Introduction
Acne is one of the most common skin disorders that
affects millions of individuals (1). Propionibacterium acnes (P.
acnes) is a major contributing factor to the inflammatory
component of acne (2). The
Gram-positive, anaerobic, lipophilic bacterium, P. acnes, is
an ubiquitous member of the skin microbiota and is found in
sebaceous follicles located on the face, chest and the back of the
majority of humans (3). Acne
presents with a number of different symptoms, including comedones,
papules, pustules, nodules, cysts and pilosebaceous inflammation
(4). The pathogenesis of acne is
characterized by i) the excess sebum production and hyperplasia of
sebaceous glands; ii) the formation of microcomedones, which is
closely associated with the hyperkeratinization of the follicular
wall and infundibulum; and iii) the induction of inflammatory
reactions, such as the acceleration of cytokine production and
arachidonic acid metabolism in keratinocytes, sebocytes and invaded
inflammatory cells (5).
P. acnes contributes to the inflammatory
reaction of acne by inducing monocytes and keratinocytes to produce
pro-inflammatory cytokines, including tumor necrosis factor
(TNF)-α, and interleukin (IL)-1β and IL-8 (6). The inductino of these cykonines by
P. acnes is mediated by Toll-like receptor (TLR)2 (7). TLRs are transmembrane proteins that
play a critical role in the innate immune response to a variety of
microbial pathogens (8). Pattern
recognition receptors of the TLR protein family have been
identified as P. acnes responsive receptors, and the
expression of TLR2 is elevated in P. acnes-infected
keratinocytes, monocytes and macrophages (9). TLR activation promotes the
production of pro-inflammatory cytokines, chemokines,
prostaglandins and leukotrienes (10).
Although the early colonization with P. acnes
plays an important role in the disease, the exact cause of acne and
how treatment affects the course of the skin disease remain unclear
(11). The many prescription
medications for acne allow for a large number of potential
combination treatments (11).
Therapy targets the 4 factors responsible for lesion formation,
i.e., increased sebum production, hyperkeratinization, colonization
by P. acnes and the resultant inflammatory reaction
(12). Topical antibiotics and
medications with bacteriostatic and anti-inflammatory properties
are effective for treating mild to moderate inflammatory acne
(13). However, several
antibiotics, apart from their antibacterial effects, exert
side-effects such as the suppression of host inflammatory responses
(14–16). Thus, more systematic and safer
agents are required.
Purified bee venom (BV) is a natural toxin produced
by honeybees (Apis mellifera L.). BV has been widely used as
a traditional medicine for various diseases (17,18). Several studies have examined the
biological and pharmacological activities of BV, including its
anti-inflammatory effects anti-rheumatoid arthritis (17), pain-relief effects (19), anti-cancer cell proliferation
(20) and immunomodulatory
activity (21). However, to the
best of our knowledge, the anti-inflammatory effects of BV on P.
acnes-induced inflammatory skin disease in an animal model have
not been reported to date. In the present study, we therefore
demonstrate the potential of BV for use as an alternative to
antibacterial therapy for the treatment of acne.
Materials and methods
Collection of BV
Colonies of natural honeybees (Apis mellifera
L). used in this study were maintained at the National
Academy of Agricultural Science, Wanju, Korea. BV was collected in
collecting device (Chung Jin Biotech Co., Ltd., Ansan, Korea) in a
sterile manner under strict laboratory conditions. In brief, the BV
collector was placed on the hive, and the bees were given enough
electric shocks which caused them to sting a glass plate from which
dried BV was later scraped off. The collected BV was diluted in
cold water and then centrifuged at 10,000 × g for 5 min at 4°C to
discard residues from the supernatant. Purified BV was lyophilized
by a freeze dryer and stored in a refrigerator for later use, as
previoulsy described (32). BV
used in the experiment was confirmed with size exclusion gel
chromatography (AKTA Explorer; GE Healthcare Bio-Sciences,
Piscataway, NJ, USA) by dissolving in 0.02 M phosphate-buffer with
0.25 M NaCl adjusted to pH 7.2 using a Superdex Peptide column
(Amersham Biosciences, Piscataway, NJ, USA and GE Healthcare).
Preparation of bacteria
P. acnes (ATCC 6919; American Type Culture
Collection, Rockville, MD, USA) were obtained from the Korean
Culture Center of Microorganisms (Seoul, Korea) and were cultured
at 37°C on Reinforced Clostridial Medium (BD Diagnostics, Sparks,
MD, USA) under anaerobic conditions at 37°C until reaching
OD600 1.0 (logarithmic growth phase). The log phase
bacterial culture was centrifuged at 5,000 × g at 4°C for 15 min.
Subsequently, the culture supernatant was removed, filtered
(through a 0.22 μm pore size filter), and used immediately or
stored at −20°C. The bacterial pellet was washed 3 times in 100 ml
of phosphate-buffered saline (PBS) and finally suspended in 10 ml
of PBS. The P. acnes suspension was incubated at 80°C for 30
min for heat-killing reaction. The heat-killed P. acnes
suspension was stored at 4°C until use.
Experimental animal model
Eight-week-old ICR mice (n=30) were supplied from
Samtako (Osan, Korea) and were individually housed in polycarbonate
cages and maintained under constant temperature (22±2°C) and
humidity conditions (55%). The mice were allowed free access to
food and water, and were maintained in an environment with a
12:12-h light/dark cycle. All surgical and experimental procedures
used in the current study were approved by the Institutional Review
Board (IRB) committee at the Catholic University of Daegu Medical
Center, Daegu, Korea and conformed to the US National Institutes of
Health guidelines for the care and use of laboratory animals.
