Introduction
The (pro)renin receptor [(P)RR also called RER]
constitutes a novel component of the renin-angiotensin system (RAS)
and exerts pivotal functions in cardio-renal pathophysiology since
it is linked to angiotensin II-dependent and also angiotensin
II-independent effects (1).
Binding of renin to the (P)RR increases its catalytic efficiency 4-
to 5-fold, whereas binding of prorenin non-proteolytically demasks
the catalytic activity of prorenin (2).
Various research studies have demonstrated that the
competitive inhibition of the binding of (pro)renin to the (P)RR by
parenteral application of a decoy peptide, which is derived from
the prosegment of prorenin, prevents the development of diabetic
nephropathy and reduces cardiac fibrosis (3–6).
Importantly, (pro)renin receptor blockade by these decoys is also
effective in angiotensin AT1 receptor (AT1R)-knockout mice
(7) and can even reverse renal
damage caused by diabetes (8).
Nevertheless, a number of authors were not able to
observe positive effects of these peptides (9,10).
These controversies may be explained by the prorenin-renin ratio
since the decoys were effective in high prorenin/low renin models
(1,11), and/or by the length of treatment
based on the observation that renoprotective effects were able to
be observed after 12 weeks even in a high renin Goldblatt model
(12,13).
In addition, the effectiveness of the decoy peptides
regarding reduction in weight gain, serum triglycerides and
hyperinsulinemia has been shown in animals fed with a high fat/high
carbohydrate diet (14,15). Beneficial effects were also
observed concerning diabetic retinopathy (16–18). Recently, it was demonstrated that
the (P)RR was upregulated at the mRNA and protein levels in murine
hearts after myocardial infarction as well as in patients with
dilated cardiomyopathy (19)
further supporting the pathophysiological importance of this
receptor. Consistently, at the molecular level, activation of the
(P)RR by (pro)renin is associated with a detrimental
transcriptional signature, e.g. linked to cardiac hypertrophy,
cardiac and renal cell death (20). Moreover, the (P)RR is upregulated
on the mRNA and protein levels in the hearts and kidneys of
diabetic rats (21,22).
Our research group revealed a novel signal
transduction pathway involving the physical interaction between the
(P)RR and the transcription factor promyelocytic leukemia zinc
finger protein (PLZF). Upon stimulation of the (P)RR with renin or
prorenin, PLZF was found to translocate into the nucleus and
repress the RER promoter itself (23,24). Regarding the ligand level,
different renin glyocoforms, which are likely linked to the
differential plasma half-lives, have been previously described
(25).
In addition, renin-independent functions of the
(P)RR have been recently described (26). In this context, it is important to
note that the (P)RR protein consists of an evolutionarily conserved
part (comprising the C-terminal 69–100 amino acids) and a large
N-terminal (pro)renin binding domain (1,27).
The ancient part is identical to the vacuolar proton-translocating
ATPase (V-ATPase) membrane sector-associated protein M8–9 (28,29). Therefore, it has been suggested
that the gene encoding the (P)RR results from a gene fusion
(30). Nevertheless, these parts
can be separated at the protein level again based on the
identification of a soluble (P)RR isoform [s(P)RR] which nearly
represents the extracellular part (31). The s(P)RR is generated by the
action of furin and/or ADAM proteases (31,32).
Considering the different protein isoforms of the
(P)RR, it has been shown that the specific V-ATPase inhibitor
bafilomycin A1 inhibits (P)RR signalling (29). In addition, the nuclear
translocation of PLZF, in the context of the angiotensin AT2
receptor (AT2R), is inhibited by the small molecule genistein
(33). Currently, genistein and
bafilomycin are the only drug-like, commercially available small
molecules directly interfering with (P)RR signal transduction.
Therefore, the aim of the present study was to
analyse (pro)renin-independent, i.e., constitutive, activity of the
(P)RR, the transcriptional and isoform-specific regulation of this
receptor as well as the effects of genistein and bafilomycin on its
signal transduction. Since constitutive receptor activity does not
exclude ligand effects (34) and
may be unveiled by inactivating post-translational modifications of
the ligand, we also analysed the effects of prorenin
deglycosylation.
Materials and methods
Cell culture
B-16V (mouse melanoma) and KELLY (human
neuroblastoma) cells (both from DSMZ, Braunschweig, Germany) were
grown in RPMI-1640 medium (Life Technologies, Darmstadt, Germany).
HeLa-S3 (human cervical carcinoma), HEK293 (human embryonic kidney)
(both from DSMZ) and HEK293T cells [American Type Culture
Collection (ATCC), Manassas, VA, USA] were cultivated in DMEM high
glucose (Life Technologies). Flp-In-293 HEK cells (Invitrogen,
Karlsruhe, Germany) were grown in DMEM high glucose supplemented
with 2 mM glutamine (PAA, Pasching, Austria). All media contained
10% fetal bovine serum (Life Technologies), 100 U/ml penicillin and
100 μg/ml streptomycin (A2212; Biochrom, Berlin, Germany). If the
experimental conditions were modified the changes are described in
the Results section. All cell lines were cultivated without
addition of an angiotensin AT1 receptor blocker (ARB) and, if not
otherwise specified, without starving in a humidified incubator
with 5% CO2 at 37°C.
All experiments using the stably transfected HeLa
cell line were performed in 48-well plates (BD Falcon, Franklin
Lakes, NJ, USA). For all transient transfections of (P)RR promoter
constructs and (P)RR expression vectors, 24-well plates (BD Falcon;
from Corning Inc., Corning, NY, USA) (in the case of KELLY cells)
and 6-well plates (BD Falcon), respectively, were used. Prorenin
was obtained from Innovative Research (Novi, MI, USA). Bafilomycin
A1 (Enzo Life Science, Lörrach, Germany) and genistein (Carl Roth
GmbH, Karlsruhe, Germany) were dissolved in 1% DMSO final if not
otherwise stated.
Subcloning and transient transfection
experiments
The full-length human (P)RR coding sequence was
cloned into pEGFP-N1 and pEGFP-C3 vectors (Clontech, Mountain View,
CA, USA) as described previously (23). A construct containing 1,100 bp
(directly upstream of the translational start site) of the human
(P)RR was subcloned into the pGL4.14 luciferase vector (Promega,
Mannheim, Germany) using previously published primers (23). Transient transfection experiments
of these expression vectors were perfomed using Genejuice (Merck,
Darmstadt, Germany) or Turbofect (Fermentas, St. Leon-Rot, Germany)
transfection reagents according to manufacturers’ protocols with 25
ng DNA/cm2.
SiRNA experiments were performed with siRNA against
(P)RR [5′-gcuccguaaucgccuguuu-3′ (sense strand); 20 nM final] or
scrambled control siRNA [5′-uuuaccgucgccuugagcu-3′ (sense strand)]
(Eurogentec, Köln, Germany) and against PLZF
[5′-ccagcaagauguuugagau-3′ (sense strand); 50 nM] or scrambled
control siRNA [5′-ucucgcagugacuauacau-3′ (sense strand)]
(Eurogentec), respectively, using HiPerfect (Qiagen, Hilden,
Germany). General efficacy of siRNA-mediated knockdown was
controlled in KELLY cells by real-time polymerase chain reaction
(PCR) ((P)RR mRNA was decreased to 10–30% relative to the scrambled
control) and Western blotting (RER protein was decreased to ~40%
relative to the scrambled control) and in double-stably transfected
HeLa cells by Western blotting.
Generation of stable cell lines
To measure the activity of the (P)RR, a
double-stable, double-monoclonal HeLa cell line was generated using
the human (P)RR promoter/pGL4.14 (firefly) and pGL4.79
(Renilla) plasmids (Promega). The (P) RR promoter sequence
and the assay principle were based on Schefe et al (23) and on a patent application of our
group (EP 1 890 152 A1 or PCT WO 2008/019735 A9). The
Renilla luciferase activity served for standardisation.
