Introduction
Colon cancer is a common malignant disease worldwide
(1). The incidence of colon
cancer has increased by 2.1% each year (2), and is increasing in most countries,
particularly in developing countries. The mortality rate of
colorectal cancer is second only to that of lung cancer in men and
breast cancer in women and has shown little sign of decreasing in
the past 20–30 years (3). In most
parts of China, the incidence of colon cancer has experienced the
highest increase among malignant tumors.
It has been confirmed that colorectal cancer is
initiated by the accumulation of genetic mutations (4). Chromatin modification strongly
affects tumorigenesis due to the dynamic regulation of
transcription. The disorganized chromatin structure promotes
oncogenesis.
In parallel with the discovery of the importance of
chromatin modification in tumorigenesis was the finding that
butyrates are potent anticancer agents. Sodium butyrate (NaBu) is a
short-chain fatty acid (SCFA), which can influence a variety of
physiological functions. The anti-neoplastic activity of SCFAs such
as butyrates in colorectal carcinogenesis is produced as a
consequence of a high fiber diet (5). Butyrate, as one main end-product of
intestinal microbial fermentation, demonstrates the protective role
of dietary fiber in the maintenance of colonic homeostasis
(6). SCFAs induce cell cycle
arrest, differentiation and apoptosis in cancer cells, several also
have anti-inflammatory activities, and a number have progressed to
clinical trials. These agents, which include NaBu and
(R)-trichostatin A (TSA), have displayed anti-proliferative and
differentiating activity in a wide variety of cancers (7–9).
Research has shown that NaBu induces tumor cell apoptosis in colon,
breast, esophageal and prostate cancer (10). Although several SCFAs have been
identified as possible anticancer agents, in regards to colon
cancer, NaBu is a particularly attractive agent that may have both
preventative and therapeutic value. Applied with other chemotherapy
drugs, the curative effect was found to be significantly improved.
Therefore, some researchers speculate that NaBu may also activate
several tumor-suppressor genes and thus play a role in the
antitumor mechanism indirectly (11).
Chromatin remodeling plays a vital role in the
normal function of tumor suppressors, such as Rb and p53, and this
function becomes dysregulated in several types of cancer.
Furthermore, changes in DNA methylation are prominent
characteristics of most cancers, and it is now known that
methylated DNA recruits complexes that alter chromatin structure
and repress gene transcription (12). It was previously demonstrated that
the apoptosis-associated speck-like protein (ASC) is a p53-target
gene which regulates the p53-Bax mitochondrial apoptotic pathway.
ASC is also known to be a target of methylation-induced gene
silencing. It participates in retinoic acid-induced apoptosis and
forms tungsten filament structure in cells. ASC can be considered
as a tumor-suppressor gene involved in the regulation of apoptosis,
caspase-induced inflammation and NF-κB activation (13). The expression of ASC can be
regulated at the gene level, mainly by enhancer methylation. In
addition, the abnormal expression of ASC has been found in
leukemia, bile duct, colorectal and prostate cancer (14,15). Genetic modulations of apoptotic
genes affect the development, carcinogenesis, tumor growth,
chemoradiosensitivity and inflammatory response. Aberrant
hypermethylation in the promoter regions of tumor-suppressor genes
has been shown to be a mechanism for the inactivation of many
genes. This transcriptional silencing involves methylation of the
promoter associated CpG islands and changes in the local chromatin
structure resulting in tightly compacted chromatin that inhibits
access to transcription factors and thus in the silencing of that
gene (16).
We previously investigated colon cancer,
particularly the LS174T cell line and found the absence of ASC. We
proposed that this may be associated with cell apoptosis. The
mechanism of colorectal cancer incidence remains unclear, yet the
overexpression of oncogenes and inactivation of tumor-suppressor
genes may be involved. We used NaBu-treated human colorectal cancer
cell line LS174T and investigated the proliferative status,
transcription of the tumor-associated gene ASC as well as their
interaction and their effects on the apoptosis rate of colon cancer
cells to assess whether NaBu could be effectively applied in the
clinical treatment of colorectal cancer in the future.
Materials and methods
Chemicals
NaBu was obtained from Sigma-Aldrich (St. Louis, MO,
USA). For cell proliferation and in vitro enzyme inhibition
studies, NaBu was diluted to different concentrations with
phosphate-buffered saline (PBS) and stored at 4°C until use.
Cell culture
Human colon cancer-derived cell line LS174T was
maintained in our laboratory. LS174T cells were maintained in
RPMI-1640 supplemented with 100 ml/l heat-inactivated fetal bovine
serum, 100 U/ml penicillin and 100 μg/ml streptomycin at
37°C in a 50 ml/l CO2 incubator.
Western blotting of ASC protein
LS174T cells were cultured with (5 mmol/l) or
without NaBu for 12, 24, 48 and 72 h or with 1, 5 and 10 mmol/l for
24 h. Cells were washed twice with ice-cold PBS, scraped and
transferred to tubes, and then lysed in 1 ml buffer A (10 mmol/l
HEPES, pH 7.4, 10 mmol/l KCl, 1.5 mmol/l MgCl2, 0.5
mmol/l DTT, 0.2 mmol/l PMSF, 1 μg/ml protease inhibitors,
0.25 g/l NP-40) for 15 min with rotation at 4°C. The nuclear pellet
was resuspended in 100 μl buffer B (20 mmol/l HEPES, pH 7.4,
420 mmol/l NaCl, 1.5 mmol/l MgCl2, 0.2 mmol/l EDTA, 250
ml/l glycerol, 0.5 mmol/l DTT, 0.2 mmol/l PMSF, 1 μg/ml
protease inhibitors) for 30 min, and the soluble nuclear protein
was collected by centrifugation. Fifty micrograms of nuclear
extracts was boiled in loading buffer (125 mmol/l Tris-HCl, pH 6.8,
40 g/l SDS, 200 g/l glycerol, 0.05 g/l bromphenol blue) for 5 min
and then loaded onto a 150 g/l SDS-polyacrylamide gel. After
electrophoresis, proteins were transferred onto nitrocellulose
membrane (0.45 μm). The following antibody was used: goat
polyclonal antibody against rabbit IgG (Santa Cruz Biotechnology,
Inc.). The binding of the antibody was detected using an ECL system
(Sigma-Aldrich), and membranes were then exposed to Kodak BioMax
film for 1 min. Antibody against β-actin (Sigma-Aldrich) was used
for the western blotting as a control for protein
concentration.