The mice were randomly divided into 6 different
groups (5 mice/group) as follows: i) NC, normal control. ii) PA,
living P. acnes [1.0×107 colony-forming units
(CFU)/20 μl in PBS] was intradermally injected into the left ears
of the mice. The right ears received an equal amount (20 μl) of
PBS. iii) PA/Vas, living P. acnes was intradermally injected
into both the left and the right ears. After the injection, 0.05 g
vaseline (Sigma, St. Louis, MO, USA) was applied to the surface of
the skin of the right ear. For the next 3 groups, PA/BV1, PA/BV10
and PA/BV100, living P. acnes was intradermally injected
into both the left and the right ears. After the injection, BV (1,
10 and 100 μg mixed with 0.05 g vaseline) was applied to the
surface of the skin of the right ear in the mice in each group. At
the end of each treatment period (24 h later), the animals were
sacrificed by cervical dislocation and the ears were excised.
Histological analysis
All tissue specimens were fixed in 10% formalin for
at least 24 h at room temperature. Following fixation,
perpendicular sections to the anterior-posterior axis of the wound
were dehydrated in graded ethanol, cleared in xylene and embedded
in paraffin. Thin sections (4-μm-thick) were mounted on glass
slides, dewaxed, rehydrated in distilled water and stained with
hematoxylin and eosin (H&E). As part of the histological
evaluation, a pathologist blinded to the previous treatment
examined all the sections under a light microscope (Nikon, Tokyo,
Japan).
Immunohistochemical staining
Paraffin-embedded tissue sections were
deparaffinized with xylene, dehydrated in gradually decreasing
concentrations of ethanol, and subsequently treated with 3%
hydrogen peroxidase in methanol for 10 min to block endogenous
peroxidase activity. Tissue sections were immersed in 10 mM sodium
citrate buffer (pH 6.0) for 5 min at 95°C. The final step was
repeated using fresh 10 mM sodium citrate solution (pH 6.0). The
sections were allowed to remain in the same solution while cooling
for 20 min and rinsed in PBS. The sections were incubated with
primary antibody (1:100) for 1 h at 37°C. Primary antibodies were
the following: anti-TNF-α (Abcam, Cambridge, UK) and anti-IL-1β
(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). The signal
was visualized using an EnVision System (Dako, Carpinteria, CA,
USA) for 30 min at 37°C. 3,3′-Diaminobenzidine tetrahydrochloride
(DAB) was used as the coloring reagent, while hematoxylin was used
as the counterstain.
Immunofluorescence staining
Paraffin-embedded ear tissue sections were
deparaffinized with xylene and dehydrated in gradually decreasing
concentrations of ethanol. The tissue sections were subsequently
placed in blocking serum (5% bovine serum albumin in PBS) at room
temperature for 1 h. Primary antibody (1:100 dilution) was added
followed by overnight incubation at 4°C, and incubation with
secondary antibody (1:200 dilution) was performed at room
temperature for 4 h. Antibodies used in the experiments were the
following: CD14, TLR2 (Santa Cruz Biotechnology, Inc.) and
anti-mouse, anti-rabbit-biotinylated secondary antibodies
conjugated with FITC or Texas Red (Invitrogen, Carlsbad, CA, USA).
The slides were mounted using VECTASHIELD® Mounting
Medium (VECTOR Laboratories, Inc., Burlingame, CA, USA). Specimens
were examined and photographed using a fluorescence microscope
(Nikon).
Western blot analysis
The tissues were lysed and homogenized in lysis
buffer (50 mM Tris pH 8.0, 150 mM NaCl, 5 mM EDTA, 0.5% NP-40, 100
mM PMSF, 1 M DTT, 10 mg/ml leupeptin and aprotinin; all from
Sigma). Following incubation on ice for 30 min, the samples were
centrifuged at 8,000 × g at 4°C for 30 min, and the supernatant was
collected. The total protein concentration was determined by
Bradford assay (Bio-Rad Laboratories, Richmond, CA, USA). Total
protein (10–50 μg) was separated on 8–12% SDS-polyacrylamide gels
and transferred onto PVDF membranes (Millipore Corp., Billerica,
MA, USA) using a standard SDS-PAGE gel electrophoresis procedure.
The membranes were blocked in 5% skim milk in TBS-T (10 mM Tris,
150 mM NaCl and 0.1% Tween-20) for 2 h at room temperature and
probed with primary antibody for 4 h. A horseradish peroxidase
(HRPO)-conjugated secondary antibody (anti-mouse, anti-rabbit and
anti-goat) was used for detection. Signals were detected using an
enhanced chemiluminescence detection system (Amersham Biosciences,
Piscataway, NJ, USA). The primary antibodies used in this study
were the following: anti-TNF-α from Abcam, and anti-IL-1β and
anti-glyceraldehyde 3-phosphate-dehydrogenase (GAPDH) from Santa
Cruz Biotechnology. All primary antibodies were diluted at 1:1,000.
Signal intensity was quantified using an image analyzer (LAS-3000;
Fujifilm, Tokyo, Japan).
Reverse-transcription polymerase chain
reaction (RT-PCR)
Total RNA was extracted from ear tissue with TRIzol
reagent (Gibco, Grand Island, NY, USA) according to the
manufacturer’s recommendations. The purity and quantity of the RNA
preparation were measured at optical densities of 260 and 280 nm.
First-strand cDNA was synthesized using oligo(dT) primers and M-MLV
reverse-transcriptase (Promega Corp., Madison, WI, USA). An aliquot
of cDNA was used for PCR using primer sets specific to mouse TNF-α,
IL-1β and GAPDH. Primer sequences were as follows: TNF-α forward,
5′-AGT GGT GCC AGC CGA TGG GTT GT-3′ and reverse, 5′-GCT GAG TTG
GTC CCC CTT CTC CAG-3′; IL-1β forward, 5′-CAT GAG CAC CTT CTT TTC
CT-3′ and reverse, 5′-TGT ACC AGT TGG GGA ACT CT-3′; GAPDH forward,
5′-GTG GAC ATT GTT GCC ATC AAC G-3′ and reverse, 5′-GAG GGA GTT GTC
ATA TTT CTC G-3′. PCR products were visualized by 1.5% agarose gel
electrophoresis with ethidium bromide staining.
