Introduction
DNA replication is essential not only for
duplication of the genome but also for maintenance of genomic
integrity during DNA repair (1,2).
Chromosomal DNA replication in eukaryotic cells requires three
distinct DNA polymerases-α, -δ (Polδ), and -ɛ (1–5).
Mammalian Polδ plays a crucial and versatile role in DNA
replication and various DNA repair processes (6). Its function as a chromosomal DNA
polymerase is dependent on the association with proliferating cell
nuclear antigen (PCNA) which functions as a molecular sliding clamp
(7,8). Similar to S. pombe enzyme,
the mammalian Polδ consists of four subunits: p125, p50 (also named
DNA polymerases δ 2 small subunit, Pold2), p66 and p12, forming a
heterotetrameric complex (4,9).
It has been reported that Pold2 serves as a scaffold for the
assembly of Polδ by interacting simultaneously with all of the
other three subunits (10). In
addition, Pold2 is also involved in the recruitment of several
proteins regulating DNA metabolism, including p21, PDIP1
(polymerase δ-interacting protein 1, PDIP1), PDIP38, PDIP46 and WRN
(Werner protein, WRN) (11–14).
PIAS2 [protein inhibitor of activated STAT (signal
transducer and activator of transcription) 2, PIAS2] belongs to the
family of PIAS proteins, which includes PIAS1, PIAS2, PIAS3 and
PIASy (15–18). PIAS proteins were initially
identified as specific cofactors inhibiting DNA binding and
transcriptional activation by the STAT family of transcription
factors (15). These proteins
typically contain an N-terminal SAP domain, an N-terminal PINIT
domain, a central RING-like zincfinger (RLD) domain and a
C-terminal acidic domain (19,20). The human PIAS2 gene encodes two
splice variants, PIASxα (also called ARIP3, androgen
receptor-interacting protein 3) and PIASxβ (also known as Miz1,
Msx-interacting-zinc finger) (16,21). The mouse PIAS2 gene has at least
five isoforms (isoforms 1–5), which differ in the N- and C-termini
as a result of alternative splicing. The mouse PIAS2 protein shows
99.4% sequence identity to the human homologue. Although PIAS2 was
originally identified as a protein that binds to and inhibits
STAT4, the functions of PIAS2 protein are clearly not limited to
the regulation of STAT4. PIAS2 is capable of interacting with and
modulating (activating and repressing) transcriptional activities
