Introduction
Rheumatoid arthritis (RA) is an inflammatory joint
disease characterized by hyperplasia of the synovial tissue and
formation of pannus, which grows invasively into the cartilage,
causing cartilage and bone destruction. Analyses of hyperplastic
synovial tissue of patients with RA have revealed a number of
features of transformed long-living cells, such as the presence of
somatic mutations, expression of oncogenes and resistance to
apoptosis (1–3).
We previously reported that the decoy receptor 3
(DcR3)/TR6/M68/tumor necrosis factor receptor (TNFR) superfamily
member 6 (TNFRSF6b) is expressed in rheumatoid fibroblast-like
synoviocytes (RA-FLS), and that DcR3 expression induced in RA-FLS
by TNFα protects cells from Fas-induced apoptosis (4). DcR3, a member of the TNFR
superfamily, lacks the transmembrane domain of conventional TNFRs
and thus can be a secreted protein (5). DcR3 is typically overexpressed in
tumor cells, including lung and colon cancers (5), gliomas, gastrointestinal tract
tumors (6) and virus-associated
leukemia (7). In addition, as
previous studies have demonstrated, DcR3 is expressed in some
normal tissues, including the colon, stomach, spleen, lymph nodes,
spinal cord, pancreas and lungs (5,6).
However, DcR3 is not expressed in NIH3T3 human fibroblast cells
(8). DcR3 has 3 ligands, Fas
ligand (FasL), LIGHT and TNF-like ligand 1A (TL1A), which are
members of the TNF superfamily (9). The overexpression of DcR3 may
benefit tumors by helping them avoid the cytotoxic and regulatory
effects of FasL (5,10), LIGHT (11) and TL1A (12). In a previous study, we suggested
that DcR3 is one of the key molecules that regulate the
proliferation of RA-FLS (4).
Previous studies have suggested that DcR3 directly
induces osteoclast formation from monocytes (13), and that DcR3 triggers the enhanced
adhesion of monocytes via reverse signaling (14). We have also reported that DcR3
induces very late antigen-4 (VLA-4) expression in THP-1
macrophages, inhibiting cycloheximide-induced apoptosis (15). As for RA-FLS, in a recent study,
we reported that DcR3 binds to membrane-bound TL1A expressed on
RA-FLS, resulting in the negative regulation of cell proliferation
induced by inflammatory cytokines (16). Therefore, we hypothesized that
DcR3 plays a role in the pathogenesis of RA, not only as a decoy
receptor, but also as a ligand via TL1A on RA-FLS. However, the
function of DcR3 as a ligand in RA-FLS is not yet well understood.
In the current study, we searched for genes in RA-FLS whose
expression was regulated by the ligation of DcR3 using cDNA
microarray. The gene expression profiles may reveal the possible
target molecules that play a significant role in the DcR3-TL1A
signaling pathway in the pathogenesis of RA.
Materials and methods
Isolation and culture of synovial
fibroblasts
RA-FLS were obtained during total knee replacement
surgery from 4 patients (samples 1–4) with RA who fulfilled the
1987 criteria of the American College of Rheumatology (formerly,
the American Rheumatism Association) (17), who had never been treated with
biological drugs. Synovial samples were collected from the patients
who provided written consent in order to participate in this study
in accordance with the World Medical Association Declaration of
Helsinki Ethical Principles for Medical Research Involving Human
Subjects. The protocol, including consent procedures, was approved
by Kobe University Graduate School of Medicine Ethics Committee.
Tissue specimens were minced and digested in Dulbecco’s modified
Eagle’s medium (DMEM; Gibco BRL, Grand Island, NY, USA) containing
0.2% collagenase (Sigma, St. Louis, MO, USA) for 2 h at 37°C with
5% CO2. The dissociated cells were cultured in DMEM
supplemented with 10% fetal bovine serum (FBS; BioWhittaker,
Walkersville, MD, USA) and 100 U/ml of penicillin/streptomycin.
Following overnight culture, the non-adherent cells were removed,
and the adherent cells were subsequently incubated further in fresh
medium. All experiments were conducted using cells from passages 3
to 4 (4).
RNA extraction
Four individual lines (samples 1–4) of primary
cultured RA-FLS (2×106 cells/well) were incubated with
1.0 μg/ml of recombinant DcR3-Fc protein or control human
IgG1 (R&D Systems, Minneapolis, MN, USA) for 12 h at 37°C with
5% CO2. Following incubation, RNA was extracted using a
QIAshredder and the RNeasy Mini kit (Qiagen, Hilden, Germany)
according to the manufacturer’s instructions. Extraction of total
RNA was performed for each sample separately.
Gene expression profiling and data
analysis
Gene expression was detected by microarray (Human
Genome U133 Plus 2.0, GeneChip® 3′ Expression Array;
Affymetrix, Santa Clara, CA, USA). The labeling of RNA probes,
hybridization and washing were carried out according to the
manufacturer’s instructions.