After the first transfection (pGL4.14) cells were selected using
hygromycin B (250 μg/ml medium; PAA) and monoclonalised using
cloning cylinders (C7983; Sigma-Aldrich, Steinheim, Germany). After
the second transfection (pGL4.79) selection was performed by
addition of G-418 sulphate (500 μg/ml medium; PAA) followed by
monoclonalisation.
A prorenin-overexpressing and prorenin-secreting
HEK293 cell line was generated using the Flp-In system (Invitrogen)
and an expression vector (pcDNA5/FRT) encoding preprorenin fused to
a C-terminal His10-tag.
Fractionated protein extraction
Nuclear and cytosolic proteins were isolated as
described previously (35).
Nuclear fractions were controlled by Wsestern blotting using an
antibody against TFIID as described below.
Protein purification and prorenin
deglycosylation
His10-tagged prorenin was purified via
metal affinity chromatography as follows. Flp-In-293-HEK cells
stably transfected with an expression vector encoding C-terminally
His10-tagged preprorenin were cultured without starving
in 1 or 4.5 g/l glucose concentrations for two weeks followed by a
two-day serum-free period to exclude interference of serum proteins
before collection of the supernatant for affinity chromatography of
prorenin. A gravity column (Empty Disposable PD-10 Column,
17-0435-01; GE Healthcare, Munich, Germany) was loaded with 5 ml of
a cobalt matrix (Talon Metal Affinity Resin; Clontech,
Saint-Germain-en-Laye, France) and equilibrated with 20 ml washing
buffer (50 mM sodium dihydrogen phosphate, 300 mM sodium chloride,
pH 8.0). Two hundred milliliters of the cell culture supernatant
was loaded on the column. The flow-through was discarded. The
matrix was washed with washing buffer containing 10 mM imidazole
(Sigma-Aldrich). Afterwards, prorenin was eluted with the same
buffer but with 250 mM imidazole. Ten fractions each 2 ml were
collected and analysed by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and Coumassie staining. The pure,
prorenin-containing fractions were pooled and dialysed in 3 liters
of phosphate-buffered saline (PBS) overnight at 4°C. Finally, the
protein solution was concentrated using filter devices excluding
proteins <30 kDa (Amicon Ultra 0.5 ml; Millipore GmbH,
Schwalbach, Germany). The concentration was determined with UV
absortion spectrometry (ND-1000; PeqLab, Erlangen, Germany).
Purified prorenin and a commercially obtained
prorenin (from Innovative Reseach) were deglycosylated using
N-glycosidase F (11365185001; Roche, Mannheim, Germany), which does
not exhibit proteolytic activity, for 2 h and resolved via SDS-PAGE
followed by Coomassie staining.
Real-time PCR
Reverse transcription was performed using M-MLV
reverse transcriptase (RNase H minus; Promega) and 1 μg RNA. PCR
was performed applying Go-Taq qPCR Master Mix (Promega) and the
following primer pairs: 5′-ATTGGC CTATACCAGGAGAG-3′ (forward) and
5′-TTCCCCATAAC GCTTCCCAA-3′ (reverse) for (P)RR and 5′-CCGCAGCTAGG
AATAATGGAATA-3′ (forward) and 5′-TCTAGCGGCGCA ATACGAAT-3′ (reverse)
for 18S rRNA. A reaction without addition of reverse transcriptase
served as the negative control. The PCR reactions were run on a
Stratagene Mx3000P (Stratagene, La Jolla, CA, USA).
Western blotting
Immunoblotting was performed as previously described
(36) but using a cell lysis
buffer containing 1× PBS (pH 7.2, without calcium and magnesium;
Invitrogen), 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% sodium
dodecyl sulfate (SDS) (all from Sigma-Aldrich) and Complete
EDTA-free cocktail tablets (Roche). The following primary
antibodies were used: anti-PLZF (ab39354; Abcam, Cambridge, UK),
anti-GFP (sc-8334; Santa Cruz Biotechnology, Inc., Heidelberg,
Germany), anti-TFIID [TATA box-binding protein (TBP)] (sc-273;
Santa Cruz Biotechnology, Inc.), anti-(P)RR (ATP6AP2; HPA003156;
Sigma-Aldrich), anti-actin (sc-1615; Santa Cruz Biotechnology,
Inc.) and anti-GAPDH (MAB374, Chemicon/Merck Millipore, Billerica,
MA, USA). Detection of the horseradish peroxidase (HRP)-labelled
secondary antibody was performed with an enhanced chemiluminescence
(ECL) reagent containing a 1:1 mixture of solution A [100 mM Tris
base (pH 8.5), 2.5 mM luminol (Sigma-Aldrich), 0.4 mM
p-coumaric acid (Sigma-Aldrich)] and solution B [100 mM Tris
base (pH 8.5), 0.02% H2O2]. Proteins were
quantified using a Bradford assay (Roti-Nanoquant; Carl Roth). Cell
culture supernatants were concentrated using Centriprep®
10K columns followed by Amicon Ultra 3K columns (both obtained from
Merck Millipore, Darmstadt, Germany) before Western blotting.
Densitometric analysis was carried out using ImageJ 1.42q software
(National Institutes of Health, USA).
Reporter gene assays
Promoter reporter assays regarding stable and
transient transfections were performed using the Dual-Luciferase
Reporter® assay system (Promega).
For transient transfections, relative luciferase
activity (RLA), defined as the mean value of the firefly
luciferase/Renilla luciferase ratios of each construct
related to the insertless reporter plasmid pGL4.14, served as
read-out. Regarding the stable transfection, the
firefly/Renilla ratio served as read-out.
Cellular phenotypic assays
Cellular proliferation was measured using the BrdU
colorimetric cell proliferation ELISA (Roche). Mitochondrial
dehydrogenase activity was determined via the Cell Proliferation
Assay XTT (AppliChem, Darmstadt, Germany). Cellular ATP
concentrations were analysed using the CellTiter-Glo Luminescent
Cell Viability Assay (Promega). The Bradford assay was obtained
commercially (Roti-Nanoquant; Carl Roth).
To measure lactate dehydrogenase (LDH) activity in
the cell culture supernatant, 50 μl medium, 200 μl NADH buffer [0.2
mM β-nicotinamide adenine dinucleotide (reduced disodium salt
hydrate; Sigma-Aldrich), 0.1 M potassium hydrogen phosphate buffer
(pH 7.4)] and, to start the reaction, 25 μl pyruvate buffer [22.7
mM sodium pyruvate (Sigma-Aldrich), 0.1 M potassium hydrogen
phosphate buffer (pH 7.4)] were mixed. After 10 sec, NADH was
quantified photometrically at 340 nM (10 times every 10 sec) in a
96-well plate reader.
For the staining of acidic cell organelles (e.g.
lysosomes or peroxisomes) HeLa cells were seeded 24 h before
stimulation in black 96-well plates (Cellstar MicroClear; Greiner,
Frickenhausen, Germany). After 1 h prestimulation with genistein or
bafilomycin A1, the dye (LysoTracker Red DND-99; Life Technologies)
was added to the culture medium (75 nM final) for 1 h. After
replacement of the supernatant by serum- and phenol red-free
medium, measurement was carried out according to the manufacturer’s
instructions using a fluorescence plate reader (Mithras 940;
Berthold, Bad Wildbad, Germany).
Statistical analysis
For comparisons of two interventions including siRNA
effects under DMSO control (Fig.
2A) a two-tailed, unpaired t-test was applied. Regarding
multiple comparisons, a one-way analysis of variance (ANOVA) with
Bonferroni post-hoc adjustment was performed. Statistical
significance was assumed at p<0.05 for t-test and ANOVA.