Reverse transcription-polymerase chain
reaction (RT-PCR) analysis
Cells were placed in 25 cm2 flasks at a
density of 2×106 cells/flask. After 24 h, cells were
exposed to complete medium in the presence (5 mmol/l) or absence of
butyrate for 1 day. Total RNA was extracted by the RNAiso reagent
(Takara). Reverse transcription was performed on 2 μg total
RNA in a reaction volume of 20 μl with 2 μl of 10X RT
buffer, 4 μl MgCl2 (25 mmol/l), 2 μl dNTP
mix (10 mmol/l), 0.5 μl RNAse inhibitor, 2 μl
dithiothreitol (0.1 mol/l), 1 μl AMV reverse transcriptase
(5 U/μl) and 1 μl downstream primer (50
μmol/l). The volume was then adjusted to 20 μl with
distilled water. PCR amplification was then performed with 10
μl cDNA solution supplemented with 10 μl of 10X PCR
buffer, 0.5 μl of upstream primers (50 μmol/l), 0.25
μl Ex Taq HS (Takara) and water to a final volume of 50
μl. PCR conditions consisted of 35 cycles for β-actin
primers 5′-cct tcc tgg gca tgg agt cct g-3′ (sense) and 5′-gga gca
atg atc ttg atc ttc-3′ (antisense); 35 cycles for ASC primers
5′-tgg gcc tgc agg aga tg-3′ (sense) and 5′-att tgg tgg gat tgc
cag-3′ (antisense). Reactions were initiated at 95°C for 5 min, and
each PCR cycle consisted of 30 sec at 95°C, 1 min at 55°C and 1 min
at 72°C, followed by one cycle of 60°C for 10 min.
MTT assay
Cell proliferation was determined by the MTT
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay.
Briefly, LS174T cells were placed at a density of 104
cells/well in a 96-well plate (Costar, Cambridge, MA, USA)
overnight, and different concentrations of NaBu were added to each
well. After the preset culture duration, medium was replaced with
50 μl of 1 mg/ml MTT (Sigma-Aldrich) solution and the plate
was incubated for another 4 h. The dark-blue formazan crystal
precipitate reduced from the yellow soluble MTT by viable cells was
dissolved in 100 μl of DMSO. The absorbance was read at 595
nm on an enzyme linked immunosorbent assay (ELISA) plate reader
(Bio-Red Instruments, Inc.). Proliferation survival rate = (OD
value of experimental hole − OD value of blank hole/OD value of
control hole − OD value of blank hole) × 100%.
DAPI staining
The LS174T cells were incubated in 15-cm2
cell culture flasks for 24 h. Then the cells were treated with
different concentrations of NaBu for the specified times as
described above. The cells were washed with 60 μl/well of
PBS for 3 times, 5 min each time. The cells were then fixed with 60
μl/well of 4% PFA (paraformaldehyde) for 20 min. The PFA was
washed out with PBS. Cells were permeabilized with 60
μl/well of 0.1% Triton X-100 for 10 min. The washes were
repeated 3 times. Incubation was carried out for 15 min at room
temperature, then rinsed 3 times in PBS, and mounted. Finally, the
cells were photographed under a fluorescence microscope.
Annexin V-FITC/PI assay
To detect apoptosis, the cells were stained with PI
and fluorescein isothiocyanate (FITC)-conjugated Annexin V using
the Annexin V-FITC Apoptosis Detection Kit I (Beyotime, China).
Annexin V-FITC identifies cells in early apoptosis by detecting
externalized phosphatidylserine and PI identifies cells that have
lost plasma membrane integrity (i.e., necrotic or late apoptotic
cells). LS174T cells were seeded at a density of 0.5×106
cells/ml/well and treated with NaBu diluted with RPMI-1640 medium
for the times indicated at 37°C in an incubator with a humidified
atmosphere containing 5% CO2. After incubation, 1 ml of
the cell suspension was transferred to a 15-ml tube and 1 ml of
cold PBS was added for washing the cells. Centrifugation was
carried out at 500 × g for 5 min at 4°C. Cells were re-suspended in
binding buffer and 2.5 ml of FITC-labeled Annexin V and 2.5 ml of
PI solution were added. Then tube was kept on ice for 10 min and
then subjected to flow cytometry (Becton-Dickinson, USA). Cells
were gated on the basis of their forward and side light scatter
with any cell debris excluded from analysis. The gated cells were
then plotted for Annexin V-FITC and PI staining in a two-way dot
plot to assess the percentage of apoptotic LS174T cells.
Results
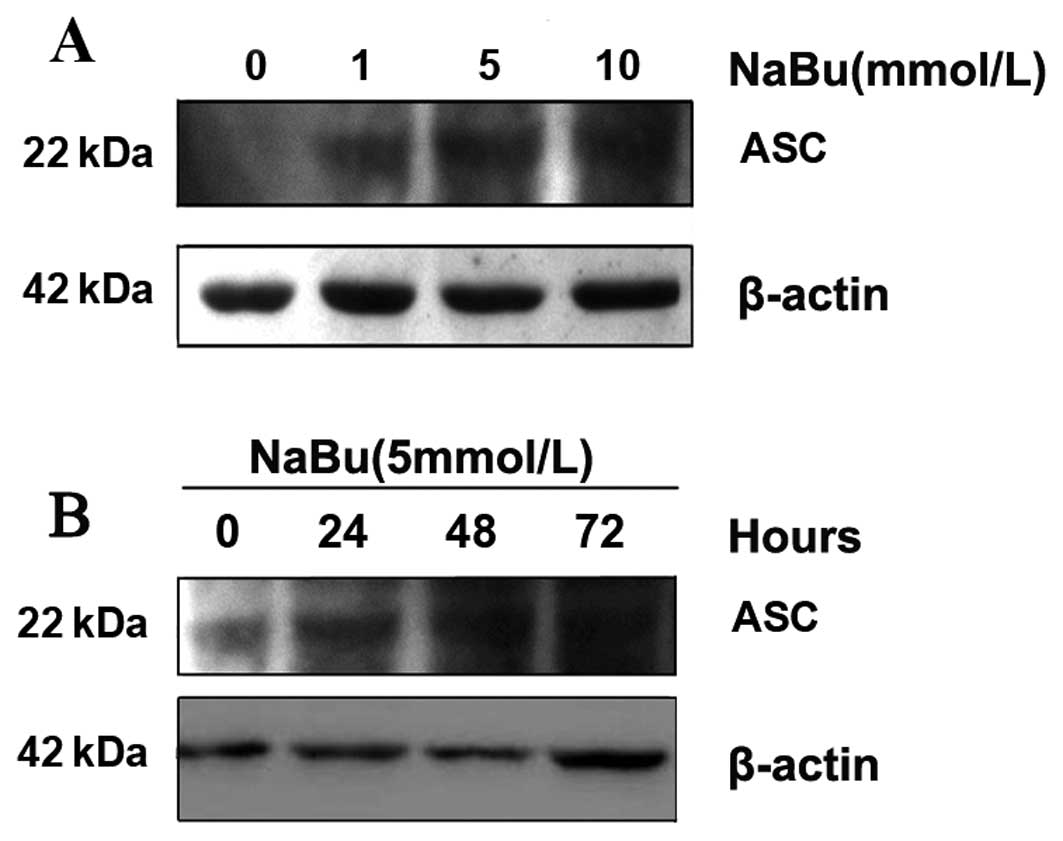
Sodium butyrate induces re-expression of
ASC protein
To examine the change in ASC expression in response
to the different dosages of NaBu and exposure times, we initially
detected the ASC protein by using western blotting. As the results
showed, following treatment with different dosages of NaBu, the
re-expression of ASC protein increased along with the gradually
increased dosages (Fig. 1A). When
1 mmol/l of NaBu was applied to the cells for 24 h, the
re-expression of ASC protein was very weak and blurred. Expresstion
was more evident when cells were exposed to 5 mmol/l and
significant at 10 mmol/l NaBu when compared with the control groups
(Fig. 1A). When using the same
concentration of NaBu (5 mmol/l) with different exposure times, the
re-expression of ASC protein increased along with prolonged time
exposure (Fig. 1B). The results
showed that the expression of ASC increased in a time- and
dose-dependent manner when exposed to NaBu.
