.68±0.56 pg/ml) and severe (11.52±1.22 pg/ml)
HFMD groups compared with that in the control group (2.82±1.25
pg/ml; Fig. 4B). However, the levels
of IL-4 and TGF-β were not identified to be significantly different
among the three groups (Fig. 4C and
D).
Discussion
At present, a significant amount of evidence
indicates that inappropriate inflammation participates in the
progression of HFMD (11,13). However, when more mild and benign
symptoms are present following infection, little information has
been unveiled concerning the adaptive immunity of the infected
host. In the present study, the T-cell profile and related immune
mediators were analyzed in CA16-infected children with HFMD, in
order to investigate the association between changes in T cells and
HFMD. The results demonstrate that CA16-infected children had
higher percentages of Th1, Th17, Tc1 and Tc17 cells as well as
plasma IFN-γ and IL-17A levels than healthy control children had.
These results indicate that the immune system is disordered
following CA16 infection. In concurrence with the present study, Li
et al observed an imbalance of T-lymphocyte subsets with
abnormal production of IFN-γ and IL-17A following EV71 infection in
children with HFMD (9). In addition,
the EV71-related encephalitis was associated with a significant
increase in the levels of pro-inflammatory cytokines, such as IL-6,
TNF-α and IL- 1β. Together, these findings imply that HFMD-related
infection, including EV71 and CA16 infection, contributes to
dysregulation of the immune system.
So far, to the best of our knowledge, no information
depicting the role of Th1 in CA16 infected HFMD patients has been
published. Th1 cytokines have been shown to have a crucial role in
the pathogenesis of EV71-infected HFMD (9). In the present study, it was
demonstrated that there was no significant difference in Th2 and
Tc2 percentages among the three groups, while IFN-γ production by T
cells (Th1 and Tc1) was significantly increased following CA16
infection. In addition, the Th1/Th2 ratio was also shown to rise in
children with HFMD. These results indicate the CA16 dysregulated
the balance between Th1 and Th2, and suggest that CA16 infection in
HFMD may associated with Th1-promoted disease.
The Th1/Th2 ratio hypothesis was proposed from
research in 1986 suggesting that mouse T-helper cells expressed
differing cytokine patterns (16).
Th1 cells drive cellular immunity while Th2 cells drive humoral
immunity. The Th1/Th2 ratio thus reflects the cellular/humoral
immunity balance following pathogen invasion. The Th1/Th2 ratio
clearly indicates immunological changes in mammalians. The Th1/Th2
ratio hypothesis explains cancer, diabetes and pathogenic infection
well (17–19). However, the Th1/Th2 ratio hypothesis
also demonstrates limitations in several diseases, including
asthma, rheumatoid arthritis and chronic hepatitis B (19–21). In
the present study, the Th1/Th2 balance was affected in children
with HFMD. However, no significant difference could be observed in
the Th1/Th2 ratio between patients with mild and severe HFMD. This
shows a certain limitation of the Th1/Th2 balance theory in
children with HFMD. Thus, changes in T-cell subsets participate in
CA16-induced HFMD. Treg and Th17 cells have been identified to
participate in multidirectional immunological reaction following
Th1/Th2 balance changes (22,23). The
present study also revealed a change in the Th17 and Th17/Treg
ratio that was associated with the disease severity. Based on these
findings, the imbalance of Th1/Th2 cells may stimulated by HFMD,
while the Th17/Treg imbalance may play a decisive function in HFMD
progression. These findings are consistent with a previous study by
Chen et al, which identified that Th17 cells increased
significantly in children with HFMD, suggesting that Th17-mediated
immune response plays a crucial role in the progression of HFMD
(24). Notably, the percentage of
Th17 cells was higher in the severe group compared with the mild
group. Thus, it was also found that the level of IL-17A, which is
secreted by Th17 cells, also increased. IL-17A is known to
contribute to the inflammatory response and induces the production
of several cytokines and chemokine mediators. As result, neutrophil
activation processes, such as differentiation, maturation and
migration, are under the control of IL-17A (25). This is in accordance with the finding
that the white blood cell count increased following CA16 infection
in the children in the present study. Li et al demonstrated
that Th17 cells and IL-17A are involved in the progression of
EV71-related HFMD (9). Thus, these
data suggest that changes in the levels of Th17 cells and
increasing levels of IL-17A secretion are key factors in
CA16-induced HFMD.