Nuclear protein extraction
Tissues were trypsinized and incubated for 15 min in
a hypo-osmotic buffer (10 mM HEPES, pH 7.9, 10 mM KCI, 1.5 mM
MgCl2, 0.2 mM EDTA, 25% glycerol, 0.5 mM DTT) for 20
min, and vortexed and centrifuged at 5,000 × g for 15 min at 4°C.
The nuclear supernatant was collected as the nuclear lysate
fraction. Each buffer also contained 0.2 mM phenylmethanesulfonyl
fluoride (PMSF) and 10 μg/ml protease inhibitors.
Electrophoretic mobility shift assay
(EMSA)
A DIG Gel Shift kit (Roche, Mannheim, Germany) was
used for EMSA. The nuclear factor-κB (NF-κB) and activator protein
(AP)-1 oligonucleotide probe (NF-κB, 5′-AGT TGA GGG GAC TTT CCC AGG
C-3′, AP-l, 5′-CGC TTG ATG ATG CAG CCG GAA-3′; only sense strands
are shown) was end-labeled with DIG-ddPUT. For binding reactions, 9
μg nuclear extract protein was mixed with binding buffer (0.5 μg
poly dI-dC, 0.1 μg poly L-lysine and 0.8 μg labeled
oligonucleotide, final volume of 20 μl) and was incubated at 37°C
for 30 min. The nuclear protein-DNA complex was separated on a 6%
non-denaturing polyacrylamide gel in TBE buffer at 80 V for 1.5–2 h
at 4°C. The samples were transferred to Hybond-XL membranes
(Amersham Biosciences) by electroblotting for 30 min. The membranes
were cross-linked for 10 min and washed with solution containing
100 mM maleic acid, 150 mM NaCl and 0.3% Tween-20, pH 7.5 for 10
min, were then moved to blocking solution (DIG Gel Shift kit) for
40 min. The membranes were incubated with anti-digoxigenin-AP Fab
fragments (1:10,000) for 30 min, and the nuclear protein-DNA bands
were developed with detection solution including 100 mM Tris-HCl,
100 mM NaCl, pH 9.5 and 100 μg/ml disodium 3-{4-methoxyspiro
[1,2-dioxetane-3,2′(5′-chloro)tricyclo(3.3.1.13,7)decan]-4-yl}
phenyl phosphate. NF-κB and AP-1 bands from the films were
quantitated using an image analyzer (LAS-3000; Fujifilm).
Statistical analysis
Data are presented as the means ± SE. The Student’s
t-test was used to assess the significance of differences between
independent experiments. A value of p<0.05 was considered to
indicate a statistically significant difference.
Results
Effects of BV on P. acnes-induced
inflammatory skin disease
To investigate the therapeutic effects of BV against
P. acnes-induced inflammatory skin disease, P. acnes
was intradermally injected into both the left and the right ears of
ICR mice. After the injection, various concentrations of BV (1, 10
and 100 μg) mixed with 0.05 g of vaseline were applied to the
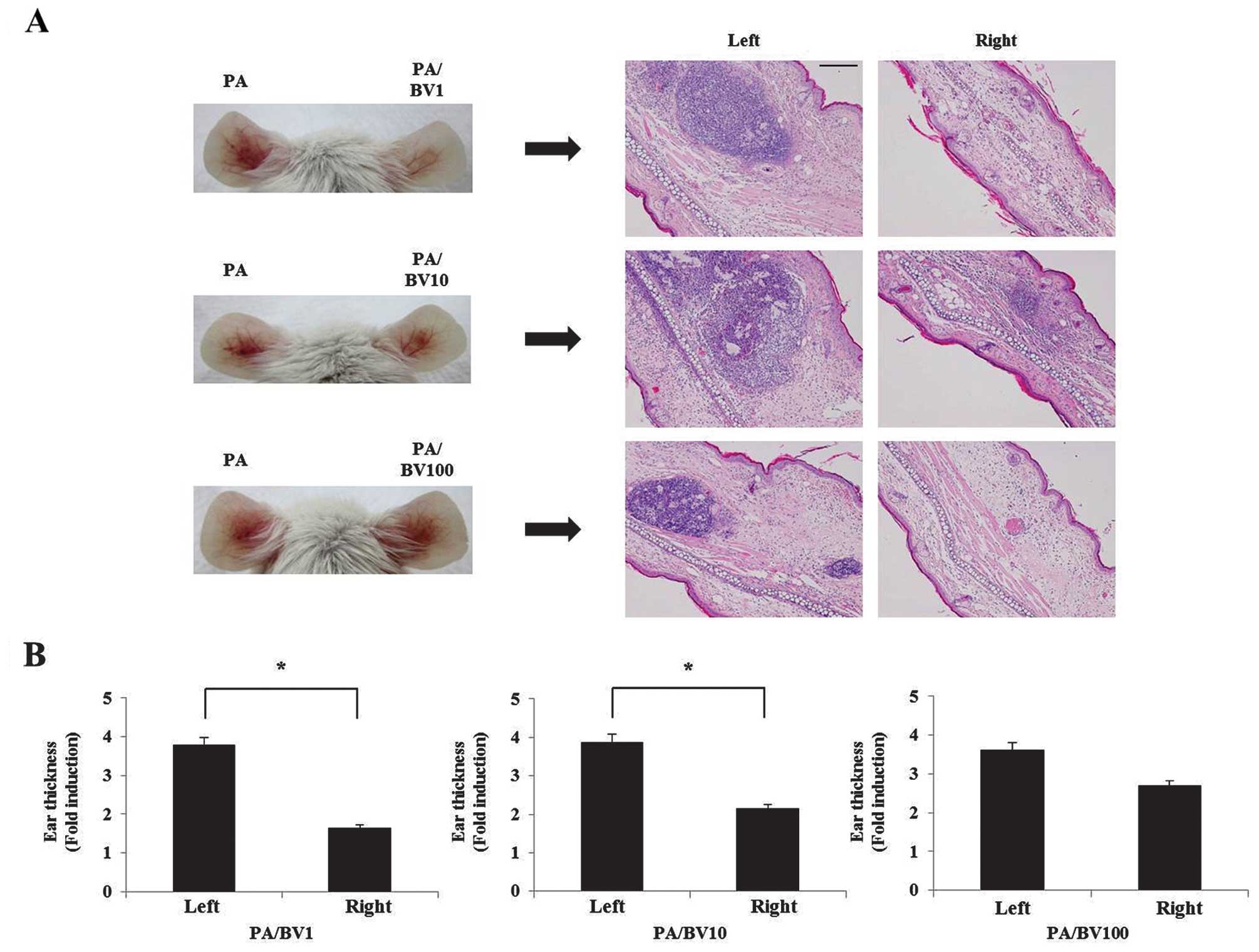
surface of the skin of the right ear. As shown in Fig. 1A, significant ear swelling,
redness and erythema were observed 24 h after the P. acnes
injection. Histological observation revealed that the P.