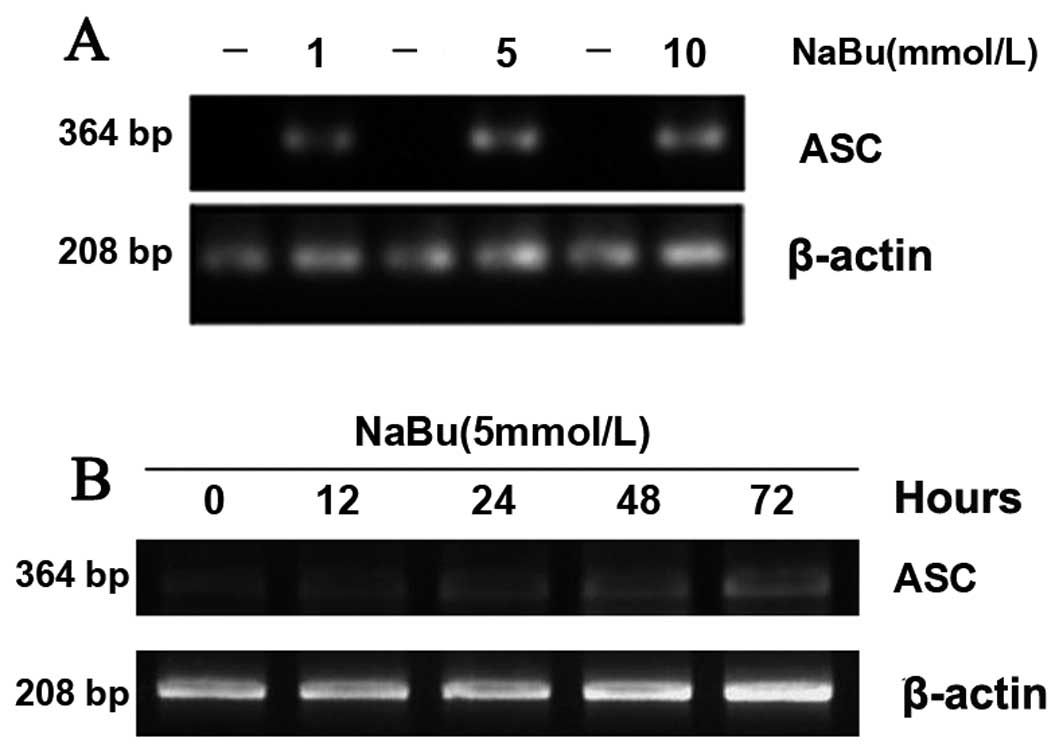
Sodium butyrate upregulates the
expression level of ASC mRNA
We next determined ASC mRNA using RT-PCR. The
expression level of ASC mRNA was upregulated following the
administration of different dosages of NaBu and increased with
gradually increasing dosages (Fig.
2A). We then used the same concentration of NaBu (5 mmol/l)
with different exposure times. The expression level of ASC mRNA was
upregulated along with the time duration (Fig. 2B). The results showed that the
mRNA expression of ASC increased in a time- and dose-dependent
manner when exposed to NaBu.
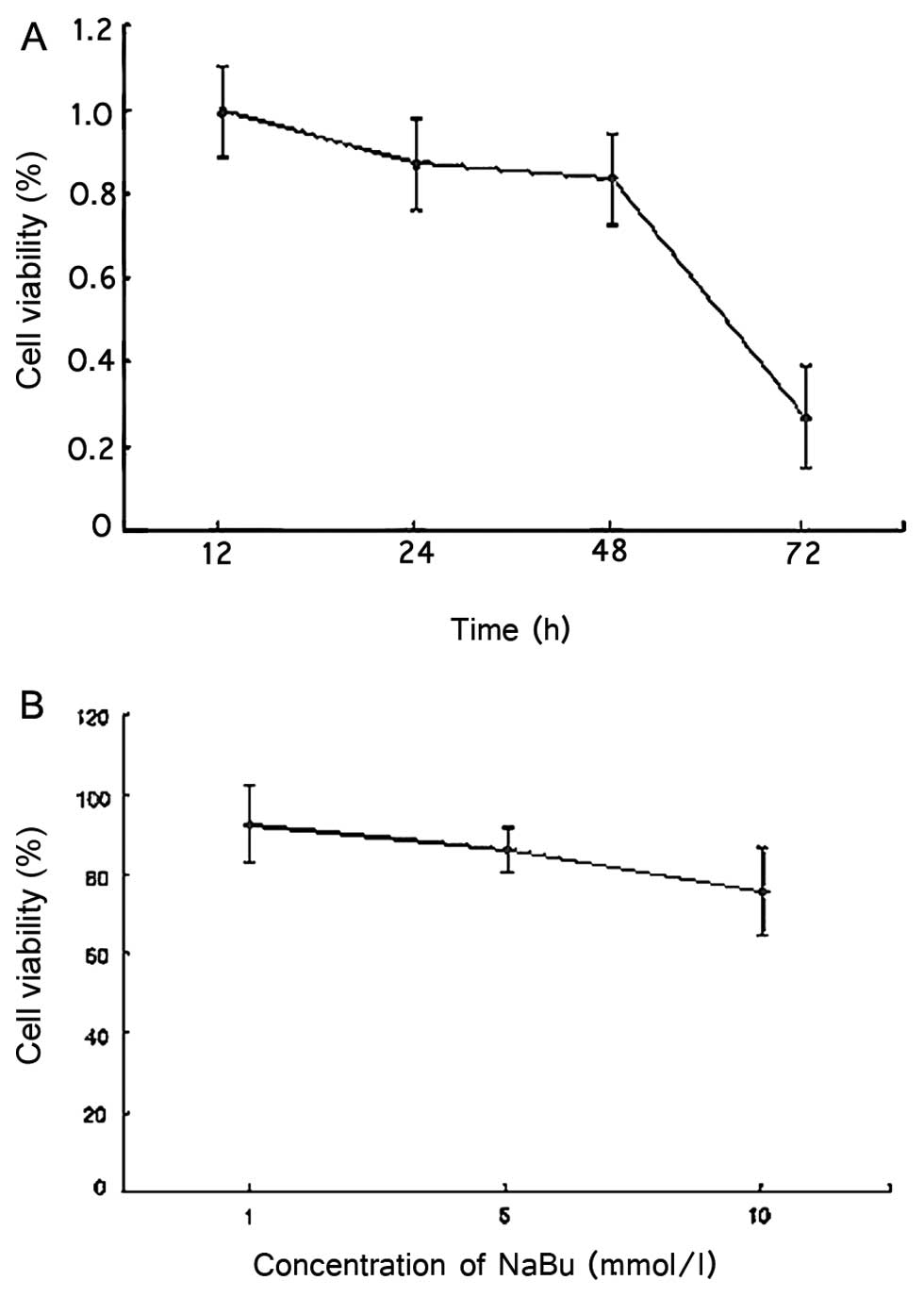
Effect of NaBu on the proliferation rate
of LS174T cells
ASC is a tumor-suppressor gene. Upregulated
expression of ASC inhibits the proliferation of tumor cells or
promotes apoptosis. Generally, a variety of genes in the genome are
re-expressed by NaBu. Thus, increasing the dosages of NaBu may
enhance the possibility of the re-expression of ASC and a reduction
in cell proliferation. We exposed LS174T cells to different dosages
of NaBu for 24 h (Fig. 3B). The
cell proliferation was determined using an MTT assay. The results
showed that cell proliferation rates were decreased by 7.57, 13.81
and 24.50% in the presence of 1, 5 and 10 mmol/l NaBu, respectively
(each group repeat 8 times). When cells were treated with the same
concentration (5 mmol/l) of NaBu (Fig. 3A), the cells proliferation rates
decrease progressively along with the time exposure. The
proliferation rates decreased by 0.56, 13.11, 16.62 and 73.17% at
12, 24, 48 and 72 h. The above results indicated an inverse
relationship between the cell proliferation rate and the exposure
time and the drug concentration.