In contrast to the acute increase in the levels of
Th17 cells and IL-17A in plasma, other inflammatory mediators such
as IL-4 and TGF-β showed no changes following CA16 infection. The
percentage of Treg cells was also unchanged in children with HFMD.
This also implies the importance of Th17 cell function in the
pathogenesis of HFMD rather than the function of Treg cells.
In addition, no significant difference could be
identified between mild and severe HFMD cases. These finding is
comparable with the results in the studies by Yang et al
(26) and Li et al (9). The authors demonstrated that the viral
load of EV71 was not correlated with certain immune factors
statistically, including IL-4 and TGF-β. These data suggest that
the viral loads in throat swabs or stool specimens may not
correlate with several T-cell subsets as well as the immune factors
in plasma. Lin et al reported that the changes in T-cell
subsets exhibited a tissue-specific profile (27). They confirmed that throat swab and
stool specimens showed a reduced viral load of EV71 than the
specific tissue that is the focus of infection, and thus are not
adequate for diagnosis. Therefore, it would be interest to
elucidate the precise tissue-specific profile of different T-cell
subsets in CA16-induced HFMD in future studies.
In conclusion, the present study reported that a
dysregulation of T-cell subsets occurs in CA16-induced HFMD, as
well as imbalances in the Th1/Th2 and Th17/Treg ratios following
infection. The imbalances contribute to increases in the levels of
IFN-γ and IL-17A accordingly. However, the molecular mechanisms of
CA16-induced HFMD remain insufficiently understood. The present
findings provide a new insight for future studies of CA16-induced
HFMD, and provide information of diagnostic and therapeutic values
for CA16 infection.
References
|
1
|
Iwai M, Masaki A, Hasegawa S, et al:
Genetic changes of coxsackievirus A16 and enterovirus 71 isolated
from hand, foot, and mouth disease patients in Toyama, Japan
between 1981 and 2007. Jpn J Infect Dis. 62:254–259.
2009.PubMed/NCBI
|
|
2
|
Zhang Y, Wang D, Yan D, et al: Molecular
evidence of persistent epidemic and evolution of subgenotype B1
coxsackievirus A16-associated hand, foot, and mouth disease in
China. J Clin Microbio. 48:619–622. 2010. View Article : Google Scholar
|
|
3
|
Ding NZ, Wang XM, Sun SW, Song Q, Li SN
and He CQ: Appearance of mosaic enterovirus 71 in the 2008 outbreak
of China. Virus Res. 145:157–161. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu LJ, Xu HM, Li XJ, et al: Co-detection
in the pathogenesis of severe hand-foot-mouth disease. Arch Virol.
157:2219–2222. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chang HW, Lin YW, Ho HM, et al: Protective
efficacy of VP1-specific neutralizing antibody associated with a
reduction of viral load and pro-inflammatory cytokines in human
SCARB2-transgenic mice. PLoS One. 8:e698582013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang SM, Lei HY, Huang MC, et al:
Modulation of cytokine production by intravenous immunoglobulin in
patients with enterovirus 71-associated brainstem encephalitis. J
Clin Virol. 37:47–52. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Belkaid Y and Rouse BT: Natural regulatory
T cells in infectious disease. Nat Immunol. 6:353–360. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Khong WX, Foo DG, Trasti SL, Tan EL and
Alonso S: Sustained high levels of interleukin-6 contribute to the
pathogenesis of enterovirus 71 in a neonate mouse model. J Virol.
85:3067–3076. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li S, Cai C, Feng J, et al: Peripheral T
lymphocyte subset imbalances in children with enterovirus
71-induced hand, foot and mouth disease. Virus Res. 180:84–91.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gocinski BL and Tigelaar RE: Roles of CD4+
and CD8+ T cells in murine contact sensitivity revealed by in vivo
monoclonal antibody depletion. J Immunol. 144:4121–4128.