of several transcription factors (22–24).
To determine the molecular mechanism of PIAS2
function, we used the yeast two-hybrid screening technique to
identify interacting proteins associated with PIAS2. Interestingly,
Pold2 was found to interact with PIAS2. Here we report evidence for
a direct interaction between Pold2 and PIAS2.
Materials and methods
Yeast two-hybrid analysis
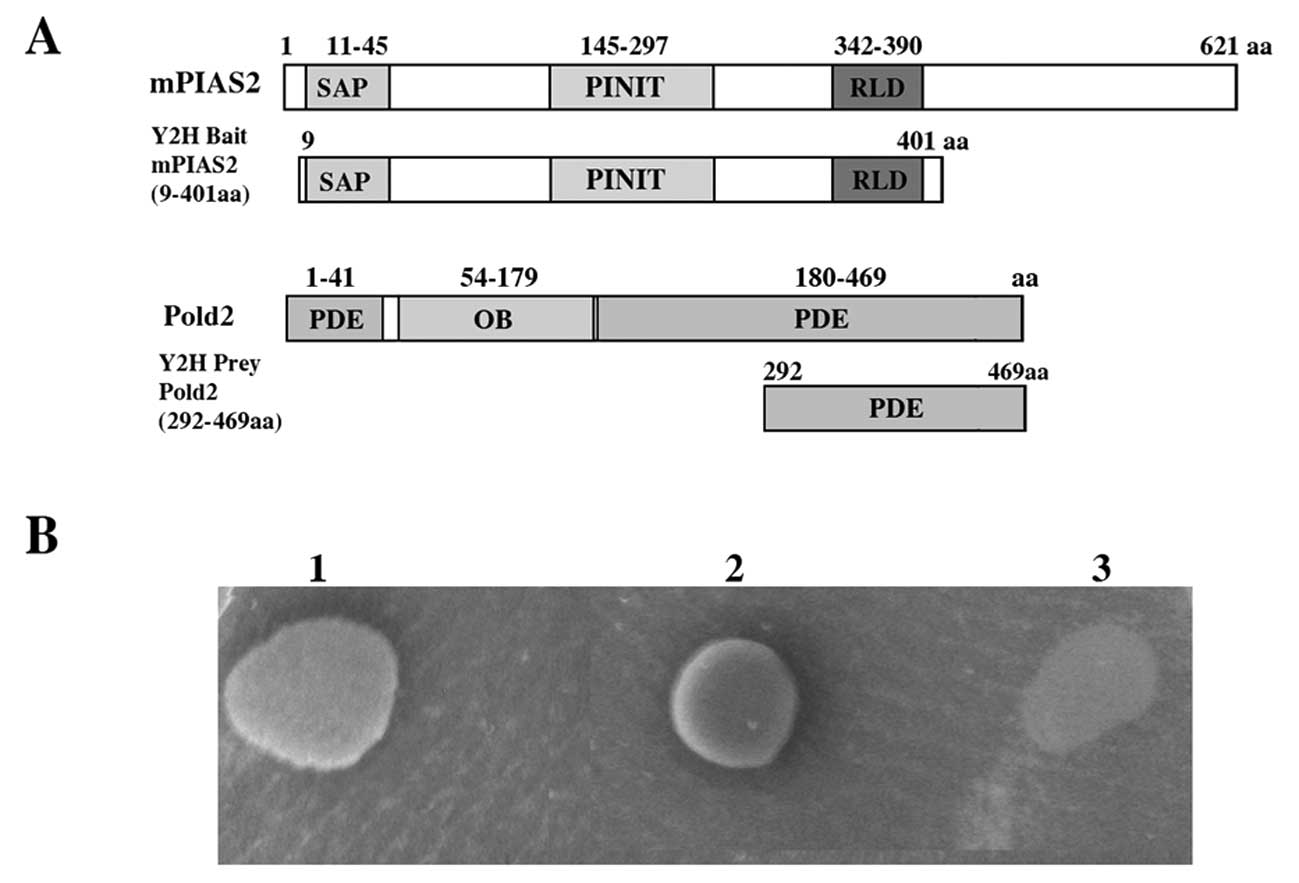
The common cDNA fragment of the mouse PIAS2 gene
[9-401 aa corresponding to nucleotides (nt) 213–1391 of NM_008602]
was cloned into the pGBKT7 vector containing the GAL4 DNA-binding
domain to generate the bait plasmid, pGBKT7-PIAS2. This construct
did not show toxic effects or autonomous transcriptional activation
following transformation into the yeast strain, Y187. The mouse
stem cell cDNA library was constructed in a pGADT7-Rec vector
containing a GAL4 activation domain using Matchmaker Library
Construction and Screening kits (Clontech, Santa Clara, USA) and
then transformed into the yeast AH109 strain. Yeast two-hybrid
screening was performed using the Matchmaker Two-Hybrid system
(Clontech). Positive clones were selected based on their ability to
grow on synthetic dropout (SD) medium/-LTHA/X-α-Gal and for X-α-Gal
activity.
Clone isolation and interaction
analysis
Plasmid DNA of prey clones was isolated (Qiagen,
Hilden, Germany) and transformed into E. coli DH5α. Prey
clones were recovered by ampicillin resistance and cDNA inserts
were identified by PCR amplification, sequencing, and BLAST
alignment. Interaction between the bait and identified prey clones
was verified by co-transforming the purified prey plasmid with the
bait pGBKT7-PIAS2 construct into the yeast AH109 strain followed by
selection on SD/-LTHA medium. Co-transforming of pGBKT7-p53 with
pGADT7-SV40 was used as a positive control. A negative control was
also included by co-transforming of pGBKT7-p53 with pGADT7 vector
into the same yeast cells.
Expression constructs and antibody
preparation
The common cDNA fragment of the mouse PIAS2 gene was
amplified by PCR and cloned into the pCMV-Myc and pDsRed-Express-1
vector (Clontech) to generate pCMV-c-Myc-PIAS2 and
pDsRed-Express-1-PIAS2 respectively. Full-length Pold2 was cloned
into pEGFP-N1 (Clontech) to generate pEGFP-N1-Pold2.