acnes injection induced a considerable increase in the number
of infiltrated inflammatory cells. By contrast, the BV-treated ears
showed noticeably reduced ear thickness, swelling, erythema and
inflammatory reactions. In particular, treatment with 1 μg of BV
resulted in a 3-fold reduction of ear thickness compared to the
ears injected only with living P. acnes (Fig. 1B). Based on these results, 1 μg of
BV was used in the subsequent experiments.
BV suppresses pro-inflammatory cytokine
expression
It has previously been demonstrated that P.
acnes stimulates the production of pro-inflammatory cytokines,
such as TNF-α and IL-1β (6).
Therefore, we investigated whether TNF-α and IL-1β are involved in
the inhibitory effects of BV on P. acnes-induced
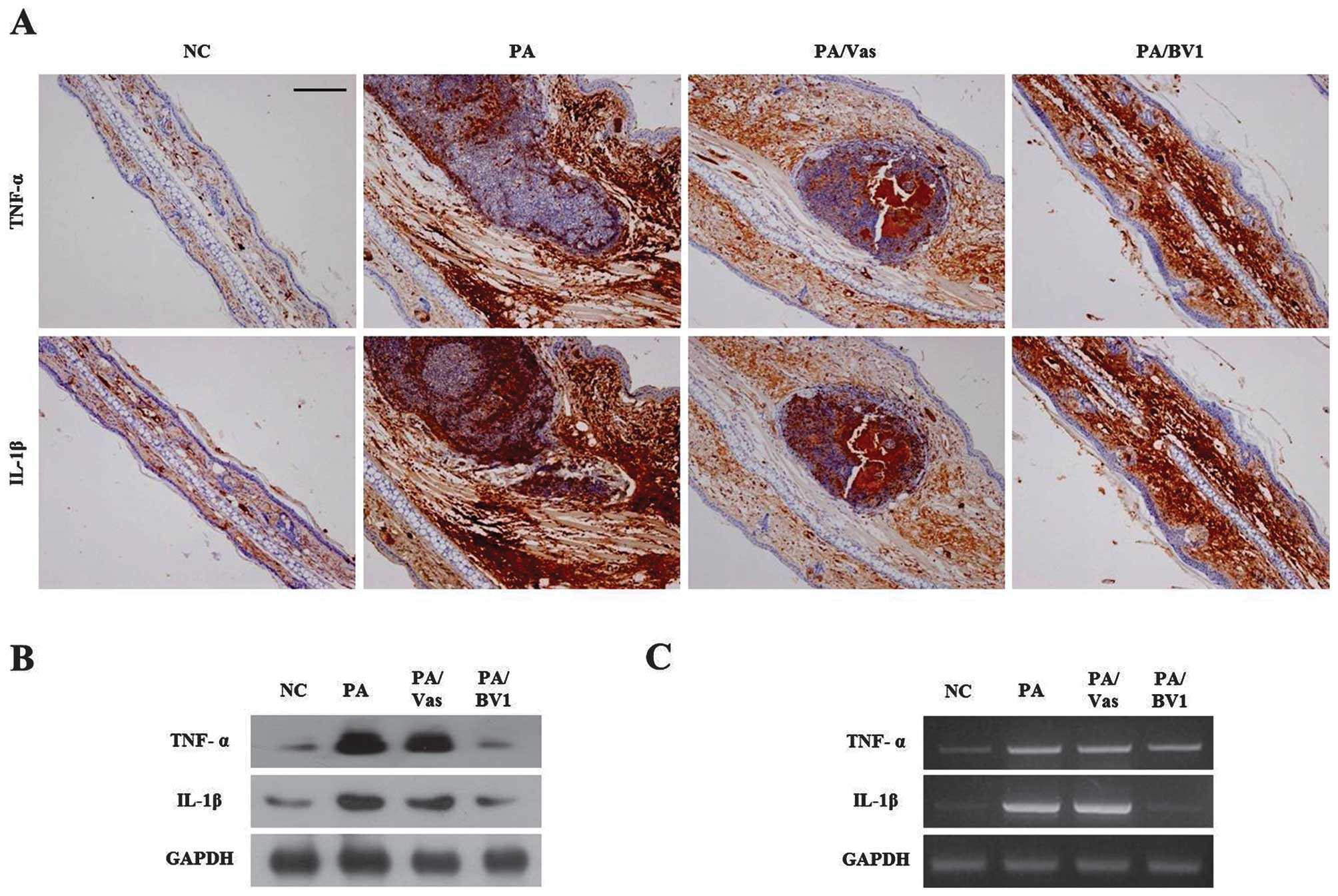
inflammatory skin disease. Immunohistochemical staining for TNF-α
and IL-1β revealed that the expression levels of these cytokines
were barely detectable in the normal skin tissue from the NC group.