Concerning plotting of the dose-response curves, a sigmoidal
regression analysis was used.
Results
Effects of genistein, bafilomycin and
siRNA interventions on (P)RR promoter activity and (P)RR
expression
We previously demonstrated that PLZF is a crucial
adapter protein of the (P)RR and its own promoter (23). Other studies have shown that the
nuclear translocation of PLZF is inhibited by genistein (33) and that bafilomycin reduces (P)RR
signal transduction (29).
Therefore, we aimed to ascertain whether genistein, bafilomycin or
siRNA against pathway components affects (P)RR promoter activity.
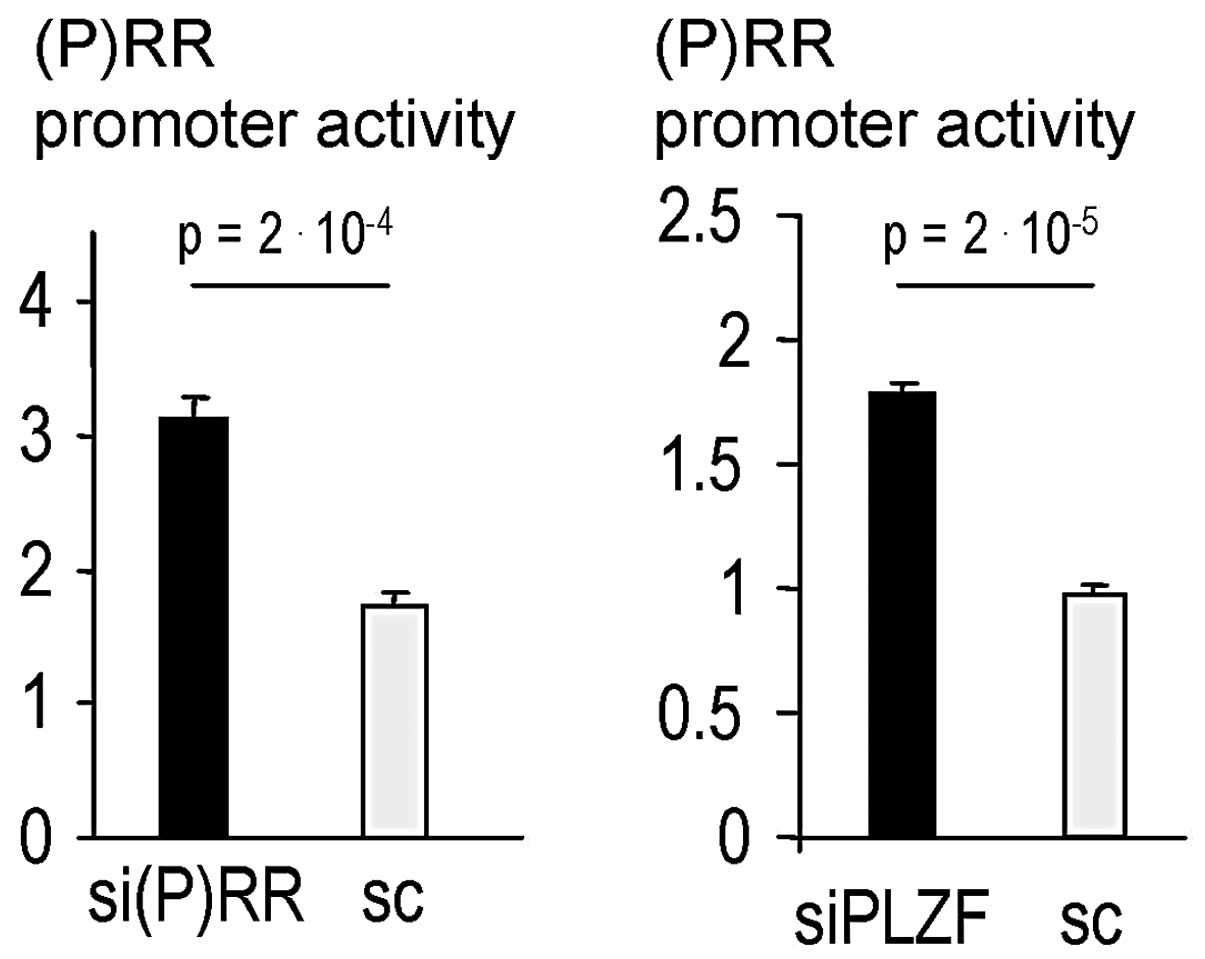
As expected, siRNA knockdown of (P)RR and PLZF both significantly
derepressed (P)RR promoter activity (Fig. 1). In addition, genistein increased
(P)RR promoter activity, whereas bafilomycin had a minor effect
(Fig. 2A and B). However, ANOVA
using the pooled data shown in Fig.
2A and B indicated that both bafilomycin concentrations
significantly increased promoter activity (p<0.01 for 0.1 μM and
p<0.001 for 1 μM). Moreover, siRNA against (P)RR and also PLZF
caused a derepression of (P)RR promoter activity in the context of
coincubations (Fig. 2A)
consistent with a repressive role of the (P)RR-PLZF pathway on the
(P)RR promoter (23).
Importantly, the effect of siRNA against PLZF was abolished by
genistein (Fig. 2A) and
bafilomycin (Fig. 2B), whereas
both substances did not inhibit the effect of siRNA against (P)RR
(Fig. 2A and B).
In addition, the EC50 value of genistein
was determined showing a potency of 2–4 μM regarding promoter
activation (Fig. 2C). In
addition, a non-sigmoidal concentration-response relationship was
also observed concerning the effects of bafilomycin on (P)RR
promoter activity (data not shown).
Finally, we analysed whether the effects of
genistein and bafilomycin can also be observed at the endogenous
transcript level. Both interventions significantly increased (P)RR
mRNA after 12 h (Fig. 2D) when
effects on cell number are unlikely.
Additionally, the effects of genistein and
bafilomycin on the protein level were analysed with respect to
full-length and soluble (P)RR. After 12 and 24 h of incubation and
upon examination of the total cellular lysates, treatment with
bafilomycin but not genistein significantly increased soluble (P)RR
(Fig. 2E). Consistently, a
significantly increased s(P)RR level was observed in the cellular
supernatant following incubation with bafilomycin (Fig. 2F).
In this context, it is important to note that the
antibody used in Fig. 2E and F
only detects the N-terminal part of the (P)RR, i.e., full-length
and s(P)RR, but not the V-ATPase-associated isoform.
Effects of glucose on (P)RR promoter
activity and (P)RR isoform expression
Based on the pathophysiological role of the (P)RR in
diabetic nephropathy, we examined its promoter regulation using
different glucose conditions. High glucose increased basal (P)RR
promoter activity ~3-fold (Fig.