The following commercially available antibodies were
used for immunoprecipitation and western blot analysis: monoclonal
anti-c-Myc antibody (Millipore Biotechnology, Bedford, MA, USA) and
rabbit anti-GFP polyclonal antibody (Epitomics, Burlingame,
USA).
Cell culture and transfection
HEK 293T and HeLa cells (human epithelial carcinoma
line) were cultured and maintained in DMEM (Invitrogen)
supplemented with 10% fetal bovine serum at 37°C in a humidified
atmosphere containing 5% CO2. Transient transfection of HEK 293T
and HeLa cells (75–90% confluence) was performed using
Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the
instructions provided by the manufacturer.
Immunoprecipitations
HEK 293T cells were transiently transfected with
pCMV-c-Myc-PIAS2 and pEGFP-N1-Pold2 constructs and maintained as
described above for 24 h. The cells were then washed with ice-cold
PBS and harvested in 500 ml lysis buffer [150 mM NaCl, 50 mM Tris
pH 8.0, 1% Nonidet P-40, 0.5% deoxycholate, and a protease
inhibitor mixture (Roche Applied Science, Mannheim, Germany)].
After incubation in lysis buffer for 60 min on ice, lysates were
clarified by centrifugation for 20 min at 14,000 rpm at 4°C.
Specific monoclonal antibodies (8 μg) were added to the
supernatant and incubated for 120 min at 4°C prior to addition of
50 μl protein G-agarose followed by an overnight incubation
at 4°C. Samples were then centrifuged for 1 min in a
microcentrifuge and washed with 1 ml lysis buffer, 1 ml washing
buffer 2 (500 mM NaCl, 50 mM Tris pH 7.5, 0.1% Nonidet P-40, 0.05%
deoxycholate) and 1 ml washing buffer (10 mM Tris pH 8.0, 0.1%
Nonidet P-40, 0.05% deoxycholate).
Western blot analysis
Initial lysates and immunoprecipitated proteins were
analyzed by SDS-PAGE and immunoblotting was carried out using
specific antibodies. Proteins were electrophoretically transferred
to nitrocellulose membranes (Amersham Biosciences, Amersham, UK)
and probed with the appropriate antibodies. Bound antibodies were
visualized with horseradish peroxidase (HRP)-conjugated anti-rabbit
or anti-mouse (Sigma) IgG antibodies using the enhanced
chemiluminescence system (ECL, Thermo Scientific, MA, USA).
Fluoresence microscope analysis
To detect PIAS2 and Pold2, HEK 293T/HeLa cells
plated on round coverslips were transiently transfected with the
pDsRed-Express-1-PIAS2 and pEGFP-N1-Pold2 constructs using
Lipofectamine 2000 reagent. After 48 h, cells were washed twice
with PBS and visualized by Nikon scanning laser confocal
microscope.
Results
Identification of Pold2 as a
PIAS2-interacting protein using a yeast two-hybrid screening
system
To identify mouse proteins that interact with PIAS2,
cDNA libraries derived from mouse stem cells were screened using a
yeast two-hybrid system with the bait plasmid pGBKT7-PIAS2. Growth
of yeast strain Y187 co-transformed with this construct and the
empty prey plasmid pGADT7 was not supported on SD/-LTHA medium
confirming that the bait was not auto-activated. A total of
2×106 primary transformants were screened, yielding over
54 positive clones, 10 of which demonstrated strong growth on
SD/-LTHA medium. These clones were isolated, sequenced and aligned
using the NCBI BLAST alignment search tool. One clone encoded Pold2
(DNA polymerase, δ 2 small subunit), spanning regions of 177 amino
acids (Pold2 292-469 aa) was found to interact with PIAS2 protein.
To confirm these results, the PIAS2-Pold2 interaction was further
investigated using a direct yeast hybrid screening system. The
yeast strain co-transformed with pGADT7-Pold2 and pGBKT7 could not
grow on SD/-LTHA medium and showed negative β-galactosidase
activity (Fig. 1B, clone 3).
While strong growth on SD/-LTHA medium was observed only in yeast
cells co-transformed with pGBKT7-PIAS2 and pGADT7-Pold2 (Fig. 1B, clone 1), indicating interaction
of Pold2 with PIAS2 in yeast.
