Introduction
Varicose veins are a common venous disease, with a
prevalence of 8.6% among individuals over 15 years of age and as
high as 16.4% among individuals over 45 years of age in China
(1). Varicose veins are
superficial vessels in the lower extremities that are abnormally
twisted, lengthened and dilated, and are often associated with
incompetent valves within the vein (2). Symptoms of varicosity include
swelling, restlessness, limb heaviness and fatigue, an
aching/throbbing sensation, burning, tingling, direct tenderness,
itching and nocturnal leg cramps (3–5).
Furthermore, varicose veins are demonstrated to be
autosomal-dominant with incomplete penetrance. Offspring have a 90%
chance of developing varicose veins if their parents have them
(6–9).
The IQ-domain GTPase-activating protein (IQGAP)
family of proteins is found in multiple organisms, including yeast,
fish, Xenopus and mammals. Three IQGAP proteins, including
IQGAP1, IQGAP2 and IQGAP3, have been identified in humans. All
three IQGAPs are comprised of distinct domains, comprising a
homology domain (CHD), a polyproline binding region (WW), an IQ
domain and a RasGTPase-activating protein-related domain (GRD)
(10).
To date, IQGAP1 is the most well characterized of
the IQGAP proteins. A list of proteins binding to IQGAP1 has been
previously reviewed (10).
Nevertheless, the binding of several of these proteins exhibits
unique features. A detailed investigation with a panel of >20
mutant proteins revealed that the interactions of Cdc42 and Rac1
with IQGAP1 differ considerably from their interactions with other
binding proteins (11). Moreover,
IQGAP1 is a scaffold protein modulating the MEK/extracellular
signal-regulated kinase (ERK) signaling pathway. IQGAP1 has been
shown to bind directly to both MEK1/2 and ERK1/2 and modulates
their activation (12,13). Subsequent studies demonstrated
that IQGAP1 also interacts with MAPK components proximal to MEK,
which is known as Raf (14).
Consistent with these findings, IQGAP1 is required for VEGF to
activate B-Raf in vascular endothelial cells (15). Additionally, IQGAP1 is involved in
the regulation of cadherin-mediated cell-cell adhesion, cell
polarization and cell migration (16).
The abnormal growth of vascular smooth muscle cells
(VSMCs) plays an essential role in intimal formation in the early
stages of atherosclerosis and restenosis, as well as in varicosity
(17). Differences in phenotype
and function between VSMCs derived from normal veins and those
derived from varicose veins have been identified. VSMCs derived
from varicose veins are characterized by significantly increased
capabilities of proliferation, migration and synthesis (18). In a previous study, it was
demonstrated that VSMCs in varicose veins were poorly
differentiated, with an increase in secretory cytoplasmic
organelles, which possibly reflects the unusual synthetic and
secretory roles of VSMCs. A reduction in filament bundles was also
observed, which suggests a decreased contractility of VSMCs in
varicose veins (19).
Currently, a variety of factors have been shown to
modulate the VSMC phenotype, such as microRNAs (miRNAs) (20), estrogen (21), oxidative stress (22) and Ca2+
(23). Among these, the dual
effects of estrogen on VSMC proliferation and migration have been
reported (24). However, the
exact mechanisms involved have not yet been elucidated. In this
study, lesion VSMCs were isolated from patients with varicose veins
and exposed to 10−8 M 17β-estradiol (E2).
Exposure to E2 induced a differential effect on the
VSMCs obtained from varicose veins and those obtained from normal
veins. Further examination revealed that IQGAP1 expression was
significantly increased in the VSMCs obtained from varicose veins
compared with the normal VSMCs. The knockdown of IQGAP1 reduced
VSMC proliferation and migration, while the overexpression of
IQGAP1 promoted both proliferation and migration. The
overexpression of IQGAP1 enhanced estrogen receptor (ER)
transcriptional activity in the VSMCs obtained from varicose veins.
Additionally, elevated levels of phosphorylated Akt and ERK were
observed in the VSMCs obtained from varicose veins.
Materials and methods
A total of 11 patients (7 females and 4 males) with
varicose veins were enrolled in the present study carried out at
the Affiliated Suzhou Hospital of Nanjing Medical University,
Suzhou, China. The age of the patients ranged from 51 to 85 years,
with an average age of 54.1 years. Varicose veins were obtained
during classic vascular stripping surgery. Surplus normal segments
of saphenous veins were obtained from 9 subjects (5 females and 4
males) and served as the non-varicose control veins. In all cases,
the vena saphena magna was harvested at a location from 5 to
15 cm below the knee. No significant differences in mean age were
obseved between the patients with varicose veins and the controls
(mean age, 54.1±8.7 and 51.3±9.2 years, respectively). Human
umbilical veins were obtained from women (7 subjects; mean age,
27.4±5.9 years) with normal pregnancies after vaginal delivery. The
present study was approved by the Ethics Committee of Suzhou
Hospital Affiliated to Nanjing Medical University. Informed consent
was obtained from all patients prior to enrollment.
Cell culture
The prepared VSMCs were cultured in phenol red-free
DMEM containing 10% fetal calf serum, 100 units/ml penicillin, 100
pg/ml streptomycin and 4 mM L-glutamine at 37°C in 5%
CO2. The cultures used for the present study had a VSMC
purity of >90% (determined by immunocytochemical staining for
α-smooth muscle actin). For PI3K or ERK inhibition, VSMCs from
varicose veins or normal controls were treated with 10 μM
LY294002 or 10 μM PD98059 for 24 h, respectively.
