Introduction
Endometriosis, characterized by the growth of the
endometrial gland and stroma outside the uterine cavity, is a
gynecological disorder affecting women of reproductive age
worldwide (1,2). Endometriotic implants arise from
retrograde menstruation of endometrial tissue through the fallopian
tubes into the peritoneal cavity. Distribution of this ectopic
tissue occurs most often within the pelvic peritoneum, but may also
occur in the pelvic viscera, rectovaginal spectrum, pleura,
abdominal wall, and rarely, the brain (3). Endometriosis affects 6–10% of women
of reproductive age and its symptoms include dysmenorrhea,
dyspareunia, chronic pelvic pain, irregular uterine bleeding and/or
infertility (4).
In endometrial tissue, the ectopic (extrauterine)
endometrium (ECE) of women with endometriosis has a similar
histological nature compared with its eutopic (intrauterine)
endometrium (EUE), although there are a number of different
biochemical and functional characteristics (5–7).
Previous studies showed that endometrium from women with
endometriosis has abnormalities in the structures, proliferation,
immune components, adhesion molecules, proteolytic enzymes and
inhibitors, steroid and cytokine production and responsiveness, as
well as mRNA expression and protein production when compared with
endometrium of women without endometriosis (8–11).
The association between the etiology and endometriosis remains to
be elucidated, however, key pathogenesis might be the
characteristics of refluxed endometrium and the peritoneal
microenvironment. To gain a better understanding of the factors
involved in the onset of endometriosis, the molecular mechanisms of
endometriosis should be investigated.
Currently, proteomic analysis is the preferred tool
used for understanding this disease as it enables the estimation of
a number of proteins in complicated protein mixtures and
quantitative distinctions of the alterations in protein profiles
(10,12,13). In addition, the combinatorial
system of two-dimensional electrophoresis (2-DE) and mass
spectrometry (MS) has been extensively applied to scan for
differentially expressed proteins. Proteomics and mass spectrometry
have been previously used to investigate endometriosis using
various sample types including serum, peritoneal fluid, eutopic and
ectopic endometrial tissue, and endometrial fluid (14–18).
The aim of this study was to investigate the
characteristic of endometriosis on a molecular level by using 2-DE
combined with MS. Global proteins in ectopic endometrial tissue
with endometriosis were compared with those from the normal
endometrial tissue. The present study may provide insight into the
pathogenesis of endometriosis as well as understanding of molecular
mechanisms involved in the progression of this disease.
Materials and methods
Subjects
This study was approved by the ethics committee of
the Institutional Review Board at the Pusan National University
Hospital (PNUH IRB 2010144). Patients and controls were required to
sign a consent form prior to participation in the study. Eutopic
endometrial tissues were collected from the patients who underwent
laparoscopy or laparotomy for myomectomy or benign ovarian cyst
other than endometriosis, such as mature cystic teratoma,
functional cyst and hemorrhagic cyst (ages 25–48; n=6). Patients
presenting with abdominal/pelvic pain, dysmenorrhea and/or
subfertility underwent laparoscopy and were histologically
diagnosed as endometriosis. These patients (aged 27–40; n=13)
comprised the endometriosis group. Samples were collected during
the proliferative phase. Biopsy specimens were obtained for to
histologically confirm the diagnosis and the extent of the disease
was staged according to the revised American Fertility Society
(rAFS) classification. The tissue samples were frozen in liquid
nitrogen and preserved at −80°C until required.
Protein sample preparation
Ectopic endometrial tissue samples stored at −80°C
were thawed, weighed and combined with lysis buffer containing 7 M
urea, 2 M thiourea, 4% w/v
3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate (CHAPS),
50 mM dithiothreitol (DTT), 0.5% (v/v) pharmalyte (pH 3–10NL) and
protease inhibitor (GE Healthcare, Piscataway, NJ, USA). The
tissues were homogenized and then agitated every 5 min for 1 h,
after which the samples were centrifuged at 17,000 rpm for 30 min
at 20°C. The supernatant was quantified by the 2-D Quant kit (GE
Healthcare) and stored at −80°C until required for further
analysis.
2-DE and protein identification by
electrospray ionization-quadrupole-time of flight/mass spectrometer
ESI-Q-TOF/MS
Individual protein samples were divided into the
endometriosis and control groups. The electrophoretic separation of
these proteins was performed as previously described (19). Briefly, proteins were diluted into
isoelectric focusing (IEF) buffer and then 100 μg of proteins were
loaded onto the pH 3–10 NL Immobiline DryStrip gels (GE Healthcare;
18 cm). The gels were used to rehydrate the strips at 20°C for 12
h. IEF was performed with an IEF electrophoresis unit (GE
Healthcare) after 12-h rehydration. The focusing conditions started
at a linear ramp from 500 to 1000 V for 1 h and a constant voltage
of 8000 V for 6 h to give a total voltage of 56,000 Vh. After
focusing, each strip was equilibrated for 15 min in an
equilibration buffer containing 50 mM Tris-HCl (pH 8.8), 6 M urea,
2% SDS, 30% glycerol and 0.002% (w/v) bromophenol blue. The first
equilibration buffer contained 1% DTT and the second 135 mM
iodoacetamide (IAA). The Ettan DALT 2-D gel system (GE Healthcare)
was used for electrophoresis along the second dimension. The
equilibrated strips were inserted into 12% SDS-PAGE gels (18 cm)
and the separated gels were stained using a PlusOne Silver staining
kit (GE Healthcare). Spot detection, pair matching and
normalization were carried out using the ProteomWeaver software 2.2
(Definiens, Munich, Germany). The ratios of the spots with ≥2-fold
changes in staining intensity were selected for ESI-Q-TOF/MS
analysis. Details of the ESI-Q-TOF/MS analysis have been described
in a previous study (20).