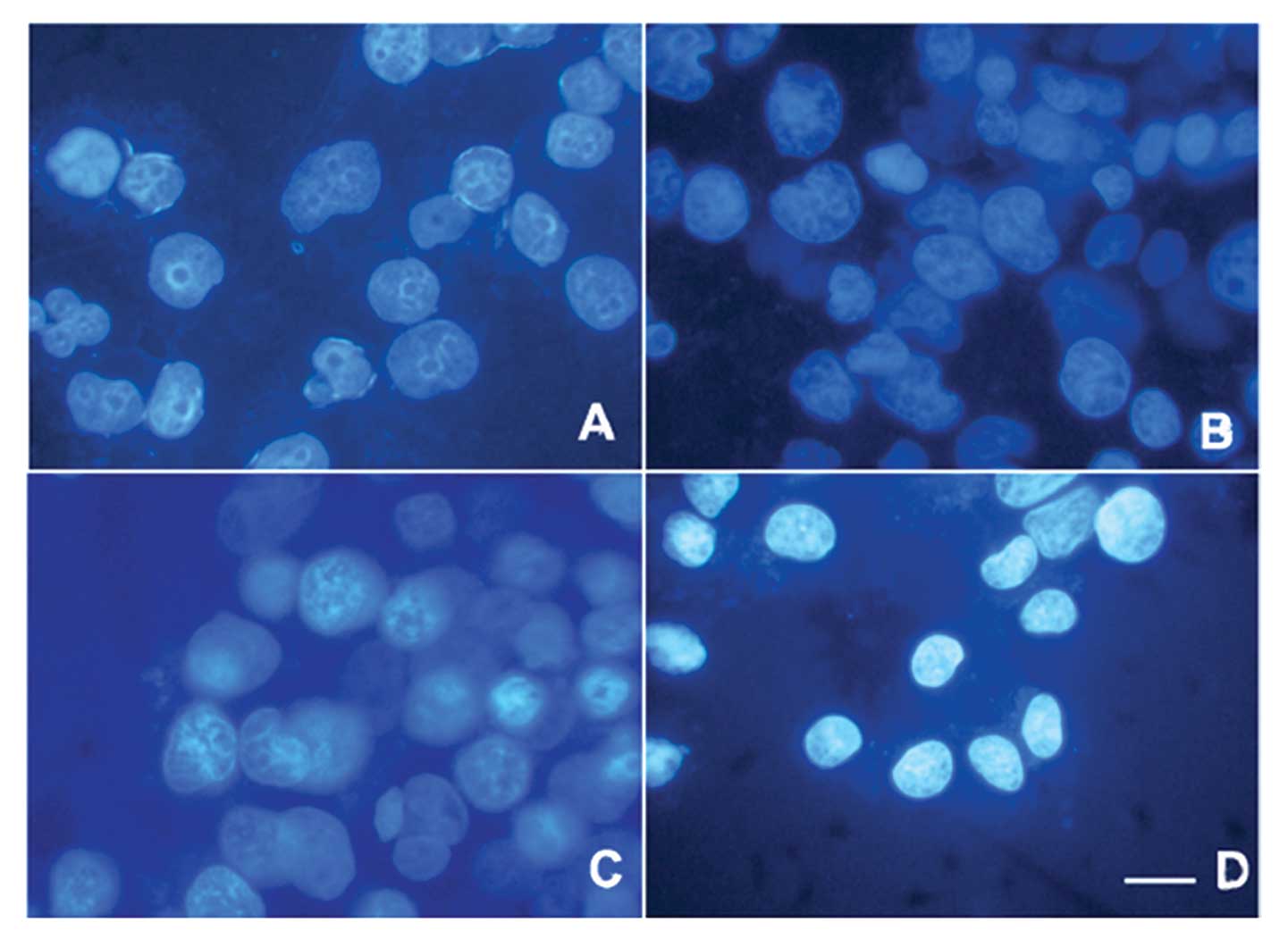
Morphological changes in the LS174T cells
as a result of NaBu-induced apoptosis
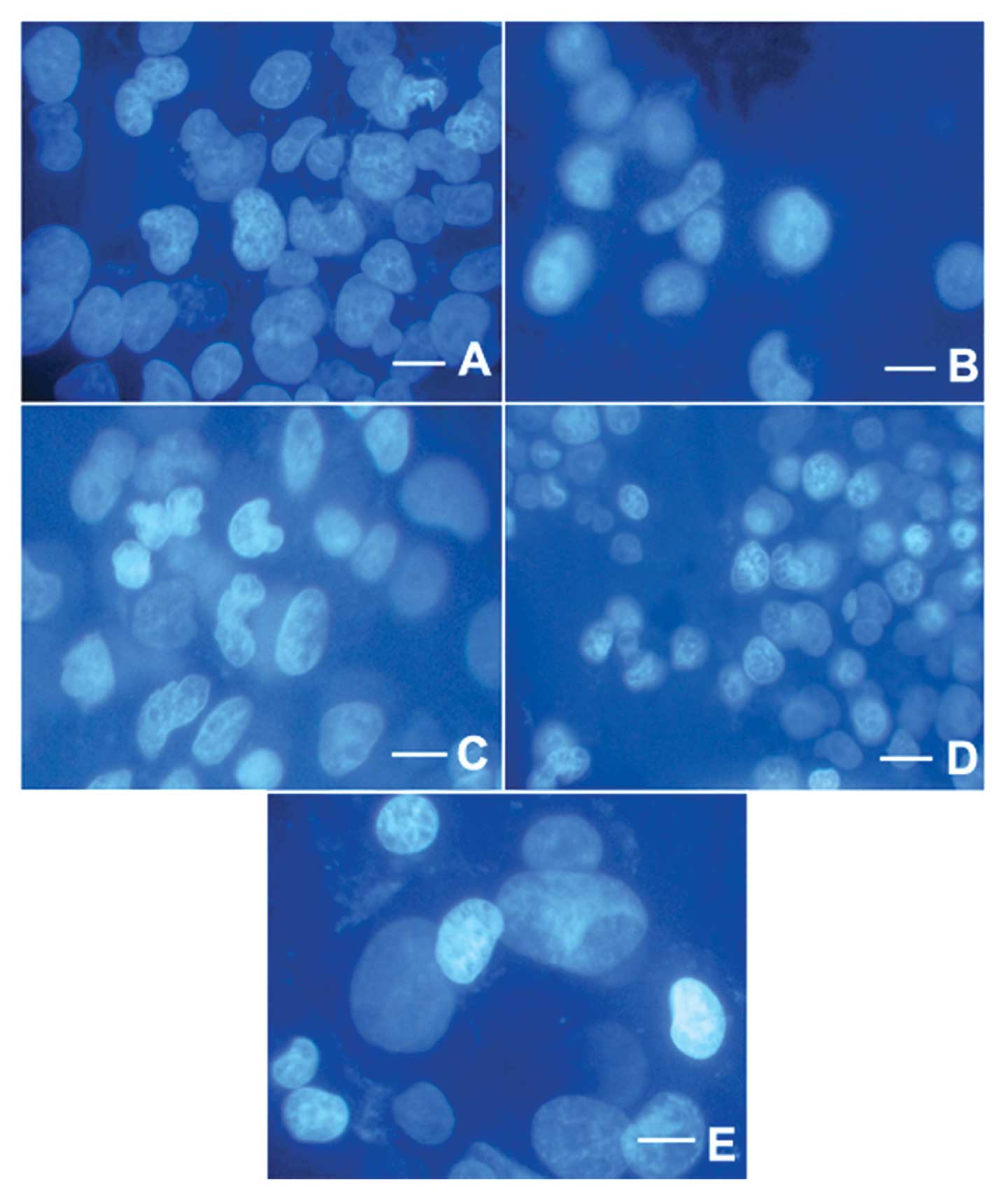
The apoptotic morphology in cellular bodies and
chromatin condensation were confirmed by DAPI staining (Figs. 4 and 5). Structures of cells in the control
group were complete and clear (Figs.
4 and 5). In contrast, in the
NaBu-treated groups, the cell nuclei became shrunken and some began
to be disrupted or exhibited an abnormal nuclear shape. This
phenomenon appeared to become gradually more severe with increased
exposure time (Fig. 5B–E) as well
as with increased dosages (Fig.
4B–D).
Apoptosis assay
For further verification of the re-expression of
ASC, we applied an apoptosis assay to detect the apoptosis level
following treatment with NaBu. To identify apoptosis of the plasma
membrane, Annexin V-FITC/PI double staining in the cells was
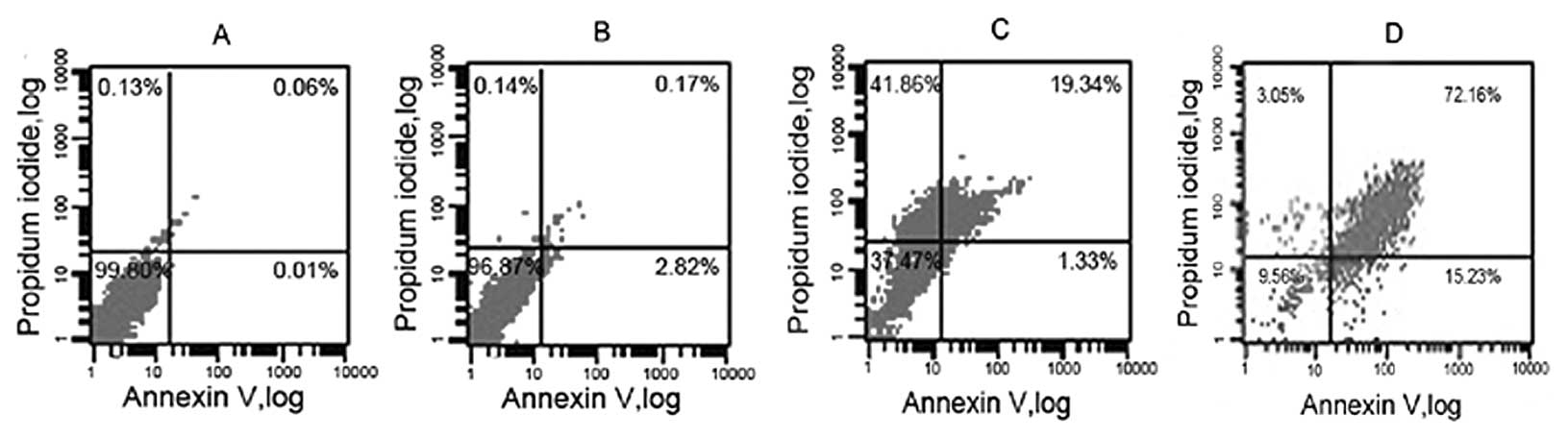
performed by flow cytometry. In the nonapoptotic viable control
cells, the Annexin V-FITC staining and PI-negative staining were
noted by dots in the bottom left quadrant (Fig. 6A). When cells were exposed to 1
mmol/l NaBu for 24 h, the change in the cell apoptosis rate was
slight (Fig. 6B). After exposure
of the cells to 5 mmol/l NaBu for 24 h, a small number of cells
showed Annexin V-FITC-positive and PI-negative staining, which
increased the number of dots in the bottom right quadrant from
0.01±1% in the control cells to 1.33±1% (Fig. 6C). The cells in this stage of
apoptosis were still viable. Following treatment with NaBu at 10
mmol/l, cells undergoing advanced apoptosis stained positive for
Annexin V-FITC and PI (upper right quadrant) and the numbers were
significantly augmented to 15.23±2% after 24 h (Fig. 6D). The population of cells
progressed to advanced apoptosis, indicating that the cells were no
longer viable. The relationship between cell apoptosis and NaBu
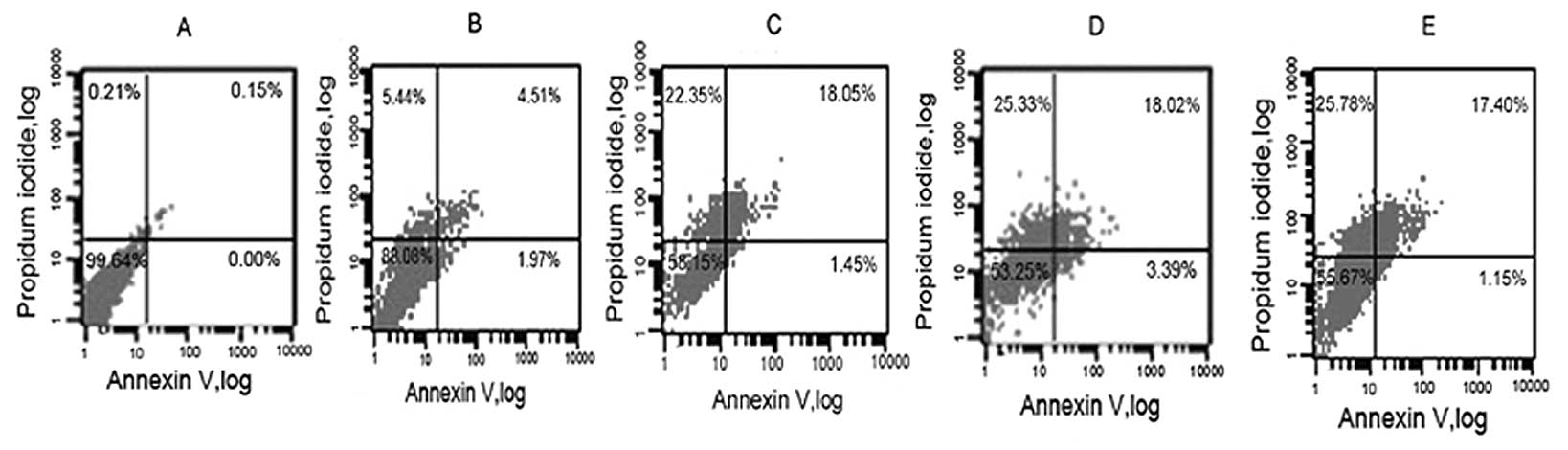
culture time was also investigated. We treated the cells with 5
mmol/l of NaBu for different exposure times. The results showed