Adenovirus construction
Briefly, IQGAP1 was amplified and subcloned into
pAdTrack-CMV, an adenoviral shuttle plasmid, whereas GFP was used
as a non-specific control. Subsequently, the recombinant shuttle
plasmids pAdTrack-CMV and pAdEasy-1 were homologously recombined in
the Escherichia coli strain BJ5183. The recombinant plasmids
obtained were transfected into HEK293 cells (Cell Bank of Shanghai
Institutes for Biological Sciences, Shanghai, China) to generate
recombinant adenovirus. The virus was amplified and purified, and
titers were determined using a p24 ELISA kit (Cell Biolabs, Inc.,
San Diego, CA, USA) before being stored at −80°C for subsequent
use.
Small-interfering RNA (siRNA)
transfection
Scrambled siRNA and siRNA targeting IQGAP1
(sc-35700) were purchased from Santa Cruz Biotechnology (Santa
Cruz, CA, USA). The cells were transfected with scrambled or IQGAP1
siRNA according to the manufacturer’s instructions. Briefly, IQGAP1
siRNA and scrambled siRNA (30 pmol) were diluted in 500 μl
DMEM and mixed with 5 μl Lipofectamine RNAi MAX (Invitrogen,
Carlsbad, CA, USA). After 15 min of incubation at room temperature,
the complexes were added to the cells to a final volume of 3 ml
medium. The cells were then harvested at the indicated times for
further analysis. The efficiency of the IQGAP1 siRNA was confirmed
by western blot analysis of Flag expression.
Transfections and luciferase assays
VSMCs from varicose veins were transfected with a
reporter construct containing a luciferase gene (ERE2-TK-LUC) after
IQGAP1 expression was knocked down by IQGAP1 siRNA. Subsequently,
10−8 M E2 was added to the culture medium. After these
cells were treated for 24 or 48 h, the luciferase activity was
assayed according to the manufacturer’s instructions using the
luciferase assay system (Promega, Madison, WI, USA).
MTT assay
VSMC growth and proliferation were evaluated by MTT
assay. The experiments were carried out in 96-well plates according
to the manufacturer’s instructions (Roche GmbH, Mannheim, Germany).
For MTT assay, tetrazolium salts were transformed by active enzymes
in the cells into intracellular formazan deposits. The cells were
incubated for 4 h with the tetrazolium salts. After this incubation
time, the purple formazan salts formed became soluble. Absorbance
was determined at 490 nm.
Migration assays
The VSMCs were plated in 6-well plates with DMEM.
Twenty-four hours after plating, the medium was replaced with DMEM
with 0.5% charcoal-stripped fetal bovine growth serum (sBGS) and 18
h after starvation, a single scratch was made using a plastic p200
pipette tip. The cells were washed once with PBS to remove
non-adhered cells, and incubated in DMEM with 0.5% sBGS containing
10−8 M E2 or the vehicle (PBS). At 24 h after
the scratch was made, bright-field images of 4 different positions
for each condition were captured and cell migration was measured as
the number of cells that had entered the scratch. Each experiment
was repeated 4 times.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA was extracted from the VSMCs using TRIzol
RNA-extraction reagent (Gibco, Rockville, MD, USA). Approximately 5
μg of total RNA for each sample was reverse-transcribed into
cDNA for RT-qPCR. Quantitative PCR (qPCR) was performed in a final
volume of 10 μl, containing 5 μl of SsoFast™ EvaGreen
Supermix (Bio-Rad Laboratories, Hercules, CA, USA), 1 μl of
cDNA (1:50 dilution) and 2 μl each of the forward and
reverse primers (1 mM). The steps in qPCR were performed as
follows: 94°C for 2 min for initial denaturation; 94°C for 20 sec,
58°C for 15 sec, and 72°C for 15 sec, and 2 sec used for plate
reading for 40 cycles. A melt curve was generated from 65 to 95°C.
β-actin was used as a quantitative and qualitative control to
normalize the gene expression. All of the primers used in this
experiment are listed in Table
I.
 | Table IList of primers for RT-qPCR. |
Table I
List of primers for RT-qPCR.
| Genes | Sequences |
|---|
| IQGAP1 | Forward:
5′-GTGAAATCATCAACACCCAC-3′ |
| Reverse:
5′-TTCCTGGTATTTGTTCTTTGG-3′ |
| β-actin | Forward:
5′-GCACCACACCTTCTACAATG-3′ |
| Reverse:
5′-TGCTTGCTGATCCACATCTG-3′ |
Western blot analysis
The VSMCs were homogenized and lysed with RIPA lysis
buffer (100 mM NaCl, 50 mM Tris-HCl pH 7.5, 1% Triton X-100, 1 mM
EDTA, 10 mM β-glycerophosphate, 2 mM sodium vanadate and protease
inhibitor). Forty micrograms of protein per lane were separated by
12% SDS-PAGE and electroblotted onto a nitrocellulose membrane
(Thermo Fisher Scientific, Waltham, MA, USA). Target proteins were
probed using primary antibodies including anti-p-Akt (sc-33437),
anti-Akt (sc-5298), anti-p-ERK (sc-16982), anti-ERK (sc-514302),
anti-IQGAP1 (sc-10792; all from Santa Cruz Biotechnology) and
anti-β-actin (612657; BD bioscience, San Jose, CA, USA). The
ECL-plus system (GE Healthcare, Pittsburgh, PA, USA) was used for
detection.
Statistical analysis
The results are expressed as the means ± SD.
Statistical significance was analyzed using one-way factorial ANOVA
or the two-tailed Student’s t-test. A value of P<0.05 was
considered to indicate a statistically significant difference. All
analyses were conducted using SPSS software (SPSS Inc., Chicago,
IL, USA).