Western blot analysis
A total of 11 individual samples in endometriosis
group (rAFS II =1, rAFS III =4, rAFS IV =6) and 4 samples from
control group were used for western blotting. Tissue protein was
separated by 10% SDS-PAGE gel electrophoresis and then transferred
for 30 min onto nitrocellulose membranes by using a
Trans-blot® SD Semi-dry Transfer cell (Bio-Rad,
Hercules, CA, USA). The membranes (Whatman, Dassel, Germany) were
immediately placed into a 5% non-fat milk solution and blocked for
1 h at room temperature. They were incubated overnight at 4°C with
vitamin D binding protein (DBP) (Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA) and flavin reductase (FR) (Santa Cruz
Biotechnology, Inc.) antibody at a dilution of 1:125 and 1:500,
respectively, in TBST. The membranes were then washed three times
with 1X TBST and incubated with horseradish peroxidase-conjugated
donkey anti-rabbit IgG (Thermo scientific, Rockford, IL, USA) and
rabbit anti-goat IgG (Abcam, Cambridge, MA, USA) at 1:10,000 and
1:1,000 dilution for 20 min at room temperature. The proteins on
the membrane were visualized by an enhanced chemiluminescence
system (ECL) detection kit (Surmodics, Eden Prairie, MN, USA) and
bands were quantified using Image J 1.43 software (http://rsb.info.nih.gov/ij/download.html). The protein
levels were normalized to those of β-actin signal on the same
membrane.
Statistical analysis
Data were expressed as means ± SEM. Comparisons of
two groups were analyzed by the Student’s t-test and those of
multiple groups were analyzed by the Tukey HSD Kramer comparison
test. JMP 7.0.1 (SAS Institute Inc., Cary, NC, USA) was used for
statistical analyses. P<0.05 was considered to indicate
statistical significance.
Results
Protein identification in ECE tissue from
women with endometriosis
A total of 406 spots were detected in the