As shown in Fig. 2A, the PA and
PA/Vas groups showed a marked increase in the number of TNF-α and
IL-1β-positive cells compared to the NC group. However, treatment
with BV significantly inhibited the number of TNF-α- and
IL-1β-positive cells compared to the PA and PA/Vas groups.
Consistent with immunohistochemical staining, the results from
western blot analysis revealed that treatment with BV suppressed
the expression levels of these cytokines in the inflamed tissue
(Fig. 2B). Moreover, the results
from RT-PCR also indicated that treatment with BV markedly
inhibited the expression of TNF-α and IL-1β in the PA/BV1 group
(Fig. 2C). These observations
suggested that BV effectively inhibited the inflammatory events in
the animal model of P. acnes-induced inflammatory skin
disease.
BV inhibits TLR2 expression in inflamed
skin tissue
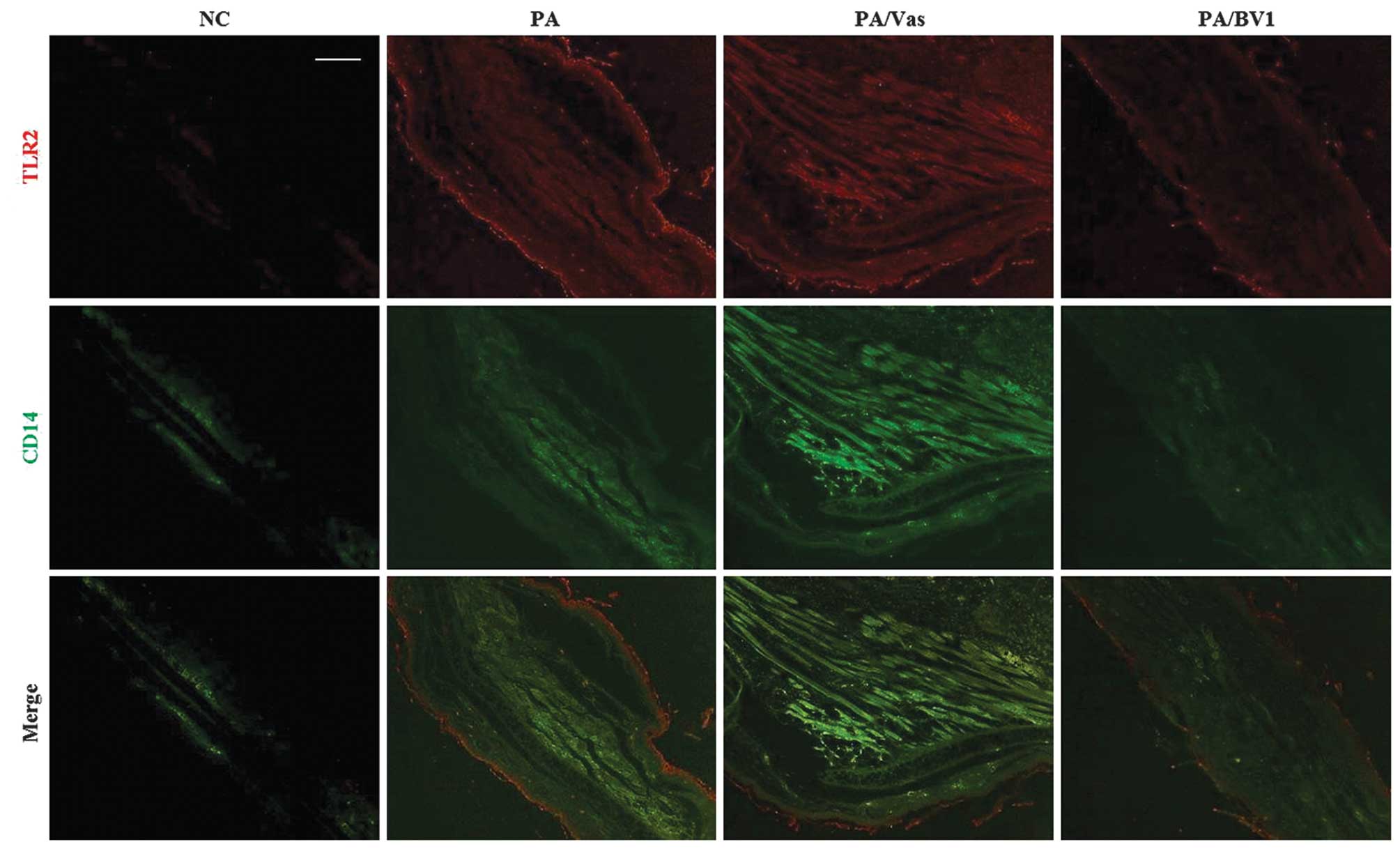
To determine whether BV effectively blocks TLR2 and
the macrophage marker of CD14, we performed double
immunoflurorescence staining. As shown in Fig. 3, the PA and PA/Vas groups showed
significantly higher TLR2 and CD14 expression levels than the NC
group. In addition, merged images show that the PA group
predominantly expressed CD14 in the dermis and TLR2 in the
epidermis, both of which are localized at different parts of the
tissue. These results demonstrated that P. acnes induced the
infiltration of macrophages and the activation of TLR2 in the
inflamed skin tissue. By contrast, treatment with BV significantly
inhibited TLR2 and CD14 expression in the P. acnes-injected
tissue. Therefore, BV effectively blocks the expression of TLR2 and
CD14 in P. acnes-induced inflammatory skin disease.
BV inhibits the DNA-binding activity of
transcription factors
Pro-inflammatory cytokines are regulated at the
transcriptional levels by several transcriptional factors,
including AP-1 and NF-κB (22).