Results
Estrogen stimulation differentially
regulates VSMC proliferation and migration
Though compelling evidence has elucidated the role
of estrogen in vascular protection, elevated estrogen levels have
been shown to be associated with vascular remodeling and varicose
veins (25,26). In addition, VSMC proliferation and
migration play important roles in vascular reforming (27). Therefore, we hypothesized that
estrogen may directly influence the physiological function of
VSMCs. In order to validate our hypothesis, VSMCs isolated from
varicose veins, normal veins and human umbilical veins were either
treated or not with 10−8 M E2 for 24 h. At
the end of the treatment, cell proliferation and migration were
detected by MTT assay and scratch wound motility assay,
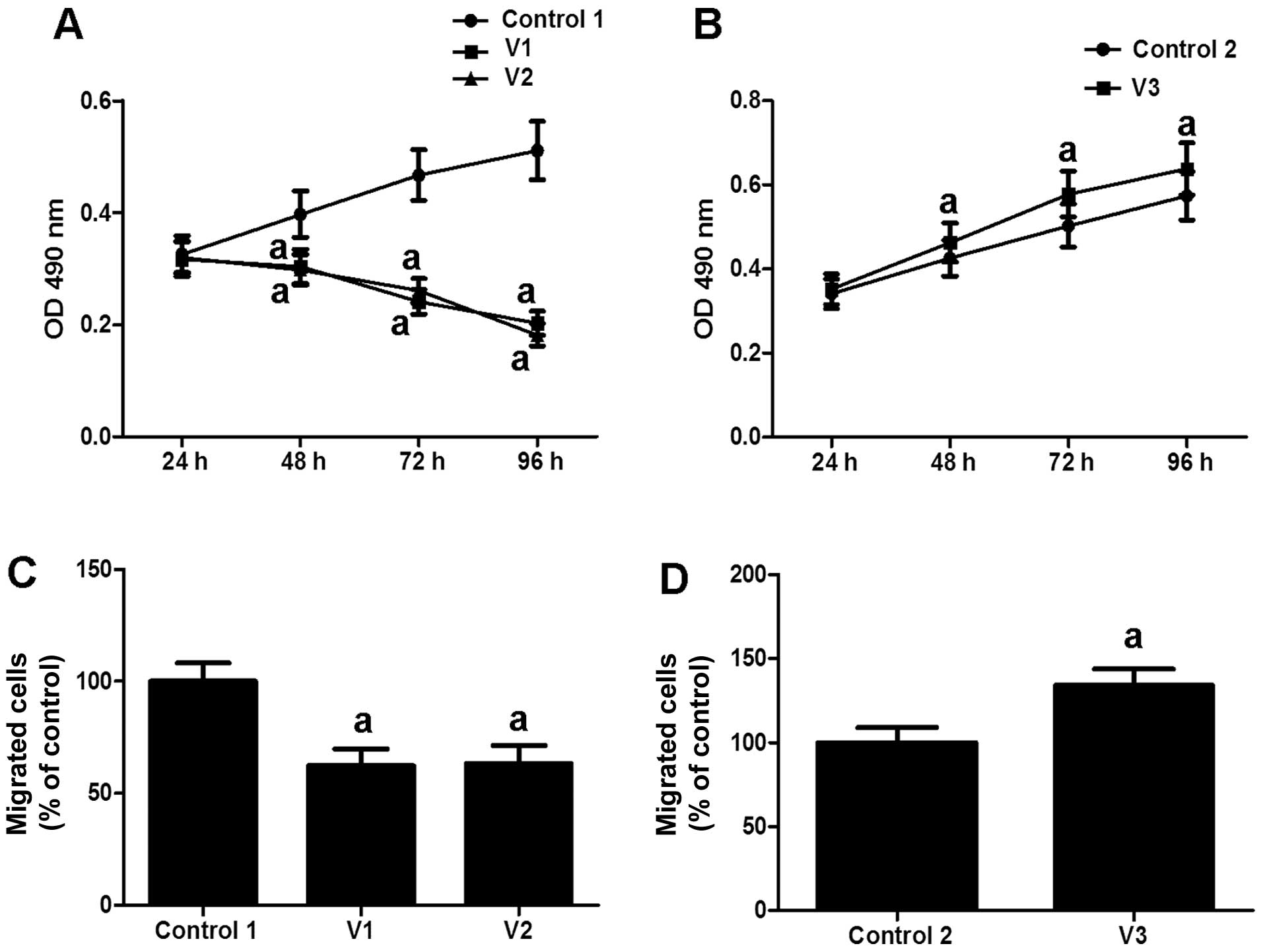
respectively. Compared with the normal controls (Control 1, 0
μM E2), estrogen markedly inhibited the
proliferation and migration rate of the VSMCs obtained from normal
veins (V1) and human umbilical veins (V2) (Fig. 1A and C). By contrast, treatment
with estrogen markedly promoted the proliferation of the VSMCs
obtained from varicose veins (V3 vs. Control 2: lesion VSMCs not
treated with E2), as well as VSMC migration (Fig. 1B and D). These results indicate
that estrogen has a dual effect on VSMC proliferation and
migration.