that the percentage NaBu-treated cells undergoing early apoptosis
and secondary necrosis was increased with the increased exposure
time (Fig. 7). The rates of cell
apoptosis and secondary necrosis were increased to 6.48% at 12 h.
At 24 h the rate was 19.5%. The viability of the cells decreased
with an increase in exposure time, but the apoptosis rate did not
increased at 72 h compared to that of 48 h (Fig. 7D and E). Thus, the results showed
a time- and dose-dependent manner in the rates of cell apoptosis
following exposure to NaBu.
Discussion
The SCFA family of transcriptional co-repressors,
such as sodium butyrate (NaBu), induces cell cycle arrest, markers
of cell differentiation and apoptosis in colon cancer cell lines
in vitro (17). At the
same time, mechanisms responsible for the resistance of certain
cancers to the effect of chemotherapeutic agents are not completely
understood. In this research, we used NaBu to investigate the
re-expression of ASC in LS174T cells. We demonstrated that the
expression of ASC was obviously recovered following treatment with
NaBu, which induced the apoptosis of LS174T cells; the
proliferation rates decreased which further confirmed the
re-expression of ASC. This suggests the NaBu plays a role in the
reactivation of ASC expression and the latter promoted the
apoptosis of LS174T cells.
To investigate the association of transcriptional
modification, ASC, a p53-target gene which regulates the p53-Bax
mitochondrial apoptotic pathway, was introduced to our
experiment.
In the mitochondrial apoptosis pathway, activated
p53 promotes apoptosis through Bax, a molecule that stimulates the
release of cytochrome-c from mitochondria, activating procaspase-9
and resulting in cell death. Expression of regulators of the
p53-Bax mitochondrial apoptosis pathway, ASC/TMS1, can become
inactivated by promoter methylation leading to the disruption of
the pathway. In addition, ASC-activated caspase-8 can further
cleave the cytosolic Bid protein, which can translocate to
mitochondria and act as an activator of the mitochondrial apoptotic
pathway. Accumulating evidence shows a wide array of functions for