Pold2 interacts with PIAS2 in vivo
The data obtained from the yeast two-hybrid screen
was confirmed by evidence of a direct interaction between PIAS2 and
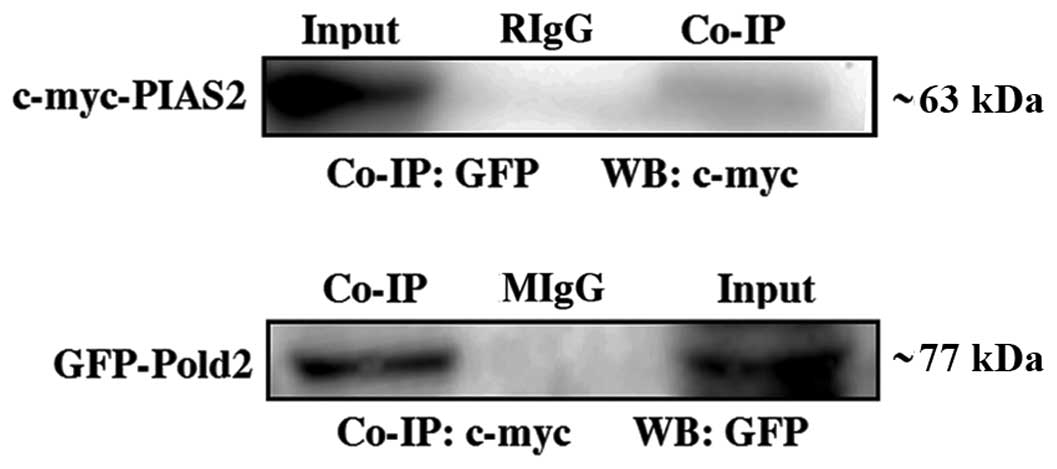
Pold2 in mammalian cells. The expression vectors for c-myc-tagged
PIAS2 and GFP-tagged Pold2 were co-transfected to human embryonic
kidney 293T cells. Twenty-four hours after transfection, the cell
extract was prepared, and the proteins in the extract were first
immunoprecipitated with the anti-c-myc, anti-GFP and non-specific
IgG, respectively. The precipitates were then immnuoblotted against
the anti-GFP or anti-c-myc antibodies, respectively (Fig. 2). The anti-c-myc antibody
precipitated c-myc-PIAS2. GFP-Pold2, on the other hand, was
detected in the immunoprecipitate with the ant-GFP antibody but not
with IgG (Fig. 2). PIAS2 protein
was also immunoprecipitated from the transfected HEK 293T cells
using a rabbit anti-GFP antibody prior to detection of PIAS2 by
anti-c-myc antibody. As shown in Fig.
2, Pold2 and PIAS2 fusion proteins were observed in the
co-immunoprecipiate and total lysate (input). These results
indicated that Pold2 can interact with PIAS2 in mammalian
cells.
Co-localization of Pold2 with PIAS2 in
HEK 293T and HeLa cells
To further confirm the interaction between Pold2 and
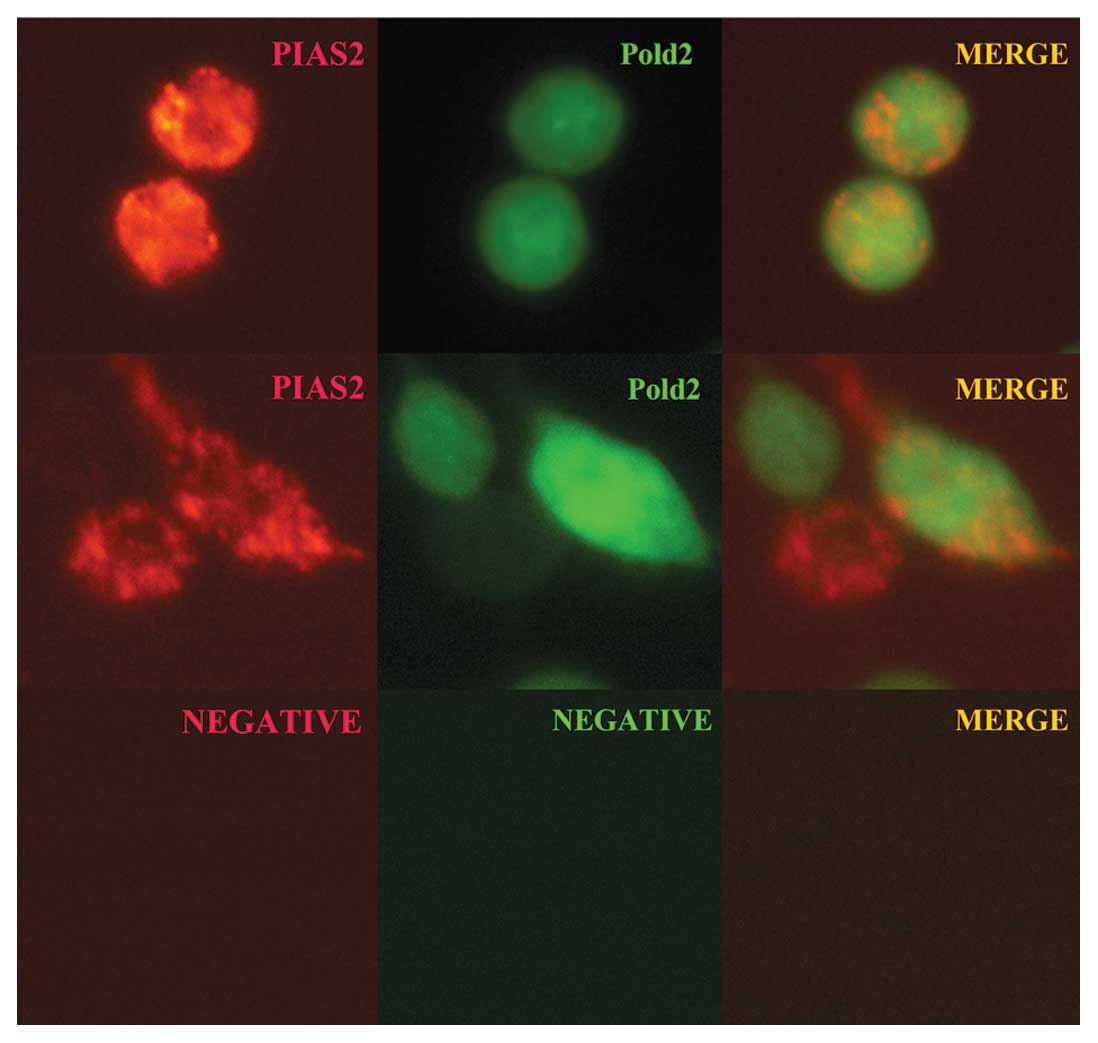
PIAS2 proteins, the co-localization was demonstrated by