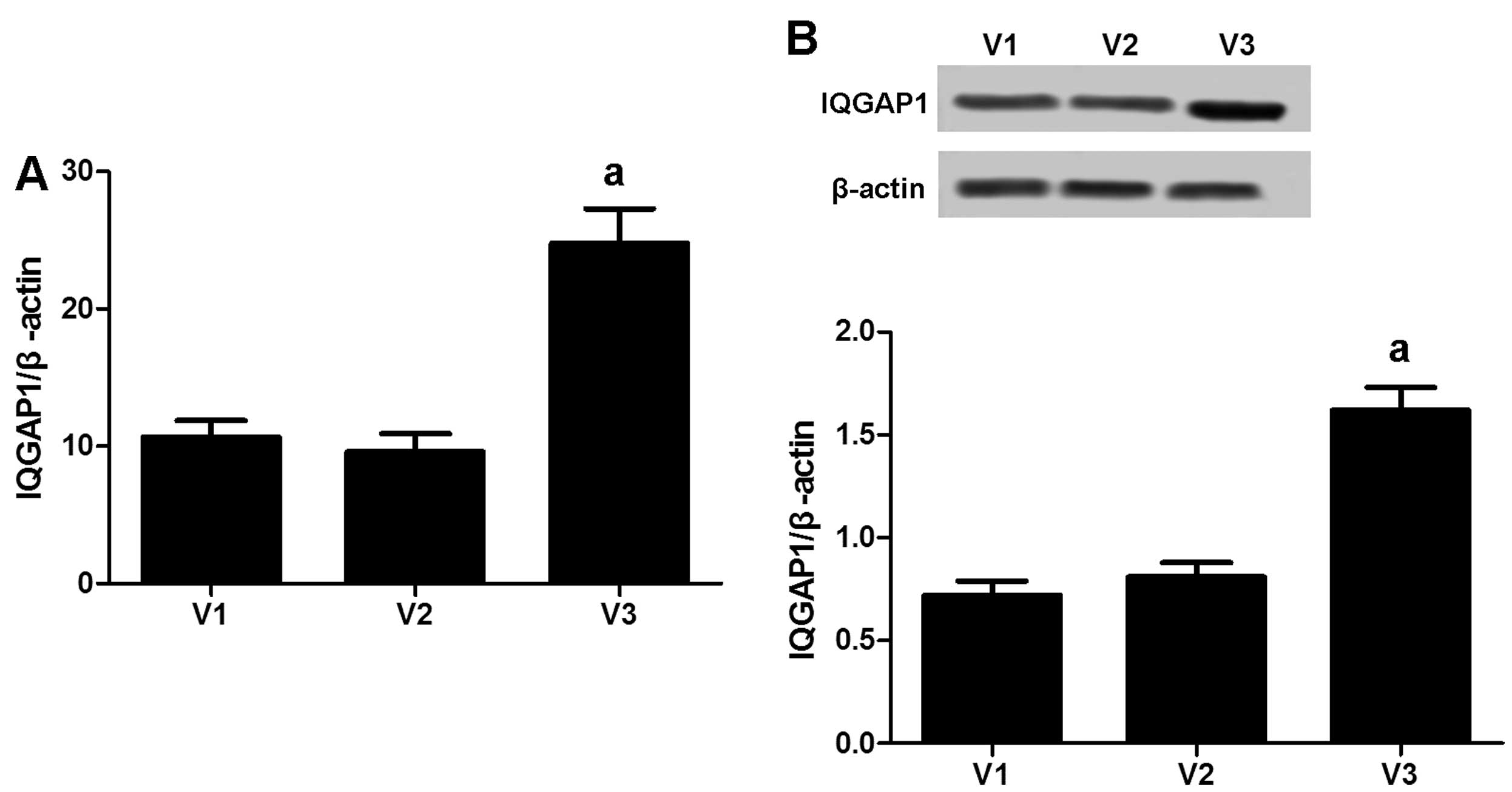
IQGAP1 is overexpressed in VSMCs derived
from varicose veins
IQGAP1 is a scaffold protein that contributes to
cell adhesion, cell migration and proliferation. To verify IQGAP1
expression in VSMCs, RT-qPCR was performed to analyze the mRNA
level of IQGAP1 in the VSMCs obtained from varicose veins (V3, of
11 patients), normal veins (V1, of 9 patients) and human umbilical
veins (V2, of 7 patients). As shown in Fig. 2A, IQGAP1 expression (190 kDa) was
significantly increased in the VSMCs obtained from varicose veins
compared with those obtained from normal veins and human umbilical
veins. These results were further confirmed by western blot
analysis (Fig. 2B).