3A). Strikingly, prorenin decreased (P)RR promoter activity
only under high glucose conditions whereas an inverse response was
observed in the cells cultured in a physiological glucose
concentration (Fig. 3A).
 | Figure 3Effects of high glucose conditions on
(pro)renin receptor [(P)RR] promoter activity and (P)RR isoform
expresssion. (A) HeLa cells double-stably transfected with (P)RR
promoter luciferase constructs were stimulated, after 24 h of
starving, with prorenin for 24 h under different glucose
concentrations (n=3 for each intervention). (B) HeLa and HEK293T
wild-type cells, cultured under the indicated conditions, were
transiently transfected with an expression vector encoding
full-length RER C- or N-terminally fused with an EGFP-tag of 26 kDa
(pEGFP-N1 or pEGFP-C3, respectively). (P)RR isoforms generated
endogenously by post-translational processing were detected by
Western blotting of total cellular proteins using an anti-GFP
antibody. Anti-actin Western blotting served as loading control. M,
molecular weight marker; D, 1% DMSO; G, 10 μM genistein; B, 0.1 μM
bafilomycin; f.l., full-length (P)RR-GFP; V, V-ATPase domain of
(P)RR fused to GFP. |
Since the (P)RR is known to be expressed in
different protein isoforms, we ascertained whether glucose and
bafilomycin also affect these protein identities of the (P)RR. For
this purpose, two expression vectors encoding full-length (P)RR N-
or C-terminally fused to GFP, respectively, were transiently
transfected into wild-type HeLa and wild-type HEK293T cells
(Fig. 3B). Regarding specificity,
the V-ATPase isoform can only be detected with the anti-GFP
antibody after transfection of a C-terminally tagged (P)RR
construct since the V-ATPase domain [~9 kDa (37)] within the full-length (P)RR
[~38–39 kDa (23)] is located at
the C-terminus. High glucose strongly increased full-length (P)RR
in HeLa but not in HEK cells. Bafilomycin, in contrast to
genistein, caused a relative shift from the full-length isoform to
the V-ATPase isoform in HeLa cells due to a decreased abundance of
the full-length form (Fig.
3B).
Effects of glucose on prorenin
glycosylation
To ascertain whether high glucose directly affects
the glycosylation of the ligand prorenin, we cultured HEK cells
stably overexpressing prorenin in high or (relatively) low glucose
medium (Fig. 4). The results
revealed that HEK cells exhibit the ability to glycosylate
prorenin. Furthermore, the two previously described N-glyosylation
sites of prorenin (38) were
confirmed as indicated by the double band in the presence of the
glycosidase. Importantly, high glucose did not alter the
glycosylation pattern of prorenin (Fig. 4).
Effects of prorenin glycosylation on
(P)RR promoter activity
We next aimed to ascertain whether deglycosylation
impacts the ligand activity of prorenin in transiently transfected
promoter assays with a sufficient transfection efficiency (as
indicated by a firefly signal of the insertless control vector over
1,000 counts) and with a full deglycosylation of both asparagines
(as confirmed by SDS-PAGE). As expected based on our previous
results (23,24), native prorenin significantly
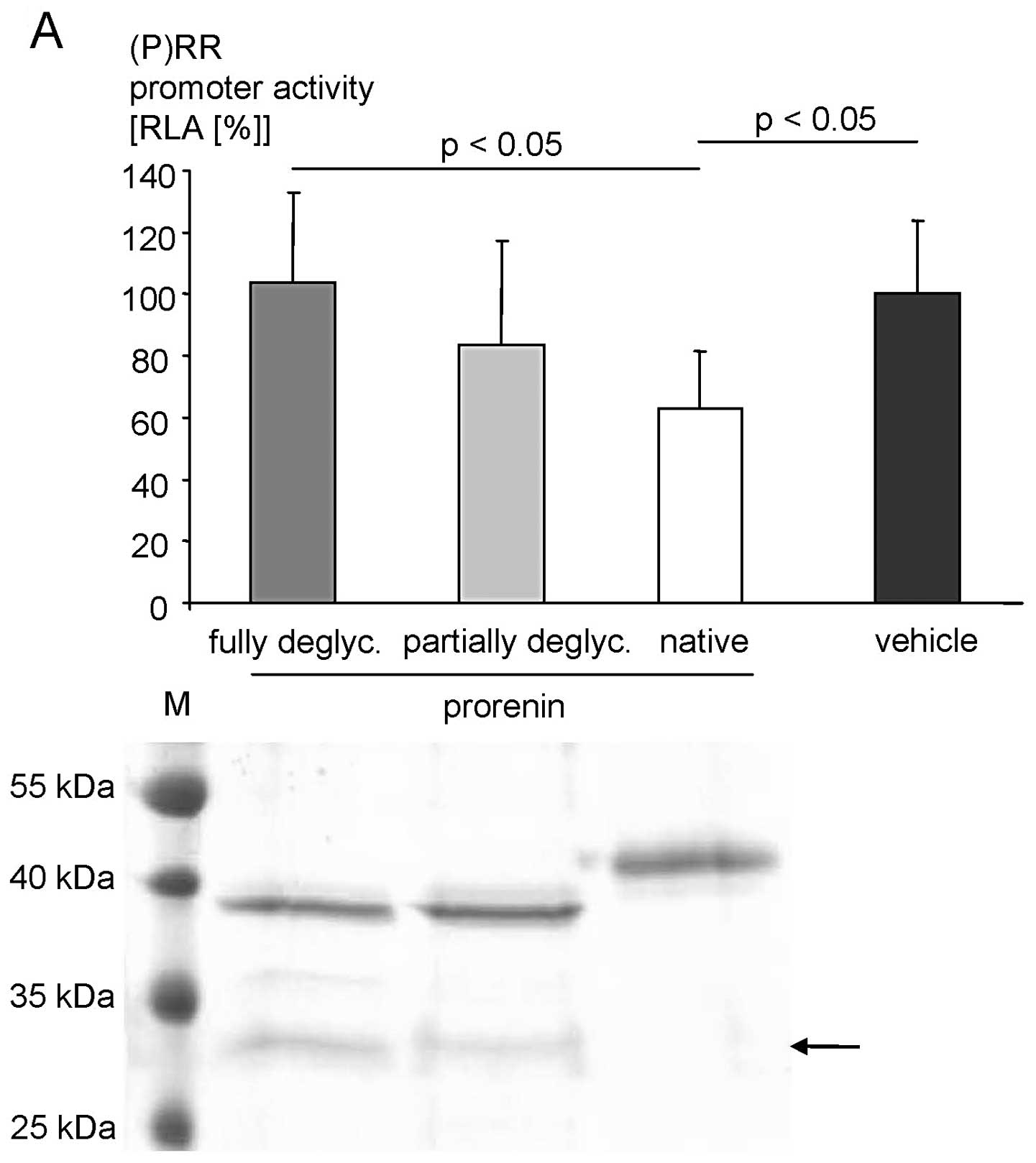
repressed (P)RR promoter activity (Fig. 5A). Complete deglycosylation, as
confirmed by SDS-PAGE, abolished this repressive effect (Fig. 5A).
Similar results were also obtained for neuronal
KELLY cells. Native prorenin strongly repressed (P)RR promoter
activity. Deglycosylation of both asparagines of prorenin abolished
the ability to repress the (P)RR promoter, whereas a mixture of
fully deglycosylated and partially deglycosylated (i.e., one of the
two glycosylation sites is completely deglycosylated) prorenin
exhibited intermediate repressive effects (Fig. 5B).
Effects of glucose, genistein and
bafilomycin on PLZF translocation
Finally, we aimed to ascertain whether genistein and
bafilomycin affect the nuclear translocation of PLZF at different
glucose concentrations. Genistein (100 μM) (Fig. 6A) and bafilomycin (1 μM) (Fig. 6B) reduced the nuclear content of
PLZF under physiological and high glucose conditions after 18 h in
the HEK293T cells. This effect of genistein was not observed after
12 h in the HEK293T and KELLY cells (data not shown). Glucose
concentration per se (i.e., under DMSO) appeared not to
alter the PLZF content (Fig.
6).
Cellular effects of the (P)RR
sub-pathways
Regarding cellular effects of the
(pro)renin-(P)RR-PLZF cascade, HEK cells stably overexpressing
prorenin were analysed regarding proliferation using a BrdU assay
(Fig. 7A). Compared to the
non-overexpressing control cells, prorenin caused an increase in
proliferation. This pro-proliferative effect was attenuated by
siRNA against (P)RR, siRNA against PLZF and by pharmacological
interventions using genistein or bafilomycin.