co-transfected pDsRed-Express-1-PIAS2 and pEGFP-N1-Pold2 into HEK
293T and HeLa cells, respectively. Two days after transfection, the
cells were detected by a Nikon scanning laser confocal microscope.
As shown in Fig. 3, GFP-Pold2
protein was uniformly distributed in HEK 293T and HeLa cells
(Fig. 3, green) and DsRed-PIAS2
protein was expressed in these cells (Fig. 3, red). The merged image (Fig. 3, yellow) revealed that Pold2 and
PIAS2 were partially co-localized in these cells, indicating that
Pold2 can interact with PIAS2 in HEK 293T and HeLa cells.
Discussion
The purpose for this study was to identify novel
proteins that interact with mouse PIAS2 using the yeast two-hybrid
system. Human PIAS2 include PIASxα and PIASxβ while mouse PIAS2
gene has at least five isoforms (isoforms 1–5), which differ in the
N- and C-termini as a result of alternative splicing. So we used
the common encoding region of mouse PIAS2 as a bait to screen a
mouse cDNA library. As a result, Pold2 protein, a subunit of the
DNA polymerase δ complex (10),
was found to be a novel interaction partner for PIAS2.
Obviously, protein-protein interactions revealed by
the yeast two-hybrid system do not necessarily indicate that the
native proteins are capable of interacting, nor do they provide any
evidence that such interactions take place in a cellular context.
In this study, we have further characterized the interaction of
Pold2 with PIAS2.