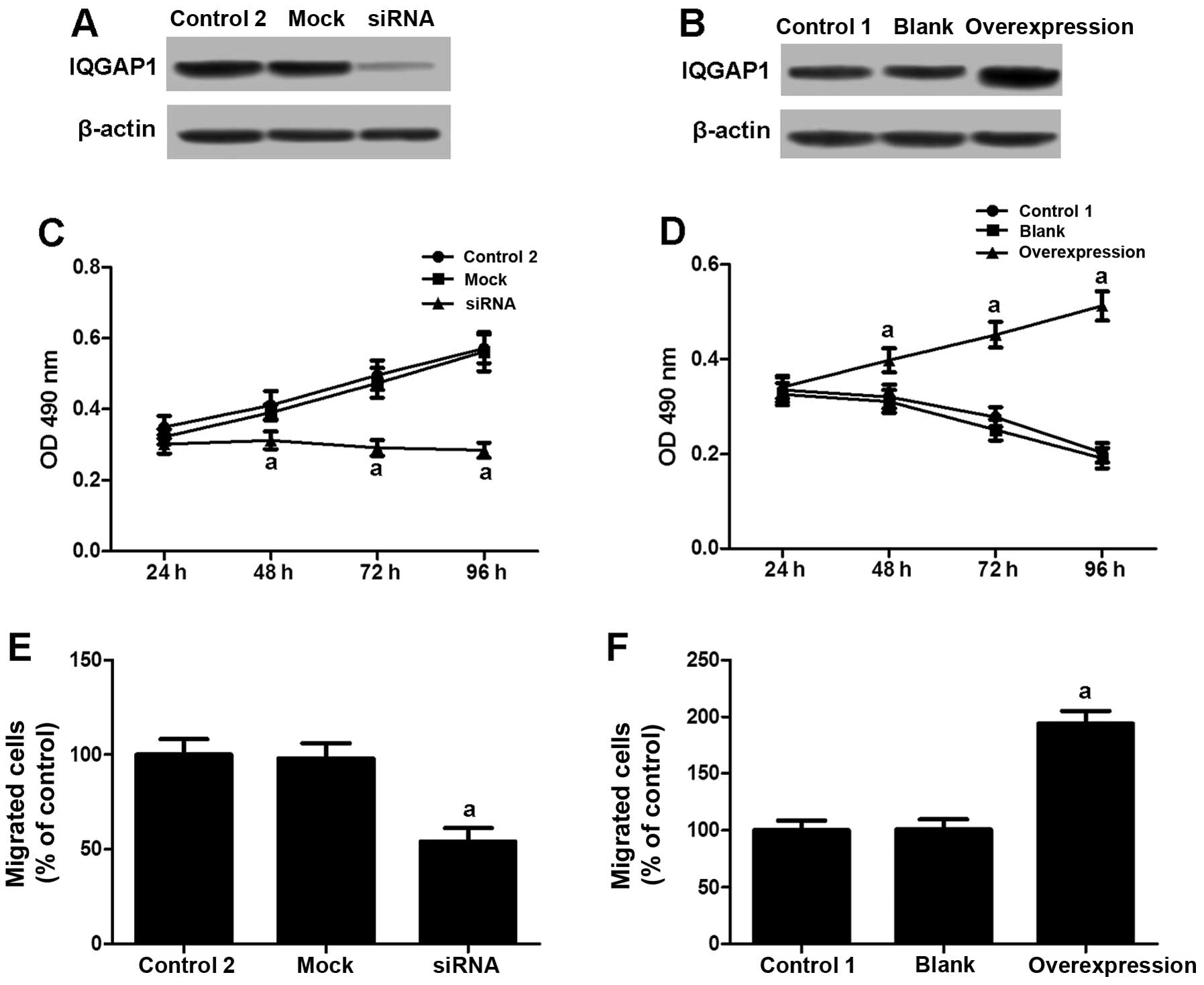
IQGAP1 is involved in the modulation of
VSMC proliferation and migration induced by estrogen
A high IQGAP1 expression was demonstrated to
contribute to cell proliferation and migration. We then
investigated whether IQGAP1 is associated with the regulation of
VSMC proliferation and migration induced by estrogen. The VSMCs
obtained from varicose veins were transfected with IQGAP1 siRNA and
treated with 10−8 M E2 (Fig. 3A). Compared with the controls
(lesion VSMCs not treated with E2; Control 2), the
depletion of IQGAP1 expression markedly reduced VSMC proliferation
(Fig. 3C) and migration (Fig. 3E). Moreover, the VSMCs obtained
from normal veins and treated with E2 (V1) transfected
with an IQGAP1 overexpression vector (Fig. 3B) attained higher proliferation
(Fig. 3D) and migration rates
(Fig. 3F). These results suggest
that IQGAP1 plays a role in the regulation of VSMCs in response to
estrogen stimulation.
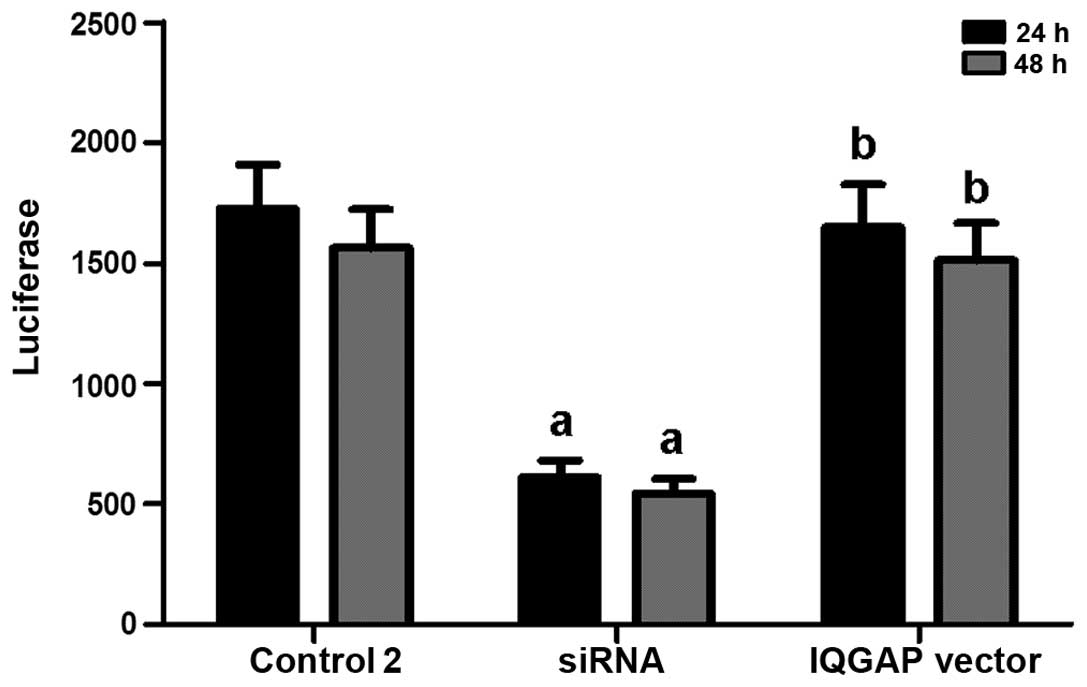
IQGAP1 enhances ERα activity induced by
estrogen
It has been reported that ER is involved in the
promotion of cell growth (24,28). Therefore, we evaluated whether
IQGAP1 exerts its effects by regulating ER transcriptional
activity. Following the knockdown of IQGAP1, the lesion VSMCs were
transiently transfected with a reporter construct containing a
luciferase gene regulated by two estrogen response elements
(ERE2-TK-LUC) to examine the variations in ERα transcriptional
activity. Compared with the controls, cells with a 45% decrease in
IQGAP1 expression levels, expressed ERα with a 65% lower
transcriptional activity (Fig.
