Introduction
Oxidative stress results from a persistent imbalance
between antioxidant defense mechanisms and the production of highly
reactive oxygen species (ROS) (1). The chronic hyperglycemia leads to
oxidative stress, which is involved in the progression of
pancreatic β-cell deterioration as well as in the development of
diabetic complications (2). Bone
complications in diabetes include an early onset of osteopenia and
osteoporosis (3,4). These conditions lead to an increase
in bone fracture and a delay in healing of fracture that affects
the quality of life (5). In
vitro studies have shown that oxidative stress inhibits
osteoblastic differentiation (6)
and induces osteoblast insults and apoptosis (7). One of the mechanisms of
diabetes-related bone disease may be the direct effects on
osteoblasts and bone turnover. An imbalance between bone-forming
osteoblasts and bone-resorbing osteoclasts leads to the
pathogenesis and etiology of certain bone metabolic diseases,
including osteoporosis and osteopetrosis (8).
Sugars that contain aldehyde groups that are
oxidized to carboxylic acids are classified as reducing sugars, and
they produce ROS through autoxidation and protein glycosylation
(9–11). The 2-deoxy-D-ribose (dRib) is a
strong reducing sugar that becomes highly reactive with proteins
(12,13). Since glucose is the least reactive
of the reducing sugars and requires long-term exposure time to
provoke oxidative stress by the cells (11), we selected dRib as a surrogate for
glucose to induce oxidative damage of MC3T3-E1 osteoblastic cells.
We previously demonstrated that dRib promotes apoptosis by
increasing oxidative stress in HIT-T15 pancreatic β-cells (12–15) and MC3T3-E1 osteoblastic cells
(16–18).
Glabridin is an isoflavan compound, one of the major
active flavonoids in licorice (19). It has been reported to exhibit
multiple pharmacological activities such as cytotoxic activity
(20), antimicrobial activity
(21) and protection from
oxidative stresses (22).
Treatment of osteoblastic cells with glabridin prevented apoptosis
and production of PGE2 and NO induced by TNF-α (23). Glabridin has been reported to
protect osteoblastic cells against antimycin A-induced cytotoxicity
(24), suggesting that the
enhancement of osteoblast function by glabridin may result in the
prevention of osteoporosis and inflammatory bone diseases. We
recently reported on glabridin-induced prevention of
osteoclastogenesis by inhibition of RANKL-induced activation of
signaling molecules and subsequent transcription factors in
osteoclast precursor cells (25).
Although there is some evidence that the health
beneficial and pharmacological effects of glabridin are due to the
activation of bone formation and the decrease in oxidative stress,
no study has investigated whether glabridin can protect
osteoblastic cells against oxidative damage mediated by dRib, as a
source of ROS. Oxidative stress is involved in the modulation of
the expression of transcription factors and cellular signaling,
which may affect osteoblast functions. In the present study, we
aimed to investigate the effects of glabridin on the oxidative
stress-induced damage and cellular dysfunction in MC3T3-E1
osteoblastic cells.
Materials and methods
Cell culture
Murine osteoblastic MC3T3-E1 cells were obtained
from the American Type Culture Collection (Rockville, MD, USA).
Cells were cultured in α-modified minimal essential medium (α-MEM;
Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine
serum (FBS; Sigma Chemical Co., St. Louis, MO, USA), 100 μU/ml
penicillin, and 100 μg/ml streptomycin. The cultures were
maintained at 37°C in a humidified 5% CO2 atmosphere and
sub-cultured by trypsinization with 0.05% trypsin-0.02% EDTA in
Ca2+- and Mg2+-free Dulbecco’s
phosphate-buffered saline (DPBS) when the cells reached
approximately 70% confluence. For assessment of cell viability,
apoptosis and ROS production, the cells were plated in 24-well
culture plates at a density of 2×104 cells/well. Two
days after culture, cells were treated with glabridin (0.01–50 μM)
for 24 h in α-MEM containing 0.5% FBS. The cells were also seeded
in 6-well culture plates at a density of 1×105
cells/well and treated with culture medium containing 10 mM
β-glycerophosphate and 50 μg/ml ascorbic acid to initiate in
vitro mineralization (26).
Cell culture medium was changed every 2 days. After 6 days, cells
were cultured with medium containing dRib and/or glabridin for 2
days to measure the alkaline phosphatase (ALP) activity, collagen
content, and gene expression.
Assessment of cell viability
Cell viability was determined by measuring cell
metabolic activity using the CCK-8 kit (Dojindo Co., Kumamoto,
Japan) (27). The CCK-8 contains
WST-8 [2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,
4-disulfophenyl)-2H-tetrazolium, monosodium salt], which produces a
water-soluble formazan dye upon reduction in the presence of an
electron carrier. The amount of yellow formazan dye generated by
the activity of dehydrogenases in cells is directly proportional to
the number of living cells. Osteoblastic MC3T3-E1 cells were plated
in 24-well cell culture plates at a density of 2×104
cells/well. At the end of the culture period, 50 μl of the CCK-8
solution was added to each well of the culture plate, which
contained 500 μl of the medium. Following a 4-h incubation,
absorbance was measured with a Zenyth 3100 multimode detector
(Anthos Labtec Instruments GmbH, Wals/Salzburg, Austria) at 450 nm
using a 650 nm filter as a reference. Cells incubated with culture
medium alone were used to determine 100% viability and were
included as a control in all the experiments to allow estimation of
the percent viability of the cell samples.
Apoptosis determination by ELISA
A cell death ELISA kit (Roche Molecular
Biochemicals, Mannheim, Germany), which quantitatively detects
cytosolic histone-associated DNA fragments, was used to measure
apoptosis according to the manufacturer’s instructions. Briefly,
cells were seeded at a density of 2×104 cells in 24-well
culture plates. The culture conditions used were the same as those
described for the cell proliferation assay. Following incubation,
cells were lysed and intact nuclei were pelleted by centrifugation.
An aliquot of supernatant was used as the source of antigen for
sandwich ELISA using a primary anti-histone monoclonal antibody
that was bound to the streptavidin-coated wells of a microtiter
plate. Subsequently, plates were treated with a second anti-DNA
monoclonal antibody coupled to peroxidase. Nucleosome levels were
quantified by determining the amount of peroxidase retained in the
immunocomplex. Peroxidase activity was determined photometrically
at 405 nm using ABTS
[2,2′-azino-di(3-ethylbenzthiazoline-6-sulfonate)] as the
substrate.
Measurement of ROS
The fluorescent probe,
chloromethyl-2,7-dichlorofluorescein diacetate (DCFDA; Molecular
Probes, Eugene, OR, USA), was used to measure intracellular ROS
levels (27). Osteoblastic
MC3T3-E1 cells were cultured for 24 h in α-MEM containing 0.5% FBS,
rinsed twice with DPBS, and then treated with 10 μM of DCFDA for 1
h. Cells were then rinsed, scraped, and their fluorescence was
measured (excitation 485 nm, emission 515 nm) using a Zenyth 3100
multimode detector.
Determination of mitochondrial membrane
potential (ΔΨm)
The ΔΨm of cells was measured using a
JC-1
(5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine
iodide) ΔΨm detection kit (Cayman Chemical Co., Ann
Arbor, MI, USA). Cells were incubated with the
ΔΨm-sensitive fluorescent dye, JC-1 for 20 min at 37°C,
washed twice in DPBS, and then red fluorescence (excitation 550 nm,
emission 600 nm) and green fluorescence (excitation 485 nm,
emission 535 nm) were measured using a Zenyth 3100 multimode
detector. Mitochondrial depolarization (i.e., loss of
ΔΨm) was indicated by a decrease in the red/green
fluorescence ratio.
Alkaline phosphatase activity
At the time of cell harvesting, the medium was
removed and the cell monolayer was gently washed twice with PBS.
The cells were then lysed with 0.2% Triton X-100 and the lysate was
centrifuged at 14,000 × g for 5 min. The cleared supernatant was
used for the measurement of ALP activity and protein concentration.
The ALP activity and protein concentration were determined using an
ALP activity assay kit (Somang Co., Korea) and a Bradford assay kit
(Bio-Rad, Hercules, CA, USA), respectively.
Collagen contents
Cellular collagen content was measured using a
Sircol Collagen Assay kit (Biocolor Ltd., Carrickfergus, Northern
Ireland, UK). This assay is a quantitative dye-binding method
designed for the analysis of collagens extracted from mammalian
tissues and cells during in vitro culture. The dye reagent
binds specifically to the [Gly-X-Y]n helical structure
found in mammalian collagens (types I–V).