Avadis 3.3 Prophetic software (Strand Life Sciences,
Bangalore, India) was used for statistical analysis. Differentially
expressed genes were extracted by a paired t-test, with a P-value
<0.05 considered to indicate a statistically significant
difference, and fold change >1.4, and ordered into hierarchical
clusters using the Euclidean algorithm as the distance measure, and
the complete algorithm as the linkage method.
Microarray data have been deposited in NCBIs Gene
Expression Omnibus (GEO) and are accessible through GEO series
accession no. GSE45665.
Results
Microarray analysis (gene expression
profiling of RA-FLS stimulated by DcR3-Fc)
Microarray data analysis revealed that DcR3
upregulated or downregulated the expression of various genes in
RA-FLS. We identified the 100 most differentially regulated genes
in the DcR3-stimulated group compared with the control
IgG1-stimulated group. Among these, 45 genes were upregulated
(Table I) and 55 genes were
downregulated (Table II).
 | Table IThe 45 genes upregulated by DcR3. |
Table I
The 45 genes upregulated by DcR3.
| Gene symbol | Representative
public ID | P-value | Fold change | Gene title |
|---|
| --- | AW612461 | 0.002695443 | 2.0972981 | --- |
| CDH2 | NM_001792 | 0.0327261 | 1.9276143 | Cadherin 2, type 1,
N-cadherin (neuronal) |
| AGPAT9 | BC006236 | 0.006782195 | 1.9184313 |
1-Acylglycerol-3-phosphate
O-acyltransferase 9 |
| LOC440944 | BC036698 | 0.039432783 | 1.8932389 | Hypothetical
protein LOC440944 |
| BIVM | BC039587 | 0.012760683 | 1.6798534 | Basic,
immunoglobulin-like variable motif-containing protein |
| ZC3H3 | D63484 | 0.02914655 | 1.6767992 | Zinc finger
CCCH-type containing 3 |
| IL12B | NM_002187 | 0.008306135 | 1.6463093 | Interleukin 12B
(natural killer cell stimulatory factor 2, cytotoxic lymphocyte
maturation factor 2, p40) |
| NUB1 | AK026433 | 0.045851827 | 1.6456505 | Negative regulator
of ubiquitin-like proteins 1 |
| LTV1 | AW236214 | 0.002476914 | 1.6453595 | LTV1 homolog (S.
cerevisiae) |
| DMRT2 | AF284225 | 0.033502746 | 1.6123677 | Doublesex and mab-3
related transcription factor 2 |
| SUZ12P | AI820796 | 0.021593563 | 1.5686668 | Suppressor of zeste
12 homolog pseudogene |
| ZBTB1 | BU950380 | 0.039786864 | 1.5227357 | Zinc finger and BTB
domain containing 1 |
| --- | N29716 | 0.00277571 | 1.5223095 | --- |
| --- | AK090762 | 0.034467466 | 1.5185958 | --- |
| --- | BG398977 | 0.015083793 | 1.5135463 | --- |
| RANBP17 | NM_022897 | 0.021518266 | 1.5035642 | RAN binding protein
17 |
| GAS5 | BF336936 | 0.041366957 | 1.499078 | Growth
arrest-specific 5 (non-protein coding) |
| --- | BC031996 | 0.011258653 | 1.4903411 | --- |
| REPS2 | AI984607 | 0.012121066 | 1.4882914 | RALBP1 associated
Eps domain containing 2 |
| LOC645158 | BC018088 | 0.013921799 | 1.4854323 | Hypothetical
protein LOC645158 |
| --- | AI332454 | 0.012677074 | 1.4752275 | --- |
| CCDC138 | AU152965 | 0.022518823 | 1.4711432 | Coiled-coil domain
containing 138 |
| hCG_1749898 | BC012486 | 0.002276273 | 1.471024 | KRTAPx protein |
| --- | AI693281 | 0.0196566 | 1.4607961 | --- |
| TUBB2B | AL533838 | 0.006628409 | 1.4584572 | Tubulin, beta
