1. Introduction
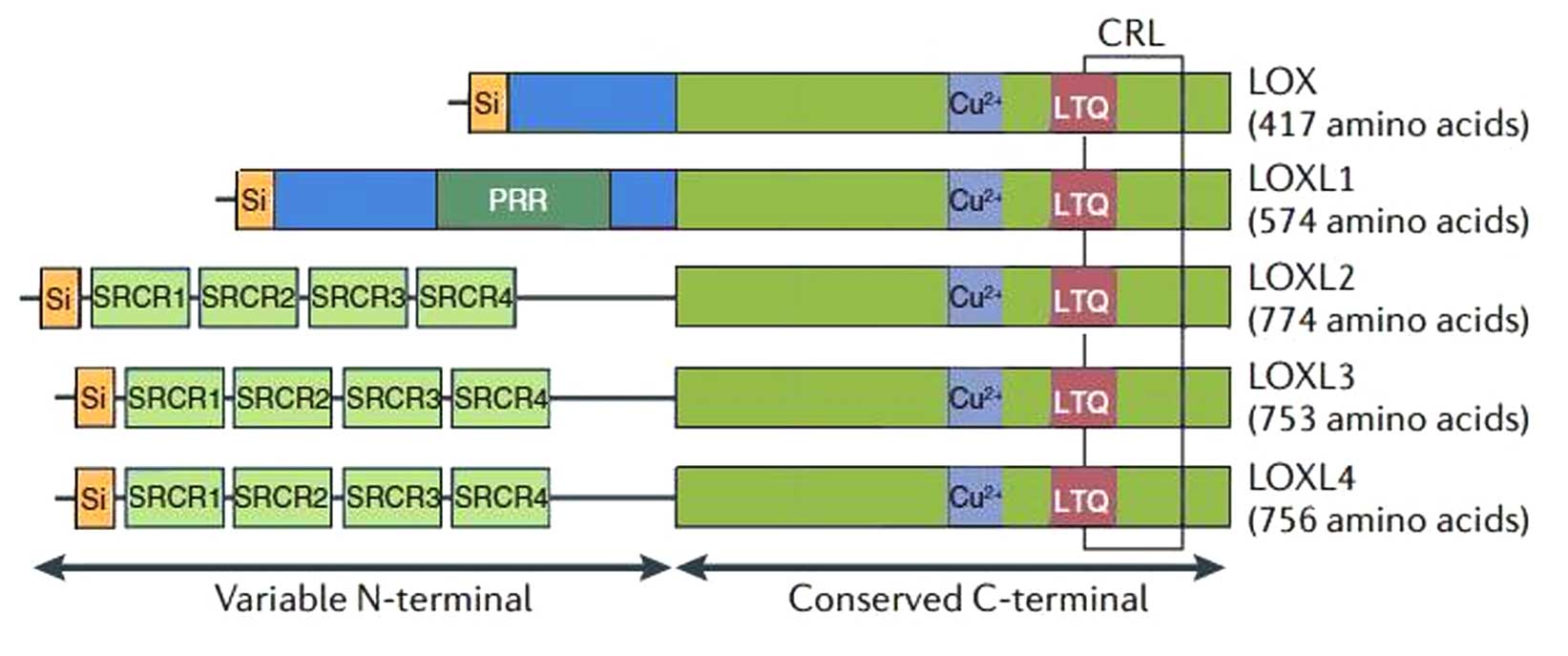
LOX-like (LOXL)2 is a member of the lysyl oxidase
(LOX) family, which is comprised of five members, the prototypical
LOX and four related members, LOXL1-4. It is a secreted
copper-dependent amine oxidase, and its main role is to catalyze
the covalent cross-link of collagen and elastin in the
extracellular matrix (ECM). This occurs through the oxidative
deamination of peptidyl lysine residues in ECM components (1,2).
Each member of the LOX family has a highly conserved carboxyl
(C)-terminal domain that contains a copper-binding motif, lysine
tyrosylquinone (LTQ) residues and a cytokine receptor-like (CRL)
domain, which is essential for catalytic activity (3). The amino-terminal regions are
different and are thought to be important in protein-protein
interactions. The prodomains in LOX and LOXL1 enable their
secretion as inactive proenzymes, which are then activated
extracellularly. However, LOXL2, LOXL3 and LOXL4 contain four
scavenger receptor cysteine-rich (SRCR) domains instead of a
pro-sequence, and SRCR domains are thought to be involved in
protein-protein interactions at the N-terminal region (4), and in the ligand binding of both
soluble proteins and membrane-bound protein receptors (5,6).
Due to similarities in the domain arrangement, LOX and LOXL1
represent one LOX subfamily, whereas LOXL2-4 constitute another LOX
subfamily (Fig. 1).