RNA extraction
Total RNA was isolated from cells using the TRIzol
reagent (Invitrogen). After isolation, RNA integrity was assessed
using an Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto,
CA, USA). The cDNAs were synthesized with the Transcriptor first
strand cDNA synthesis kit (Roche Diagnostics GmbH, Mannheim,
Germany) and stored at −70°C until further processing. All
procedures were performed according to the manufacturer’s
instructions.
Real-time RT-PCR
Real-time PCR was performed to verify the
differential expression of selected genes using a Roche LightCycler
480 System (Roche Diagnostics GmbH) and the TaqMan method using the
Roche Universal Probe Library (UPL) kit. Relative gene expression
was determined by employing the comparative CT method. All
reactions were carried out in a total volume of 20 μl of reaction
mixture containing 10.0 μl of 2× UPL master mix, 1.0 μl of 5′
primer (10 pmol/μl), 1.0 μl of 3′ primer (10 pmol/ml), 0.2 μl of
UPL probe, 1.0 μl of cDNA and 6.8 μl of sterile water. The thermal
cycling conditions for PCR were an initial denaturation for 10 min
at 95°C, followed by 40 cycles of 94°C for 10 sec and 60°C for 30
sec. The primers summarized in Table
I were designed by the Roche ProbeFinder assay tool. For the
RT-PCR analysis, duplicate PCRs were carried out for each cDNA.
Negative controls (except templates) were included in the PCR
reaction to ensure specific amplification. LightCycler 480 software
version 1.2 (Roche Diagnostics GmbH) was used for the analysis of
the quantitative PCR. The values obtained from each sample were
normalized to HPRT (hypoxanthine guanine phosphoribosyl
transferase) expression. Levels of each gene expression in all
experimental groups were compared to the expression levels of the
control group.
 | Table IPrimer sequences used in this
study. |
Table I
Primer sequences used in this
study.
| Gene | Accession no. | Forward primer | Reverse primer |
|---|
| AKT1 | NM_009652.3 | 5′-TCG TGT GGC AGG
ATG TGT AT-3′ | 5′-ACC TGG TGT CAG
TCT CAG AGG-3′ |
| AKT2 | NM_001110208.1 | 5′-CGA CCC AAC ACC
TTT GTC A-3′ | 5′-GAT AGC CCG CAT
CCA CTC T-3′ |
| AKT3 | NM_011785.3 | 5′-TGG ACC ACT GTT
ATA GAG AGA ACA TTT-3′ | 5′-TGG ATA GCT TCC
GTC CAC TC-3′ |
| ALP | NM_007431.2 | 5′-GGC CAG CTA CAC
CAC AAC A-3′ | 5′-CTG AGC GTT GGT
GTT ATA TGT CTT-3′ |
| BMP2 | NM_007553.2 | 5′-GGT CAC AGA TAA
GGC CAT TGC-3′ | 5′-GCT TCC GCT GTT
TGT GTT TG-3′ |
| BMP4 | NM_007554.2 | 5′-GAG GAG TTT CCA
TCA CGA AGA-3′ | 5′-GCT CTG CCG AGG
AGA TCA-3′ |
| BMP7 | NM_007557.2 | 5′-CGA TAC CAC CAT
CGG GAG TTC-3′ | 5′-AAG GTC TCG TTG
TCA AAT CGC-3′ |
| Collagen | NM_007742.3 | 5′-AGA CAT GTT CAG
CTT TGT GGA C-3′ | 5′-GCA GCT GAC TTC
AGG GAT G-3′ |
| GPX1 | NM_008160.6 | 5′-GGT TTC CCG TGC
AAT CAG T-3′ | 5′-TCG GAC GTA CTT
GAG GGA AT-3′ |
| GPX4 | NM_001037741.2 | 5′-TAA GAA CGG CTG
CGT GGT-3′ | 5′-GTA GGG GCA CAC
ACT TGT AGG-3′ |
| OPG | NM_008764.3 | 5′-ATG AAC AAG TGG
CTG TGC TG-3′ | 5′-CAG TTT CTG GGT
CAT AAT GCA A-3′ |
| OPN | NM_001204201.1 | 5′-TGA GAT TGG CAG
TGA TTT GC-3′ | 5′-ATC TGG GTG CAG
GCT GTA AA-3′ |
| Osteocalcin | NM_031368.4 | 5′-CAC CAT GAG GAC
CCT CTC TC-3′ | 5′-TGG ACA TGA AGG
CTT TGT CA-3′ |
| PI3K | NM_020272.2 | 5′-TTT GGG AGA CTG
AAT CTC TGG-3′ | 5′-GTG GCA TCC TTT
ACA ATC TCG-3′ |
| SOD1 | NM_011434.1 | 5′-CCA TCA GTA TGG
GGA CAA TAC A-3′ | 5′-GGT CTC CAA CAT
GCC TCT CT-3′ |
| SOD2 | NM_013671.3 | 5′-GAC CCA TTG CAA
GGA ACA A-3′ | 5′-GTA GTA AGC GTG
CTC CCA CAC-3′ |
| SOD3 | NM_011435.3 | 5′-GGG GAG GCA ACT
CAG AGG-3′ | 5′-TGG CTG AGG TTC
TCT GCA C-3′ |
Statistical analysis
The results are expressed as the means ± SD.
Statistical analysis was performed using one-way ANOVA with a
subsequent Tukey’s multiple comparison test. A P-value <0.05 was
considered to indicate statistically significant differences.
Statistical analysis was performed using SAS software (SAS
Institute Inc., Cary, NC, USA).
Results
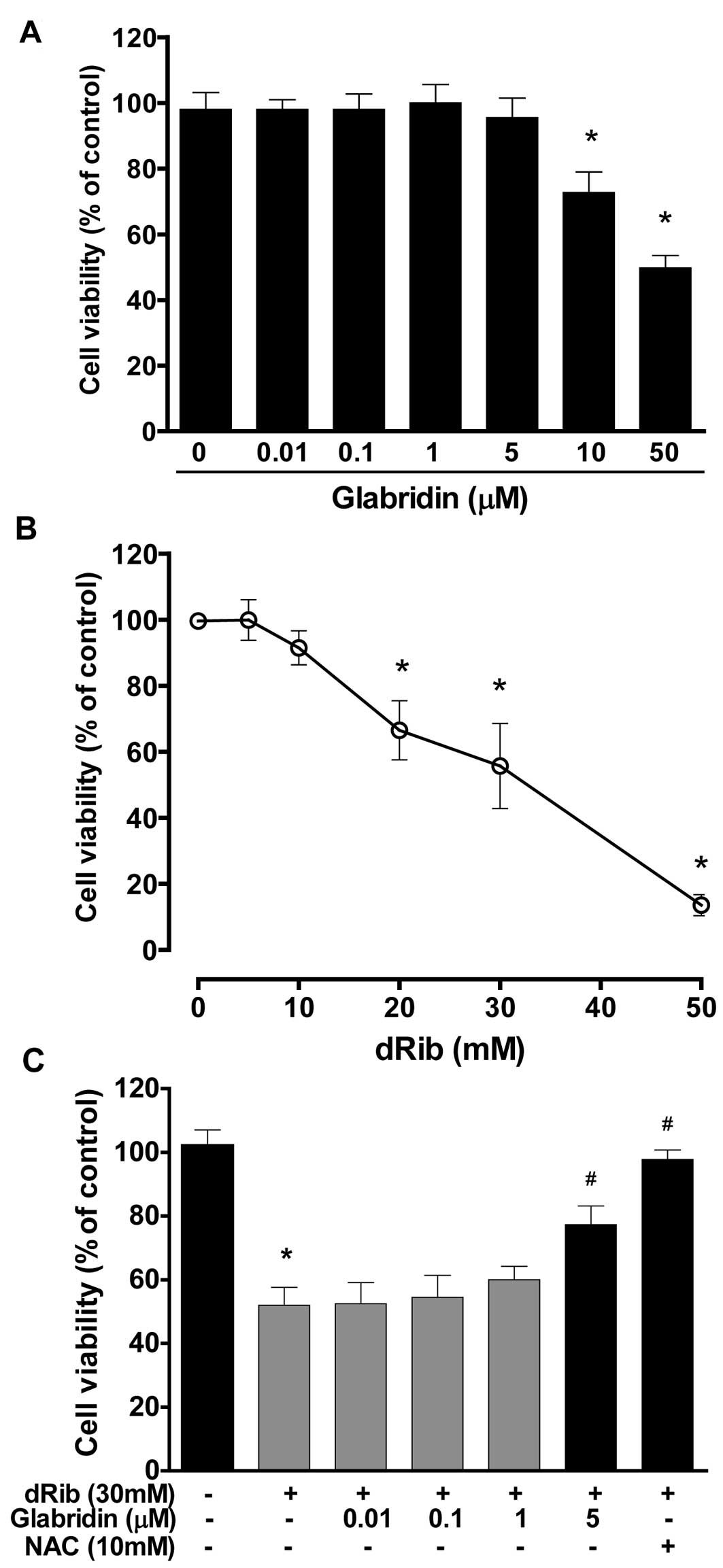
Glabridin inhibits dRib-induced decrease
in MC3T3-E1 osteoblastic cell survival
To evaluate the effect of glabridin itself on
MC3T3-E1 osteoblastic cell survival, cells were incubated in α-MEM
containing 0.5% FBS with increasing concentrations of glabridin
(0.01–50 μM) for 24 h and then cell viability was determined using
the CCK-8 assay. Glabridin at concentrations of 0.01–5 μM had no
effect on cell viability, while higher doses (>10 μM) were found
to be cytotoxic in a dose-dependent manner (Fig. 1A). Therefore, we chose the highest
non-toxic concentration of glabridin (5 μM) for all subsequent cell
culture experiments. To evaluate the effect of dRib on MC3T3-E1
osteoblastic cell survival, cells were incubated for 24 h in α-MEM
containing 0.5% FBS with increasing concentrations of dRib (10–50
mM). We observed a dose-dependent decrease in cell viability in
cells exposed to various concentrations of dRib for 24 h (Fig. 1B). Based on the results of these
cytotoxicity studies, we used 30 mM of dRib in subsequent
biochemical assays. At this concentration, ~50% inhibition of cell
viability occurred in 24 h under our experimental culture
conditions. To determine whether glabridin had an effect on the
dRib-induced decrease in cell survival, cells were preincubated
with glabridin for 30 min and then cultured in the presence of 30
mM of dRib for 24 h. The CCK-8 assays revealed that glabridin (5
μM) partially reversed the dRib-mediated reduction in cell
viability (Fig. 1C). The
antioxidant N-acetyl-L-cysteine (NAC) was used to investigate the
mechanism of dRib-induced cell damage. Pretreatment of cells with
10 mM of NAC almost completely reversed the dRib-induced
cytotoxicity.