TMS1/ASC, from triggering apoptosis to regulating the activity of
NF-κB to activating inflammatory caspases (18). Aberrant NF-κB activity can promote
tumorigenesis by stimulating the proliferation of tumor cells,
enhancing the angiogenic and metastatic potential of tumors and
inhibiting apoptosis (19). It
was suggested that ASC is a tumor-suppressor gene as noted by its
pro-apoptotic function. Induced expression of ASC inhibits cell
proliferation and results in DNA fragmentation in a time-dependent
manner. It was reported that the methylation-mediated silencing of
the TMS1 gene is accomplished through the binding protein MBDs
(methyl-CpG binding proteins) (20). CpG island methylation has been
shown to be essential for normal development, X-chromosome
inactivation, imprinting and the suppression of parasitic DNA
sequences (21,22). Gene silencing associated with the
aberrant methylation of promoter region of CpG islands is an
acquired epigenetic alteration that serves as an alternative to
genetic events in the inactivation of tumor suppressor and other
genes in human cancers.
NaBu has been applied in cellular and molecular
research (23,24), particularly in antitumor research
(25–28). It plays a vital role in vitamin
D-induced apoptosis through PTEN upregulation, which has potential
benefit in gastric cancer therapy (29). Furthermore, NaBu activates Notch1
signaling, reduces tumor markers (e.g. ASCL1 and CgA) and induces
cell cycle arrest significantly, alters tumor cell proliferation
and apoptosis in pheochromocytoma, which were indicated by the
levels of cyclin D1, p21, cleaved PARP and cleaved caspase-3
proteins (30). NaBu also
increases expression of the coxsackie and adenovirus receptor in
colon cancer cells (31).
Although some researchers have reported that NaBu induces human
colon carcinoma HT-29 cell apoptosis through a mitochondrial
pathway (32), we found a new
relational molecular, ASC, whose expression was recovered in LS174T
cells.
In this study, we investigated the effect of NaBu on
ASC expression and the tumor proliferation in LS174T cells.