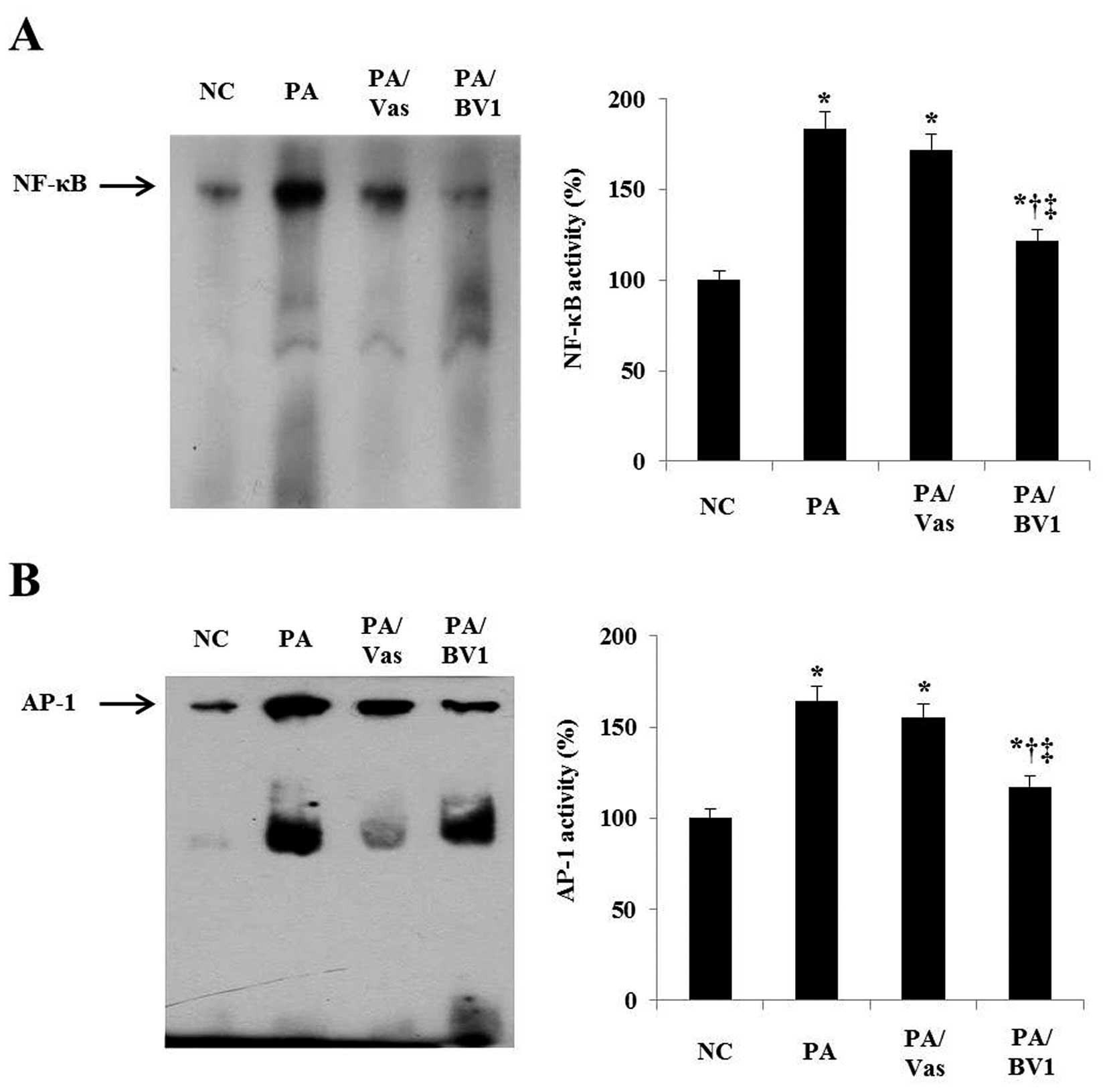
Therefore, we examined the DNA-binding activity of NF-κB and AP-1
in inflamed skin tissue treated with or without BV by EMSA. As
shown in Fig. 4A and B, the
expression of NF-κB and AP-1 was increased in the PA and PA/Vas
groups, but those levels were significantly inhibited in the PA/BV1
group. These results indicate that BV inhibits the expression of
inflammatory cytokines, such as TNF-α and IL-1β by suppressing the
DNA-binding activity of NF-κB and AP-1 in the inflamed skin tissue.
Therefore, these results suggest that BV is effective in inhibiting
NF-κB and AP-1 activation in inflamed skin tissue, which provides
the molecular basis for the decreased expression of
chemokines/cytokines in the inflamed skin treated with BV following
P. acnes injection.
Discussion
The majority of serious skin inflammatory diseases
are due to infection by various microorganisms, including the
Gram-positive P. acnes and Staphylococcus and the
Gram-negative, Pseudomonas aeruginosa (23,24). Colonization of the pilosebaceous
follicle by P. acnes is a major factor for the inflammatory
reaction in acne vulgaris. Therefore, P. acnes has been a
major target of therapy in inflammatory acne (25). A number of skin disorders with an
inflammatory component are treated with steroids and/or antibiotics
in both oral and topical formulations (26). Topical therapies, including
retinoids, benzoyl peroxide, azelaic acid and antibiotics, when
used in combination, usually improve the condition of mild to
moderate acne (11,23). However, the long-term use of these
drugs has potential side-effects, such as hypertension,
immunosuppression (steroids) and the development of antibiotic
resistance (27–29). Thus, safer and natural agents are
required.
BV, a natural toxinm contains a variety of peptides,
including melittin, apamin, adolapin and mast cell degranulating
peptide along with enzymes, biological amines and non-peptide
components (18). A high
concentration of BV (100 μg/ml) may lead to the instability of
human lymphocytes. By contrast, low concentrations of BV are not
genotoxic and do not produce oxidative damage (30). As a traditional alternative
medical approach, BV therapy has been used to reduce pain and in
the treatment of inflammatory diseases, such as rheumatoid
arthritis and multiple sclerosis since ancient times (17). In a previous study, we
investigated whether BV reduces atherosclerotic lesion formation
via anti-inflammatory mechanisms (31). Previous studies have also
demonstrated the antimicrobial activities of BV against P.
acnes-induced inflammation in vitro (32). However, the anti-inflammatory
effects of BV on P. acnes-induced inflammatory skin disease
using an animal model have not yet been fully elucidated.