endometriosis group (n=13) and a total of 385 spots were identified
in the control group (n=6). Among these proteins, 50 spots were
found to be differentially expressed with spot density changes of
approximately ≥2-fold. Twenty spots were decreased whereas 15 spots
were increased in the endometriosis group compared with those in
the control group. Moreover, eight spots were expressed only in the
control group and seven spots were expressed only in the
endometriosis group (Fig. 1).
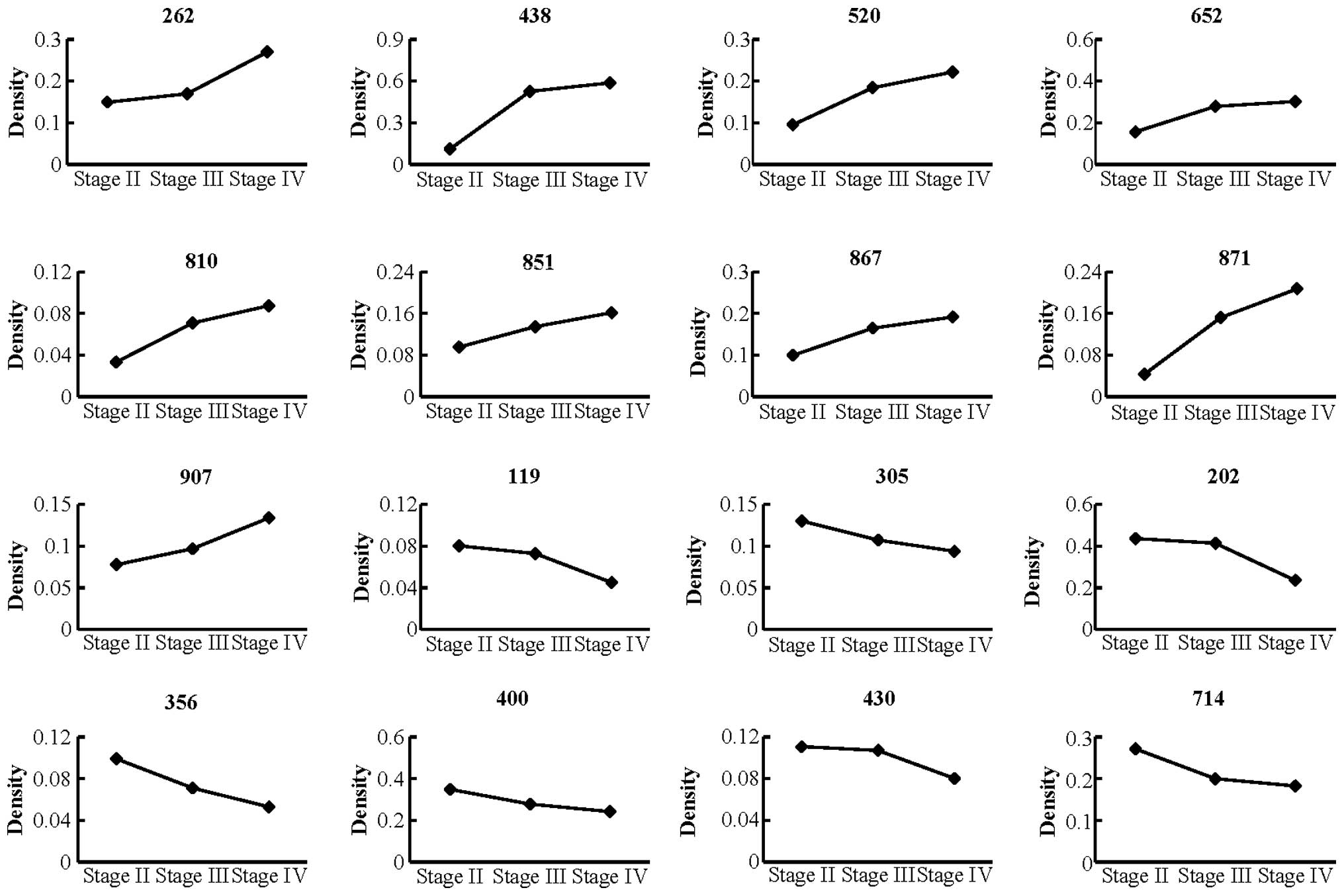
Sixteen selected spots were compared according to clinical stage
(stage II=1, III=4 and IV=8) of endometriosis through 2-DE analysis
(Fig. 2). Among these spots, the
density of 9 spots gradually increased whereas the density of 7
spots gradually decreased from stage II to IV (Fig. 2). All 16 spots were selected and
identified by ESI-Q-TOF/MS (Table
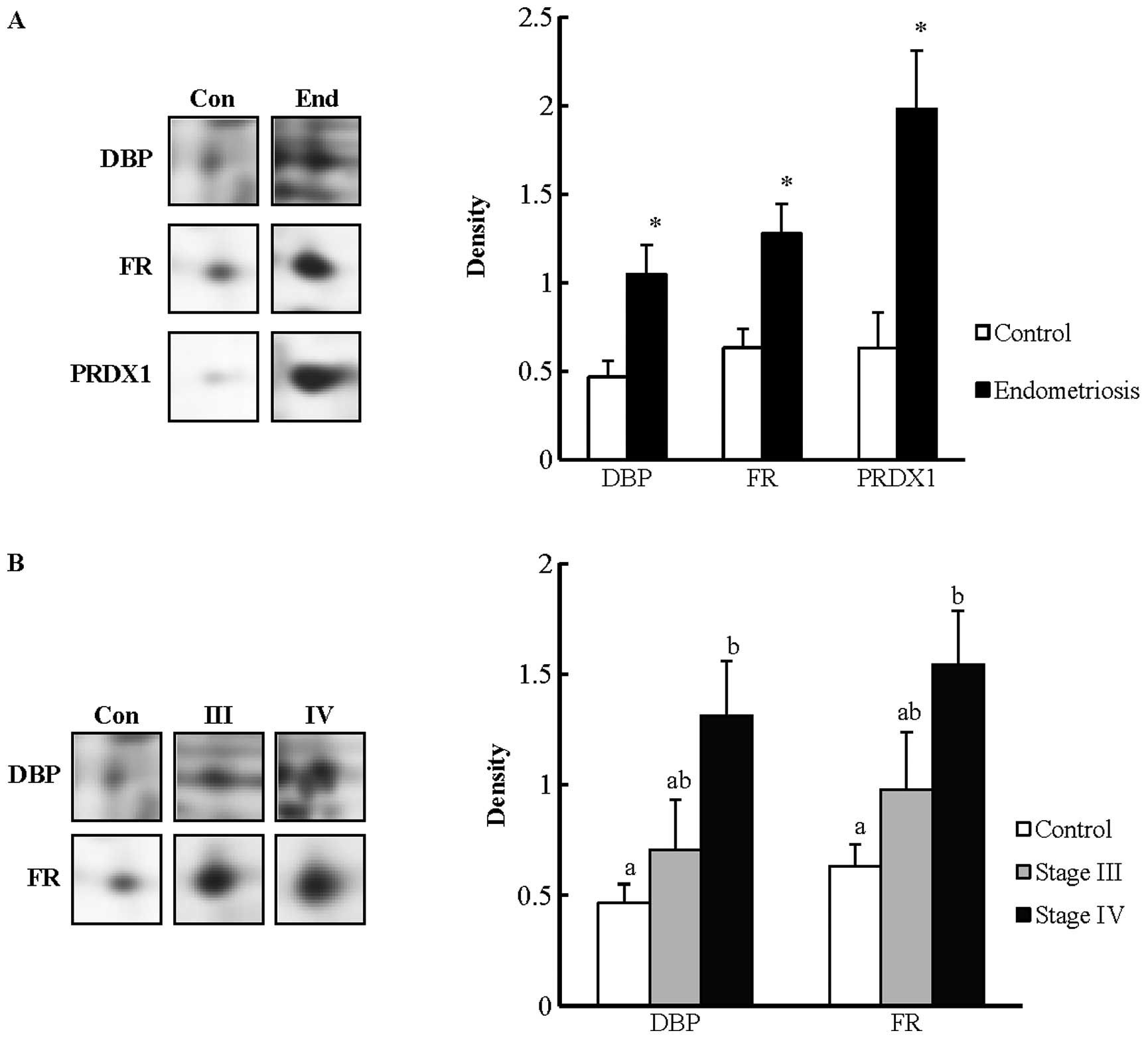
I). The densities of three candidate proteins are shown in
Fig. 3. The three proteins were
statistically increased in the endometriosis group compared with
the control group (P<0.05). The proteins were DBP (520), FR
(867) and peroxiredoxin-1 (PRDX1) (400). In addition, the
expression of those proteins was investigated according to the
clinical stages. The results showed that DBP and FR were statically
significant (P<0.05) and the two proteins were selected as
validation proteins in the subsequent western blotting
analysis.
 | Table IIdentification of differentially
expressed protein spots in ectopic endometrial tissue by ESI/Q-TOF
MS. |
Table I
Identification of differentially
expressed protein spots in ectopic endometrial tissue by ESI/Q-TOF
MS.
| Spot no. | Accession no. | Protein name | Score | Mol. wt.
(kDa)/pI | Expression | Molecular
functions |
|---|
| 119 | gi|74718831 | Glyoxalase
domain | 242 | 34.8/5.40 | Down | Glycolytic
enzyme |