The ability of Pold2 to interact with PIAS2 was
demonstrated by co-immunoprecipitation. For this purpose, both
tagged forms of Pold2 and PIAS2 eukaryotic expression vectors were
co-transfected into human embryonic kidney 293T cells. The whole
cell extracts were used for co-immunoprecipitation. The experiments
revealed that Pold2 and PIAS2 could be reciprocally
co-immunoprecipitated from total cell lysates of the transfected
HEK 293T cells. These studies strongly support the view that the
interaction of Pold2 with PIAS2 is physiological, i.e. that this
interaction takes place in a cellular context.
By using an overexpression system, we further
provided evidence of a direct interaction between GFP-Pold2 and
DsRed-PIAS2. Microscopy showed that Pold2 protein was distributed
in the cytoplasm and nuclei of HEK 293T and HeLa cells, and PIAS2
protein expression was scattered in these cells. The merged image
revealed that Pold2 and PIAS2 were partial co-localized in these
cells. These results suggested that Pold2 can interact with PIAS2
in mammalian cells.
Mouse Pold2 was cloned from a mouse cDNA embryo
library by PCR (25). It was
located on chromosome 11, band A2 and encoded a 469 amino acid
protein. The homology of the mouse protein with human and bovine
homologues was extremely high and the interaction of the 125-kDa
subunit and the 50-kDa subunit of Polδ was also highly conserved
between mouse and human (25).
The crystal structure showed that there are three domains in Pold2:
two phosphodiesterase-like (PDE) domains and one
oligonucleotide/oligosaccharide binding (OB) domain. The flanking
regions of Pold2 PDE provide a larger surface area and the
flexibility to form many protein-protein interactions (26). In our screening, the C-terminal
PDE domain of Pold2, which spanned from 292 to 469 amino acids, was
found to interact with PIAS2. It has also been reported that there
were many proteins interacting with Pold2 protein and fulfilling
their function. One of them is p21Cip/WAF1/Sdi1 (p21).
p21 was the first cyclin-dependent kinase inhibitor (CKIs) to be
identified (27,28). The N-terminal of p21 can bind to
and inhibit the cyclin-dependent protein kinases (CDKs) which are
necessary for G1 to S phase transition as well as S phase
progression (29). A second
cyclin binding site has also been identified near the C terminus of
p21. The C-terminal region of p21 was also shown to bind
proliferating cell nuclear antigen (PCNA) and to inhibit DNA
replication mediated by Polδ (30). p21 was also found to interact with
the small subunit of Polδ, Pold2 (11). This ability indicated that p21 may
be recruited to the replication complex via an interaction with
Pold2. This interaction could provide the basis for the targeting
or anchoring of p21 to the replication complex, thereby
facilitating the actions of p21. The latter could include several
possibilities, i.e., the inhibition of Polδ by binding of p21 to
PCNA within the replication complex, or by inhibition of
phosphorylation of replication complex proteins by the CDK2/cyclin
A kinase (11).
As a member of PIAS2, Miz1 was a mutant PIASxβ with
deletion of amino acids 1–133. This mutant protein was initially
described as a protein that interacts with the carboxy terminus of
the Myc oncoprotein (31). Miz1
was able to activate transcription of genes encoding the cell cycle
inhibitors p21, leading to cell cycle arrest (31). Induction of p21 expression
required binding of Miz1 to the core p21 promoter and, as a
consequence, factors that control Miz1 could affect p21 expression
(32). Miz1 could also repress
transcription when it formed complexes with other transcription
factors. For instance, Miz1 could repress transcription of the p21
gene in a complex with the Myc oncoprotein. Myc repressed
transcription of the p21 gene through binding to Miz1. As a result,
high levels of Myc suppressed cell cycle arrest and favoured
apoptosis in response to DNA damage (32). At its amino-terminus, Miz1
contained a POZ protein-protein interaction domain, which can
mediate both homo and heterodimerization among POZ domain proteins.
Miz1 heterodimerized with Zbtb4 repressed p21 expression and
inhibits cell cycle arrest in response to p53 activation (33). In this study, we found that Pold2
could interact with PIAS2. The yeast-two hybrid bait also contained
the full length of Miz1 POZ domain. So there was a possibility that
PIAS2 could heterodimerize with Pold2 and regulated the function of
p21. The latter could be recruited to the Polδ complex and thereby
modulate signaling that could affect Polδ activity. Further study
is required to provide more information and evidence for
understanding the functional significance of the Pold2-PIAS2
interaction.