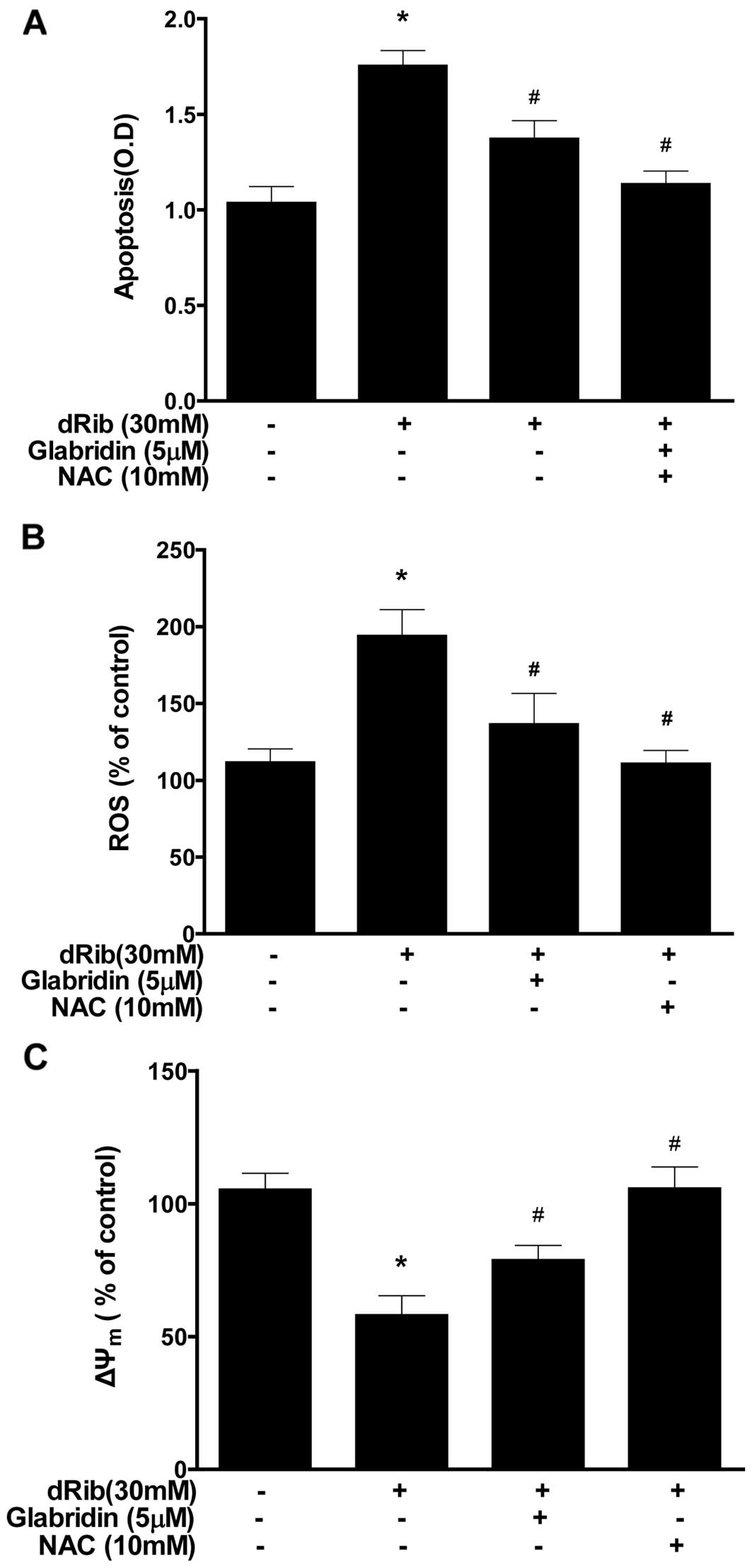
Effect of glabridin on the dRib-induced
ROS generation, apoptosis and intrinsic ΔΨm
The oxidative stress caused by dRib in MC3T3-E1
osteoblastic cells was evaluated by measuring ROS generation,
apoptosis and ΔΨm. Oxidative stress may initiate a
mitochondrial permeability transition event, which is an early
mediator of cellular apoptosis. When cells were treated with 30 mM
of dRib, an increase in ROS generation and apoptosis was observed,
while a decrease in ΔΨm was noted. Treatment with
glabridin (5 μM) in the presence of 30 mM of dRib attenuated all
the dRib-induced effects (Fig.
2). We used the antioxidant NAC to investigate the effect of
oxidative stress in the cells. It was observed that NAC prevented
the dRib-induced cellular effects.