| 202 | gi|62897681 | Calreticulin
precursor | 114 | 48.1/4.29 | Down | Apoptosis
regulation |
| 262 | gi|16418467 | Myosin
regulatory | 6 | 19.8/7.78 | Up | Regulation of both
smooth muscle and nonmuscle cell contractile activity via its
phosphorylation |
| 305 | gi|62089188 | Lactoylglutathione
lyase | 55 | 20.8/5.12 | Down | Ion transport |
| 356 | gi|55664663 | Voltage-dependent
anion | 382 | 31.6/7.50 | Down | Apoptosis
regulation |
| 400 | gi|55959887 |
Peroxiredoxin-1 | 453 | 22.1/8.27 | Down | Apoptosis
regulation |
| 430 | gi|36038 | Rho
GDP-dissociation | 272 | 23.2/5.01 | Down | Apoptosis
regulation |
| 438 | gi|48145547 | Ferritin light
chain | 670 | 20.0/5.50 | Up | Cellular
homeostasis |
| 520 | gi|455970 | Vitamin D-binding
protein | 41 | 53.0/5.40 | Up | Carrier vitamin D
sterols |
| 652 | gi|3641398 | NADP-dependent
isocitrate | 79 | 46.7/6.53 | Down | Citric acid cycle
enzyme |
| 714 | gi|119630158 | Carbonyl
reductase | 89 | 30.4/8.55 | Down |
NADPH-dependent |
| 851 | gi|48255905 | Transgelin
(TAGLN) | 158 | 22.6/8.87 | Up | Calcium
interactions |
| 867 | gi|4502419 | Flavin reductase
(NADPH) | 92 | 22.1/7.13 | Up | Oxidoreductase |
| 871 | gi|48145547 | Ferritin light
chain (FTL) | 446 | 20.0/5.50 | Up | Cellular
homeostasis |
| 907 | gi|5453559 | ATP synthase
subunit d | 15 | 18.5/5.21 | Up | Ion transport |
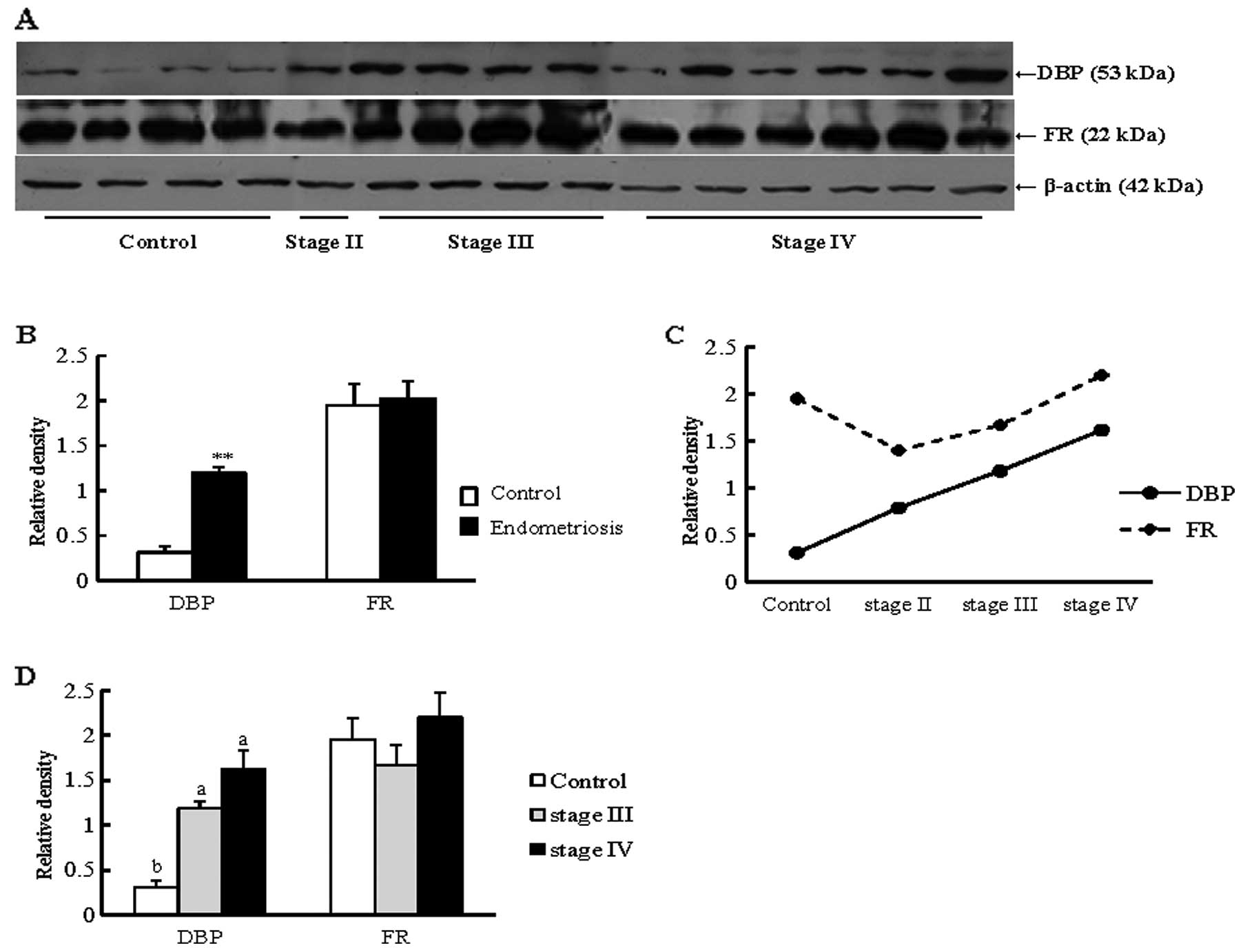
Validation test by western blotting
Results of the western blot analysis revealed that
the expression of DBP was significantly increased in the
endometriosis group when compared with that in the control group
(P<0.01) (Fig. 4A and B). This
protein also showed an increased pattern in stages II and IV of the
endometriosis group compared with the control group (Fig. 4C and D). Another selected
candidate protein, FR, exhibited an increase in the endometriosis