4). However, ERα activity in the VSMCs was restored by the
IQGAP1 expression vector (Fig.
4).
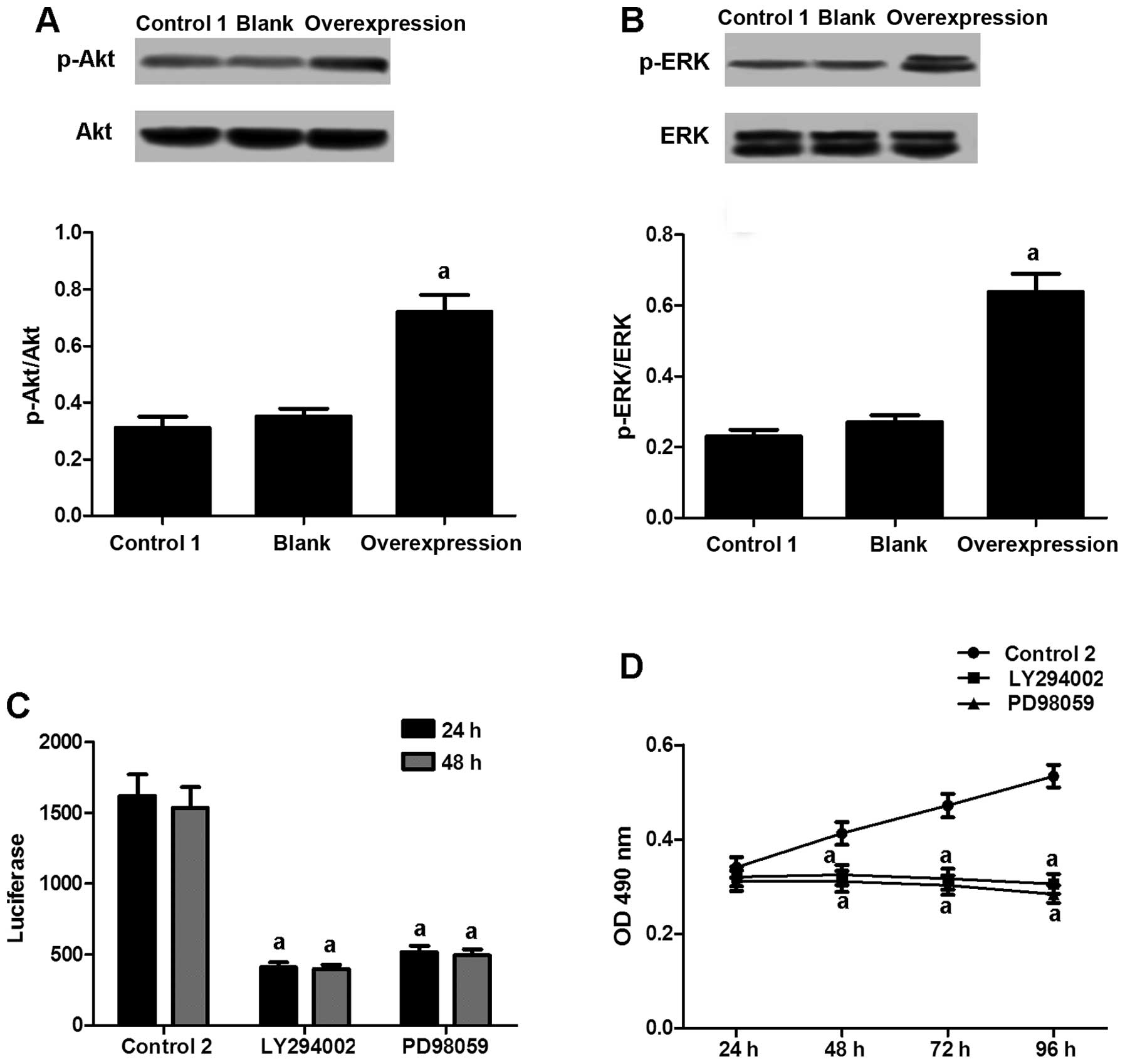
IQGAP1 activates ERα in VSMCs through the
activation of kinases
To examine the mechanisms through which IQGAP1
promotes ERα activity, we assessed the phosphorylation levels of
kinases that promote cell growth, including Akt and ERK. Elevated
phosphorylation levels of Akt (Fig.
5A) and ERK (Fig. 5B) were
observed in the normal VSMCs transfected with the IQGAP1
overexpression vector. However, treatment with the PI3K (Akt
kinase) inhibitor, LY294002, or the ERK inhibitor, PD98059,
distinctly suppressed ERα transcriptional activity (Fig. 5C), as well as the proliferation
rate of the lesion VSMCs (Fig.
5D). Hence, our results provide evidence of the mediation of
ERα activation and VSMC proliferation by kinases.