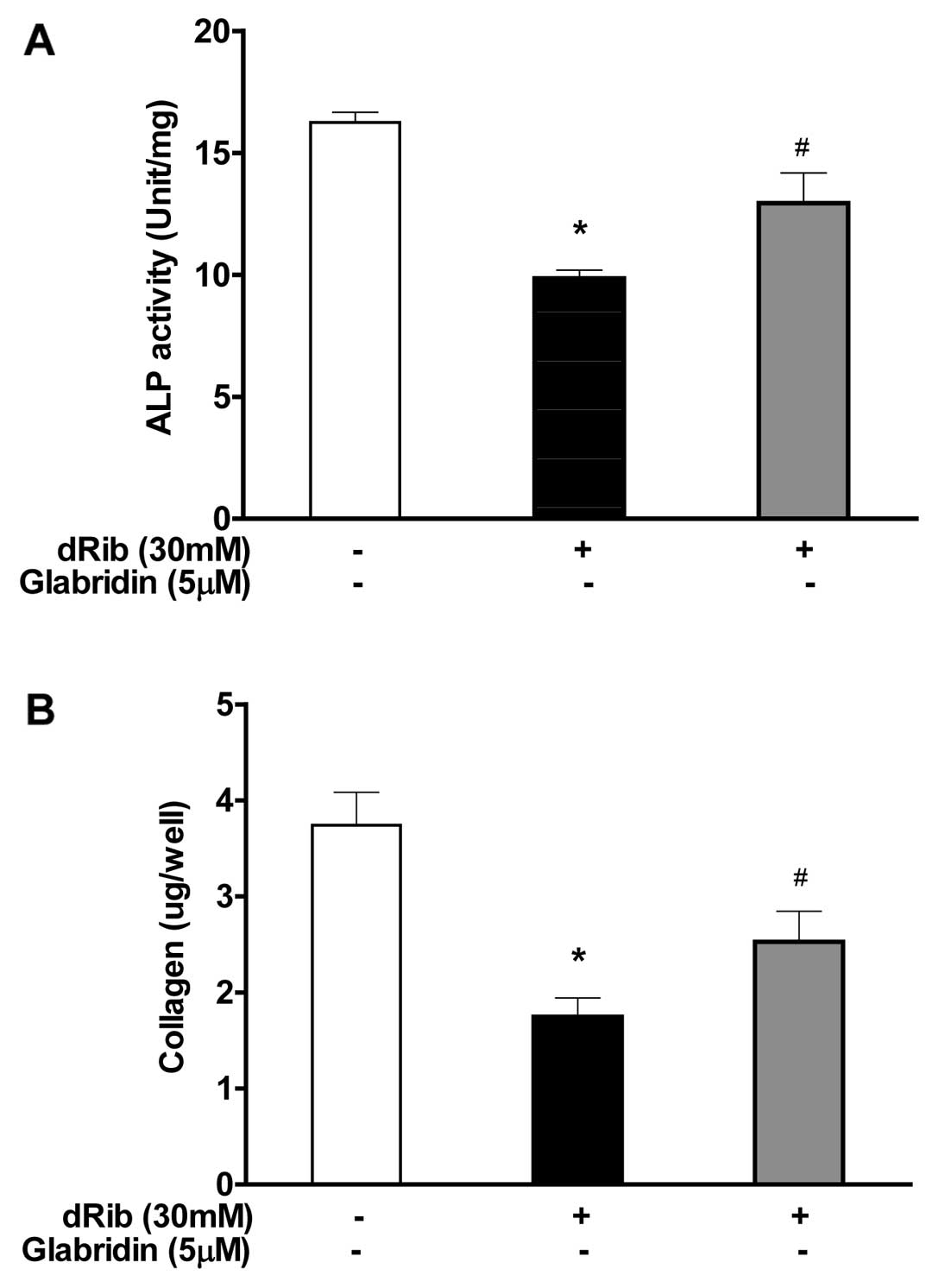
Glabridin inhibits dRib-induced decrease
in ALP activity and collagen contents in MC3T3-E1 osteoblastic
cells
ALP activity and collagen content were measured to
study the effect of glabridin on the osteoblastic cell
differentiation. The reducing sugar dRib was found to have an
inhibitory effect on osteoblastic differentiation markers, however,
when osteoblasts were treated with 5 μM of glabridin in the
presence of 30 mM of dRib, significant increase in the major
osteoblast-specific ALP activity and collagen contents was observed
(Fig. 3).
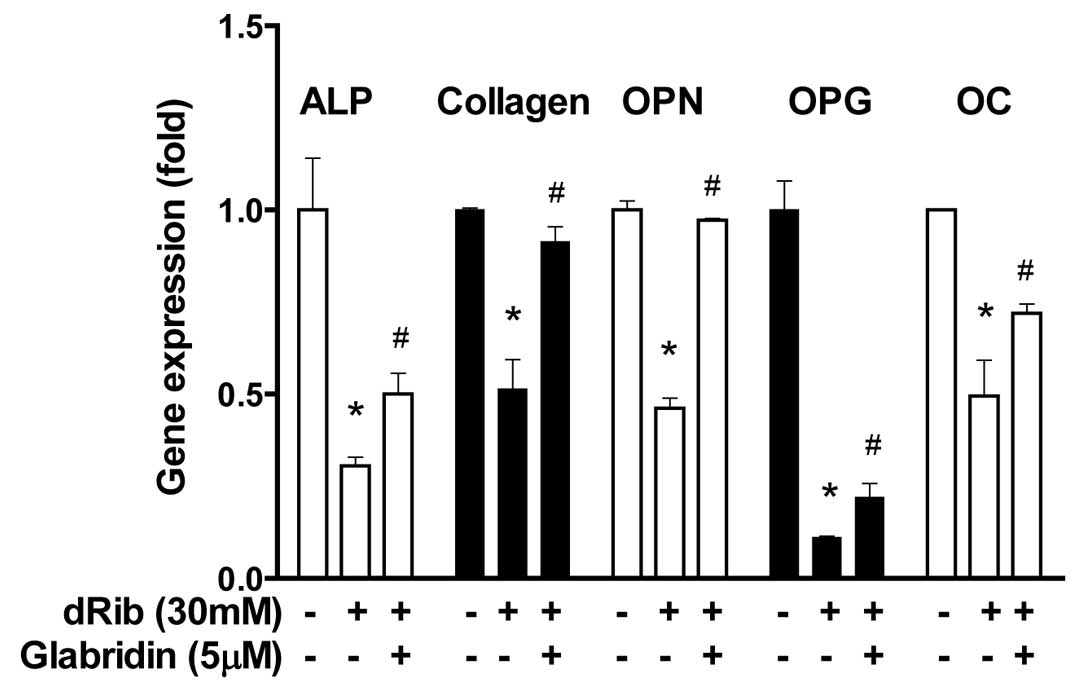
Glabridin inhibits dRib-induced decrease
in differentiation genes in MC3T3-E1 osteoblastic cells
To study the differentiated function of the
osteoblastic cells at the transcriptional level, we analyzed the
gene expression of a number of molecular markers of osteoblast
differentiation. Five differentiation markers [ALP, collagen,
osteopontin (OPN), osteoprotegerin (OPG) and osteocalcin (OC)] were
observed to be downregulated in response to the dRib induction,
however, treatment with 5 μM of glabridin partially inhibited
dRib-induced downregulation of gene expression of differentiation
markers (Fig. 4).
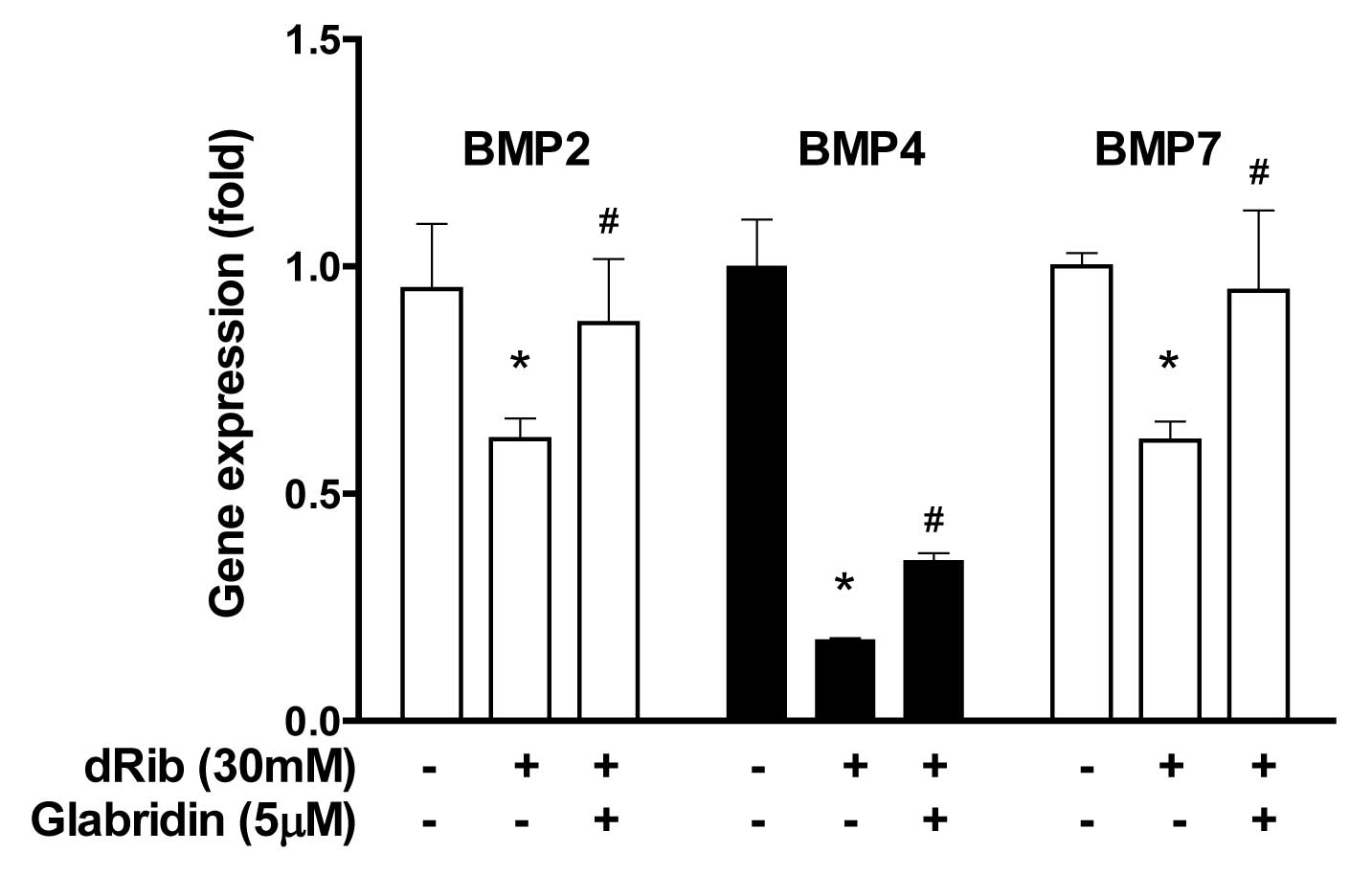
Glabridin inhibits dRib-induced decrease
in bone morphogenetic protein (BMP) genes in MC3T3-E1 osteoblastic
cells
BMPs are known to be the most potent regulators of
osteoblastic differentiation among numerous local factors. The
reducing sugar dRib was found to have an inhibitory effect on the
gene expression of BMPs. However, when osteoblasts were treated
with 5 μM of glabridin in the presence of 30 mM of dRib, BMPs
including BMP2, BMP4 and BMP7 were significantly increased
(Fig. 5).