group compared with the control group, although the difference not
significant (Fig. 4).
Discussion
In the present study, 2-DE combined with MS were
used to explore proteins that may be involved in the progression of
endometriosis. Two proteins identified in the ECE tissue were
gradually increased according to disease progression (stage II to
IV) in the 2-DE gel image analysis. Spots were identified by MS as
DBP and FR. In the subsequent validation test, using western
blotting, DBP showed a significantly higher expression level in the
endometriosis group compared with the control group. However, no
significant difference was found for FR.
DBP is ~58 kDa in size and is genetically related to
serum albumin, α-fetoprotein and afamin (21–23). It is a polymorphic serum
glycoprotein with lots of functions that include highly specific
binding of vitamin D sterols, G-actin, fatty acids and chemotactic
agents. It is produced in the liver and is located predominantly in
the serum (24–26). It has two large domains (I and II)
and a shorter domain at the COOH terminus (domain III) (26). Although it is derived from the
plasma (or serum), it is expressed in various tissues and is
synthesized in the cell membrane-associated lymphocytes, monocytes
and neutrophils (26–28). It is associated with macrophage
activation, increases monocyte and neutrophil chemotaxis to
C5-derived peptides and plays a role as an actin scavenger protein
(29,30). However, the connection between DBP
and endometriosis has not been studied in detail, with only a few
papers reporting its expression in endometriosis. In previous
studies, DBP presented various expression aspects in urine,
peritoneal fluid and plasma (31,32). In recent studies, proteomic
analysis of the serum and urine showed that the concentration of
DBP is elevated in endometriosis patients compared with controls
(33,34).