Discussion
To date, the exact etiology of varicose veins has
not been clarified. However, increasing numbers of risk factors,
such as hormonal changes, obesity, leg injury, prolonged standing,
a highly refined diet and tight undergarments have been reported to
be involved in the development of this disease (8,25,26). Hormonal changes, such as elevated
estrogen levels, which occur during pregnancy, have been reported
to play a major role in the development of varicose veins (27). Alterations in hormonal levels
inducing hypertrophy and the growth of the SMC layer in varicose
veins have been suggested in different studies (28–31). However, augmented estrogen levels
have also been reported to reduce SMC migration and proliferation
(17,32,33). These dual or opposite effects of
estrogen have also been demonstrated in other cell types (34); however, the mechanisms invovled
have not yet been clearly defined. Notably, in the present study we
found that the proliferation and migration of the VSMCs obtained
from normal veins were inhibited by treatment with E2,
while those of the lesion VSMCs were stimulated. These results
indicate that estrogen may have dual effects on VSMC proliferation
and migration. This effect has also been noted by Zhang et
al (24), who claimed that
the growth effect induced by estrogen on VSMCs was largely
dependent on the estrogen concentration. Additionally, Song et
al (35) deduced that the
growth-regulating effect of estrogen on VSMCs was dependent on the
phenotypic state of the cells.
In the present study, IQGAP1 was shown to be
overexpressed in VSMCs obtained from varicose veins compared with
those obtained from normal veins or human umbilical veins.
Accumulating evidence has identified the role of IQGAP1 in the
modulation of cell proliferation and migration. Mataraza et
al (36) demonstrated that
the overexpression of IQGAP1 in mammalian cells promoted cell
migration in a Cdc42- and Rac1-dependent manner. Furthermore,
IQGAP1, through interaction with platelet-derived growth factor
receptor (PDGFR) and focal adhesion (FA) signaling proteins, has
been shown to promote the activation of PDGFR in FAs, as well as FA
formation, which results in VSMC migration and neointimal formation
following injury (37). Notably,
the present study demonstrated that the high expression of IQGAP1
overcame the inhibitory effects on VSMC proliferation and migration
induced by E2 in VSMCs derived from normal veins.
We also observed that ERα transcriptional activity
was decreased following the knockdown of IQGAP1 expression in the
lesion VSMCs, which suggests a regulatory role of IQGAP1 in ERα
activation. Further investigation suggested that ERα activation and
VSMC proliferation may be attributed to a triggering of kinase
activation by IQGAP1. Distinct co-regulatory proteins are recruited
to ERα and regulate ERα function by serving as co-repressors or
co-activators (38). Accumulating
evidence indicates that ERα is associated with interconnected
networks of proteins that maintain the structure and function of
the receptor and influence estrogen responsive gene expression.
Erdemir et al (39)
identified that IQGAP1 may directly interact with the nuclear
receptor ERα and modulate its transcriptional activation of pS2, PR
and cyclin D1.
Collectively, the present study demonstrates the
differential modulation by estrogen of VSMC proliferation and
migration, in which IQGAP1 plays an essential role. The high
expression of IQGAP1 is associated with the increased proliferation
and migration of VSMCs exposed to E2. These effects may
be attributed to a regulation of ERα activity, as well as kinase
activation by IQGAP1. As the induction of VSMC proliferation and
migration is essential for the progression of varicosity, IQGAP1
may thus be an effective target for the treatment of
varicosity.
References
|
1
|
Teng H, Dong J, Li Z and Huang J: Effect
of vascular endothelial cells in the occurrence and development of
lower extremity varicose veins. China Medical Herald. 10:46–49.
2013.
|
|
2
|
Raffetto JD and Khalil RA: Mechanisms of
varicose vein formation: valve dysfunction and wall dilation.
Phlebology. 23:85–98. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ruckley CV: Socioeconomic impact of
chronic venous insufficiency and leg ulcers. Angiology. 48:67–69.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Carpentier PH, Cornu-Thenard A, Uhl JF,
Partsch H and Antignani PL: Appraisal of the information content of
the C classes of CEAP clinical classification of chronic venous
disorders: a multicenter evaluation of 872 patients. J Vasc Surg.
37:827–833. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Langer RD, Ho E, Denenberg JO, Fronek A,
Allison M and Criqui MH: Relationships between symptoms and venous
disease: the San Diego population study. Arch Intern Med.
165:1420–1424. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kurz X, Kahn SR, Abenhaim L, et al:
Chronic venous disorders of the leg: epidemiology, outcomes,
diagnosis and management. Summary of an evidence-based report of
the VEINES task force Venous Insufficiency Epidemiologic and
Economic Studies. Int Angiol. 18:83–102. 1999.PubMed/NCBI
|
|
7
|
Hamdan A: Management of varicose veins and
venous insufficiency. Jama. 308:2612–2621. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee AJ, Evans CJ, Allan PL, Ruckley CV and
Fowkes FG: Lifestyle factors and the risk of varicose veins:
Edinburgh Vein Study. J Clin Epidemiol. 56:171–179. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cornu-Thenard A, Boivin P, Baud JM, De
Vincenzi I and Carpentier PH: Importance of the familial factor in
varicose disease. Clinical study of 134 families. J Dermatol Surg
Oncol. 20:318–326. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
White CD, Erdemir HH and Sacks DB: IQGAP1
and its binding proteins control diverse biological functions. Cell
Signal. 24:826–834. 2012. View Article : Google Scholar :
|
|
11
|
Owen D, Campbell LJ, Littlefield K, et al:
The IQGAP1-Rac1 and IQGAP1-Cdc42 interactions: interfaces differ
between the complexes. J Biol Chem. 283:1692–1704. 2008. View Article : Google Scholar
|
|
12
|
Roy M, Li Z and Sacks DB: IQGAP1 binds
ERK2 and modulates its activity. J Biol Chem. 279:17329–17337.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Roy M, Li Z and Sacks DB: IQGAP1 is a
scaffold for mitogen-activated protein kinase signaling. Mol Cell
Biol. 25:7940–7952. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ren JG, Li Z and Sacks DB: IQGAP1
modulates activation of B-Raf. Proc Natl Acad Sci USA.