In addition, the presence of various protein domains
in LOX family members suggests their involvement in multiple
biological functions beyond ECM stabilization. Although all members
are involved in different stages of tumor progression (7), and have a high sequence homology,
the precise molecular functions of LOXL1, LOXL3, LOXL4 are not yet
fully understood. Previous studies on LOXL2 in tumors have,
however, made some progress: LOXL2 is currently thought to be
involved in developmental regulation, cell adhesion, senescence,
cell migration and invasion, metastasis, epithelial-mesenchymal
transition (EMT) and malignant transformation (4,7,8).
In this review, we summarize the broader areas of the progress made
in relation to LOXL2, particularly its functions and mechanisms of
action in tumors.
2. Functions of LOXL2 in cancer
LOXL2 may be a tumor suppressor
The human LOXL2 gene is located on chromosome
8p21-22, and is initially synthesized as an 87-kDa proenzyme of 774
amino acids. In 1997, the LOXL2 gene was originally identified as a
downregulated transcript in various non-adherent tumor cell lines
when compared to adherent tumor cell lines (9). Subsequently, several studies have
also identified LOXL2 as a downregulated transcript in cell lines
and tumors. Investigations into the function of LOXL2 have
indicated that the gene is a candidate for tumor suppression
(7). There are several reports
that support a role for LOXL2 as a tumor suppressor. A nearly
60-fold decrease in LOXL2 mRNA expression was found to have
occurred in RAS-transformed rat fibroblasts (10). The study by Rost et al
(11) demonstrated that the
downregulation of LOXL2 mRNA expression was observed in several
cell lines and tissue samples of head and neck squamous cell
carcinoma (HNSCC). Via a large-scale serial analysis of gene
expression, downregulated LOXL2 expression was also observed in
ovarian tumors (12,13). Recently, a study on non-small cell
lung cancer (NSCLC) by Zhan et al (14) revealed that, compared with the
surrounding normal tissues, LOXL2 mRNA and protein expression
levels were downregulated in tumor samples from patients with
NSCLC. However, the upregulation LOXL2 of expression has been
reported in breast, colon, esophageal and pancreatic carcinoma cell
lines, and gastric cancer (15,16–18). Despite being classed as a secreted
protein, LOXL2 has been shown to have both intracellular and
extracellular functions, and can induce EMT. These seemingly
opposing reports suggest that LOXL2 has complex and paradoxical
roles and may function both as a tumor suppressor and promoter of
metastasis, depending on its localization and tumor type, and also
has the ability to induce EMT (19).
LOXL2 promotes invasion and metastasis in
cancer
In contrast to the above-mentioned findings, it has