Our results show that the expression of DBP in ECE
tissue is increased in patients with endometriosis in accordance
with rAFS. Hunt et al demonstrated that DBP was synthesized
by DBP gene transcription in liver, kidney and brain (in order of
abundance) (35). DBP was
synthesized in the liver at low levels until 40 weeks after
conception and the expression presumed adult levels of 500 μg/ml in
serum (36). It was also detected
in the lung, heart, stomach, spleen, uterus and brain using PCR of
DBP cDNA (35). Results obtained
in the present study suggest that DBP transcription occurs actively
in ECE and the DBP expression level accelerates with its
deterioration into a deeper stage.
DBP is known as a chemotactic factor that recruits
neutrophils, monocytes and fibroblasts, as it plays an important
role in the immune system (28,37,38). DBP is the precursor of
macrophage-activating factor (MAF). This activity was confirmed
when mouse peritoneal cells were stimulated with
lysophosphatidylcholine, ensuring macrophages with increased
phagocytic activity (29,39). DBP is converted to its active
form, and is partially deglycosylated by β-galactosidase and
sialidase activities of the B- and T-cells, producing DBP-MAF
(29). It is a powerful activator
of macrophage functions and is involved in immune responses. The
conversion of DBP to DBP-MAF may be reduced in malignancies by the
action of α-N-acetylgalactosaminidase (40). Thus, activation of DBP-MAF
suggests that it has a positive effect on the pathogenesis of the
disease. Thus, the action of DBP-MAF may also have a similarly good
impact on endometriosis in the form of cancer. However, several
studies have shown differential DBP-MAF function among DBP
genotypes (41–44). DBP has three major allele products
(GC1F, GC1S and GC2) (47).
Several studies have found that the GC1F form of DBP is correlated
with increased chronic obstructive pulmonary disease (COPD) risk
for Asians, whereas the GC2 form is associated with reduced COPD in
whites (41–43,46,47). In addition to COPD, GC2 was also
associated with increased risk to bronchiectasis (41). Another study revealed that DBP
genotypes have different glycosylation rates. Borges et al
showed that the allele products of GC1 were glycosylated at a total
rate of 10–30% and those of GC2 were glycosylated at a rate of 1–5%
(48). In the endometriosis
study, there were related DBP genotypes. Faserl et al
demonstrated that the DBP allele products in serum were different
because of the reduced ability of DBP to convert to DBP-MAF, the
essential macrophage activator. According to that study, the form
of DBP encoded by the GC1 allele was much more readily converted to
DBP-MAF, while that encoded by GC2 was not readily converted, i.e.,
none of the women in the control group expressed only GC2 allele
products, whereas they were expressed in a much higher percentage
of women with endometriosis. As stated above, overexpression of the
GC2 allele in endometriosis patients may act on the scavenger
function of macrophages (33). In
view of our study, increased DBP expression in endometriosis
patients and stages suggests that GC2 allele products may yield the
survival and implantation of ectopic endometrial tissues in the
peritoneal cavity or other places.
In conclusion, DBP may be crucial in the progression
of endometriosis. However, additional studies are necessary to
determine the association between DBP genotypes and the development
and progress of endometriosis.
Acknowledgements
This study was supported by a grant of the Korea
Health Technology R&D Project (A101367), Ministry of Health and
Welfare and the Next-Generation BioGreen 21 Program (PJ00819105),
Rural Development Administration, Republic of Korea.
References
|
1
|
Farquhar C: Endometriosis. BMJ.
334:249–253. 2007. View Article : Google Scholar
|
|
2
|
Meuleman C, Vandenabeele B, Fieuws S,
Spiessens C, Timmerman D and D’Hooghe T: High prevalence of
endometriosis in infertile women with normal ovulation and
normospermic partners. Fertil Steril. 92:68–74. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sasson IE and Taylor HS: Stem cells and
the pathogenesis of endometriosis. Ann N Y Acad Sci. 1127:106–115.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Eskenazi B and Warner ML: Epidemiology of
endometriosis. Obstet Gynecol Clin North Am. 24:235–258. 1997.
View Article : Google Scholar
|
|
5
|
Melega C, Balducci M, Bulletti C, Galassi
A, Jasonni VM and Flamigni C: Tissue factors influencing growth and
maintenance of endometriosis. Ann N Y Acad Sci. 622:256–265. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Noble LS, Simpson ER, Johns A and Bulun
SE: Aromatase expression in endometriosis. J Clin Endocrinol Metab.
81:174–179. 1996.PubMed/NCBI
|
|
7
|
Tseng JF, Ryan IP, Milam TD, Murai JT,
Schriock ED, Landers DV and Taylor RN: Interleukin-6 secretion in
vitro is up-regulated in ectopic and eutopic endometrial stromal
cells from women with endometriosis. J Clin Endocrinol Metab.