104:10465–10469. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meyer RD, Sacks DB and Rahimi N:
IQGAP1-dependent signaling pathway regulates endothelial cell
proliferation and angiogenesis. PLoS One. 3:e38482008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Noritake J, Watanabe T, Sato K, Wang S and
Kaibuchi K: IQGAP1: a key regulator of adhesion and migration. J
Cell Sci. 118:2085–2092. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li QY, Chen L, Zhu YH, Zhang M, Wang YP
and Wang MW: Involvement of estrogen receptor-beta in farrerol
inhibition of rat thoracic aorta vascular smooth muscle cell
proliferation. Acta Pharmacol Sin. 32:433–440. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xiao Y, Huang Z, Yin H, Lin Y and Wang S:
In vitro differences between smooth muscle cells derived from
varicose veins and normal veins. J Vasc Surg. 50:1149–1154. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Renno WM, Saleh F and Wali M: A journey
across the wall of varicose veins: what physicians do not often see
with the naked eye. Med Princ Pract. 15:9–23. 2006. View Article : Google Scholar
|
|
20
|
Cheng Y, Liu X, Yang J, et al:
MicroRNA-145, a novel smooth muscle cell phenotypic marker and
modulator, controls vascular neointimal lesion formation. Circ Res.
105:158–166. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang X, Zhang Y, Hou D, et al:
17beta-estradiol inhibits oleic acid-induced rat VSMC proliferation
and migration by restoring PGC-1alpha expression. Mol Cell
Endocrinol. 315:74–80. 2010. View Article : Google Scholar
|
|
22
|
Branchetti E, Poggio P, Sainger R, et al:
Oxidative stress modulates vascular smooth muscle cell phenotype
via CTGF in thoracic aortic aneurysm. Cardiovasc Res. 100:316–324.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Afroze T, Yang G, Khoshbin A, et al:
Calcium efflux activity of plasma membrane Ca2+ ATPase-4
(PMCA4) mediates cell cycle progression in vascular smooth muscle
cells. J Biol Chem. 289:7221–7231. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang L, Zhu C, Zhang X, Wan Y and Song J:
Dual effects of estrogen on vascular smooth muscle cells:
receptor-mediated proliferative vs. metabolite-induced
pro-senescent actions. Steroids. 76:309–316. 2011. View Article : Google Scholar
|
|
25
|
Hobbs JT: Letter: varicose veins in
developing countries. Lancet. 2:2591976. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ahti TM, Makivaara LA, Luukkaala T, Hakama
M and Laurikka JO: Lifestyle factors and varicose veins: does
cross-sectional design result in underestimate of the risk?
Phlebology. 25:201–206. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ciardullo AV, Panico S, Bellati C, et al:
High endogenous estradiol is associated with increased venous
distensibility and clinical evidence of varicose veins in
menopausal women. J Vasc Surg. 32:544–549. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wali MA and Eid RA: Changes of elastic and
collagen fibers in varicose veins. Int Angiol. 21:337–343.
2002.
|
|
29
|
Bastos AN, Alves MM, Monte-Alto-Costa A,
et al: α-smooth muscle actin, fibrillin-1, apoptosis and
proliferation detection in primary varicose lower limb veins of
women. Int Angiol. 30:262–271. 2011.PubMed/NCBI
|
|
30
|
Han HC: Twisted blood vessels: symptoms,
etiology and biomechanical mechanisms. J Vasc Res. 49:185–197.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kowalewski R, Malkowski A, Sobolewski K
and Gacko M: Evaluation of transforming growth factor-beta
signaling pathway in the wall of normal and varicose veins.
Pathobiology. 77:1–6. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ueda K, Lu Q, Baur W, Aronovitz MJ and
Karas RH: Rapid estrogen receptor signaling mediates
estrogen-induced inhibition of vascular smooth muscle cell
proliferation. Arterioscler Thromb Vasc Biol. 33:1837–1843. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao J, Imbrie GA, Baur WE, et al:
Estrogen receptor-mediated regulation of microRNA inhibits
proliferation of vascular smooth muscle cells. Arterioscler Thromb
Vasc Biol. 33:257–265. 2013. View Article : Google Scholar :
|
|
34
|
Lewis-Wambi JS and Jordan VC: Estrogen
regulation of apoptosis: how can one hormone stimulate and inhibit?
Breast Cancer Res. 11:2062009. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Song J, Wan Y, Rolfe BE, Campbell JH and
Campbell GR: Effect of estrogen on vascular smooth muscle cells is
dependent upon cellular phenotype. Atherosclerosis. 140:97–104.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mataraza JM, Briggs MW, Li Z, Entwistle A,
Ridley AJ and Sacks DB: IQGAP1 promotes cell motility and invasion.
J Biol Chem. 278:41237–41245. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kohno T, Urao N, Ashino T, et al: IQGAP1
links PDGF receptor-beta signal to focal adhesions involved in
vascular smooth muscle cell migration: role in neointimal formation
after vascular injury. Am J Physiol Cell Physiol. 305:C591–C600.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Heldring N, Pike A, Andersson S, et al:
Estrogen receptors: how do they signal and what are their targets.
Physiol Rev. 87:905–931. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Erdemir HH, Li Z and Sacks DB: IQGAP1
binds to estrogen receptor-alpha and modulates its function. J Biol
Chem. 289:9100–9112. 2014. View Article : Google Scholar : PubMed/NCBI
|