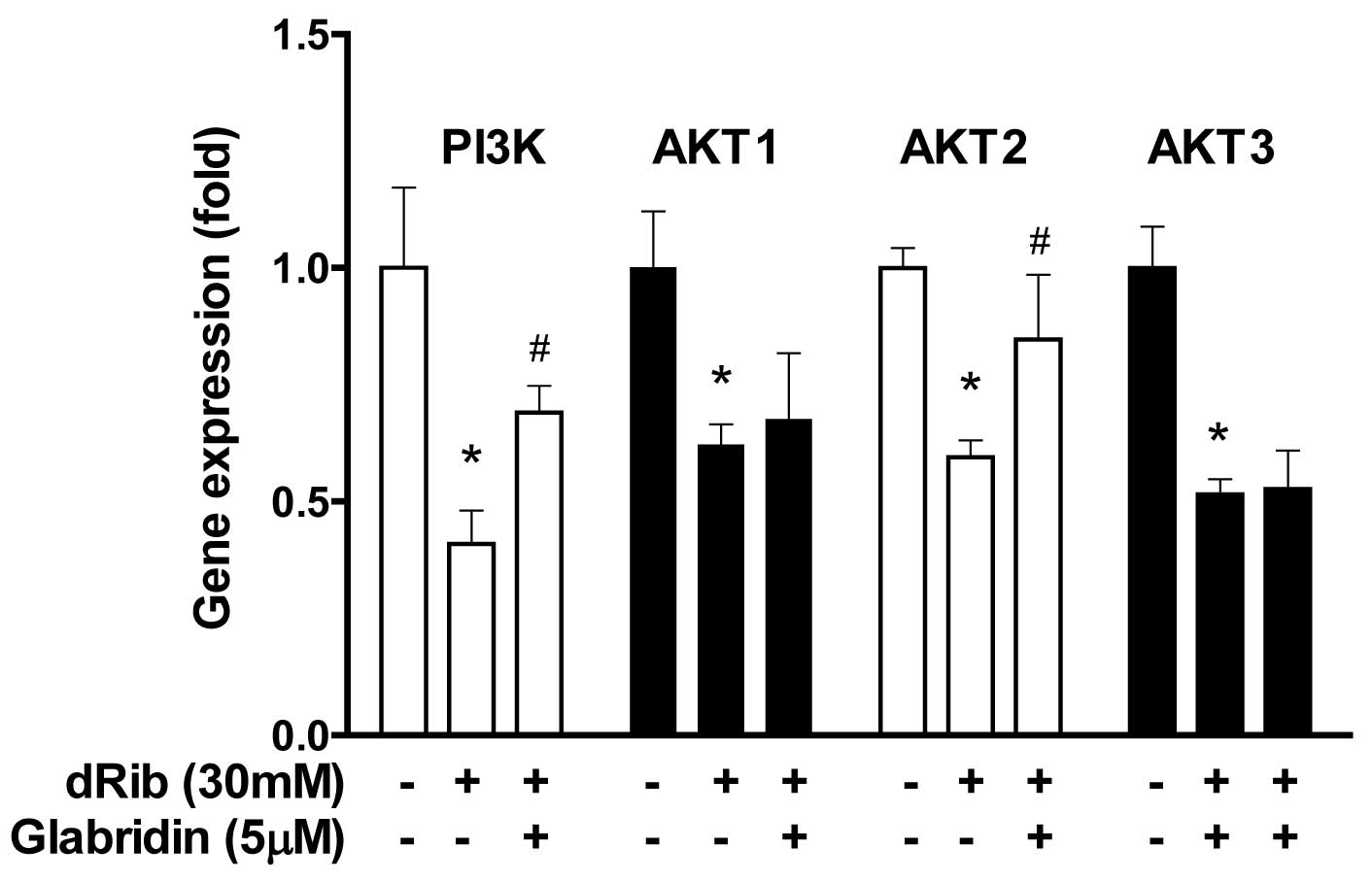
Glabridin inhibits dRib-induced decrease
in phosphatidylinositol 3′-kinase (PI3K) and AKT2 genes in MC3T3-E1
osteoblastic cells
We examined the effect of glabridin on the PI3K/AKT
genes, which were involved in cellular survival pathways. When
osteoblasts were treated with 30 mM of dRib, the gene expression of
PI3K, AKT1, AKT2 and AKT3 was downregulated. However, treatment
with 5 μM of glabridin in the presence of 30 mM dRib increased the
gene expression of PI3K and AKT2 but not AKT1, AKT3 (Fig. 6).