been reported by several groups that the mRNA and protein levels of
LOXL2 are increased in invasive or metastatic breast, pancreatic
and liver carcinoma cell lines (17,20,21). LOXL2 has been characterized as a
promoter of tumor cell invasiveness (22). Moreover, in immunocompetent or
immunocompromised orthotopic and transgenic breast cancer models,
the genetic, chemical or antibody-mediated inhibition of LOXL2 has
been shown to result in decreased metastasis (15). These studies and the emerging
evidence suggests that in these situations, LOXL2 may function as a
promoter of invasion and metastasis rather than a tumor
suppressor.
In 2009, Brekhman and Neufeld (23), using a novel asymmetric 3D
in-vitro invasion assay and a quantitative analysis of the
directional invasive behavior of tumor cells in a 3D environment
mimicking the tumor microenvironment, found that the inhibition of
endogenous LOXL2 expression in several types of tumor cells
strongly inhibited their invasiveness, and further demonstrated
that LOXL2 was a potent inducer of invasiveness. In 2013, a study
by Ahn et al (20), using
an immunohistochemical study of tissue specimens from 309 patients
with breast cancer and reverse transcription polymerase chain
reaction (RT-PCR) analysis of human breast cancer cell lines,
confirmed that a higher LOXL2 expression was associated with the
invasiveness of basal-like breast cancer cells and lower survival
rates of breast cancer patients. Subsequently, in 2014, Wong et
al (21), integrating the
clinical relevance, molecular regulation and functional
implications, demonstrated that LOXL2 promoted intrahepatic
metastasis and extrahepatic metastasis. The relevant mechanisms are
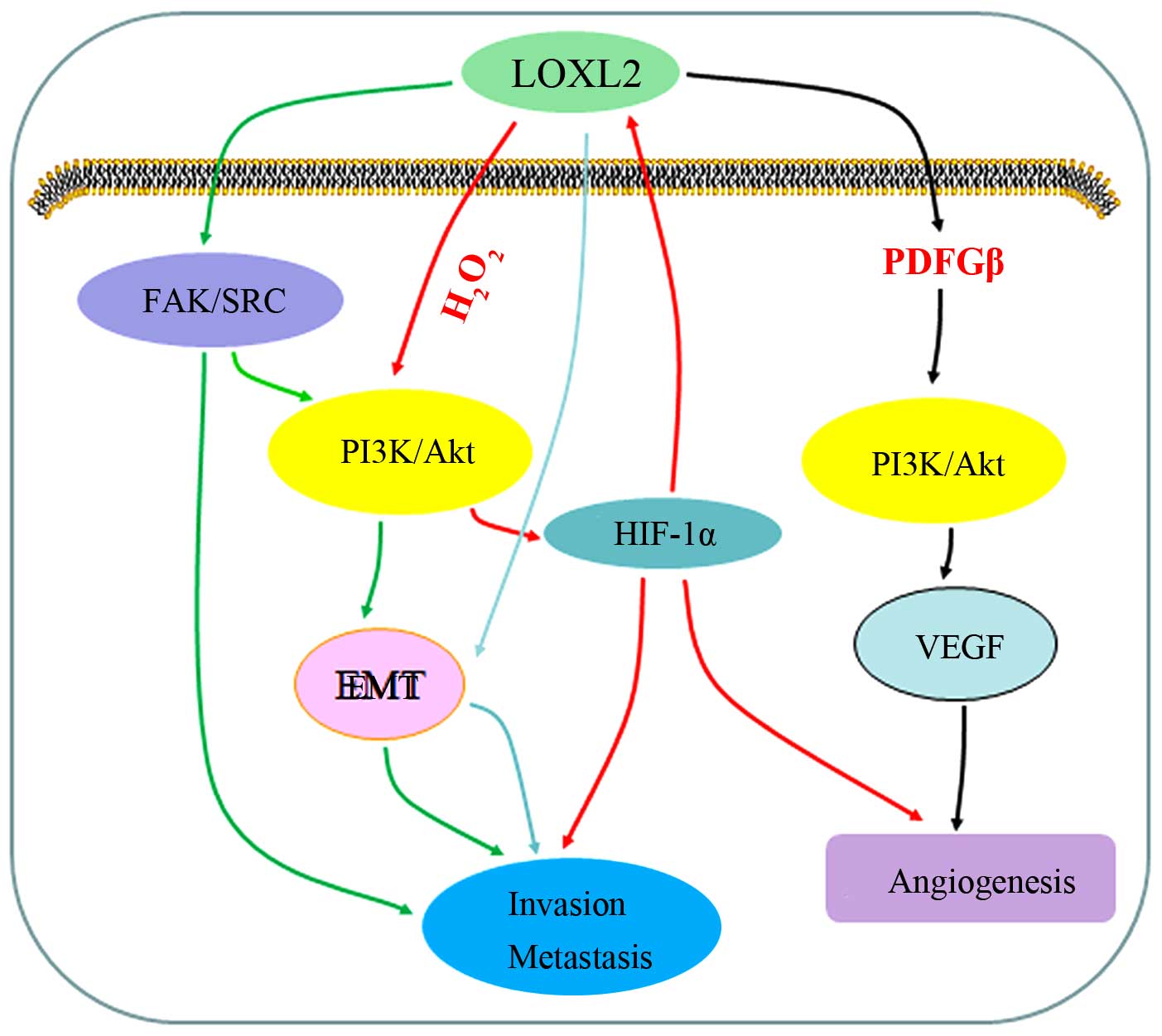
discussed below and are summarized in Fig. 2.
LOXL2 promotes cancer-associated
fibrosis
Fibrosis may play a role in tumor progression by
promoting cell transformation and metastasis directly (24). Several studies on the
extracellular functions of the LOX family in physiological and
pathological processes have found that the LOX family plays a
critical role in organs containing fibrillar collagen and/or
elastic fibers, such as the skin, lungs, the cardiovascular system,
cartilage, and the fibrous lamina propria in the small intestines
and stomach (25–27). For example, when LOXL1 is knocked
down, normal elastic fibers are not deposited in the uterine tract
postpartum, and mice lacking LOXL1 develop pelvic organ prolapse,
enlarged air spaces in the lungs, loose skin and vascular
abnormalities with concomitant tropoelastin accumulation (27). In clinical practice, a reduced LOX
activity owing to the diminished availability of copper, was found
in two X-linked recessively inherent disorders, Menkes disease and
occipital horn syndrome (28).
The above-mentioned findings indicate that LOXL2 promotes fibrosis.