2B |
| SLC9A9 | AA029791 | 0.004298657 | 1.4567653 | Solute carrier
family 9 (sodium/hydrogen exchanger), member 9 |
| --- | AA505135 | 0.023416178 | 1.4560933 | --- |
| SLC16A6 | AI873273 | 0.032126337 | 1.4534916 | Solute carrier
family 16, member 6 (monocarboxylic acid transporter 7) |
| --- | AL080112 | 0.035647828 | 1.4486992 | --- |
| LOC100128988 | AI761436 | 0.035967685 | 1.4464797 | Similar to
hCG2018847 |
| ZNF252 | AU145662 | 0.015798416 | 1.4447424 | Zinc finger protein
252 |
| FGFR1OP2 | R91766 | 0.021976791 | 1.4398756 | FGFR1 oncogene
partner 2 |
| --- | R26931 | 0.03931969 | 1.4398652 | --- |
| ZBTB10 | BG483802 | 0.006168698 | 1.4324573 | Zinc finger and BTB
domain containing 10 |
| C3AR1 | U62027 | 0.031616967 | 1.4275972 | Complement
component 3a receptor 1 |
| ZER1 | NM_006336 | 0.000529948 | 1.4270489 | Zer-1 homolog
(C. elegans) |
| THRB | BF431989 | 0.027043225 | 1.4229552 | Thyroid hormone
receptor, beta (erythroblastic leukemia viral (v-erb-a) oncogene
homolog 2, avian) |
| FLJ35220 | AI311040 | 0.008202907 | 1.422941 | Hypothetical
protein FLJ35220 |
| CDH10 | NM_006727 | 0.018871933 | 1.4211183 | Cadherin 10, type 2
(T2-cadherin) |
| PEX13 | BC040953 | 0.00670011 | 1.4182297 | Peroxisomal
biogenesis factor 13 |
| C7orf58 | NM_024913 | 0.04462639 | 1.4087468 | Chromosome 7 open
reading frame 58 |
| DOK3 | BC004564 | 0.039809346 | 1.4068991 | Docking protein
3 |
| --- | CA776505 | 0.038875684 | 1.4046313 | --- |
| EGR3 | NM_004430 | 0.008657368 | 1.4038599 | Early growth
response 3 |
| ZNF681 | BG281940 | 0.009279283 | 1.4019198 | Zinc finger protein
681 |
| Table IIThe 55 genes downregulated by
DcR3. |
Table II
The 55 genes downregulated by
DcR3.
| Gene symbol | Representative
public ID | P-value | Fold change | Gene title |
|---|
| TPH1 | NM_004179 | 0.022398373 | 2.4520018 | Tryptophan
hydroxylase 1 |
| TREML4 | AK090633 | 0.028559439 | 1.9765018 | Triggering receptor
expressed on myeloid cells-like 4 |
| CEP70 | AI285884 | 0.038745213 | 1.8690827 | Centrosomal protein
70 kDa |
| CCNB2 | AK023404 | 0.040380865 | 1.8138049 | Cyclin B2 |
| CCDC121 | NM_024584 | 0.023371078 | 1.8025432 | Coiled-coil domain
containing 121 |
| ZNF563 | NM_145276 | 0.01811626 | 1.7816929 | Zinc finger protein
563 |
| PANK2 | AV703394 | 0.025317192 | 1.769196 | Pantothenate kinase
2 |
| --- | N46436 | 0.047841128 | 1.7669531 | --- |
| ZFP28 | AW590434 | 0.011407225 | 1.7393188 | Zinc finger protein
28 homolog (mouse) |
| LOC284926 | BG828817 | 0.04181046 | 1.7324702 | Hypothetical
protein LOC284926 |
| SLC24A1 | AF026132 | 0.001774447 | 1.7150456 | Solute carrier
family 24 (sodium/potassium/calcium exchanger), member 1 |
| --- | AU146924 | 0.04154287 | 1.699999 | --- |
| SOS2 | L20686 | 0.019266233 | 1.6804754 | Son of sevenless
homolog 2 (Drosophila) |
| UBE3B | AL096740 | 0.048379965 | 1.6700492 | Ubiquitin protein
ligase E3B |
| ELL | AL521391 | 0.03850427 | 1.6655054 | Elongation factor
RNA polymerase II |
| SEZ6L2 | AF131749 | 0.008593195 | 1.6527929 | Seizure related 6
homolog (mouse)-like 2 |
| MB | NM_005368 | 0.024074124 | 1.650637 | Myoglobin |