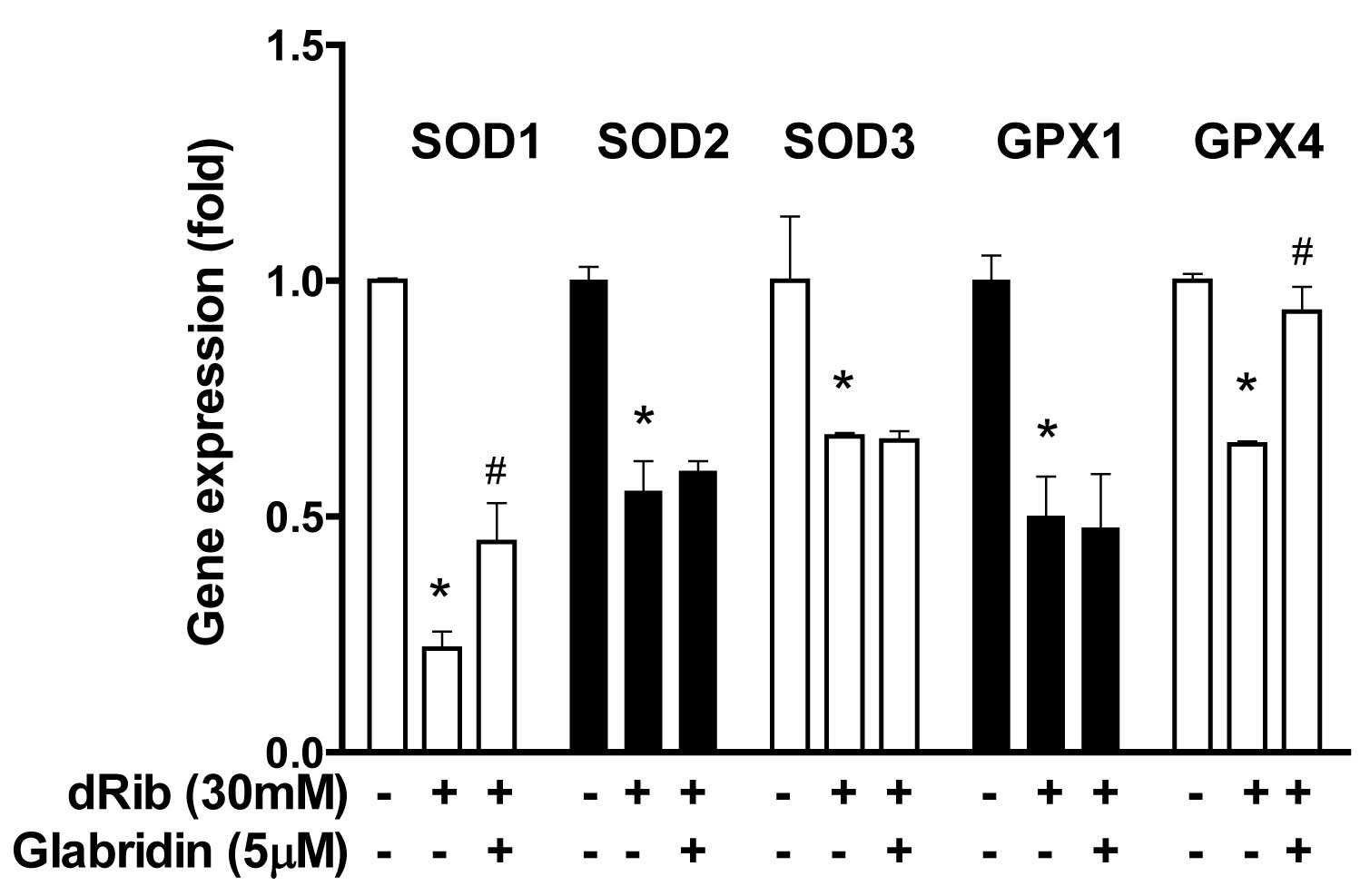
Glabridin inhibits dRib-induced decrease
in antioxidant enzyme gene in MC3T3-E1 osteoblastic cells
In addition to the biochemical aspects of oxidative
stress, the gene expression of antioxidative enzymes was
investigated. When osteoblasts were treated with 30 mM of dRib,
gene expression of SOD1, SOD2, SOD3, GPX1 and GPX4 was
downregulated. However, treatment with 5 μM of glabridin in the
presence of 30 mM of dRib led to an increase in the gene expression
of SOD1 and GPX4, but not SOD2, SOD3, or GPX1 (Fig. 7).
Discussion
In the present study, we investigated the effects of
glabridin on dRib-induced oxidative damage and cellular dysfunction
using an MC3T3-E1 osteoblastic cell culture model. The dRib is a
strong reducing sugar that is highly reactive with proteins, in
contrast to glucose which has the lowest reducing capacity of all
the monosaccharides (9).
Therefore, we chose dRib as a surrogate for glucose to study the
effects of oxidative stress in osteoblastic cells. One of the
mechanisms of diabetes-related bone disease may be the direct
effects of high glucose on osteoblastic cells. This hypothesis is
explained by in vitro studies, which state that
hyperglycemia has been shown to inhibit osteoblastic cell
proliferation and differentiation (28,29), indicating that extracellular high
glucose would directly impair osteoblastic functions resulting in
bone disease. In our previous studies, we found that dRib induces
cellular damage in pancreatic β-cells by increasing oxidative
stress and protein glycation (12–15). Recently, we also reported that
dRib induces cellular dysfunction and apoptosis in the MC3T3-E1
mouse osteoblastic cell line by increasing oxidative stress
(18). In this study, glabridin
partially reversed the dRib-mediated reduction in cell viability
(Fig. 1). The antioxidant NAC was
used to investigate the mechanism of dRib-induced cell damage.
Pretreatment of cells with NAC almost completely reversed the
dRib-induced cytotoxicity. These findings suggest that the
dRib-induced cytotoxicity was most likely due to oxidative
stress-induced effects. Our recent finding showed that the
antioxidants NAC and α-lipoic acid (ALA) almost reversed the
dRib-mediated reduction in viability of MC3T3-E1 osteoblastic cells
(18). When cells were treated
with dRib, ROS generation and apoptosis increased, while
ΔΨm decreased. Treatment with glabridin in the presence
of dRib attenuated all the dRib-induced effects (Fig. 2). The antioxidant NAC prevented
the dRib-induced cellular effects. These data are consistent with
our previous studies, which stated that the antioxidants NAC and
ALA protected pancreatic β-cells and MC3T3-E1 osteoblastic cells
against oxidative stress as shown in terms of reduction in ROS
generation and apoptosis (12–14,18,27). These findings indicate that
glabridin can function as an antioxidant and thereby protect
MC3T3-E1 osteoblastic cells from dRib-induced oxidative cell
damage. We used JC-1 staining to assess ΔΨm, which is a
marker of mitochondrial oxidative phosphorylation activity. The
JC-1 is a lipophilic and cationic dye that permeates plasma and
mitochondrial membranes of cells. A low JC-1 ratio indicates a low
amount of the aggregated form of JC-1 in the mitochondria, which
correlates with a high amount of ROS (30). Oxidative stress may initiate a
mitochondrial permeability transition event, which is an early
indication of the initiation of cellular apoptosis. This process is
typically defined as a collapse in the electrochemical gradient
across the mitochondrial membrane, as measured by the change in the
ΔΨm (31).
Mitochondrial dysfunction is a consequence of oxidative damage
caused by increased oxidant levels. In our study, pretreatment of
cells with glabridin attenuated marked decrease in dRib-induced
ΔΨm (Fig. 2C). These
results indicate that glabridin protects mitochondrial functions by
functioning as an antioxidant in osteoblastic cells.
The present study demonstrated attenuation of
dRib-induced decrease in major osteoblast differentiation markers,
such as ALP activity and collagen contents, in the presence of
glabridin (Fig. 3). The MC3T3-E1
cells are the osteoblastic precursor cell lines, which are cloned
from calvariae of newborn mice (32); they are the most frequently used
cell lines to study osteoblast differentiation, as well as the
function of mature osteoblastic cells. Osteoblast differentiation
is the primary event of bone formation. Bone ALP is a glycoprotein
localized in the plasma membrane of osteoblastic cells, which is
also one of the osteoblastic phenotype markers (33). Alterations in this activity have
been observed in osteoporosis and other metabolic bone diseases.
High levels of ALP activity are shown in both preosteoblasts and
osteoblasts in vivo and in differentiating osteoblasts in
vitro. Osteoblast cells produce type I collagen, which is the
most abundant protein in the bone matrix, serves an early marker of
osteoblast differentiation, and is the major organic component of
mineralized bone matrix (34).
In this study, five differentiation markers (ALP,
collagen, OPN, OPG, OC) were found to be downregulated by dRib,
however, glabridin partially inhibited dRib-induced downregulation
of differentiation genes (Fig.
4). OPN is a major acidic phosphorylated glycoprotein secreted
by osteoblasts and acts as a regulator of bone formation (35). OPG, produced by osteoblasts, is
one of the regulators of bone metabolism and inhibits bone
resorption by regulating function of osteoclast (36). OC is also known as a marker of
bone formation and is the most abundant non-collagenous protein in
bone (37). Thus, these molecular
markers are important and are the best-known regulators of
osteoblast function.
When osteoblasts were treated with glabridin in the
presence of dRib, a significant increase in the bone morphogenetic
proteins BMP2, BMP4 and BMP7 was observed (Fig. 5). The MC3T3-E1 cells are highly
BMP-responsive and can complete differentiation processes in the
long-term cultures. Previous studies have demonstrated that BMPs
stimulate ALP activity, collagen synthesis, parathyroid hormone
(PTH) responsiveness, and osteocalcin production in osteoblastic
cells (38–40), suggesting that BMPs stimulate
differentiation function of osteoblastic cells. It has been
suggested that glabridin exerts differentiation function of
osteoblastic cells by stimulation of BMP production.