In addition, we examined the (pro)renin-independent
cellular effects of the (P)RR. Repression of (P)RR expression by
siRNA in wild-type neuronal cells in a system without (pro)renin
supplementation significantly decreased cell number (Fig. 7B), whereas PLZF knockdown by siRNA
had no effect (data not shown). Finally, the dose-dependent
proliferative effects of genistein and bafilomycin were analysed in
wild-type KELLY cells (Fig. 7C).
Bafilomycin strongly decreased proliferation in contrast to
genistein in a cellular system without incubation of recombinant or
purified (pro)renin.
To analyse whether the altered proliferation is
translated into different cell numbers, we performed XTT and ATP
assay in addition to total protein measurements under the cell
culture conditions as in Fig. 7B and
C since these assays are indicative of total cell count
(Fig. 7D). The shapes of the
dose-response curves including the rightward shift between
genistein and bafilomycin were observed in all of the assays and in
the different cell types indicating that bafilomycin reduced the
cell number through inhibition of proliferation. Furthermore, a
minor contribution of direct cytotoxicity to the cell
number-reducing effects of bafilomycin A1 was demonstrated using a
LDH assay (Fig. 7D).
Finally, we examined whether small molecule
interventions using genistein or bafilomycin and gene silencing
approaches using siRNA exhibit similar effects. Affecting the (P)RR
by either bafilomycin or siRNA, in the absence of (pro)renin, both
strongly reduced cell number in a concentration-response
relationship (Fig. 8). In
contrast, affecting PLZF by either exposure to genistein or siRNA,
in the absence of prorenin, did not alter the cell number with the
exception of 100 μM genistein (Fig.
8).
Concerning the intracellular phenotypic impacts of
genistein and bafilomycin, we determined intravesicular pH
regulation using Lysotracker fluorescence dye in HeLa cells. To
exclude effects on cell number, incubation of these substances was
restricted to 2 h. Fig. 9
indicates that bafilomycin increased lysosomal/peroxisomal pH with
an EC50 of ~2 nM whereas genistein had no effect.
Discussion
There are three major findings of this study
regarding the (P)RR signal transduction cascade. First, ligand
glycosylation is a crucial determinant of intrinsic activity.
Second, glucose concentration affects (P)RR signalling at different
levels. Third, the (P)RR exhibits constitutive, PLZF-independet
pro-proliferative effects.
In the present study, we demonstrated that the
steady-state glucose concentration does not affect the
glycosylation pattern of prorenin (Fig. 4), in contrast to HbA1c glycation
(i.e., non-enzymatic glycosylation) by glucose (39). The glycosylation pattern itself is
crucial for the ligand activity of prorenin since deglycosylation
abolishes the effect of prorenin on the (P)RR signal transduction
(Fig. 5). Furthermore, a mixture
of fully deglycosylated and partially deglycosylated (i.e., one of
the two glycosylation sites was completely deglycosylated) prorenin
is sufficient to mediate half-maximal intrinsic activity (Fig. 5B). To the best of our knowledge,
this is the first report demontrating that the intrinsic activity
of prorenin depends on its glycosylation.
In contrast to the glucose-independent ligand
glycosylation, high glucose conditions increased the basal (i.e.,
without prorenin stimulation) (P)RR activity as measured by
promoter assay, and reversed the ability of prorenin regarding
(P)RR activation (Fig. 3A). This
increased basal (P)RR receptor activity under high glucose was
likely caused by an increased receptor expression based on the
observation that glucose induced (P)RR at the protein level in HeLa
cells (Fig. 3B). The increased
full-length (P)RR protein expression appeared not to alter the
(P)RR isoform ratio in HeLa cells (Fig. 3B).
The strong upregulation of full-length (P)RR protein
expression in HeLa cells cultivated in high glucose conditions
observed in this study is consistent with previous studies which
demonstrated an upregulation of (P)RR mRNA and protein in hearts
(21) as well as kidneys
(22) of diabetic rats, of (P)RR
protein in kidneys of patients with diabetic nephropathy (40), and of (P)RR mRNA and protein in
rat mesangial cells by glucose (41). Mechanistically, NF-κB, AP-1 and
Sp1/Sp3 appear to be involved in the (P)RR promoter regulation by
high glucose (42). The authors
showed that an exchange of c-jun, c-fos, NF-κB p65 and NF-κB p50 on
cis-elements of the (P)RR promoter mediated the glucose
responsiveness of this gene, indicating a complex promoter
regulation in which more than one transcription factor was
involved. Furthermore, dissocation of V-ATPase subunits was found
to be promoted by low glucose levels (43) which in turn may affect the
regulation of the (P)RR gene, since it encodes an essential
accessory protein of V-ATPases (44).
Focusing on the regulatory levels, bafilomycin and
genistein increased (P)RR promoter activity and (P)RR mRNA
(Fig. 2A–D). Bafilomycin but not
genistein decreased exogenous (i.e., under control of a CMV
promoter) full-length (P)RR protein in HeLa cells (Fig. 3B). Concerning endogenous (P)RR,
soluble (P)RR was strongly increased by bafilomycin in the total
lysates and in the supernatant (Fig.
2E and F). This clearly increased s(P)RR expression may be
reflected by an increased translation of full-length (P)RR followed
by direct processing into the soluble isoform and a rapid
degradation of the V-ATPase-associated isoform. This would also
explain why the full-length (P)RR protein expression was almost
unaltered by bafilomycin (Fig.
2E) and is consistent with a recent conclusion by Fukushima
et al, that the elevation of plasma s(P)RR level may
indicate an upregulation of the full-length form (45).
Here, we were able to confirm the previous data of
our group (23,24) that revealed that prorenin mediates
pro-proliferative effects via the (P)RR-PLZF axis (Fig. 7A) which is also consistent with a
microarray analysis linking (pro)renin stimulation with a gene
signature associated with cardiac hypertrophy (20). Furthermore, research indicates
that (pro)renin increases DNA synthesis (46) and proliferation (47) of vascular smooth muscle cells, and
that prorenin can increase protein and DNA synthesis in
cardiomyocytes underlying myocyte hypertrophy and proliferation
(48). In addition, it was found
that prorenin increased the proliferation of endothelial cells and
that melanoma xenografts stably transfected with prorenin had an
increased tumour growth in vivo compared to mock controls
likely involving the (P)RR since an angiotensin AT1 receptor
blocker was ineffective in vitro (49).
Our data obtained in neuronal, melanoma, hepatoma
and epithelial cells in the absence of stimulation with the ligand
(pro)renin indicate that the (P)RR exerts additional constitutive,
cell type-independent pro-proliferative/pro-survival effects
(Fig. 7C and D). These effects
are independent of PLZF as indicated by our experiments using siRNA
against PLZF or genistein. This is in contrast to the essential
role of PLZF in (pro)renin-induced pro-proliferation as discussed
above (Fig. 10). This
constitutive function of the (P)RR is consistent with the phenotype
of the cardiomyocyte-specific (P)RR knockout which is characterised
by cardiac cell death (50) and
also with the phenotype of the podocyte-specific (P)RR knockout
which is characterised by non-apoptotic podocyte cell death
(51) since both phenotypes can
be explained by a V-ATPase dysfunction. Moreover, repression of
(P)RR expression by siRNA reduced the viability of cultured
cardiomyoblasts in an experimental setting without (pro)renin
incubation (21). The role of the
constitutive (P)RR function in cellular survival is also supported
by the central nervous system necrosis observed in zebrafish with
(P)RR gene mutagenesis (52) but
also in zebrafish with genetic alterations in different V-ATPase
subunits (30). Furthermore, the
ligand renin is only expressed in mammalian and nonmammalian
vertebrates (53) but not in
invertebrates such as C. elegans, which is not viable when
(P)RR is lacking (27). The
importance of the constitutive activity is further supported by the
fact that at least plasma (pro)renin levels and, therefore, even
more (pro)renin concentrations in cell culture medium with a serum
content of 10%, are too low to be of biological relevance with
respect to (P)RR activation (54).