In conclusion, evidence was presented here to
demonstrate there was a direct interaction between PIAS2 and Pold2,
the small subunit of Polδ both in vitro and in vivo.
Although its functions were currently unknown, these findings
indicated that many proteins might be involved the action of
Polδ.
Acknowledgements
This study was supported by grants
from the National Natural Science Foundation of China (no. 31071020
and no. 81000272) and the Natural Science Foundation of Jiangsu
Province of China (no. BK2009192).
References
|
1.
|
U HubscherM GiovanniS SpadariEukaryotic
DNA polymerasesAnnu Rev
Biochem71133163200210.1146/annurev.biochem.71.090501.150041
|
|
2.
|
MD SuttonGC WalkerManaging DNA
polymerases: coordinating DNA replication, DNA repair, and DNA
recombinationProc Natl Acad Sci
USA9883428349200110.1073/pnas.11103699811459973
|
|
3.
|
SP BellA DuttaDNA replication in
eukaryotic cellsAnnu Rev
Biochem71333374200210.1146/annurev.biochem.71.110601.13542512045100
|
|
4.
|
MY LeeCK TanKM DowneyAG SoFurther studies
on calf thymus DNA polymerase δ purified to homogeneity by a new
procedureBiochemistry23190619131984
|
|
5.
|
SA MacNeillS MorenoN ReynoldsP NursePA
FantesThe fission yeast Cdcl protein, a homologue of the small
subunit of DNA polymerase 8, binds to Pol3 and Cdc27EMBO
J154613462819968887553
|
|
6.
|
YI PavlovPV ShcherbakovaIB RogozinRoles of
DNA polymerases in replication, repair, and recombination in
eukaryotesInt Rev
Cytol25541132200610.1016/S0074-7696(06)55002-817178465
|
|
7.
|
R MossiU HubscherClamping down on clamps
and clamp loaders-the eukaryotic replication factor CEur J
Biochem25420921619989660172
|
|
8.
|
Z KelmanPCNA: structure, functions and
interactionsOncogene14629640199710.1038/sj.onc.12008869038370
|
|
9.
|
B XieN MazloumL LiuA RahmehH LiMY
LeeReconstitution and characterization of the human DNA polymerase
delta four-subunit
holoenzymeBiochemistry411313313142200210.1021/bi026270712403614
|
|
10.
|
H LiB XieY ZhouA RahmehS TrusaS ZhangY
GaoEY LeeMY LeeFunctional roles of p12, the fourth subunit of human
DNA polymerase δJ Biol Chem2811474814755200616510448
|
|
11.
|
H LiB XieA RahmehY ZhouMY LeeDirect
interaction of p21 with p50, the small subunit of human DNA
polymerase deltaCell
Cycle5428436200610.4161/cc.5.4.242516479163
|
|
12.
|
H HeCK TanKM DowneyAG SoA tumor necrosis
factor α- and interleukin 6-inducible protein that interacts with
the small subunit of DNA polymerase δ and proliferating cell
nuclear antigenProc Natl Acad Sci USA9811979119842001
|
|
13.
|
L LiuEM Rodriguez-BelmonteN MazloumB XieMY
LeeIdentification of a novel protein, PDIP38, that interacts with
the p50 subunit of DNA polymerase δ and proliferating cell nuclear
antigenJ Biol Chem2781004110047200312522211
|
|
14.
|
AM SzekelyYH ChenC ZhangJ OshimaSM
WeissmanWerner protein recruits DNA polymerase δ to the
nucleolusProc Natl Acad Sci USA971136511370200011027336
|
|
15.
|
B LiuJ LiaoX RaoSA KushnerCD ChungDD
ChangK ShuaiInhibition of Stat1-mediated gene activation by
PIAS1Proc Natl Acad Sci
USA951062610631199810.1073/pnas.95.18.106269724754
|
|
16.
|
N KotajaS AittomäkiO SilvennoinenJJ
PalvimoOA JänneARIP3 (androgen receptor-interacting protein 3) and
other PIAS (protein inhibitor of activated STAT) proteins differ in
their ability to modulate steroid receptor-dependent
transcriptional activationMol
Endocrinol1419862000200010.1210/mend.14.12.0569
|
|
17.