In the present study, we investigated whether BV
regulates the expression of pro-inflammatory-related genes in a
mouse model of skin tissue inflammation. A previous study suggested
that P. acnes induces inflammation through the activation of
TLR, particularly TLR2 and TLR4 (10). The activation of TLR2 and TLR4 has
been shown to stimulate the release of pro-inflammatory cytokines,
such as IL-1α, IL-1β, IL-6, IL-8 and TNF-α (33). More specifically, TLR2 mediates
host responses to peptidoglycan from Gram-positive bacteria,
whereas TLR4 mediates host responses to lipopolysaccharide (LPS)
from Gram-negative bacteria (34). The inflammation triggered through
TLR2 is important in the pathogenesis of acne (25). Therefore, the regulation of TLR2
expression may provide a feasible target for the treatment of
inflammatory acne lesions (2). To
examine this hypothesis, we determined the effects of BV on the
expression of the pro-inflammatory cytokines, TNF-α and IL-1β, as
well as TLR2 and CD14 in inflamed skin tissue. Our data indicate
that BV is able to reduce various inflammatory events observed in
the skin layers. Of note, BV regulated the production of TNF-α and
IL-1β at the protein level and downregulated the mRNA expression of
TNF-α and IL-1β. Consistent with these findings, TLR2 and CD14
expression was also similarly suppressed by treatment with BV.
Monocytes, macrophages, sebocytes and keratinocytes
are stimulated by P. acnes through a TLR2-mediated NF-κB
signaling pathway during inflammatory reactions in skin (10,35,36). The activation of TLR can also lead
to the activation of other transcription factors, such as AP-1
(37). The transcription factors,
NF-κB and AP-1, have been reported to be activated in acne lesions
(38). In this study, to explore
the potential mechanisms underlying the inhibitory effects of BV on
cytokine production, we investigated the nuclear activation of
NF-κB and AP-1 in P. acnes-induced skin inflammation. The
injection of P. acnes potentially increased the expression
of nuclear NF-κB and AP-1 in the skin tissue. However, treatment
with BV markedly reduced the nuclear expression of NF-κB and AP-1
in the inflamed skin. Our results indicate that BV exerts an
inhibitory effect against inflammatory skin diseases.
In conlcusion, we demonstrate that BV blocks several
skin inflammatory responses, such as the production of TNF-α and
IL-1β. The most important findings of this study are that BV
inhibits the P. acnes-stimulated production and expression
of pro-inflammatory cytokines through the modulation of
TLR2-mediated NF-κB and AP-1 signaling in inflamed skin tissue.
Taken together, our results indicate that BV exerts an inhibitory
effect on inflammatory skin diseases. Moreover, this in vivo
acne-like model may be useful for the pharmacological evaluation of
anti-acne agents. In conclusion, this study suggests that BV has
potential for use as an anti-acne agent and may be useful in the
pharmaceutical and cosmetics industries. Further studies are
required in order to fully elucidate the efficacy and mechanisms of
action of individual constituents of BV for the treatment of P.
acnes-induced inflammation.
Acknowledgements
This study was supported by a grant from the
Next-Generation BioGreen 21 Program (no. PJ009519), Rural
Development Administration, Korea.
References
|
1
|
Tsai HH, Lee WR, Wang PH, Cheng KT, Chen
YC and Shen SC: Propionibacterium acnes-induced iNOS and
COX-2 protein expression via ROS-dependent NF-κB and AP-1
activation in macrophages. J Dermatol Sci. 69:122–131. 2013.
View Article : Google Scholar
|
|
2
|
Jung MK, Ha S, Son JA, et al:
Polyphenon-60 displays a therapeutic effect on acne by suppression
of TLR2 and IL-8 expression via down-regulating the ERK1/2 pathway.
Arch Dermatol Res. 304:655–663. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mak TN, Fischer N, Laube B, et al:
Propionibacterium acnes host cell tropism contributes to
vimentin-mediated invasion and induction of inflammation. Cell
Microbiol. 14:1720–1733. 2012. View Article : Google Scholar
|
|
4
|
Nakatsuji T, Kao MC, Fang JY, et al:
Antimicrobial property of lauric acid against Propionibacterium
acnes: its therapeutic potential for inflammatory acne
vulgaris. J Invest Dermatol. 129:2480–2488. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pawin H, Beylot C, Chivot M, et al:
Physiopathology of acne vulgaris: recent data, new understanding of
the treatments. Eur J Dermatol. 14:4–12. 2004.PubMed/NCBI
|
|
6
|
Vowels BR, Yang S and Leyden JJ: Induction
of proinflammatory cytokines by a soluble factor of
Propionibacterium acnes: implications for chronic
inflammatory acne. Infect Immun. 63:3158–3165. 1995.PubMed/NCBI
|
|
7
|
Kim J: Review of the innate immune
response in acne vulgaris: activation of Toll-like receptor 2 in
acne triggers inflammatory cytokine responses. Dermatology.
211:193–198. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Medzhitov R, Preston-Hurlburt P and
Janeway CA Jr: A human homologue of the Drosophila Toll protein
signals activation of adaptive immunity. Nature. 388:394–397. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jugeau S, Tenaud I, Knol AC, et al:
Induction of toll-like receptors by Propionibacterium acnes.
Br J Dermatol. 153:1105–1113. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim J, Ochoa MT, Krutzik SR, et al:
Activation of toll-like receptor 2 in acne triggers inflammatory
cytokine responses. J Immunol. 169:1535–1541. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Williams HC, Dellavalle RP and Garner S:
Acne vulgaris. Lancet. 379:361–372. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Titus S and Hodge J: Diagnosis and
treatment of acne. Am Fam Physician. 86:734–740. 2012.
|
|
13
|
Feldman S, Careccia RE, Barham KL and
Hancox J: Diagnosis and treatment of acne. Am Fam Physician.