Indeed, accumulating copper upregulates the expression of LOXL2 in
Wilson's disease, and then leads to liver fibrosis (29). In fibrotic processes, an excessive
deposition of collagen in particular, changes the physical,
biochemical and biomechanical matrix properties, which eventually
leads to defective organ function and organ failure (30). Moreover, targeting LOXL2 with an
inhibitory monoclonal antibody (AB0023) has been shown to be
efficacious in both primary and metastatic xenograft models of
cancer, as well as in models of liver and lung fibrosis (31).
LOXL2 is a prognostic marker for
cancer
LOXL2 was, for the first time, described as a
prognostic marker for laryngeal squamous cell carcinomas (32). Peinado et al (32) examined the protein expression of
LOXL2 by immunohistochemistry in a cohort of 256 human laryngeal
squamous cell carcinomas, and found that increased expression of
LOXL2 correlated with local recurrence, and moreover, that high
levels of LOXL2 were associated with decreased overall and
disease-free survival in patients with laryngeal squamous cell
carcinomas, lung squamous cell carcinoma and lymph node-negative
(N0) breast adenocarcinomas. Subsequently, several studies
demonstrated that the high expression or overexpression of LOXL2
correlated with a poor prognosis and decreased survival in gastric
cancer (18), colon tumor
(16,33), basal-like breast carcinoma
(34), pancreatic carcinoma
(17) and hepatocellular
carcinoma (21).
3. Mechanisms of action of LOXL2 in
cancer
LOXL2 promotes the activation of focal
adhesion kinase (FAK)/Src in cancer
The involvement of LOXL2 in tumor cell invasion and
metastasis provides a powerful intervention tool for the prevention
of tumor cell dissemination. A previous study on the underlying
mechanisms through which LOXL2 contributes to tumor progression
demonstrated that LOXL2 expression promotes the activation of
associated Src kinase and FAK, and leads to the activation of
associated downstream signaling cascades (18). Indeed, studies have suggested that
Src and FAK are activated via LOX-mediated increased collagen
crosslinking, resulting in increased matrix stiffness, which is
affected by transmembrane integrins that then stimulate the
downstream activation of FAK and Src (35,36). In addition, the activation of Src
and FAK increases tumor cell proliferation and invasion and is
related to the formation of distant metastasis (18,35,36). Moreover, using anti-LOXL2 antibody
has been shown to suppress the activity of FAK and Src, and to
prevent gastric cancer metastases, thus confirming the mechanisms
of action of LOXL2 (18).
LOXL2 increases the expression of
vascular endothelial growth factor (VEGF) in cancer
Angiogenesis contributes to primary and metastatic
cancer growth, and it is necessary for tumor progression (37,38). The VEGF signaling pathway plays a
key role in promoting angiogenesis, not only under normal
conditions, but also in tumor pathogenesis. Thus, it has become a
major intervention tool in targeted cancer therapy (39,40). A study on the effects of LOX
expression on tumor-driven angiogenesis, demonstrated that the
regulation of VEGF mRNA and protein expression is carried out
through the platelet-derived growth factor β (PDGFRβ)-mediated
activation of protein kinase B (Akt) (41). In clinical practice, the use of
angiogenesis inhibitors (bevacizumab and sunitinib) or PDGFβ- or
Akt-mediated signaling inhibitors, has been shwon to prevent the
LOX-mediated increase in VEGF mRNA and protein expression, and to
abrogate the LOX-mediated stimulation of endothelial cells both
in vivo and in vitro (41). Moreover, a recent report suggested
that the inhibition of LOXL2 activity inhibits angiogenesis in part
by affecting VEGF signaling in endothelial cells (42).
LOXL2 and hypoxia-inducible factor-1α
(HIF-1α) reinforce each other in cancer
Several human cancers contain hypoxic regions. Tumor
cells adapt to hypoxic environments better than normal cells
(43). Under hypoxic conditions,
the hypoxia transcription factor, HIF-1α, binds to the hypoxia
responsive element (HRE) in the promoter region of numerous target
genes, including LOX and LOXL2 (44,45). A previous study using chromatin
immunoprecipitation (ChIP) with rabbit polyclonal antiserum to
HIF-1α suggested that LOXL2 is also significantly affected by
HIF-1α (46). LOX overexpression
increases HIF-1α expression under hypoxic conditions; the
LOX-induced increase in HIF-1α expression means they reinforce each
other to promote tumor progression (47); tumor cells have greater
proliferative, invasive and metastatic abilities and express higher
levels of LOX or LOXL2 (23,46). The tumor hypoxic environment can
also drive LOXL2 to be involved in angiogenesis. During
angiogenesis, LOXL2 contributes to the reorganization of collagen
and fibronectin associated with ECM remodeling by HIF-1α (48).