According to the results, the expression of ASC increased in a
time- and dose-dependent manner when exposed to NaBu. The cell
proliferation rates decreased and the apoptosis rates
correspondingly increased. Heerdt et al proposed that the
apoptosis noted may be the result of the effect of NaBu on
mitochondria (33). It was
established that mitochondrial activities are essential for the
initiation of butyrate-induced apoptosis as well as cell cycle
arrest in colonic carcinoma cells (34). These resulted in changes to the
membrane potential of mitochondria and led to the activation of
caspases. Recent observations showed that apoptosis by NaBu was
dependent on the activation and translocation to the mitochondria
of the pro-apoptotic Bcl-2 family member Bid, also NaBu
downregulates expression of Bcl-2 in colon cancer cells (35). By regulating Bcl-2 the
mitochondrial membrane potential changes, the NaBu-induced
activation of the caspase cascade may be an important part of
apoptosis induction.
NaBu can directly inhibit cell growth and
differentiation, promote apoptosis or inhibit tumor growth by
inhibiting angiogenesis indirectly. Chopin et al (36) showed that NaBu-induced apoptosis
in MCF-7 cells was independent of the G1-phase blockage, suggesting
that the underlying mechanisms of cell cycle blockage and induction
of apoptosis occurs through different pathways. At present, it is
well established that NaBu can initiate apoptosis through several
different pathways. Nonetheless, all primarily execute cell death
through activation of the intrinsic mitochondria-mediated pathway.
For example, an early p53-dependent and a late p53-independent form
of cell death have been associated with exposure to these
compounds. Although most studies have focused on its induction of
G1 cell cycle arrest, butyrate also induces a G2 arrest and the
present study was undertaken to examine this aspect of butyrate
action as a sensitive G2 cell cycle checkpoint and the
consequential occurrence of aberrant mitosis (37).
NaBu and other SCFAs are naturally occurring
products produced by fiber fermentation in the colon and have been
found to inhibit the growth of colon cancer cells both in
vivo and in vitro (38). In addition, when SCFAs were
applied in combination with a variety of chemotherapy drugs, better
co-treatment efficacy was noted. Yang et al (39) reported that the combination of
using DNA methyltransferase inhibitors and SCFAs can reactivate
MLH1, TIMP3, CDKN2B, CDKMN2A, ARHI and other tumor-suppressor
genes. These events promoted apoptosis in tumor cells. Nimmanapalli
et al (40) found that the
combination of using SAHA with Gleevec enhanced chemotherapy
sensitivity of Gleevec-resistant chronic myeloid leukemia cells.
Compared to transformed cells, normal cells are not sensitive to
SCFAs. NaBu causes changes in gene expression and the regulation of
proteins. Furthermore, it alters physiological functions of
drug-induced transformed cells (41,42).
Co-treatment of NaBu with a variety of chemotherapy
drugs may be beneficially utilized in clinical practice. The
present study revealed its important clinical value. Research on
NaBu and ASC expression may help to develop a strategy to improve
drug safety and efficacy and significantly enhance the efficiency
of drugs applied in cancer therapy.
Acknowledgements
This study was supported by grants
from the Chinese National Natural Science Foundation Projects
(31270867) and the Chinese State Key Program in Basic Research
(2012CB822103).
References
|
1.
|
JJ SungJY LauKL GohWK Leungthe Asia
Pacific Working Group on Colorectal CancerIncreasing incidence of
colorectal cancer in Asia: implications for screeningLancet
Oncol11871876200510.1016/S1470-2045(05)70422-816257795
|
|
2.
|
E JullumstrøA WibeS LydersenTH EdnaColon
cancer incidence, presentation, treatment and outcomes over 25
yearsColorectal Dis13512518201120128833
|
|
3.
|
PP BoyleJS LangmanABC of colorectal
cancer:
epidemiologyBMJ321805808200010.1136/bmj.321.7264.80511009523
|
|
4.
|
NS FearnheadJL WildingWF BodmerGenetics of
colorectal cancer: hereditary aspects and overview of colorectal
tumorigenesisBr Med Bull642743200210.1093/bmb/64.1.2712421723
|
|
5.
|
W DuWY LiR LuJY FangFolate and fiber in
the prevention of colorectal cancer: between shadows and the
lightWorld J
Gastroenterol16921926201010.3748/wjg.v16.i8.92120180229
|
|
6.
|
M BordonaroDL LazarovaAC
SartorelliButyrate and Wnt signaling: a possible solution to the
puzzle of dietary fiber and colon cancer risk?Cell
Cycle711781183200810.4161/cc.7.9.581818418037
|
|
7.
|
P MarksRA RifkindVM RichonR BreslowT
MillerWK KellyHistone deacetylases and cancer: causes and
therapiesNat Rev Cancer1194202200110.1038/3510607911902574
|
|
8.
|
PT MeinkeP LiberatorHistone deacetylase: a
target for antiproliferative and antiprotozoal agentsCurr Med
Chem8211235200110.2174/092986701337378711172676
|
|
9.
|
UH WeidleA GrossmannInhibition of histone
deacetylases: a new strategy to target epigenetic modifications for
anticancer treatmentAnticancer Res2014711485200010928059
|
|
10.
|
EI BukreevaND AksenovAA BardinVA
PospelovTV PospelovaEffect of histone deacetylase inhibitor sodium
butyrate (NaB) on transformants E1a + cHa-Ras expressing wild-type
p53 with suppressed transactivation
functionTsitologiia516977052009(In Russian)
|
|
11.
|
H UchidaT MaruyamaT AraseM OnoT NagashimaH
MasudaH AsadaY YoshimuraHistone acetylation in reproductive organs:
significance of histone deacetylase inhibitors in gene
transcriptionReprod Med
Biol4115122200510.1111/j.1447-0578.2005.00101.x
|
|
12.
|
H CedarY BergmanLinking DNA methylation
and histone modification: patterns and paradigmsNat Rev
Genet10295304200910.1038/nrg254019308066
|
|
13.
|
L Bouchier-HayesSJ MartinCARD games in
apoptosis and immunityEMBO
Rep3616621200210.1093/embo-reports/kvf13912101092
|
|
14.
|
AR StoneW BoboDJ BratNS DeviEG Van MeirPM
VertinoAberrant methylation and down-regulation of TMS1/ASC in
human glioblastomaAm J
Pathol16511511161200410.1016/S0002-9440(10)63376-715466382
|
|
15.
|
A VirmaniA RathiK SugioUG SathyanarayanaS
ToyookaFC KischelV TonkA PadarT TakahashiJA RothAberrant
methylation of TMS1 in small-cell, non-small cell lung cancer and
breast cancerInt J Cancer106198204200310.1002/ijc.1120612800194
|
|
16.
|
MT McCabeJC BrandesPM VertinoCancer DNA
methylation: molecular mechanisms and clinical implicationsClin
Cancer Res1539273937200910.1158/1078-0432.CCR-08-278419509173
|
|
17.
|
AJ WilsonDS ByunN PopovaLB MurrayK
L’ItalienY SowaD ArangoA VelcichLH AugenlichtJM MariadasonHistone
deacetylase 3 (HDAC3) and other class I HDACs regulate colon cell
maturation and p21 expression and are deregulated in human colon
cancerJ Biol
Chem2811354813558200610.1074/jbc.M51002320016533812
|
|
18.