The extent to which (pro)renin-independent effects
are mediated by the Wnt pathway, considering the essential role of
the (P)RR in its signalling (26), and/or by the V-ATPase isoform
remains to be elucidated. Consistent with a role of Wnt in the
basal (i.e., ligand-independent) effects of the (P)RR observed
here, it was shown that (P)RR function within the Wnt pathway is
renin-independent (26).
The overall (i.e., ligand-dependent and/or
ligand-independent) role of the (P)RR in cellular survival is also
illustrated by the observation that embryonic stem (ES) cells
deficient in the (P)RR gene are incompatible with the development
of chimeric mice when injected into blastocysts (51,55) and by the likely involvement of the
(P)RR in the growth of glioma cells (56).
The present study is the first to simultaneously
address the effects of the small molecules genistein and
baflomycin. Genistein is a phytoestrogen known to interact with
estrogen receptors α (ERα) and ERβ leading to activation of ER
responsive genes (57,58). Furthermore, genistein can inhibit
tyrosine kinases (59) and
aspects of Wnt signalling (60)
in addition to its effects on histone modifications and DNA
methylation (i.e., epigenetic modulations) (61) as well as on NF-κB (62) and Smad (63) signal transductions. In the context
of our data, it is important to note that genistein inhibits the
nuclear translocation of PLZF (33). We also observed that genistein
inhibited nuclear translocation of PLZF in HEK293T cells indicating
the contribution of this genistein-mediated mechanism in our
experiments. Since HeLa-S3 cells do not express ERα and ERβ
receptors (data not shown) the effects of genistein on this cell
type are non-ER-mediated. Similar to our data that genistein does
not significantly inhibit cell growth, the MTT assay-determined
IC50 value of genistein in a panel of cancer cell lines
was in the two-digit micromolar range (64). Consistently, a recent clinical
phase II trial indicated that genistein did not increase the
survival of pancreatic cancer patients (65).
Bafilomycin A1 is a specific V-ATPase inhibitor
which can inhibit the ligand- (i.e., prorenin- and renin-)
dependent (29) and
Wnt-associated (26) signal
transduction of the (P)RR.
In accordance with our data, bafilomycin A1 was
found to decrease the growth of different tumour cell lines in
vitro, with an IC50 of 5 nM regarding cellular
viability of pancreatic cancer cells (68), as well as xenograft growth in
vivo (66–68). Nevertheless, its toxicity excludes
its use in clinical trials (69,70).
Concerning cardiovascular indications, it was
recently demonstrated that genistein reduced proteinuria,
albuminuria and glomerular deposits in streptozotocin-induced
diabetic mice (71), similar
effects as observed with the anti-prorenin decoy peptides discussed
above (3). In addition to these
beneficial effects, genistein was found to protect pancreatic β
cells from high glucose-induced apoptosis (72). In contrast, bafilomycin A1 reduced
pancreatic islet size and impaired glucose tolerance in animal
models (73,74) hypothetically linking this
bacterial toxin to the development of type I diabetes (75).
Based on our data, we conclude that genistein is a
small molecular mimetic of siRNA against PLZF, whereas bafilomycin
exerted similar effects as siRNA against (P)RR for the following
reasons. First, genistein, bafilomycin as well as siRNAs against
PLZF or (P)RR all increased (P)RR promoter activity (Figs. 1 and 2B). Secondly, neither PLZF silencing by
siRNA nor genistein exerted significant effects on cell number
(Fig. 8). In all our cellular
phenotypic assays regarding total protein concentration, cell
number and proliferation, genistein had no effects except at the
highest (100 μM) concentration used (Fig. 7C and D). Since this concentration
is far beyond the EC50 (Fig. 2B), this indicates putative
unspecific effects at 100 μM, which is consistent with the
threshold of 5 μM genistein regarding a non-physiological in
vitro concentration (76). In
contrast, both siRNA against (P)RR and also bafilomycin similarly
reduced cell number (Fig. 8).
Third, other groups have shown that wild-type podocytes treated
with bafilomycin A1 are characterised by similar morphologic and pH
changes compared to podocytes with (P)RR deletion (51).
We further conclude that (P)RR and PLZF functions
are not identical, despite a similar impact on promoter feedback
(Fig. 1) and a similar effect on
prorenin-induced proliferation (24), for the following reasons. First,
as discussed above, (P)RR affected cell number/proliferation in
contrast to PLZF. Second, genistein and bafilomycin had distinct
effects on (P)RR isoform expression (Fig. 3B). Third, bafilomycin, as
expected, increased lysosomal/peroxisomal pH as indicated by a
reduction in Lysotracker fluorescence emission in contrast to
genistein (Fig. 9).
To conclude, our data indicate that the (P)RR does
not only exert angiotensin II-independent (2, 23)
but also (pro)renin-independent (i.e., constitutive) effects. By
employing dose-response analyses and various cellular assays, the
present study is the first detailed description of the
constitutive, pro-proliferative/pro-survial actions of this
receptor. Furthermore, the novel finding that glycosylation of
prorenin is crucial regarding its ability to initiate a signal
transduction at the (P)RR was demonstrated.
Acknowledgements
The study was supported by grants from the BMBF
(GO-Bio programme, no. 0315092 and VIP-programme, no. 0275
03V0367), the Investitionsbank Berlin (IBB) [ProFIT programme, no.
10138510; Europäischer Fonds für regionale Entwicklung (EFRE) of
the European Union (EU)] and by the Stiftung Charité.
References
|
1
|
Funke-Kaiser H, Zollmann FS, Schefe JH and
Unger T: Signal transduction of the (pro)renin receptor as a novel
therapeutic target for preventing end-organ damage. Hypertens Res.
33:98–104. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nguyen G, Delarue F, Burckle C, Bouzhir L,
Giller T and Sraer JD: Pivotal role of the renin/prorenin receptor
in angiotensin II production and cellular responses to renin. J
Clin Invest. 109:1417–1427. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ichihara A, Hayashi M, Kaneshiro Y, et al:
Inhibition of diabetic nephropathy by a decoy peptide corresponding
to the ‘handle’ region for nonproteolytic activation of prorenin. J
Clin Invest. 114:1128–1135. 2004.PubMed/NCBI
|
|
4
|
Ichihara A, Kaneshiro Y, Takemitsu T, et
al: Nonproteolytic activation of prorenin contributes to
development of cardiac fibrosis in genetic hypertension.
Hypertension. 47:894–900. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kaneshiro Y, Ichihara A, Sakoda M, et al:
Slowly progressive, angiotensin II-independent glomerulosclerosis
in human (pro)renin receptor-transgenic rats. J Am Soc Nephrol.
18:1789–1795. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Susic D, Zhou X, Frohlich ED, Lippton H
and Knight M: Cardiovascular effects of prorenin blockade in
genetically spontaneously hypertensive rats on normal and high-salt
diet. Am J Physiol Heart Circ Physiol. 295:H1117–H1121. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ichihara A, Suzuki F, Nakagawa T, et al:
Prorenin receptor blockade inhibits development of
glomerulosclerosis in diabetic angiotensin II type 1a
receptor-deficient mice. J Am Soc Nephrol. 17:1950–1961. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takahashi H, Ichihara A, Kaneshiro Y, et
al: Regression of nephropathy developed in diabetes by (pro)renin
receptor blockade. J Am Soc Nephrol. 18:2054–2061. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Muller DN, Klanke B, Feldt S, et al:
(Pro)renin receptor peptide inhibitor ‘handle-region’ peptide does
not affect hypertensive nephrosclerosis in Goldblatt rats.