LOXL2 is associated with
phosphatidylinositol 3-kinase (PI3K)/Akt in cancer
A previous study (47) suggested a mechanism of LOX, namely
that it regulates H2O2 production, a
by-product of the enzymatic reaction, and activates the signaling
pathway of PI3K/Akt, then increases HIF-1α protein synthesis; thus
LOX production further constitutes a positive regulation loop and
promotes tumor proliferation. In a breast cancer model,
LOX-mediated H2O2 production has also been
shown to participate in the activation of Src and FAK (49). Indeed, FAK and Src are upstream
signaling cascade components of PI3K (50). LOXL2 can also produce
H2O2 on the regeneration of catalytic
activity and the presence of H2O2 can promote
Src and FAK activation (18).
LOXL2 drives EMT in cancer
EMT, an essential process in development, is
described as a key event in tumor progression, facilitating the
dissociation of cancer cells from the primary tumors concomitant to
the acquisition of cell migration that provides tumor cells with
the ability to invade into the adjacent tissues (51). Moreover, EMT is considered to be
relevant for other stages of the metastatic cascade, such as
intravasation or extravasation (52). EMT is currently considered as a
focal and transient event occurring at specific tumor regions and
being perhaps more relevant to specific tumor types (51–53). Indeed, LOXL2 is originally
described as affirmatively promoting EMT inducer Snail1 stability
and functional activity, driving EMT and tumor progression through
the downregulation of E-cadherin expression at the transcriptional
level (54). A previous study
reported that LOXL2 also mediates EMT through a Snail1-independent
mechanism, related to the downregulation of epidermal
differentiation and cell polarity components (34). Recently, Cuevas et al
provided evidence that catalytically inactive LOXL2 mutants, one of
them unable to be secreted, induce and sustain a full EMT process,
indicating that intracellular LOXL2 drives EMT independent of its
enzymatic activity (55). All the
above-mentioned data support the hypothesis that LOXL2 induces
EMT.
Of note, previous studies have demonstrated that
some factors trigger EMT by activating the PI3K/Akt signaling
pathway (56–58), and thus, LOXL2 can also possibly
induce EMT through PI3K/Akt. Thereby, LOXL2 promotes tumor
progression possibly by activating multiple signal pathways through
a variety of mechanisms, both biochemical and biome-chanical. Thus,
we propose that the mechanisms of action of LOXL2 are applicable to
many common cancer models (Fig.
2), meaning that solid tumors have a high commonality.
4. Future perspectives
A malignant tumor is frequently diagnosed at an
advanced stage, as a result of tumor cell invasion and metastases,
and is usually indicative of a poor prognosis for patients. The
diagnosis of advanced stage of carcinoma is a devastating
experience for both patients and their families. The important role
of LOXL2 in a number of common tumors is unquestionable. Its
effects on cell invasion, metastasis, and the induction of
angiogenesis and malignant transformation have increased its
viability as a possible target for therapeutic intervention. For
instance, a study group from California demonstrated that a
monoclonal antibod (AB0023) against LOXL2 was efficacious in both
primary and metastatic xenograft models (31). Moreover, AB0024, which is a novel
humanized LOXL2 antibody, has already entered phase I clinical
trials in patients with solid tumors (ClinicalTrials.gov.) (59). Even though some molecular-targeted
drugs have been developed, their effects have not yet reached our
expectations. Therefore, more comprehensive studies are warranted
to broaden our understanding of the mechanisms through which LOXL2
modulates the cancer microenvironment and exerts its promoting
effects on cancer progression and metastasis, and to develop novel
anticancer therapeutics. Although the future is full of challenges,
the development of inhibitors targeting LOXL2 brings us hope that
effective treatments for primary and metastatic tumors will be
feasible in the near future.
References
|
1
|
Kagan HM and Trackman PC: Properties and
function of lysyl oxidase. Am J Respir Cell Mol Biol. 5:206–210.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Smith-Mungo LI and Kagan HM: Lysyl
oxidase: properties, regulation and multiple functions in biology.
Matrix Biol. 16:387–398. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lopez KM and Greenaway FT: Identification
of the copper-binding ligands of lysyl oxidase. J Neural Transm.
118:1101–1109. 2011. View Article : Google Scholar
|
|
4
|
Lucero HA and Kagan HM: Lysyl oxidase: an
oxidative enzyme and effector of cell function. Cell Mol Life Sci.
63:2304–2316. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hohenester E, Sasaki T and Timpl R:
Crystal structure of a scavenger receptor cysteine-rich domain
sheds light on an ancient superfamily. Nat Struct Biol. 6:228–232.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martínez VG, Moestrup SK, Holmskov U,
Mollenhauer J and Lozano F: The conserved scavenger receptor
cysteine-rich super-family in therapy and diagnosis. Pharmacol Rev.