|
BB McConnellPM VertinoTMS1/ASC: the cancer
connectionApoptosis9518200410.1023/B:APPT.0000012117.32430.0c14739594
|
|
19.
|
M KarinY CaoFR GretenZW LiNF-kappaB in
cancer: from innocent bystander to major culpritNat Rev
Cancer2301310200210.1038/nrc78012001991
|
|
20.
|
JN OuJ TorrisaniA UnterbergerN ProvençalK
ShikimiM KarimiTJ EkströmM SzyfHistone deacetylase inhibitor
Trichostatin A induces global and gene-specific DNA demethylation
in human cancer cell linesBiochem
Pharmacol7312971307200710.1016/j.bcp.2006.12.03217276411
|
|
21.
|
MF FragaE UriolL Borja DiegoM BerdascoM
EstellerMJ CañalR RodríguezHigh-performance capillary
electrophoretic method for the quantification of
5-methyl-2′-deoxycytidine in genomic DNA: application to plant,
animal and human cancer tissuesElectrophoresis23167716812002
|
|
22.
|
SB BaylinM EstellerMR RountreeKE BachmanK
SchuebelJG HermanAberrant patterns of DNA methylation, chromation
and gene expression in cancerHum Mol
Genet10687692200110.1093/hmg/10.7.68711257100
|
|
23.
|
M ZhouP LiL TanS QuQL YingH
SongDifferentiation of mouse embryonic stem cells into hepatocytes
induced by a combination of cytokines and sodium butyrateJ Cell
Biochem109606614201010.1002/jcb.2244220039312
|
|
24.
|
M RenL YanCZ ShangJ CaoLH LuJ MinH
ChengEffects of sodium butyrate on the differentiation of
pancreatic and hepatic progenitor cells from mouse embryonic stem
cellsJ Cell Biochem109236244201019911386
|
|
25.
|
M BartelsCR GeestM BieringsM BuitenhuisPJ
CofferHistone deacetylase inhibition modulates cell fate decisions
during myeloid
differentiationHaematology9510521060201010.3324/haematol.2009.00887020107159
|
|
26.
|
PA MarksVM RichonRA RifkindHistone
deacetylase inhibitors: inducers of differentiation or apoptosis of
transformed cellsJ Natl Cancer
Inst9212101216200010.1093/jnci/92.15.121010922406
|
|
27.
|
R PiekarzS BatesA review of depsipeptide
and other histone deacetylase inhibitors in clinical trialsCurr
Pharm Des1022892298200410.2174/138161204338398015279609
|
|
28.
|
HJ ChoSY KimKH KimWK KangJI KimST OhJS
KimCH AnThe combination effect of sodium butyrate and
5-Aza-2′-deoxycytidine on radiosensitivity in RKO colorectal cancer
and MCF-7 breast cancer cell linesWorld J Surg
Oncol749200919460134
|
|
29.
|
L PanAF MatloobJ DuH PanZ DongJ ZhaoY
FengY ZhongB HuangJ LuVitamin D stimulates apoptosis in gastric
cancer cells in synergy with trichostatin A/sodium butyrate-induced
and 5-aza-2′-deoxycytidine-induced PTEN upregulationFEBS
J277989999201020089040
|
|
30.
|
MA CayoAK CayoSM JarjourH ChenSodium
butyrate activates Notch1 signaling, reduces tumor markers and
induces cell cycle arrest and apoptosis in pheochromocytomaAm J
Transl Res1178183200919956429
|
|
31.
|
K KüsterC GrötzingerA KoschelA FischerB
WiedenmannM AndersSodium butyrate increases expression of the
coxsackie and adenovirus receptor in colon cancer cellsCancer
Invest28268274201019863349
|
|
32.
|
L WangHS LuoH XiaSodium butyrate induces
human colon carcinoma HT-29 cell apoptosis through a mitochondrial
pathwayJ Int Med
Res37803811200910.1177/14732300090370032319589263
|
|
33.
|
BG HeerdtMA HoustonGM AnthonyLH
AugenlichtInitiation of growth arrest and apoptosis of MCF-7
mammary carcinoma cells by tributyrin, a triglyceride analogue of
the short-chain fatty acid butyrate, is associated with
mitochondrial activityCancer Res59158415911999
|
|
34.
|
L YanX YangNE DavidsonRole of DNA
methylation and histone acetylation in steroid receptor expression
in breast cancerJ Mammary Gland Biol
Neoplasia6183192200110.1023/A:101130870751211501578
|
|
35.
|
RJ FerranteJK KubilusJ LeeH RyuA BeesenB
ZuckerK SmithNW KowallRR RatanR Luthi-CarterSM HerschHistone
deacetylase inhibition by sodium butyrate chemotherapy ameliorates
the neurodegenerative phenotype in Huntington’s disease miceJ
Neurosci2394189427200314561870
|
|
36.
|
V ChopinRA ToillonN JouyX Le
BourhisP21waf1/cip1 is dispensable for G1 arrest, but indispensable
for apoptosis induced by sodium butyrate in MCF-7 breast cancer
cellsOncogene232129200410.1038/sj.onc.120702014712207
|
|
37.
|
K FukasawaP53, cyclin-dependent kinase and
abnormal amplification of centrosomesBiochim Biophys
Acta17861523200818472015
|
|
38.
|
SY ArcherJ JohnsonHJ KimQ MaH MouV
DaesetyS MengRA HodinThe histone deacetylase inhibitor butyrate
downregulates cyclin B1 gene expression via a p21/WAF-1-dependent
mechanism in human colon cancer cellsAm J Physiol Gastrointest
Liver Physiol289G696G7032005
|
|
39.
|
X YangDL PhillipsAT FergusonWG NelsonJG
HermanNE DavidsonSynergistic activation of functional estrogen
receptor (ER)-alpha by DNA methyltransferase and histone
deacetylase inhibition in human ER-alpha-negative breast cancer
cellsCancer Res61702570292001
|
|
40.
|
R NimmanapalliE O’BryanM HuangP BaliPK
BurnetteT LoughranJ TepperbergR JoveK BhallaMolecular
characterization and sensitivity of STI-571 (imatinib mesylate,
Gleevec)-resistant, Bcr-Abl-positive, human acute leukemia cells to
SRC kinase inhibitor PD180970 and
17-allylamino-17-demethoxygeldanamycinCancer Res62576157692002
|
|
41.
|
T HeinzelOH KrämerPharmacodynamic markers
for histone deacetylase inhibitor developmentDrug Discov Today Dis
Mech4277283200710.1016/j.ddmec.2008.06.003
|
|
42.
|
K YamaguchiA LantowskiAJ DannenbergK
SubbaramaiahHistone deacetylase inhibitors suppress the induction
of c-Jun and its target genes including COX-2J Biol
Chem2803256932577200510.1074/jbc.M50320120015994313
|