69:2123–2130. 2004.
|
|
14
|
Labro MT: Immunomodulation by
antibacterial agents. Is it clinically relevant? Drugs. 45:319–328.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ayangco L and Sheridan PJ:
Minocycline-induced staining of torus palatinus and alveolar bone.
J Periodontol. 74:669–671. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ramakrishna J, Johnson AR and Banner BF:
Long-term minocycline use for acne in healthy adolescents can cause
severe autoimmune hepatitis. J Clin Gastroenterol. 43:787–790.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Billingham ME, Morley J, Hanson JM,
Shipolini RA and Vernon CA: Letter: An anti-inflammatory peptide
from bee venom. Nature. 245:163–164. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Son DJ, Lee JW, Lee YH, Song HS, Lee CK
and Hong JT: Therapeutic application of anti-arthritis,
pain-releasing, and anti-cancer effects of bee venom and its
constituent compounds. Pharmacol Ther. 115:246–270. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kwon YB, Lee JD, Lee HJ, et al: Bee venom
injection into an acupuncture point reduces arthritis associated
edema and nociceptive responses. Pain. 90:271–280. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pérez-Payá E, Houghten RA and Blondelle
SE: The role of amphipathicity in the folding, self-association and
biological activity of multiple subunit small proteins. J Biol
Chem. 270:1048–1056. 1995.PubMed/NCBI
|
|
21
|
Park HJ, Lee SH, Son DJ, et al:
Antiarthritic effect of bee venom: inhibition of inflammation
mediator generation by suppression of NF-kappaB through interaction
with the p50 subunit. Arthritis Rheum. 50:3504–3515. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Khalaf H, Jass J and Olsson PE:
Differential cytokine regulation by NF-kappaB and AP-1 in Jurkat
T-cells. BMC Immunol. 11:262010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dryden MS: Skin and soft tissue infection:
microbiology and epidemiology. Int J Antimicrob Agents. 34(Suppl
1): S2–S7. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sohail MR, Gray AL, Baddour LM, Tleyjeh IM
and Virk A: Infective endocarditis due to Propionibacterium
species. Clin Microbiol Infect. 15:387–394. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dessinioti C and Katsambas AD: The role of
Propionibacterium acnes in acne pathogenesis: facts and
controversies. Clin Dermatol. 28:2–7. 2010.
|
|
26
|
Grange PA, Raingeaud J, Calvez V and Dupin
N: Nicotinamide inhibits Propionibacterium acnes-induced
IL-8 production in keratinocytes through the NF-kappaB and MAPK
pathways. J Dermatol Sci. 56:106–112. 2009.PubMed/NCBI
|
|
27
|
Fivenson DP, Breneman DL, Rosen GB, Hersh
CS, Cardone S and Mutasim D: Nicotinamide and tetracycline therapy
of bullous pemphigoid. Arch Dermatol. 130:753–758. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zemtsov A and Neldner KH: Successful
treatment of dermatitis herpetiformis with tetracycline and
nicotinamide in a patient unable to tolerate dapsone. J Am Acad
Dermatol. 28:505–506. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Handfield-Jones S, Jones S and Peachey R:
High dose nicotinamide in the treatment of necrobiosis lipoidica.
Br J Dermatol. 118:693–696. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee WR, Park JH, Kim KH, Park YY, Han SM
and Park KK: Protective effects of melittin on transforming growth
factor-beta1 injury to hepatocytes via anti-apoptotic mechanism.
Toxicol Appl Pharmacol. 256:209–215. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee WR, Kim SJ, Park JH, et al: Bee venom
reduces atherosclerotic lesion formation via anti-inflammatory
mechanism. Am J Chin Med. 38:1077–1092. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Han SM, Lee KG, Yeo JH, Baek HJ and Park
KK: Antibacterial and anti-inflammatory effects of honeybee
(Apis mellifera) venom against acne-inducing bacteria. J Med
Plants Res. 4:459–464. 2010.
|
|
33
|
Lyte P, Sur R, Nigam A and Southall MD:
Heat-killed Propionibacterium acnes is capable of inducing
inflammatory responses in skin. Exp Dermatol. 18:1070–1072.
2009.
|
|
34
|
Mandell L, Moran AP, Cocchiarella A, et
al: Intact gram-negative Helicobacter pylori,
Helicobacter felis, and Helicobacter hepaticus
bacteria activate innate immunity via toll-like receptor 2 but not
toll-like receptor 4. Infect Immun. 72:6446–6454. 2004.PubMed/NCBI
|
|
35
|
Köllisch G, Kalali BN, Voelcker V, et al:
Various members of the Toll-like receptor family contribute to the
innate immune response of human epidermal keratinocytes.
Immunology. 114:531–541. 2005.PubMed/NCBI
|
|
36
|
Kurokawa I, Danby FW, Ju Q, et al: New
developments in our understanding of acne pathogenesis and
treatment. Exp Dermatol. 18:821–832. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Scharf S, Hippenstiel S, Flieger A,
Suttorp N and N’Guessan PD: Induction of human β-defensin-2 in
pulmonary epithelial cells by Legionella pneumophila:
involvement of TLR2 and TLR5, p38 MAPK, JNK, NF-kappaB, and AP-1.
Am J Physiol Lung Cell Mol Physiol. 298:L687–L695. 2010.
|
|
38
|
Kang S, Cho S, Chung JH, Hammerberg C,
Fisher GJ and Voorhees JJ: Inflammation and extracellular matrix
degradation mediated by activated transcription factors nuclear
factor-kappaB and activator protein-1 in inflammatory acne lesions
in vivo. Am J Pathol. 166:1691–1699. 2005. View Article : Google Scholar
|