63:967–1000. 2011. View Article : Google Scholar
|
|
7
|
Payne SL, Hendrix MJ and Kirschmann DA:
Paradoxical roles for lysyl oxidases in cancer - a prospect. J Cell
Biochem. 101:1338–1354. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Barker HE, Cox TR and Erler JT: The
rationale for targeting the LOX family in cancer. Nat Rev Cancer.
12:540–552. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Saito H, Papaconstantinou J, Sato H and
Goldstein S: Regulation of a novel gene encoding a lysyl
oxidase-related protein in cellular adhesion and senescence. J Biol
Chem. 272:8157–8160. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zuber J, Tchernitsa OI, Hinzmann B,
Schmitz AC, Grips M, Hellriegel M, Sers C, Rosenthal A and Schäfer
R: A genome-wide survey of RAS transformation targets. Nat Genet.
24:144–152. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rost T, Pyritz V, Rathcke IO, Görögh T,
Dünne AA and Werner JA: Reduction of LOX- and LOXL2-mRNA expression
in head and neck squamous cell carcinomas. Anticancer Res. 23(2B):
1565–1573. 2003.PubMed/NCBI
|
|
12
|
Hough CD, Sherman-Baust CA, Pizer ES,
Montz FJ, Im DD, Rosenshein NB, Cho KR, Riggins GJ and Morin PJ:
Large-scale serial analysis of gene expression reveals genes
differentially expressed in ovarian cancer. Cancer Res.
60:6281–6287. 2000.PubMed/NCBI
|
|
13
|
Ono K, Tanaka T, Tsunoda T, Kitahara O,
Kihara C, Okamoto A, Ochiai K, Takagi T and Nakamura Y:
Identification by cDNA microarray of genes involved in ovarian
carcinogenesis. Cancer Res. 60:5007–5011. 2000.PubMed/NCBI
|
|
14
|
Zhan P, Shen XK, Qian Q, Zhu JP, Zhang Y,
Xie HY, Xu CH, Hao KK, Hu W, Xia N, et al: Down-regulation of lysyl
oxidase-like 2 (LOXL2) is associated with disease progression in
lung adenocarcinomas. Med Oncol. 29:648–655. 2012. View Article : Google Scholar
|
|
15
|
Barker HE, Chang J, Cox TR, Lang G, Bird
D, Nicolau M, Evans HR, Gartland A and Erler JT: LOXL2-mediated
matrix remodeling in metastasis and mammary gland involution.
Cancer Res. 71:1561–1572. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fong SF, Dietzsch E, Fong KS, Hollosi P,
Asuncion L, He Q, Parker MI and Csiszar K: Lysyl oxidase-like 2
expression is increased in colon and esophageal tumors and
associated with less differentiated colon tumors. Genes Chromosomes
Cancer. 46:644–655. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rückert F, Joensson P, Saeger HD,
Grützmann R and Pilarsky C: Functional analysis of LOXL2 in
pancreatic carcinoma. Int J Colorectal Dis. 25:303–311. 2010.
View Article : Google Scholar
|
|
18
|
Peng L, Ran YL, Hu H, Yu L, Liu Q, Zhou Z,
Sun YM, Sun LC, Pan J, Sun LX, et al: Secreted LOXL2 is a novel
therapeutic target that promotes gastric cancer metastasis via the
Src/FAK pathway. Carcinogenesis. 30:1660–1669. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cano A, Santamaría PG and Moreno-Bueno G:
LOXL2 in epithelial cell plasticity and tumor progression. Future
Oncol. 8:1095–1108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ahn SG, Dong SM, Oshima A, Kim WH, Lee HM,
Lee SA, Kwon SH, Lee JH, Lee JM, Jeong J, et al: LOXL2 expression
is associated with invasiveness and negatively influences survival
in breast cancer patients. Breast Cancer Res Treat. 141:89–99.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wong CC, Tse AP, Huang YP, Zhu YT, Chiu
DK, Lai RK, Au SL, Kai AK, Lee JM, Wei LL, et al: Lysyl
oxidase-like 2 is critical to tumor microenvironment and metastatic
niche formation in hepatocellular carcinoma. Hepatology.
60:1645–1658. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Akiri G, Sabo E, Dafni H, Vadasz Z,
Kartvelishvily Y, Gan N, Kessler O, Cohen T, Resnick M, Neeman M
and Neufeld G: Lysyl oxidase-related protein-1 promotes tumor
fibrosis and tumor progression in vivo. Cancer Res. 63:1657–1666.
2003.PubMed/NCBI
|
|
23
|
Brekhman V and Neufeld G: A novel
asymmetric 3D in-vitro assay for the study of tumor cell invasion.