| --- | BC040628 | 0.049794715 | 1.6501505 | --- |
| MCOLN3 | NM_018298 | 0.030422723 | 1.6494503 | Mucolipin 3 |
| --- | AK021551 | 0.04994646 | 1.6114371 | --- |
| --- | BF062156 | 0.03806778 | 1.5880637 | --- |
| FBXL17 | AW002273 | 0.003356424 | 1.5799221 | F-box and
leucine-rich repeat protein 17 |
| ZNF117 | BF107006 | 0.048431396 | 1.5662925 | Zinc finger protein
117 |
| C1orf230 | AV746331 | 0.007892164 | 1.5605123 | Chromosome 1 open
reading frame 230 |
| --- | AA650017 | 0.049496956 | 1.5536357 | --- |
| VKORC1L1 | NM_173517 | 0.01724287 | 1.5391829 | Vitamin K epoxide
reductase complex, subunit 1-like 1 |
| MIR155HG | BG231961 | 0.026413728 | 1.5365719 | MIR155 host gene
(non-protein coding) |
| KCNAB1 | L39833 | 0.024623908 | 1.534346 | Potassium
voltage-gated channel, shaker-related subfamily, beta member 1 |
| --- | AI870634 | 0.048582062 | 1.5243825 | --- |
| YTHDC2 | AW975818 | 0.008661097 | 1.5151424 | YTH domain
containing 2 |
| CCNO | BC004877 | 0.012114909 | 1.5041811 | Cyclin O |
| C5orf24 | AW068615 | 0.029783456 | 1.5023562 | Chromosome 5 open
reading frame 24 |
| --- | AV648424 | 0.032394256 | 1.5009412 | --- |
| ADRBK2 | NM_005160 | 0.034059193 | 1.4894675 | Adrenergic, beta,
receptor kinase 2 |
| PIH1D2 | AI744716 | 0.0245486 | 1.477943 | PIH1 domain
containing 2 |
| --- | BF224218 | 0.036218014 | 1.4755102 | --- |
| --- | BF591554 | 0.042058036 | 1.474276 | --- |
| GRIN2A | N48896 | 0.006276551 | 1.4638788 | Glutamate receptor,
ionotropic, N-methyl D-aspartate 2A |
|
RABL2A///RABL2B | NM_007082 | 0.00963415 | 1.4549305 | RAB, member of RAS
oncogene family-like 2A///RAB, member of RAS oncogene family-like
2B |
| MYH14 | BC000676 | 0.009328251 | 1.4530606 | Myosin, heavy chain
14 |
| LOC727820 | AW340595 | 0.018635018 | 1.4494766 | Hypothetical
protein LOC727820 |
| --- | AW074143 | 0.04609772 | 1.448378 | --- |
| ATP5G2 | X69909 | 0.005548685 | 1.4429137 | ATP synthase,
H+ transporting, mitochondrial F0 complex, subunit C2
(subunit 9) |
| MDM4 | AW269813 | 0.031021323 | 1.4353052 | Mdm4 p53 binding
protein homolog (mouse) |
| C11orf84 | AI866590 | 0.02536254 | 1.433334 | Chromosome 11 open
reading frame 84 |
| STYX | AW968935 | 0.000515506 | 1.4238193 |
Serine/threonine/tyrosine interacting
protein |
| S100A14 | NM_020672 | 0.03042584 | 1.4226245 | S100 calcium
binding protein A14 |
| --- | BE549780 | 0.020434508 | 1.4224106 | --- |
| NEURL4 | AL136870 | 0.03654641 | 1.4173305 | Neuralized homolog
4 (Drosophila) |
| --- | AI803010 | 0.009107939 | 1.4132907 | --- |
| ETV7 | AF218365 | 0.007053576 | 1.4130081 | Ets variant 7 |
| RBBP9 | AL121893 | 0.01381168 | 1.4109538 | Retinoblastoma
binding protein 9 |
| --- | AI298755 | 0.048068948 | 1.4096705 | --- |
| LOC100130855 | AK093077 | 0.008585751 | 1.4052048 | Hypothetical
protein LOC100130855 |
| TSFM | AI796813 | 0.005840706 | 1.4036938 | Ts translation
elongation factor, mitochondrial |
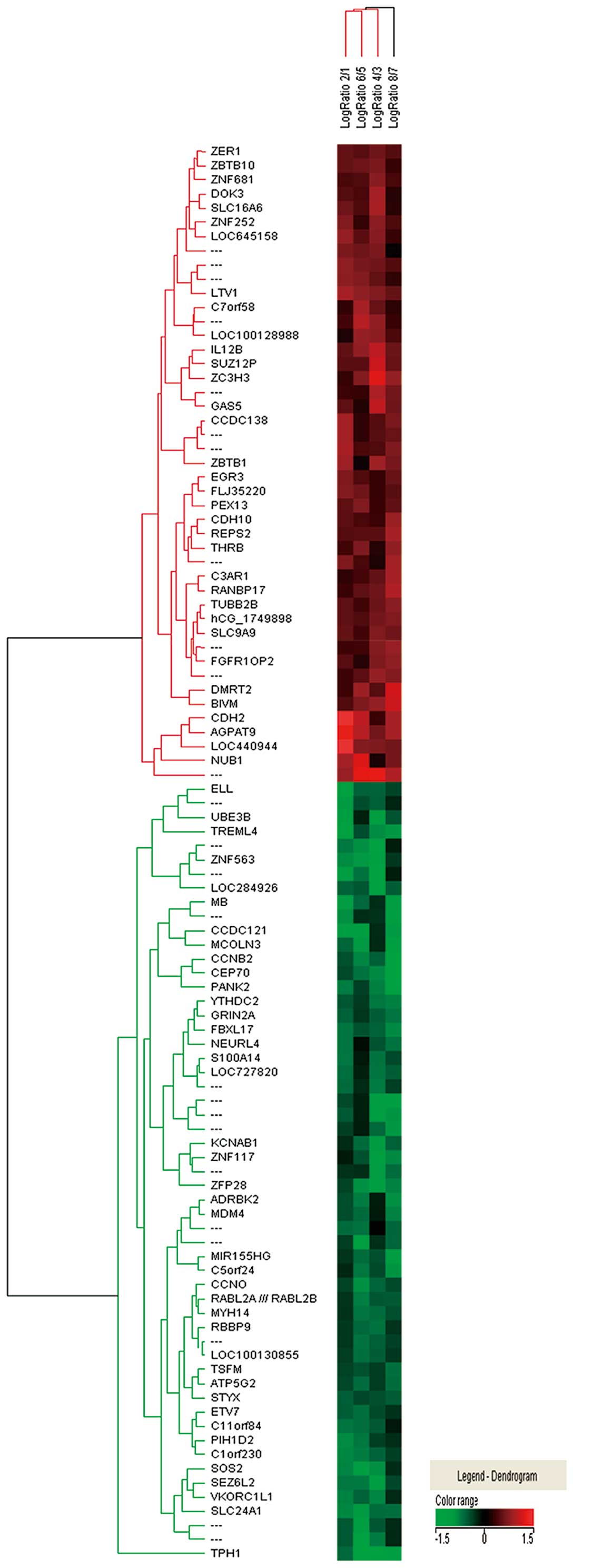
Hierarchical clustering analysis
The upregulated and downregulated genes were
classified into 7 and 10 categories according to their biological
functions, respectively (Fig. 1).