Glabridin was also found to induce the activation of
PI3K and AKT2, which were inhibited by dRib (Fig. 6). Since these signalings are
involved in cellular survival pathways, glabridin might be
cytoprotective for osteoblastic cells during oxidative stress
responses. The phosphatidylinositol 3′-kinase (PI3K)-AKT (protein
kinase B) signaling pathway is known to be activated by many types
of cellular stimuli or toxic insults and regulates fundamental
cellular functions such as transcription, translation,
proliferation, growth, and survival (41). One of the important functions of
activated PI3K in cells is the inhibition of apoptosis (42). The AKT is a good candidate for
mediating these PI3K-dependent cell survival responses. AKT has
been implicated as an anti-apoptotic factor in many different cell
death paradigms, including the withdrawal of extracellular
signaling factors, oxidative and osmotic stress, irradiation, and
the treatment of cells with chemotherapeutic drugs and ischemic
shock (43). Zinc has been
reported to inhibit hydrogen peroxide-induced MC3T3-E1 osteoblastic
cells via the mitogen activated protein kinase (MAPK) and PI3K/AKT
pathways. The various flavonoids including deoxyactein (44), honokiol (45), apocynin (46) have been demonstrated to impart
protective effects against antimycin A (an inhibitor of
mitochondrial electron transport)-induced oxidative cell damage via
the activation of PI3K and/or AKT in MC3T3-E1 osteoblastic
cells.
When osteoblasts were treated with glabridin in the
presence of dRib, significant increases in the gene expression of
SOD1 and GPX4 were observed (Fig.
7). In general, excess ROS must be promptly eliminated from the
cell by a variety of antioxidant defense mechanisms. Cellular
antioxidant enzymes and other redox molecules serve to
counterbalance ROS generated in the cell. Superoxide dismutase
(SOD), which catalyzes the dismutation of the superoxide anion into
hydrogen peroxide and molecular oxygen, is one of the most
important antioxidant enzymes (47). SOD enzymes are classified into
three groups: CuZn-SOD (SOD1), which is located in the cytoplasm,
Mn-SOD (SOD2), which is located in the mitochondria, and EC-SOD
(SOD3), which is located in extracellular location. Glutathione
peroxidase (GPx) catalyzes the reduction of hydroperoxides,
including hydrogen peroxides, by reduced glutathione and functions
to protect the cell from oxidative damage. Glutathione peroxidase 1
(GPX1) is the most abundant version, found in the cytoplasm of
nearly all mammalian tissues, whose preferred substrate is hydrogen
peroxide. GPX4 has a high preference for lipid hydroperoxides. It
has been reported that various flavonoids increased the activity of
antioxidant enzymes in osteoblastic cells. Quercetin can diminish
oxidative human osteoblastic cell damage by scavenging the radicals
and by upregulating the expression of heme oxygenase-1 (HO-1) and
SOD-1 exposed to cigarette smoke medium (48). An extract of total flavonoids from
persimmon leaves significantly decreased the level of reactive
oxygen species (ROS) and malondialdehyde (MDA), while increasing
the activity of catalase (CAT), SOD and GPX in MC3T3-E1 cells.
Simvastatin abated oxidative stress through enhancing catalase,
HO-1, and SOD activity and suppressing NADPH oxidase activity in an
aged and ovariectomized rat model (49). Another study demonstrated that
intracellular redox imbalance caused by SOD1 deficiency played a
pivotal role in the development and progression of bone fragility
both in vivo and in vitro (50).
In contrast to the antioxidant effects of glabridin
in osteoblastic cells as noted in this study, glabridin has also
been shown to confer anticancer effects in other cell types.
Previously, it has been demonstrated that glabridin exhibited
effective inhibition of cell metastasis by decreasing cancer cell
migration and invasion of human non-small cell lung cancer A549
cells (51). Glabridin has also
been reported to exhibit effective inhibition of cell metastasis by
decreasing cancer cell migration and invasion of MDA-MB-231 human
breast adenocarcinoma cells (52). Thus, glabridin appears to have
different biological effects on different cell types. The mechanism
through which glabridin exerts these cell-specific effects remains
to be elucidated.
In summary, we have shown that glabridin attenuates
dRib-induced cell damage in MC3T3-E1 osteoblastic cells due to its
antioxidant activity and improves differentiation function, which
may promote bone recovery in diabetes-related bone diseases.
Acknowledgements
This study was supported by the ACE program through
the National Research Foundation of Korea (NRF) grant funded by the
Korean Ministry of Education, Science and Technology (MEST) (no.
20110028203).
References
|
1
|
Robertson RP, Harmon J, Tran PO, Tanaka Y
and Takahashi H: Glucose toxicity in beta-cells: type 2 diabetes,
good radicals gone bad, and the glutathione connection. Diabetes.
52:581–587. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Robertson RP: Chronic oxidative stress as
a central mechanism for glucose toxicity in pancreatic islet beta
cells in diabetes. J Biol Chem. 279:42351–42354. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
López-Ibarra PJ, Pastor MM,
Escobar-Jiménez F, Pardo MD, González AG, Luna JD, Requena ME and
Diosdado MA: Bone mineral density at time of clinical diagnosis of
adult-onset type 1 diabetes mellitus. Endocr Pract. 7:346–351.
2001.PubMed/NCBI
|
|
4
|
Tuominen JT, Impivaara O, Puukka P and
Ronnemaa T: Bone mineral density in patients with type 1 and type 2
diabetes. Diabetes Care. 22:1196–1200. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Herskind AM, Christensen K,
Nørgaard-Andersen K and Andersen JF: Diabetes mellitus and healing
of closed fractures. Diabetes Metab. 18:63–64. 1992.PubMed/NCBI
|
|
6
|
Bai XC, Lu D, Bai J, Zheng H, Ke ZY, Li XM
and Luo SQ: Oxidative stress inhibits osteoblastic differentiation
of bone cells by ERK and NF-kappaB. Biochem Biophys Res Commun.
314:197–207. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fatokun AA, Stone TW and Smith RA:
Hydrogen peroxide-induced oxidative stress in MC3T3-E1 cells: the
effects of glutamate and protection by purines. Bone. 39:542–551.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Seeman E: Reduced bone formation and
increased bone resorption: rational targets for the treatment of
osteoporosis. Osteoporos Int. 14(Suppl 3): S2–S8. 2003.PubMed/NCBI
|
|
9
|
Thornalley P, Wolff S, Crabbe J and Stern
A: The autoxidation of glyceraldehyde and other simple
monosaccharides under physiological conditions catalysed by buffer
ions. Biochim Biophys Acta. 797:276–287. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kaneto H, Fujii J, Myint T, Miyazawa N,
Islam KN, Kawasaki Y, Suzuki K, Nakamura M, Tatsumi H, Yamasaki Y
and Taniguchi N: Reducing sugars trigger oxidative modification and
apoptosis in pancreatic beta-cells by provoking oxidative stress
through the glycation reaction. Biochem J. 320:855–863.
1996.PubMed/NCBI
|
|
11
|
Bunn HF and Higgins PJ: Reaction of
monosaccharides with proteins: possible evolutionary significance.
Science. 213:222–224. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Koh G, Suh KS, Chon S, Oh S, Woo JT, Kim
SW, Kim JW and Kim YS: Elevated cAMP level attenuates
2-deoxy-d-ribose-induced oxidative damage in pancreatic beta-cells.