1990.PubMed/NCBI
|
|
11
|
Lucey DR, Clerici M and Shearer GM: Type 1
and type 2 cytokine dysregulation in human infectious, neoplastic,
and inflammatory diseases. Clin Microbiol Rev. 9:532–562.
1996.PubMed/NCBI
|
|
12
|
Jutel M, Akdis M, Budak F, et al: IL-10
and TGF-β cooperate in the regulatory T cell response to mucosal
allergens in normal immunity and specific immunotherapy. Eur J
Immunol. 33:1205–1214. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Prabhakara R, Harro JM, Leid JG, Keegan
AD, Prior ML and Shirtliff ME: Suppression of the inflammatory
immune response prevents the development of chronic biofilm
infection due to methicillin-resistant Staphylococcus aureus.
Infect Immun. 79:5010–5018. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiao XL, Wu H, Li YJ, et al: Simultaneous
detection of enterovirus 70 and coxsackievirus A24 variant by
multiplex real-time RT-PCR using an internal control. J Virol
Methods. 159:23–28. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ooi MH, Wong SC, Podin Y, et al: Human
enterovirus 71 disease in Sarawak, Malaysia: a prospective
clinical, virological, and molecular epidemiological study. Clin
Infect Dis. 44:646–656. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mosmann TR, Cherwinski H, Bond MW, Giedlin
MA and Coffman RL: Two types of murine helper T cell clone. I.
Definition according to profiles of lymphokine activities and
secreted proteins. J Immunol. 136:2348–2357. 1986.PubMed/NCBI
|
|
17
|
Myint AM, Leonard BE, Steinbusch HW and
Kim YK: Th1, Th2, and Th3 cytokine alterations in major depression.
J Affect Disord. 88:167–173. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Romagnani S, Maggi E and Del Prete G: An
alternative view of the Th1/Th2 switch hypothesis in HIV infection.
AIDS Res Hum Retroviruses. 10:iii–ix. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kuo ML, Huang JL, Yeh KW, Li PS and Hsieh
KH: Evaluation of Th1/Th2 ratio and cytokine production profile
during acute exacerbation and convalescence in asthmatic children.
Ann Allergy Asthma Immunol. 86:272–276. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Okazaki K, Uchida K, Ohana M, et al:
Autoimmune-related pancreatitis is associated with autoantibodies
and a Th1/Th2-type cellular immune response. Gastroenterology.
118:573–581. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Taams LS, Palmer DB, Akbar AN, Robinson
DS, Brown Z and Hawrylowicz CM: Regulatory T cells in human disease
and their potential for therapeutic manipulation. Immunology.
118:1–9. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Weaver CT and Hatton RD: Interplay between
the TH17 and TReg cell lineages: a (co-) evolutionary perspective.
Nat Rev Immunol. 9:883–889. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu J and Paul WE: Heterogeneity and
plasticity of T helper cells. Cell Res. 20:4–12. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen J, Tong J, Liu H, et al: Increased
frequency of Th17 cells in the peripheral blood of children
infected with enterovirus 71. J Med Virol. 84:763–767. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ley K, Smith E and Stark MA:
IL-17A-producing neutrophil-regulatory Tn lymphocytes. Immunol Res.
34:229–242. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang X, Wu Q, Wen B, Dai H, Zhuang Y and
Yang J: Prevalent characteristics in children with hand-foot-mouth
disease in the Kunming area in 2010. Zhongguo Dang Dai Er Ke Za
Zhi. 14:192–194. 2012.[(In Chinese)]. PubMed/NCBI
|
|
27
|
Lin YW, Chang KC, Kao CM, Chang SP, Tung
YY and Chen SH: Lymphocyte and antibody responses reduce
enterovirus 71 lethality in mice by decreasing tissue viral loads.
J Virol. 83:6477–6483. 2009. View Article : Google Scholar : PubMed/NCBI
|