BMC Cancer. 9:4152009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cox TR and Erler JT: Remodeling and
homeostasis of the extracellular matrix: Implications for fibrotic
diseases and cancer. Dis Model Mech. 4:165–178. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mäki JM, Räsänen J, Tikkanen H, Sormunen
R, Makikallio K, Kivirikko KI and Soininen R: Inactivation of the
lysyl oxidase gene Lox leads to aortic aneurysms, cardiovascular
dysfunction, and perinatal death in mice. Circulation.
106:2503–2509. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mäki JM, Sormunen R, Lippo S,
Kaarteenaho-Wiik R, Soininen R and Myllyharju J: Lysyl oxidase is
essential for normal development and function of the respiratory
system and for the integrity of elastic and collagen fibers in
various tissues. Am J Pathol. 167:927–936. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu X, Zhao Y, Gao J, Pawlyk B, Starcher
B, Spencer JA, Yanagisawa H, Zuo J and Li T: Elastic fiber
homeostasis requires lysyl oxidase-like 1 protein. Nat Genet.
36:178–182. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mäki JM: Lysyl oxidases in mammalian
development and certain pathological conditions. Histol
Histopathol. 24:651–660. 2009.PubMed/NCBI
|
|
29
|
Vadasz Z, Kessler O, Akiri G,
Gengrinovitch S, Kagan HM, Baruch Y, Izhak OB and Neufeld G:
Abnormal deposition of collagen around hepatocytes in Wilson's
disease is associated with hepatocyte specific expression of lysyl
oxidase and lysyl oxidase like protein-2. J Hepatol. 43:499–507.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nieto MA: The ins and outs of the
epithelial to mesenchymal transition in health and disease. Annu
Rev Cell Dev Biol. 27:347–376. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Barry-Hamilton V, Spangler R, Marshall D,
McCauley S, Rodriguez HM, Oyasu M, Mikels A, Vaysberg M, Ghermazien
H, Wai C, et al: Allosteric inhibition of lysyl oxidase-like-2
impedes the development of a pathologic microenvironment. Nat Med.
16:1009–1017. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Peinado H, Moreno-Bueno G, Hardisson D,
Pérez-Gómez E, Santos V, Mendiola M, de Diego JI, Nistal M,
Quintanilla M, Portillo F and Cano A: Lysyl oxidase-like 2 as a new
poor prognosis marker of squamous cell carcinomas. Cancer Res.
68:4541–4550. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Offenberg H, Brünner N, Mansilla F,
Orntoft Torben F and Birkenkamp-Demtroder K: TIMP-1 expression in
human colorectal cancer is associated with TGF-B1, LOXL2, INHBA1,
TNF-AIP6 and TIMP-2 transcript profiles. Mol Oncol. 2:233–240.
2008. View Article : Google Scholar
|
|
34
|
Moreno-Bueno G, Salvador F, Martín A,
Floristán A, Cuevas EP, Santos V, Montes A, Morales S, Castilla MA,
Rojo-Sebastián A, et al: Lysyl oxidase-like 2 (LOXL2), a new
regulator of cell polarity required for metastatic dissemination of
basal-like breast carcinomas. EMBO Mol Med. 3:528–544. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Baker AM, Cox TR, Bird D, Lang G, Murray
GI, Sun XF, Southall SM, Wilson JR and Erler JT: The role of lysyl
oxidase in SRC-dependent proliferation and metastasis of colorectal
cancer. J Natl Cancer Inst. 103:407–424. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Baker AM, Bird D, Lang G, Cox TR and Erler
JT: Lysyl oxidase enzymatic function increases stiffness to drive
colorectal cancer progression through FAK. Oncogene. 32:1863–1868.
2013. View Article : Google Scholar
|
|
37
|
Ferrara N and Kerbel RS: Angiogenesis as a
therapeutic target. Nature. 438:967–974. PubMed/NCBI
|
|
38
|
Kerbel RS: Tumor angiogenesis. N Engl J
Med. 358:2039–2049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jubb AM and Harris AL: Biomarkers to
predict the clinical efficacy of bevacizumab in cancer. Lancet
Oncol. 11:1172–1183. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kerbel R and Folkman J: Clinical
translation of angiogenesis inhibitors. Nat Rev Cancer. 2:727–739.