|
CD ChungJ LiaoB LiuX RaoP JayP BertaK
ShuaiSpecific inhibition of Stat3 signal transduction by
PIAS3Science27818031805199710.1126/science.278.5344.18039388184
|
|
18.
|
C ZhangX YuanL YueJ FuL LuoZ YinPIASy
interacts with p73alpha and regulates cell cycle in HEK293
cellsCell
Immunol263235240201010.1016/j.cellimm.2010.04.00520471636
|
|
19.
|
AD SharrocksPIAS proteins and
transcriptional regulation-more than just SUMO E3 ligases?Genes
Dev20754758200610.1101/gad.142100616600908
|
|
20.
|
K ShuaiRegulation of cytokine signaling
pathways by PIAS proteinsCell
Res16196202200610.1038/sj.cr.731002716474434
|
|
21.
|
T AroraB LiuH HeJ KimTL MurphyKM MurphyRL
ModlinK ShuaiPIASx is a transcriptional co-repressor of signal
transducer and activator of transcriptionJ Biol
Chem2782132721330200310.1074/jbc.C30011920012716907
|
|
22.
|
Y PengJ LeeC ZhuZ SunA novel role for
protein inhibitor of activated STAT (PIAS) proteins in modulating
the activity of Zimp7, a novel PIAS-like protein, in androgen
receptor-mediated transcriptionJ Biol
Chem2851146511475201010.1074/jbc.M109.07932720159969
|
|
23.
|
M GrossB LiuJ TanFS FrenchM CareyK
ShuaiDistinct effects of PIAS proteins on androgen-mediated gene
activation in prostate cancer
cellsOncogene2038803887200110.1038/sj.onc.120448911439351
|
|
24.
|
K TakahashiT TairaT NikiC SeinoSM
Iguchi-ArigaH ArigDJ-1 positively regulates the androgen receptor
by impairing the binding of PIASxα to the receptorJ Biol
Chem2763755637563200111477070
|
|
25.
|
R HindgesU HübscherCloning, chromosomal
localization, and interspecies interaction of mouse DNA polymerase
delta small subunit
(PolD2)Genomics444551199710.1006/geno.1997.48389286699
|
|
26.
|
AG BaranovskiyND BabayevaVG ListonIB
RogozinEV KooninYI PavlovDG VassylyevTH TahirovX-ray structure of
the complex of regulatory subunits of human DNA polymerase
deltaCell Cycle730263036200810.4161/cc.7.19.672018818516
|
|
27.
|
J HarperG AdamiN WeiK KeyomarsiS
ElledgeThe p21 Cdk-interacting protein Cip1 is a potent inhibitor
of G1 cyclin-dependent
kinasesCell75805816199310.1016/0092-8674(93)90499-G8242751
|
|
28.
|
A NodaY NingS VenableO Pereira-SmithJ
SmithCloning of senescent cell-derived inhibitors of DNA synthesis
using an expression screenExp Cell
Res2119098199410.1006/excr.1994.10638125163
|
|
29.
|
J ChenPK JacksonMW KirschnerA
DuttaSeparate domains of p21 involved in the inhibition of Cdk
kinase and PCNANature374386388199510.1038/374386a07885482
|
|
30.
|
JM GulbisZ KelmanJ HurwitzM O’DonnellJ
KuriyanStructure of the C-terminal region of p21(WAF1/CIP1)
complexed with human
PCNACell87297306199610.1016/S0092-8674(00)81347-18861913
|
|
31.
|
S AdhikaryM EilersTranscriptional
regulation and transformation by Myc proteinsNat Rev Mol Cell
Biol6635645200510.1038/nrm170316064138
|
|
32.
|
S HeroldM WanzelV BeugerC FrohmeD BeulT
HillukkalaJ SyvaojaHP SaluzF HaenelM EilersNegative regulation of
the mammalian UV response by Myc through association with Miz-1Mol
Cell10509521200210.1016/S1097-2765(02)00633-012408820
|
|
33.
|
A WeberJ MarquardtD ElziN ForsterS StarkeA
GlaumD YamadaPA DefossezJ DelrowRN EisenmanH ChristiansenM
EilersZbtb4 represses transcription of P21CIP1 and controls the
cellular response to p53 activationEMBO
J2715631574200810.1038/emboj.2008.8518451802
|