Hypertension. 51:676–681. 2008.
|
|
10
|
Feldt S, Maschke U, Dechend R, Luft FC and
Muller DN: The putative (pro)renin receptor blocker HRP fails to
prevent (pro)renin signaling. J Am Soc Nephrol. 19:743–748. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nguyen G and Muller DN: The biology of the
(pro)renin receptor. J Am Soc Nephrol. 21:18–23. 2010. View Article : Google Scholar
|
|
12
|
Ryuzaki M, Ichihara A, Ohshima Y, et al:
Involvement of activated prorenin in the pathogenesis of slowly
progressive nephropathy in the non-clipped kidney of two kidney,
one-clip hypertension. Hypertens Res. 34:301–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kiyomoto H and Moriwaki K: Chronic
blockade of the (pro)renin receptor ameliorates the kidney damage
in the non-clipped kidney of Goldblatt hypertension. Hypertens Res.
34:289–291. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nagai Y, Ichihara A, Nakano D, et al:
Possible contribution of the non-proteolytic activation of prorenin
to the development of insulin resistance in fructose-fed rats. Exp
Physiol. 94:1016–1023. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lavoi JL: Methods of treating or
preventing obesity and obesity-related hypertension.
Patentanmeldung WO 2009/143619 A1. 2009
|
|
16
|
Satofuka S, Ichihara A, Nagai N, et al:
Role of nonproteolytically activated prorenin in pathologic, but
not physiologic, retinal neovascularization. Invest Ophthalmol Vis
Sci. 48:422–429. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Satofuka S, Ichihara A, Nagai N, et al:
(Pro)renin receptor-mediated signal transduction and tissue
renin-angiotensin system contribute to diabetes-induced retinal
inflammation. Diabetes. 58:1625–1633. 2009. View Article : Google Scholar
|
|
18
|
Wilkinson-Berka JL, Heine R, Tan G,
Tikellis C, Cooper ME, Nguyen G and Miller AG: The role of the
(pro)renin receptor in developing ischaemic and diabetic retina. J
Renin Angiotensin Aldosterone Syst. 9(Suppl 1): S82008.
|
|
19
|
Mahmud H, Sillje HH, Cannon MV, van Gilst
WH and de Boer RA: Regulation of the (pro)renin-renin receptor in
cardiac remodelling. J Cell Mol Med. 16:722–729. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Melnyk RA, Tam J, Boie Y, Kennedy BP and
Percival MD: Renin and prorenin activate pathways implicated in
organ damage in human mesangial cells independent of angiotensin II
production. Am J Nephrol. 30:232–243. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Connelly KA, Advani A, Kim S, et al: The
cardiac (pro)renin receptor is primarily expressed in myocyte
transverse tubules and is increased in experimental diabetic
cardiomyopathy. J Hypertens. 29:1175–1184. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Siragy HM and Huang J: Renal (pro)renin
receptor upregulation in diabetic rats through enhanced angiotensin
AT1 receptor and NADPH oxidase activity. Exp Physiol. 93:709–714.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schefe JH, Menk M, Reinemund J, et al: A
novel signal transduction cascade involving direct physical
interaction of the renin/prorenin receptor with the transcription
factor promyelocytic zinc finger protein. Circ Res. 99:1355–1366.
2006. View Article : Google Scholar
|
|
24
|
Schefe JH, Neumann C, Goebel M, et al:
Prorenin engages the (pro)renin receptor like renin and both ligand
activities are unopposed by aliskiren. J Hypertens. 26:1787–1794.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Katz SA, Opsahl JA, Abraham PA and Gardner
MJ: The relationship between renin isoelectric forms and renin
glycoforms. Am J Physiol. 267:R244–R252. 1994.PubMed/NCBI
|
|
26
|
Cruciat CM, Ohkawara B, Acebron SP, et al:
Requirement of prorenin receptor and vacuolar
H+-ATPase-mediated acidification for Wnt signaling.
Science. 327:459–463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bader M: The second life of the (pro)renin
receptor. J Renin Angiotensin Aldosterone Syst. 8:205–208. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ludwig J, Kerscher S, Brandt U, et al:
Identification and characterization of a novel 9.2-kDa membrane
sector-associated protein of vacuolar proton-ATPase from chromaffin
granules. J Biol Chem. 273:10939–10947. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Advani A, Kelly DJ, Cox AJ, et al: The
(pro)renin receptor: site-specific and functional linkage to the
vacuolar H+-ATPase in the kidney. Hypertension.
54:261–269. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sihn G, Rousselle A, Vilianovitch L,
Burckle C and Bader M: Physiology of the (pro)renin receptor: Wnt
of change? Kidney Int. 78:246–256. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cousin C, Bracquart D, Contrepas A, Corvol
P, Muller L and Nguyen G: Soluble form of the (pro)renin receptor
generated by intracellular cleavage by furin is secreted in plasma.
Hypertension. 53:1077–1082. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yoshikawa A, Aizaki Y, Kusano K, et al:
The (pro)renin receptor is cleaved by ADAM19 in the Golgi leading
to its secretion into extracellular space. Hypertens Res.
34:599–605. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Senbonmatsu T, Saito T, Landon EJ, et al:
A novel angiotensin II type 2 receptor signaling pathway: possible
role in cardiac hypertrophy. EMBO J. 22:6471–6482. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Seifert R and Wenzel-Seifert K:
Constitutive activity of G-protein-coupled receptors: cause of
disease and common property of wild-type receptors. Naunyn
Schmiedebergs Arch Pharmacol. 366:381–416. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Funke-Kaiser H, Reichenberger F, Köpke K,
et al: Differential binding of transcription factor E2F-2 to the
endothelin-converting enzyme-1b promoter affects blood pressure
regulation. Hum Mol Genet. 12:423–433. 2003. View Article : Google Scholar
|
|
36
|
Seidel K, Kirsch S, Lucht K, et al: The
promyelocytic leukemia zinc finger (PLZF) protein exerts
neuroprotective effects in neuronal cells and is dysregulated in
experimental stroke. Brain Pathol. 21:31–43. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jansen EJ and Martens GJ: Novel insights
into V-ATPase functioning: distinct roles for its accessory
subunits ATP6AP1/Ac45 and ATP6AP2/(pro)renin receptor. Curr Protein
Pept Sci. 13:124–133. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Holzman TF, Chung CC, Edalji R, et al:
Recombinant human prorenin from CHO cells: expression and
purification. J Protein Chem. 9:663–672. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wautier JL and Schmidt AM: Protein
glycation: a firm link to endothelial cell dysfunction. Circ Res.
95:233–238. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Takahashi K, Yamamoto H, Hirose T, et al:
Expression of (pro)renin receptor in human kidneys with end-stage
kidney disease due to diabetic nephropathy. Peptides. 31:1405–1408.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang J and Siragy HM: Glucose promotes
the production of interleukine-1beta and cyclooxygenase-2 in
mesangial cells via enhanced (Pro)renin receptor expression.
Endocrinology. 150:5557–5565. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Huang J and Siragy HM: Regulation of
(pro)renin receptor expression by glucose-induced mitogen-activated
protein kinase, nuclear factor-kappaB, and activator protein-1
signaling pathways. Endocrinology. 151:3317–3325. 2010. View Article : Google Scholar
|
|
43
|
Sennoune SR and Martinez-Zaguilan R:
Vacuolar H+-ATPase signaling pathway in cancer. Curr
Protein Pept Sci. 13:152–163. 2012.
|
|
44
|
Krop M, Lu X, Danser AH and Meima ME: The
(pro)renin receptor. A decade of research: what have we learned?