2002. View
Article : Google Scholar : PubMed/NCBI
|
|
41
|
Baker AM, Bird D, Welti JC, Gourlaouen M,
Lang G, Murray GI, Reynolds AR, Cox TR and Erler JT: Lysyl oxidase
plays a critical role in endothelial cell stimulation to drive
tumor angiogenesis. Cancer Res. 73:583–594. 2013. View Article : Google Scholar :
|
|
42
|
Zaffryar-Eilot S, Marshall D, Voloshin T,
Bar-Zion A, Spangler R, Kessler O, Ghermazien H, Brekhman V,
Suss-Toby E, Adam D, et al: Lysyl oxidase-like-2 promotes tumour
angiogenesis and is a potential therapeutic target in angiogenic
tumours. Carcinogenesis. 34:2370–2379. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cassavaugh J and Lounsbury KM:
Hypoxia-mediated biological control. J Cell Biochem. 112:735–744.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang GL, Jiang BH, Rue EA and Semenza GL:
Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS
heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci
USA. 92:5510–5514. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Postovit LM, Abbott DE, Payne SL, Wheaton
WW, Margaryan NV, Sullivan R, Jansen MK, Csiszar K, Hendrix MJ and
Kirschmann DA: Hypoxia/reoxygenation: A dynamic regulator of lysyl
oxidase-facilitated breast cancer migration. J Cell Biochem.
103:1369–1378. 2008. View Article : Google Scholar
|
|
46
|
Schietke R, Warnecke C, Wacker I, Schödel
J, Mole DR, Campean V, Amann K, Goppelt-Struebe M, Behrens J,
Eckardt KU and Wiesener MS: The lysyl oxidases LOX and LOXL2 are
necessary and sufficient to repress E-cadherin in hypoxia: insights
into cellular transformation processes mediated by HIF-1. J Biol
Chem. 285:6658–6669. 2010. View Article : Google Scholar :
|
|
47
|
Pez F, Dayan F, Durivault J, Kaniewski B,
Aimond G, Le Provost GS, Deux B, Clézardin P, Sommer P, Pouysségur
J and Reynaud C: The HIF-1-inducible lysyl oxidase activates HIF-1
via the Akt pathway in a positive regulation loop and synergizes
with HIF-1 in promoting tumor cell growth. Cancer Res.
71:1647–1657. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bignon M, Pichol-Thievend C, Hardouin J,
Malbouyres M, Bréchot N, Nasciutti L, Barret A, Teillon J, Guillon
E, Etienne E, et al: Lysyl oxidase-like protein-2 regulates
sprouting angiogenesis and type IV collagen assembly in the
endothelial basement membrane. Blood. 118:3979–3989. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Payne SL, Fogelgren B, Hess AR, Seftor EA,
Wiley EL, Fong SF, Csiszar K, Hendrix MJ and Kirschmann DA: Lysyl
oxidase regulates breast cancer cell migration and adhesion through
a hydrogen peroxide–mediated mechanism. Cancer Res. 65:11429–11436.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kim BR, Dong SM, Seo SH, Lee JH, Lee JM,
Lee SH and Rho SB: Lysyl oxidase-like 2 (LOXL2) controls
tumor-associated cell proliferation through the interaction with
MARCKSL1. Cell Signal. 26:1765–1773. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Peinado H, Olmeda D and Cano A: Snail, Zeb
and bHLH factors in tumour progression: an alliance against the
epithelial phenotype? Nat Rev Cancer. 7:415–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Peinado H, Del Carmen Iglesias-de la Cruz
M, Olmeda D, Csiszar K, Fong KS, Vega S, Nieto MA, Cano A and
Portillo F: A molecular role for lysyl oxidase-like 2 enzyme in
snail regulation and tumor progression. EMBO J. 24:3446–3458. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Cuevas EP, Moreno-Bueno G, Canesin G,
Santos V, Portillo F and Cano A: LOXL2 catalytically inactive
mutants mediate epithelial-to-mesenchymal transition. Biol Open.
3:129–137. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang F, Ruan XJ and Zhang HY: BDE-99
(2,2′,4,4′,5-pentabro-modiphenyl ether) triggers
epithelial-mesenchymal transition in colorectal cancer cells via
PI3K/Akt/Snail signaling pathway. Tumori. 101:238–245. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lin JJ, Zhao TZ, Cai WK, Yang YX, Sun C,
Zhang Z, Xu YQ, Chang T and Li ZY: Inhibition of histamine receptor
3 suppresses glioblastoma tumor growth, invasion, and
epithelial-to-mesenchymal transition. Oncotarget. 6:17107–17120.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Rafael D, Doktorovová S, Florindo HF,
Gener P, Abasolo I, Schwartz S Jr and Videira MA: EMT blockage
strategies: Targeting Akt dependent mechanisms for breast cancer
metastatic behaviour modulation. Curr Gene Ther. 15:300–312. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
ClinicalTrials.gov: First-in-human study
of AB0024 to evaluate safety and tolerability in adults with
advanced solid tumors. https://clinicaltrials.gov/ct2/show/NCT01323933.
Accessed March 25, 2013.
|