The upregulated genes were associated with protein complex
assembly, cell motility, regulation of transcription, cellular
protein catabolic processes, cell membrane, nucleotide binding and
glycosylation. The upregulated genes belonging to each cluster are
listed in Table III. The
downregulated genes were associated with transcription regulator
activity, RNA biosynthetic processes, cytoskeleton, zinc finger
region, protein complex assembly, phosphate metabolic processes,
mitochondrion, ion transport, nucleotide binding and cell
fractionation. The downregulated genes belonging to each cluster
are listed in Table IV.
| Table IIIFunctional categories of the 45
upregulated genes classified into 7 categories. |
Table III
Functional categories of the 45
upregulated genes classified into 7 categories.
| Functional
category | Genes |
|---|
| Protein complex
assembly | CDH2, RANBP17,
REPS2, TUBB2B, PEX13 |
| Cell motility | CDH2, IL12B,
PEX13 |
| Regulation of
transcription | CDH2, ZC3H3, NUB1,
DMRT2, ZBTB1, REPS2, SLC9A9, ZBTB10, THRB, CDH10, EGR3, ZNF681 |
| Cellular protein
catabolic processes | ZER1 |
| Cell membrane | CDH2, AGPAT9,
RANBP17, SLC9A9, SLC16A6, C3AR1, CDH10, PEX13, DOK3 |
| Nucleotide
binding | RANBP17,
TUBB2B, |
| Glycosylation | CDH2, IL12B,
SLC9A9, C3AR1, CDH10, C7orf58 |
| Table IVFunctional categories of the 55
downregulated genes classified into 10 categories. |
Table IV
Functional categories of the 55
downregulated genes classified into 10 categories.
| Functional
category | Genes |
|---|
| Transcription
regulator activity | ZNF563, ZFP28, ELL,
ZNF117, CCNO, MDM4, ETV7, TSFM |
| RNA biosynthetic
processes | ELL, ZNF117, ETV7,
TSFM |
| Cytoskeleton | Cep70, CCNB2,
GRIN2A, MYH14 |
| Zinc finger
region | TPH1, ZNF563,
ZFP28, SLC24A1, SOS2, ELL, MB, ZNF117, KCNAB1, GRIN2A, MDM4,
S100A14, ETV7, RBBP9, TSFM |
| Protein complex
assembly | MDM4 |
| Phosphate metabolic
processes | ADRBK2, ATP5G2,
STYX |
| Mitochondrion | PANK2, ATP5G2,
TSFM |
| Ion transport | TREML4, SLC24A1,
SEZ6L2, MB, MCOLN3, VKORC1L1, KCNAB1, GRIN2A, ATP5G2 |
| Nucleotide
binding | PANK2, YTHDC2,
ADRBK2, RABL2A///RABL2B, MYH14 |
| Cell
fractionation | CCNB2, SLC24A1,
GRIN2A |
Discussion
Among the 3 ligands of DcR3, TL1A (TNFSF15) is
expressed by endothelial cells (12), macrophages (18,19), T cells (20,21), monocytes (22,23), dendritic cells (23), chondrocytes (24) and synovial fibroblasts (24), and contributes to the pathogenesis
of cancer and autoimmune diseases via the apoptotic, stress,
mitogenic and inflammation pathway by binding death receptor 3
(DR3) and DcR3 (12,25). The 3 ligands of DcR3 have been
reported to contribute to the pathogenesis of RA (4,24,26,27). In these studies, DcR3 was
considered a decoy receptor for ligands. We previously demonstrated
that DcR3 binds to membrane-bound TL1A expressed on RA-FLS when it
acts as a ligand in the pathogenesis of RA (16).