Pflugers Arch. 465:87–97. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fukushima A, Kinugawa S, Homma T, et al:
Increased plasma soluble (pro)renin receptor levels are correlated
with renal dysfunction in patients with heart failure. Int J
Cardiol. 168:4313–4314. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Batenburg WW, Lu X, Leijten F, Maschke U,
Muller DN and Danser AH: Renin- and prorenin-induced effects in rat
vascular smooth muscle cells overexpressing the human (pro)renin
receptor: does (pro)renin-(pro)renin receptor interaction actually
occur? Hypertension. 58:1111–1119. 2011. View Article : Google Scholar
|
|
47
|
Sakoda M, Ichihara A, Kaneshiro Y, et al:
(Pro)renin receptor-mediated activation of mitogen-activated
protein kinases in human vascular smooth muscle cells. Hypertens
Res. 30:1139–1146. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Saris JJ, van den Eijnden MM, Lamers JM,
Saxena PR, Schalekamp MA and Danser AH: Prorenin-induced myocyte
proliferation: no role for intracellular angiotensin II.
Hypertension. 39:573–577. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Uraoka M, Ikeda K, Nakagawa Y, et al:
Prorenin induces ERK activation in endothelial cells to enhance
neovascularization independently of the renin-angiotensin system.
Biochem Biophys Res Commun. 390:1202–1207. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kinouchi K, Ichihara A, Sano M, et al: The
(pro)renin receptor/ATP6AP2 is essential for vacuolar
H+-ATPase assembly in murine cardiomyocytes. Circ Res.
107:30–34. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Riediger F, Quack I, Qadri F, et al:
Prorenin receptor is essential for podocyte autophagy and survival.
J Am Soc Nephrol. 22:2193–2202. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Amsterdam A, Nissen RM, Sun Z, Swindell
EC, Farrington S and Hopkins N: Identification of 315 genes
essential for early zebrafish development. Proc Natl Acad Sci USA.
101:12792–12797. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liang P, Jones CA, Bisgrove BW, et al:
Genomic characterization and expression analysis of the first
nonmammalian renin genes from zebrafish and pufferfish. Physiol
Genomics. 16:314–322. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Reudelhuber TL: The interaction between
prorenin, renin and the (pro)renin receptor: time to rethink the
role in hypertension. Curr Opin Nephrol Hypertens. 21:137–141.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Bader M: The (pro)renin receptor,
(P)RR/ATP6AP2, a bifunctional protein? J Renin Angiotensin
Aldosterone Syst. 9(Suppl 1): S52008.
|
|
56
|
Juillerat-Jeanneret L, Celerier J, Chapuis
Bernasconi C, et al: Renin and angiotensinogen expression and
functions in growth and apoptosis of human glioblastoma. Br J
Cancer. 90:1059–1068. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Rusin A, Krawczyk Z, Grynkiewicz G, Gogler
A, Zawisza-Puchalka J and Szeja W: Synthetic derivatives of
genistein, their properties and possible applications. Acta Biochim
Pol. 57:23–34. 2010.PubMed/NCBI
|
|
58
|
Soucy NV, Parkinson HD, Sochaski MA and
Borghoff SJ: Kinetics of genistein and its conjugated metabolites
in pregnant Sprague-Dawley rats following single and repeated
genistein administration. Toxicol Sci. 90:230–240. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Spinozzi F, Pagliacci MC, Migliorati G, et
al: The natural tyrosine kinase inhibitor genistein produces cell
cycle arrest and apoptosis in Jurkat T-leukemia cells. Leuk Res.
18:431–439. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Su Y, Simmen FA, Xiao R and Simmen RC:
Expression profiling of rat mammary epithelial cells reveals
candidate signaling pathways in dietary protection from mammary
tumors. Physiol Genomics. 30:8–16. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhang Y and Chen H: Genistein, an
epigenome modifier during cancer prevention. Epigenetics.
6:888–891. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Gullett NP, Ruhul Amin AR, Bayraktar S, et
al: Cancer prevention with natural compounds. Semin Oncol.
37:258–281. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Pavese JM, Farmer RL and Bergan RC:
Inhibition of cancer cell invasion and metastasis by genistein.
Cancer Metastasis Rev. 29:465–482. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Rusin A, Zawisza-Puchalka J, Kujawa K, et
al: Synthetic conjugates of genistein affecting proliferation and
mitosis of cancer cells. Bioorg Med Chem. 19:295–305. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
El-Rayes BF, Philip PA, Sarkar FH, et al:
A phase II study of isoflavones, erlotinib, and gemcitabine in
advanced pancreatic cancer. Invest New Drugs. 29:694–699. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
McSheehy PM, Troy H, Kelland LR, Judson
IR, Leach MO and Griffiths JR: Increased tumour extracellular pH
induced by Bafilomycin A1 inhibits tumour growth and mitosis in
vivo and alters 5-fluorouracil pharmacokinetics. Eur J Cancer.
39:532–540. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kinoshita K, Waritani T, Noto M, et al:
Bafilomycin A1 induces apoptosis in PC12 cells independently of
intracellular pH. FEBS Lett. 398:61–66. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ohta T, Arakawa H, Futagami F, et al:
Bafilomycin A1 induces apoptosis in the human pancreatic cancer
cell line Capan-1. J Pathol. 185:324–330. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Lee CM and Tannock IF: Inhibition of
endosomal sequestration of basic anticancer drugs: influence on
cytotoxicity and tissue penetration. Br J Cancer. 94:863–869. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
http://www.clinicaltrials.gov/ct2/results?term=bafilomycin&Search=Search.
accessed 19/12/2013
|
|
71
|
Elmarakby AA, Ibrahim AS, Faulkner J,
Mozaffari MS, Liou GI and Abdelsayed R: Tyrosine kinase inhibitor,
genistein, reduces renal inflammation and injury in
streptozotocin-induced diabetic mice. Vascul Pharmacol. 55:149–156.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhong WW, Liu Y and Li CL: Mechanisms of
genistein protection on pancreas cell damage in high glucose
condition. Intern Med. 50:2129–2134. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Hettiarachchi KD, Zimmet PZ and Myers MA:
The effects of repeated exposure to sub-toxic doses of
plecomacrolide antibiotics on the endocrine pancreas. Food Chem
Toxicol. 44:1966–1977. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Myers MA, Hettiarachchi KD, Ludeman JP,
Wilson AJ, Wilson CR and Zimmet PZ: Dietary microbial toxins and
type 1 diabetes. Ann NY Acad Sci. 1005:418–422. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Lammi N, Karvonen M and Tuomilehto J: Do
microbes have a causal role in type 1 diabetes? Med Sci Monit.
11:RA63–RA69. 2005.PubMed/NCBI
|
|
76
|
Klein CB and King AA: Genistein
genotoxicity: critical considerations of in vitro exposure dose.
Toxicol Appl Pharmacol. 224:1–11. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Labbaye C, Spinello I, Quaranta MT, et al:
A three-step pathway comprising PLZF/miR-146a/CXCR4 controls
megakaryopoiesis. Nat Cell Biol. 10:788–801. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhang W and Liu HT: MAPK signal pathways
in the regulation of cell proliferation in mammalian cells. Cell
Res. 12:9–18. 2002. View Article : Google Scholar
|
|
79
|
Barker N and Clevers H: Mining the Wnt
pathway for cancer therapeutics. Nat Rev Drug Discov. 5:997–1014.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Sennoune SR, Luo D and Martinez-Zaguilan
R: Plasmalemmal vacuolar-type H+-ATPase in cancer
biology. Cell Biochem Biophys. 40:185–206. 2004. View Article : Google Scholar
|
|
81
|
Huang Y, Noble NA, Zhang J, Xu C and
Border WA: Renin-stimulated TGF-beta1 expression is regulated by a
mitogen-activated protein kinase in mesangial cells. Kidney Int.
72:45–52. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Balakumar P and Jagadeesh G: Potential
cross-talk between (pro)renin receptors and Wnt/frizzled receptors
in cardiovascular and renal disorders. Hypertens Res. 34:1161–1170.
2011. View Article : Google Scholar : PubMed/NCBI
|