81:1118–1122. 1996.PubMed/NCBI
|
|
8
|
Vinatier D, Cosson M and Dufour P: Is
endometriosis an endometrial disease? Eur J Obstet Gynecol Reprod
Biol. 91:113–125. 2000. View Article : Google Scholar
|
|
9
|
Sharpe-Timms KL: Endometrial anomalies in
women with endometriosis. Ann N Y Acad Sci. 943:131–147. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kao LC, Germeyer A, Tulac S, Lobo S, Yang
JP, Taylor RN, Osteen K, Lessey BA and Giudice LC: Expression
profiling of endometrium from women with endometriosis reveals
candidate genes for disease-based implantation failure and
infertility. Endocrinology. 144:2870–2881. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hunter RH, Cicinelli E and Einer-Jensen N:
Peritoneal fluid as an unrecognised vector between female
reproductive tissues. Acta Obstet Gynecol Scand. 86:260–265. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Eyster KM, Boles AL, Brannian JD and
Hansen KA: DNA microarray analysis of gene expression markers of
endometriosis. Fertil Steril. 77:38–42. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Arimoto T, Katagiri T, Oda K, Tsunoda T,
Yasugi T, Osuga Y, Yoshikawa H, Nishii O, Yano T, Taketani Y and
Nakamura Y: Genome-wide cDNA microarray analysis of gene-expression
profiles involved in ovarian endometriosis. Int J Oncol.
22:551–560. 2003.PubMed/NCBI
|
|
14
|
Gupta S, Agarwal A, Sekhon L, Krajcir N,
Cocuzza M and Falcone T: Serum and peritoneal abnormalities in
endometriosis: potential use as diagnostic markers. Minerva
Ginecol. 58:527–551. 2006.PubMed/NCBI
|
|
15
|
Zhang H, Niu Y, Feng J, Guo H, Ye X and
Cui H: Use of proteomic analysis of endometriosis to identify
different protein expression in patients with endometriosis versus
normal controls. Fertil Steril. 86:274–282. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ferrero S, Gillott DJ, Remorgida V,
Anserini P, Leung KY, Ragni N and Grudzinskas JG: Proteomic
analysis of peritoneal fluid in women with endometriosis. J
Proteome Res. 6:3402–3411. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fowler PA, Tattum J, Bhattacharya S,
Klonisch T, Hombach- Klonisch S, Gazvani R, Lea RG, Miller I,
Simpson WG and Cash P: An investigation of the effects of
endometriosis on the proteome of human eutopic endometrium: a
heterogeneous tissue with a complex disease. Proteomics. 7:130–142.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ametzazurra A, Matorras R, García-Velasco
JA, Prieto B, Simón L, Martínez A and Nagore D: Endometrial fluid
is a specific and non-invasive biological sample for protein
biomarker identification in endometriosis. Hum Reprod. 24:954–965.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang T, Lee HG, Hwang JH, Oh JJ, Lim JN,
Kang HS, Joo JK and Lee KS: Myoglobin: a promising exogenous
reference marker using in proteomics analysis. Food Sci Biotechnol.
22:393–398. 2013. View Article : Google Scholar
|
|
20
|
Hwang JH, Oh JJ, Wang T, Jin YC, Lee JS,
Choi JR, Lee KS, Joo JK and Lee HG: Identification of biomarkers
for endometriosis in eutopic endometrial cells from patients with
endometriosis using a proteomics approach. Mol Med Rep. 8:183–194.
2013.PubMed/NCBI
|
|
21
|
Yang F, Brune JL, Naylor SL, Cupples RL,
Naberhaus KH and Bowman BH: Human group-specific component (Gc) is
a member of the albumin family. Proc Natl Acad Sci USA.
82:7994–7998. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cooke NE and David EV: Serum vitamin
D-binding protein is a third member of the albumin and alpha
fetoprotein gene family. J Clin Invest. 76:2420–2424. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lichenstein HS, Lyons DE, Wurfel MM,
Johnson DA, McGinley MD, Leidli JC, Trollinger DB, Mayer JP, Wright
SD and Zukowski MM: Afamin is a new member of the albumin,
alpha-fetoprotein, and vitamin D-binding protein gene family. J
Biol Chem. 269:18149–18154. 1994.PubMed/NCBI
|
|
24
|
Ray R: Molecular recognition in vitamin
D-binding protein. Proc Soc Exp Biol Med. 212:305–312. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Haddad JG: Plasma vitamin D-binding
protein (Gc-globulin): multiple tasks. J Steroid Biochem Mol Biol.
53:579–582. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Swamy N, Head JF, Weitz D and Ray R:
Biochemical and preliminary crystallographic characterization of
the vitamin D sterol- and actin-binding by human vitamin D-binding
protein. Arch Biochem Biophys. 402:14–23. 2002. View Article : Google Scholar
|
|
27
|
McLeod JF and Cooke NE: The vitamin
D-binding protein, alpha-fetoprotein, albumin multigene family:
detection of transcripts in multiple tissues. J Biol Chem.