Genome-wide gene expression cDNA microarray is a
powerful technique used to investigate the pathophysiology of a
variety of diseases, including tumors (28–30), immune-mediated diseases (31,32) and inflammatory diseases (33–35). Using microarray, Chang et
al revealed that genes characteristically expressed by
tumor-associated macrophages were upregulated by DcR3 (30). In the current study, we first
demonstrated the expression profiles of genes in RA-FLS regulated
by DcR3.
We demonstrated that DcR3 regulates the expression
of genes that are mainly associated with the upregulation of the
protein complex assembly, cell motility and the regulation of
transcription, and the downregulation of transcription regulator
activity, RNA biosynthetic processes and cytoskeleton. We then
focused on the following genes: cadherin 2, type 1, N-cadherin
(neuronal) (CDH2), interleukin 12B (natural killer cell stimulatory
factor 2, cytotoxic lymphocyte maturation factor 2, p40) (IL12B),
tryptophan hydroxylase 1 (TPH1), centrosomal protein 70 kDa (Cep70)
and Zinc finger proteins as these genes were highly regulated,
either upregulated or downregulated, and belonged to major
functional clustering categories.
As for each gene, CDH2 has been reported to be
associated with cell attachment and migration (36), metastatic potential (37), osteoblast differentiation
(38) and the proliferation of
RA-FLS (39).
IL12B encodes the IL-12B p40 subunit of IL-12 and
IL-23 cytokines. IL-12 induces Th1 immune responses, and is thus
linked with autoimmune diseases (40), while IL-23 is linked with
autoimmune diseases via Th17 immune responses (41). IL-12 (42) and IL-23 (43,44) have also been reported to be
involved in the pathogenesis of RA.
TPH1 is a rate-limiting enzyme involved in the
synthesis of serotonin, and has been reported to be associated with
the pathogenesis of RA through the inflammatory pathway (45) and bone biology (46–48).
Cep70 was discovered in a proteomic study of the
centrosome (49). Centrosomal
activity is indispensable for the execution of cytokinesis and the
progression of the cell cycle (50). Cep70 is crucial for mitotic
spindle assembly (51) and
promotes microtubule polymerization by increasing microtubule
elongation (52).
Zinc finger proteins are involved in a broad range
of biological activities, including double-stranded DNA binding,
single-stranded DNA and RNA recognition, as well as coordinating
protein-protein interactions (53).
In the current study, we first reported the
expression profiles of genes in RA-FLS regulated by DcR3. Combined
with our previous findings that DcR3 serves as a ligand by binding
to membrane-bound TL1A on RA-FLS, our data demonstrate that DcR3
may regulate the gene expression of various key molecules in RA-FLS
by binding to TL1A, thus affecting the pathogenesis of RA, such as
proliferation, apoptosis, inflammation and bone biology. Further
studies on the genes detected in the current study may provide a
deeper understanding of the pathogenesis and treatment of RA by
DcR3-TL1A signaling.
Acknowledgements
The authors thank Ms. Kyoko Tanaka, Ms. Minako
Nagata, and Ms. Maya Yasuda for providing technical assistance.
This study was supported by a Grant-in-Aid from the Health Science
Research Grant of the Japanese Ministry of Education, Science and
Culture (no. 24592261).
References
|
1
|
Chou CT, Yang JS and Lee MR: Apoptosis in
rheumatoid arthritis - expression of Fas, Fas-L, p53, and Bcl-2 in
rheumatoid synovial tissues. J Pathol. 193:110–116. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tak PP, Zvaifler NJ, Green DR and
Firestein GS: Rheumatoid arthritis and p53: how oxidative stress
might alter the course of inflammatory diseases. Immunol Today.
21:78–82. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yamanishi Y, Boyle DL, Rosengren S, Green
DR, Zvaifler NJ and Firestein GS: Regional analysis of p53
mutations in rheumatoid arthritis synovium. Proc Natl Acad Sci USA.
99:10025–10030. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hayashi S, Miura Y, Nishiyama T, et al:
Decoy receptor 3 expressed in rheumatoid synovial fibroblasts
protects the cells against Fas-induced apoptosis. Arthritis Rheum.
56:1067–1075. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pitti RM, Marsters SA, Lawrence DA, et al:
Genomic amplification of a decoy receptor for Fas ligand in lung
and colon cancer. Nature. 396:699–703. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bai C, Connolly B, Metzker ML, et al:
Overexpression of M68/DcR3 in human gastrointestinal tract tumors
independent of gene amplification and its location in a four-gene
cluster. Proc Natl Acad Sci USA. 97:1230–1235. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ohshima K, Haraoka S, Sugihara M, et al:
Amplification and expression of a decoy receptor for fas ligand
(DcR3) in virus (EBV or HTLV-I) associated lymphomas. Cancer Lett.
160:89–97. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen J, Zhang L and Kim S: Quantification
and detection of DcR3, a decoy receptor in TNFR family. J Immunol
Methods. 285:63–70. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi G, Wu Y, Zhang J and Wu J: Death decoy
receptor TR6/DcR3 inhibits T cell chemotaxis in vitro and in vivo.