Arch Biochem Biophys. 438:70–79. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Koh G, Lee DH and Woo JT: 2-Deoxy-D-ribose
induces cellular damage by increasing oxidative stress and protein
glycation in a pancreatic beta-cell line. Metabolism. 59:325–332.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee YJ, Suh KS, Choi MC, Chon S, Oh S, Woo
JT, Kim SW, Kim JW and Kim YS: Kaempferol protects HIT-T15
pancreatic beta cells from 2-deoxy-D-ribose-induced oxidative
damage. Phytother Res. 24:419–423. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Suh KS, Oh S, Woo JT, Kim SW, Kim JW, Kim
YS and Chon S: Apigenin attenuates 2-deoxy-D-ribose-induced
oxidative cell damage in HIT-T15 pancreatic β-cells. Biol Pharm
Bull. 35:121–126. 2012.PubMed/NCBI
|
|
16
|
Choi EM and Kim YH: Hesperetin attenuates
the highly reducing sugar-triggered inhibition of osteoblast
differentiation. Cell Biol Toxicol. 24:225–231. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee KH and Choi EM: Myricetin, a naturally
occurring flavonoid, prevents 2-deoxy-D-ribose induced dysfunction
and oxidative damage in osteoblastic MC3T3-E1 cells. Eur J
Pharmacol. 591:1–6. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Suh KS, Choi EM, Kwon M, Chon S, Oh S, Woo
JT, Kim SW, Kim JW and Kim YS: Kaempferol attenuates
2-deoxy-d-ribose-induced oxidative cell damage in MC3T3-E1
osteoblastic cells. Biol Pharm Bull. 32:746–749. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chin YW, Jung HA, Liu Y, Su BN, Castoro
JA, Keller WJ, Pereira MA and Kinghorn AD: Anti-oxidant
constituents of the roots and stolons of licorice (Glycyrrhiza
glabra). J Agric Food Chem. 55:4691–4697. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fukai T, Sakagami H, Toguchi M, Takayama
F, Iwakura I, Atsumi T, Ueha T, Nakashima H and Nomura T: Cytotoxic
activity of low molecular weight polyphenols against human oral
tumor cell lines. Anticancer Res. 20:2525–2536. 2000.PubMed/NCBI
|
|
21
|
Fukai T, Marumo A, Kaitou K, Kanda T,
Terada S and Nomura T: Anti-Helicobacter pylori flavonoids from
licorice extract. Life Sci. 71:1449–1463. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Haraguchi H, Yoshida N, Ishikawa H, Tamura
Y, Mizutani K and Kinoshita T: Protection of mitochondrial
functions against oxidative stresses by isoflavans from
Glycyrrhiza glabra. J Pharm Pharmacol. 52:219–223. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Choi EM: The licorice root derived
isoflavan glabridin increases the function of osteoblastic MC3T3-E1
cells. Biochem Pharmacol. 70:363–368. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Choi EM: Glabridin protects osteoblastic
MC3T3-E1 cells against antimycin A induced cytotoxicity. Chem Biol
Interact. 193:71–78. 2011. View Article : Google Scholar
|
|
25
|
Kim HS, Suh KS, Sul D, Kim BJ, Lee SK and
Jung WW: The inhibitory effect and the molecular mechanism of
glabridin on RANKL-induced osteoclastogenesis in RAW264.7 cells.
Int J Mol Med. 29:169–177. 2012.PubMed/NCBI
|
|
26
|
Kanno S, Anuradha CD and Hirano S:
Localization of zinc after in vitro mineralization in osteoblastic
cells. Biol Trace Elem Res. 83:39–47. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Suh KS, Chon S, Oh S, Kim SW, Kim JW, Kim
YS and Woo JT: Prooxidative effects of green tea polyphenol
(−)-epigallocatechin-3-gallate on the HIT-T15 pancreatic beta cell
line. Cell Biol Toxicol. 26:189–199. 2010.
|
|
28
|
Balint E, Szabo P, Marshall CF and Sprague
SM: Glucose-induced inhibition of in vitro bone mineralization.
Bone. 28:21–28. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Terada M, Inaba M, Yano Y, Hasuma T,
Nishizawa Y, Morii H and Otani S: Growth-inhibitory effect of a
high glucose concentration on osteoblast-like cells. Bone.
22:17–23. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Szilágyi G, Simona L, Koska P, Telek G and
Nagy Z: Visualization of mitochondrial membrane potential and
reactive oxygen species via double staining. Neurosci Lett.
399:206–209. 2006.PubMed/NCBI
|
|
31
|
Salido M, Gonzalez JL and Vilches J: Loss
of mitochondrial membrane potential is inhibited by bombesin in
etoposide-induced apoptosis in PC-3 prostate carcinoma cells. Mol
Cancer Ther. 6:1292–1299. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sudo H, Kodama HA, Amagai Y, Yamamoto S
and Kasai S: In vitro differentiation and calcification in a new
clonal osteogenic cell line derived from newborn mouse calvaria. J
Cell Biol. 96:191–198. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bellows CG, Aubin JE and Heersche JN:
Initiation and progression of mineralization of bone nodules formed
in vitro: the role of alkaline phosphatase and organic phosphate.
Bone Miner. 14:27–40. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Domon S, Shimokawa H, Yamaguchi S and Soma
K: Temporal and spatial mRNA expression of bone sialoprotein and
type I collagen during rodent tooth movement. Eur J Orthod.
23:339–348. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen Y, Bal BS and Gorski JP: Calcium and
collagen binding properties of osteopontin, bone sialoprotein, and
bone acidic glycoprotein-75 from bone. J Biol Chem.
267:24871–24878. 1992.PubMed/NCBI
|
|
36
|
Khosla S: Minireview: the OPG/RANKL/RANK
system. Endocrinology. 142:5050–5055. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Price PA and Nishimoto SK:
Radioimmunoassay for the vitamin K-dependent protein of bone and
its discovery in plasma. Proc Natl Acad Sci USA. 77:2234–2238.
1980. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Takuwa Y, Ohse C, Wang EA, Wozney JM and
Yamashita K: Bone morphogenetic protein-2 stimulates alkaline
phosphatase activity and collagen synthesis in cultured
osteoblastic cells, MC3T3-E1. Biochem Biophys Res Commun.
174:96–101. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nakase T, Takaoka K, Masuhara K, Shimizu
K, Yoshikawa H and Ochi T: Interleukin-1β enhances and tumor
necrosis factor-α inhibits bone morphogenetic protein-2-induced
alkaline phosphatase activity in MC3T3-E1 osteoblastic cells. Bone.
21:17–21. 1997.
|
|
40
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yao R and Cooper GM: Requirement for
phosphatidylinositol-3 kinase in the prevention of apoptosis by
nerve growth factor. Science. 267:2003–2006. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Franke TF, Kaplan DR and Cantley LC: PI3K:
downstream AKTion blocks apoptosis. Cell. 88:435–437. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Choi EM: Deoxyactein isolated from
Cimicifuga racemosa protects osteoblastic MC3T3-E1 cells
against antimycin A-induced cytotoxicity. J Appl Toxicol. Dec
19–2011.(Epub ahead of print). View Article : Google Scholar : 2011.
|
|
44
|
Choi EM: Honokiol protects osteoblastic
MC3T3-E1 cells against antimycin A-induced cytotoxicity. Inflamm
Res. 60:1005–1012. 2011. View Article : Google Scholar
|
|
45
|
Choi EM and Lee YS: Protective effect of
apocynin on antimycin A-induced cell damage in osteoblastic
MC3T3-E1 cells. J Appl Toxicol. May 2–2011.(Epub ahead of print).
View Article : Google Scholar : 2011.
|
|
46
|
Zelko IN, Mariani TJ and Folz RJ:
Superoxide dismutase multigene family: a comparison of the CuZn-SOD
(SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures,
evolution, and expression. Free Radic Biol Med. 33:337–349. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Braun KF, Ehnert S, Freude T, Egaña JT,
Schenck TL, Buchholz A, Schmitt A, Siebenlist S, Schyschka L,
Neumaier M, Stöckle U and Nussler AK: Quercetin protects primary
human osteoblasts exposed to cigarette smoke through activation of
the antioxidative enzymes HO-1 and SOD-1. Sci World J.
11:2348–2357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sun L, Zhang J, Lu X, Zhang L and Zhang Y:
Evaluation to the antioxidant activity of total flavonoids extract
from persimmon (Diospyros kaki L.) leaves. Food Chem
Toxicol. 49:2689–2696. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yin H, Shi ZG, Yu YS, Hu J, Wang R, Luan
ZP and Guo DH: Protection against osteoporosis by statins is linked
to a reduction of oxidative stress and restoration of nitric oxide
formation in aged and ovariectomized rats. Eur J Pharmacol.
674:200–206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Nojiri H, Saita Y, Morikawa D, Kobayashi
K, Tsuda C, Miyazaki T, Saito M, Marumo K, Yonezawa I, Kaneko K,
Shirasawa T and Shimizu T: Cytoplasmic superoxide causes bone
fragility owing to low-turnover osteoporosis and impaired collagen
cross-linking. J Bone Miner Res. 26:2682–2694. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tsai YM, Yang CJ, Hsu YL, Wu LY, Tsai YC,
Hung JY, Lien CT, Huang MS and Kuo PL: Glabridin inhibits
migration, invasion, and angiogenesis of human non-small cell lung
cancer A549 cells by inhibiting the FAK/rho signaling pathway.
Integr Cancer Ther. 10:341–349. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hsu YL, Wu LY, Hou MF, Tsai EM, Lee JN,
Liang HL, Jong YJ, Hung CH and Kuo PL: Glabridin, an isoflavan from
licorice root, inhibits migration, invasion and angiogenesis of
MDA-MB-231 human breast adenocarcinoma cells by inhibiting focal
adhesion kinase/Rho signaling pathway. Mol Nutr Food Res.
55:318–327. 2011. View Article : Google Scholar : PubMed/NCBI
|