264:21760–21769. 1989.PubMed/NCBI
|
|
28
|
Kew RR and Webster RO: Gc-globulin
(vitamin D-binding protein) enhances the neutrophil chemotactic
activity of C5a and C5a des Arg. J Clin Invest. 82:364–369. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamamoto N and Homma S: Vitamin D3 binding
protein (group-specific component) is a precursor for the
macrophage-activating signal factor from
lysophosphatidylcholine-treated lymphocytes. Proc Natl Acad Sci
USA. 88:8539–8543. 1991. View Article : Google Scholar
|
|
30
|
Chishimba L, Thickett DR, Stockley RA and
Wood AM: The vitamin D axis in the lung: a key role for vitamin
D-binding protein. Thorax. 65:456–462. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Borkowski J, Gmyrek GB, Madej JP, Nowacki
W, Goluda M, Gabryś M, Stefaniak T and Chełmońska-Soyta A: Serum
and peritoneal evaluation of vitamin D-binding protein in women
with endometriosis. Postepy Hig Med Dosw (Online). 62:103–109.
2008.PubMed/NCBI
|
|
32
|
Ferrero S, Gillott DJ, Anserini P,
Remorgida V, Price KM, Ragni N and Grudzinskas JG: Vitamin D
binding protein in endometriosis. J Soc Gynecol Investig.
12:272–277. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Faserl K, Golderer G, Kremser L, Lindner
H, Sarg B, Wildt L and Seeber B: Polymorphism in vitamin D-binding
protein as a genetic risk factor in the pathogenesis of
endometriosis. J Clin Endocrinol Metab. 96:E233–E241. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cho S, Choi YS, Yim SY, Yang HI, Jeon YE,
Lee KE, Kim H, Seo SK and Lee BS: Urinary vitamin D-binding protein
is elevated in patients with endometriosis. Hum Reprod. 27:515–522.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hunt JL and Licht P: Identification and
structural characterization of a novel member of the vitamin D
binding protein family. Comp Biochem Physiol B Biochem Mol Biol.
121:397–406. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Haddad JG, Harper KD, Guoth M, Pietra GG
and Sanger JW: Angiopathic consequences of saturating the plasma
scavenger system for actin. Proc Natl Acad Sci USA. 87:1381–1385.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Perez HD, Kelly E, Chenoweth D and Elfman
F: Identification of the C5a des Arg cochemotaxin. Homology with
vitamin D-binding protein (group-specific component globulin). J
Clin Invest. 82:360–363. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Piquette CA, Robinson-Hill R and Webster
RO: Human monocyte chemotaxis to complement-derived chemotaxins is
enhanced by Gc-globulin. J Leukoc Biol. 55:349–354. 1994.PubMed/NCBI
|
|
39
|
Yamamoto N, Homma S and Millman I:
Identification of the serum factor required for in vitro activation
of macrophages. Role of vitamin D3-binding protein (group specific
component, Gc) in lysophospholipid activation of mouse peritoneal
macrophages. J Immunol. 147:273–280. 1991.
|
|
40
|
Yamamoto N, Naraparaju VR and Asbell SO:
Deglycosylation of serum vitamin D3-binding protein leads to
immunosuppression in cancer patients. Cancer Res. 56:2827–2831.
1996.PubMed/NCBI
|
|
41
|
Wood AM, Bassford C, Webster D, Newby P,
Rajesh P, Stockley RA and Thickett DR: Vitamin D-binding protein
contributes to COPD by activation of alveolar macrophages. Thorax.
66:205–210. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ishii T, Keicho N, Teramoto S, Azuma A,
Kudoh S, Fukuchi Y, Ouchi Y and Matsuse T: Association of
Gc-globulin variation with susceptibility to COPD and diffuse
panbronchiolitis. Eur Respir J. 18:753–757. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ito I, Nagai S, Hoshino Y, Muro S, Hirai
T, Tsukino M and Mishima M: Risk and severity of COPD is associated
with the group-specific component of serum globulin 1F allele.
Chest. 125:63–70. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dimeloe S and Hawrylowicz C: A direct role
for vitamin D-binding protein in the pathogenesis of COPD? Thorax.
66:189–190. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chun RF: New perspectives on the vitamin D
binding protein. Cell Biochem Funct. 30:445–456. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Horne SL, Cockcroft DW and Dosman JA:
Possible protective effect against chronic obstructive airways
disease by the GC2 allele. Hum Hered. 40:173–176. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Schellenberg D, Paré PD, Weir TD, Spinelli
JJ, Walker BA and Sandford AJ: Vitamin D binding protein variants
and the risk of COPD. Am J Respir Crit Care Med. 157:957–961. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Borges CR, Jarvis JW, Oran PE and Nelson
RW: Population studies of vitamin D binding protein
microheterogeneity by mass spectrometry lead to characterization of
its genotype- dependent O-glycosylation patterns. J Proteome Res.
7:4143–4153. 2008. View Article : Google Scholar
|