J Immunol. 171:3407–3414. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tsuji S, Hosotani R, Yonehara S, et al:
Endogenous decoy receptor 3 blocks the growth inhibition signals
mediated by Fas ligand in human pancreatic adenocarcinoma. Int J
Cancer. 106:17–25. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu KY, Kwon B, Ni J, Zhai Y, Ebner R and
Kwon BS: A newly identified member of tumor necrosis factor
receptor superfamily (TR6) suppresses LIGHT-mediated apoptosis. J
Biol Chem. 274:13733–13736. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Migone TS, Zhang J, Luo X, et al: TL1A is
a TNF-like ligand for DR3 and TR6/DcR3 and functions as a T cell
costimulator. Immunity. 16:479–492. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang CR, Wang JH, Hsieh SL, Wang SM, Hsu
TL and Lin WW: Decoy receptor 3 (DcR3) induces osteoclast formation
from monocyte/macrophage lineage precursor cells. Cell Death
Differ. 11(Suppl 1): S97–S107. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hsu MJ, Lin WW, Tsao WC, et al: Enhanced
adhesion of monocytes via reverse signaling triggered by decoy
receptor 3. Exp Cell Res. 292:241–251. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tateishi K, Miura Y, Hayashi S, Takahashi
M and Kurosaka M: DcR3 protects THP-1 macrophages from apoptosis by
increasing integrin alpha4. Biochem Biophys Res Commun.
389:593–598. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Takahashi M, Miura Y, Hayashi S, Tateishi
K, Fukuda K and Kurosaka M: DcR3-TL1A signalling inhibits
cytokine-induced proliferation of rheumatoid synovial fibroblasts.
Int J Mol Med. 28:423–427. 2011.PubMed/NCBI
|
|
17
|
Arnett FC, Edworthy SM, Bloch DA, et al:
The American Rheumatism Association 1987 revised criteria for the
classification of rheumatoid arthritis. Arthritis Rheum.
31:315–324. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kamada N, Hisamatsu T, Honda H, et al:
TL1A produced by lamina propria macrophages induces Th1 and Th17
immune responses in cooperation with IL-23 in patients with Crohn’s
disease. Inflamm Bowel Dis. 16:568–575. 2010.PubMed/NCBI
|
|
19
|
Bamias G, Martin C III, Marini M, et al:
Expression, localization, and functional activity of TL1A, a novel
Th1-polarizing cytokine in inflammatory bowel disease. J Immunol.
171:4868–4874. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Prehn JL, Mehdizadeh S, Landers CJ, et al:
Potential role for TL1A, the new TNF-family member and potent
costimulator of IFN-gamma, in mucosal inflammation. Clin Immunol.
112:66–77. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Papadakis KA, Zhu D, Prehn JL, et al:
Dominant role for TL1A/DR3 pathway in IL-12 plus IL-18-induced
IFN-gamma production by peripheral blood and mucosal
CCR9+ T lymphocytes. J Immunol. 174:4985–4990. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cassatella MA, Pereira-da-Silva G, Tinazzi
I, et al: Soluble TNF-like cytokine (TL1A) production by immune
complexes stimulated monocytes in rheumatoid arthritis. J Immunol.
178:7325–7333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Prehn JL, Thomas LS, Landers CJ, Yu QT,
Michelsen KS and Targan SR: The T cell costimulator TL1A is induced
by FcgammaR signaling in human monocytes and dendritic cells. J
Immunol. 178:4033–4038. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang J, Wang X, Fahmi H, et al: Role of
TL1A in the pathogenesis of rheumatoid arthritis. J Immunol.
183:5350–5357. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sethi G, Sung B and Aggarwal BB:
Therapeutic potential of VEGI/TL1A in autoimmunity and cancer. Adv
Exp Med Biol. 647:207–215. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bamias G, Siakavellas S, Stamatelopoulos
K, Chryssochoou E, Papamichael C and Sfikakis P: Circulating levels
of TNF-like cytokine 1A (TL1A) and its decoy receptor 3 (DcR3) in
rheumatoid arthritis. Clin Immunol. 129:249–255. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Edwards JR, Sun SG, Locklin R, et al:
LIGHT (TNFSF14), a novel mediator of bone resorption, is elevated
in rheumatoid arthritis. Arthritis Rheum. 54:1451–1462. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khan J, Simon R, Bittner M, et al: Gene
expression profiling of alveolar rhabdomyosarcoma with cDNA
microarrays. Cancer Res. 58:5009–5013. 1998.PubMed/NCBI
|
|
29
|
Espinosa I, Catasus L, Canet B, D’Angelo
E, Munoz J and Prat J: Gene expression analysis identifies two
groups of ovarian high-grade serous carcinomas with different
prognosis. Mod Pathol. 24:846–854. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chang YC, Chen TC, Lee CT, et al:
Epigenetic control of MHC class II expression in tumor-associated
macrophages by decoy receptor 3. Blood. 111:5054–5063. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li J, Yang S, Lu S, et al: Differential
gene expression profile associated with the abnormality of bone
marrow mesenchymal stem cells in aplastic anemia. PLoS One.
7:e477642012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Whitney LW, Becker KG, Tresser NJ, et al:
Analysis of gene expression in mutiple sclerosis lesions using cDNA
microarrays. Ann Neurol. 46:425–428. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Heller RA, Schena M, Chai A, et al:
Discovery and analysis of inflammatory disease-related genes using
cDNA microarrays. Proc Natl Acad Sci USA. 94:2150–2155. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee SK, Jeon EK, Kim YJ, et al: A global
gene expression analysis of the peripheral blood mononuclear cells
reveals the gene expression signature in psoriasis. Ann Dermatol.
21:237–242. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
van der Pouw Kraan TC, van Gaalen FA,
Kasperkovitz PV, et al: Rheumatoid arthritis is a heterogeneous
disease: evidence for differences in the activation of the STAT-1
pathway between rheumatoid tissues. Arthritis Rheum. 48:2132–2145.
2003.PubMed/NCBI
|
|
36
|
Akitaya T and Bronner-Fraser M: Expression
of cell adhesion molecules during initiation and cessation of
neural crest cell migration. Dev Dyn. 194:12–20. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kashima T, Nakamura K, Kawaguchi J, et al:
Overexpression of cadherins suppresses pulmonary metastasis of
osteosarcoma in vivo. Int J Cancer. 104:147–154. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Marie PJ: Role of N-cadherin in bone
formation. J Cell Physiol. 190:297–305. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nonomura Y, Mizoguchi F, Suzuki A, et al:
Hypoxia-induced abrogation of contact-dependent inhibition of
rheumatoid arthritis synovial fibroblast proliferation. J
Rheumatol. 36:698–705. 2009. View Article : Google Scholar
|
|
40
|
Hasko G and Szabo C: IL-12 as a
therapeutic target for pharmacological modulation in
immune-mediated and inflammatory diseases: regulation of T helper
1/T helper 2 responses. Br J Pharmacol. 127:1295–1304. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Paradowska-Gorycka A, Grzybowska-Kowalczyk
A, Wojtecka-Lukasik E and Maslinski S: IL-23 in the pathogenesis of
rheumatoid arthritis. Scand J Immunol. 71:134–145. 2010. View Article : Google Scholar
|
|
42
|
Swaak AJ, van den Brink HG and Aarden LA:
Cytokine production in whole blood cell cultures of patients with
rheumatoid arthritis. Ann Rheum Dis. 56:693–695. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu FL, Chen CH, Chu SJ, et al:
Interleukin (IL)-23 p19 expression induced by IL-1beta in human
fibroblast-like synoviocytes with rheumatoid arthritis via active
nuclear factor-kappaB and AP-1 dependent pathway. Rheumatology
(Oxford). 46:1266–1273. 2007. View Article : Google Scholar
|
|
44
|
Kim HR, Cho ML, Kim KW, et al:
Up-regulation of IL-23p19 expression in rheumatoid arthritis
synovial fibroblasts by IL-17 through PI3-kinase-, NF-kappaB- and
p38 MAPK-dependent signalling pathways. Rheumatology (Oxford).
46:57–64. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kular L, Pakradouni J, Kitabgi P, Laurent
M and Martinerie C: The CCN family: a new class of inflammation
modulators? Biochimie. 93:377–388. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yadav VK and Ducy P: Lrp5 and bone
formation. Ann NY Acad Sci. 1192:103–109. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gustafsson BI, Thommesen L, Stunes AK, et
al: Serotonin and fluoxetine modulate bone cell function in vitro.
J Cell Biochem. 98:139–151. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ducy P and Karsenty G: The two faces of
serotonin in bone biology. J Cell Biol. 191:7–13. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Andersen JS, Wilkinson CJ, Mayor T,
Mortensen P, Nigg EA and Mann M: Proteomic characterization of the
human centrosome by protein correlation profiling. Nature.
426:570–574. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Doxsey S, Zimmerman W and Mikule K:
Centrosome control of the cell cycle. Trends Cell Biol. 15:303–311.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Shi X, Sun X, Liu M, Li D, Aneja R and
Zhou J: CEP70 protein interacts with gamma-tubulin to localize at
the centrosome and is critical for mitotic spindle assembly. J Biol
Chem. 286:33401–33408. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Shi X, Wang J, Yang Y, Ren Y, Zhou J and
Li D: Cep70 promotes microtubule assembly in vitro by increasing
microtubule elongation. Acta Biochim Biophys Sin (Shanghai).
44:450–454. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Leon O and Roth M: Zinc fingers: DNA
binding and protein-protein interactions. Biol Res. 33:21–30. 2000.
View Article : Google Scholar : PubMed/NCBI
|