Introduction
Osteoarthritis (OA), a cause of disability, is
characterized by changes in chondrocyte gene expression and
extracellular matrix (ECM) destruction (1,2).
Chondrocytes are the only cell type present in articular cartilage,
and their biosynthetic activities are necessary to maintain the
stability of the cartilage. Several lines of evidence indicate that
chondrocyte death is a key player in cartilage degeneration
(3–5). Chondrocyte death may occur due to
apoptosis; it is therefore possible that chondrocyte loss caused by
apoptosis plays a crucial role in the pathogenesis of OA (6,7).
Recent studies have also demonstrated that endoplasmic reticulum
(ER) stress, upstream of the mitochondria-dependent apoptotic
signal transduction pathway, plays a role in chondrocyte apoptosis
(8,9).
ER stress, as a new apoptotic pathway, results from
an imbalance between the load of unfolded proteins in the ER and
the capacity of the ER (10). It
is considered that there are three apoptotic pathways triggered by
ER stress: the C/EBP-homologous protein (Chop)/growth arrest and
DNA damage-inducible gene 153 (Gadd153) pathway, the c-JUN
NH2-terminal kinase (JNK) pathway and the caspase
pathway (11). The transcription
of the Chop gene is mediated by three signal transmission passages:
PKR-like ER kinase (PERK), activating transcription factor (Atf)6
and inositol-requiring protein 1 (IRE1) (12). The three protein molecules
activate the apoptotic genes, Gadd153 and TRAF2 (the gene for the
activation of JNK), and initiate chondrocyte apoptosis (12,13). In addition, the apoptotic genes,
caspase-3 and caspase-9, are activated through the caspase pathway,
thereby executing the final stages of apoptosis (14). Several critical signaling
molecules, such as binding immunoglobulin protein (Bip), X-box
binding protein 1 (Xbp1), Atf4, Bcl-2-associated X protein (Bax)
and B-cell lymphoma 2 (Bcl-2) have been shown to participate in the
process of apoptosis induced by ER stress (15). Thus, decreasing apoptosis through
by regulating the effects of related signaling molecules is crucial
to preventing the progressive degeneration of articular
cartilage.
Bushen Zhuangjin decoction (BZD) is a well-known
formulation that nourishes the Gan (liver) and the Shen (kidney)
and also strengthens tendons and bones, and has been long used in
China (16). Clinical studies
have demonstrated that BZD has a positive effect on the treatment
of OA, and experimental studies have found that BZD decreases the
apoptosis of chondrocytes and promotes chondrocyte proliferation
(17,18). However, the specific mechanisms
responsible for the inhibitory effects of BZD on ER stress-mediated
chondrocyte apoptosis remain to be elucidated. Therefore, in this
study, we aimed to determine whether BZD inhibits the tunicamycin
(TM)-induced apoptosis of chondrocytes mediated by ER stress and to
elucidate the underlying mechanisms.
Materials and methods
Preparation of the BZD aqueous
extract
BZD consists of the following 10 herbs: 12 g Shu Di
Huang (steamed Chinese foxglove, Radix Rehmanniae Glutinosae
Conquitae), 10 g Niu Xi (Achyranthes bidentata), 12 g Dang
Gui (Angelica sinensis), 12 g Fu Ling (Poria cocos),
12 g Xu Duan (Dipsacus), 12 g Shan Zhu Yu (Cornus
officinalis), 10 g Bai Shao (white peony root, Radix
Paeoniae Alba), 10 g Du Zhong (Eucommia bark), 10 g
Wu Jia Pi (Acanthopanax root bark) and 5 g Qing Pi (immature
tangerine peel, Pericarpium Citri Reticulatae Viride). All
herbs were purchased from the Third People's Hospital of Fujian
University of Traditional Chinese Medicine (FJUTCM; Fuzhou, China)
and were identified by the Teaching and Research section of FJUTCM.
The components were mixed and extracted with standard methods
according to Chinese Pharmacopoeia (Chinese Pharmacopoeia
Committee, 2010). The herbs of BZD were extracted with distilled
water by a refluxing method and were then filtered and
concentrated. The filtrate of BZD was evaporated using a rotary
evaporator (model RE-2000; Shanghai Yarong Biochemistry Instrument
Factory, Shanghai, China) and was then dried to a constant weight
with a vacuum drying oven (model DZF-300; Shanghai Yiheng
Scientific Instrument Co., Ltd., Shanghai, China). The water
extract of BZD was finally dissolved in phosphate-buffered saline
(PBS; HyClone Laboratories, Inc., Logan, UT, USA) to the
concentration of 10 mg/ml and was stored at −20°C. The working
concentrations of BZD were prepared by diluting the stock solution
in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal
bovine serum (FBS) (both from HyClone Laboratories, Inc.) and then
by filtering it through a 0.22-µm filter and storing it at
4°C.
Chondrocyte culture and
identification
Articular chondrocytes were isolated and cultured as
previously described (19). In
brief, 4 SD rats were sacrificed by cervical dislocation. The
articular cartilage was cut out from the bilateral knee joints,
peeled off under sterile conditions and rinsed with PBS (HyClone
Laboratories, Inc.) 3 times. The cartilage was cut into sections of
1 mm3 and digested with 0.2% collagenase II
(Sigma-Aldrich, St. Louis, MO, USA) in a 37°C 5% CO2
incubator. The supernatant fluid was collected every 90 min, and
centrifuged for 5 min (1,000 rpm/min). The precipitate was
resuspended with DMEM containing 10% FBS (both from HyClone
Laboratories, Inc.) and transferred onto a 25 mm2
culture flask and cultured in a 37°C 5% CO2 incubator;
these were termed the primary chondrocytes. The Sprague Dawley (SD)
rats used in this study were purchased from Shanghai SLAC
Laboratory Animal Co., Ltd. (Shanghai, China). The present study
was approved by the Institutional Animal Care and Use Committee of
FJUTCM. The morphological changes and growth characteristics of the
chondrocytes were recorded under a phase contrast microscope
(Olympus, Tokyo, Japan). To identify the chondrocytes,
immunocytochemical staining for type II collagen was performed.
Briefly, the cells were seeded onto sterilized coverslips and
placed into wells of a 12-well plate (2×105 cells/
well), and cultured in a 37°C 5% CO2 incubator for 48 h.
The cells were fixed with 4% paraformaldehyde (Sigma-Aldrich) at
4°C for 30 min, and blocked with 10% bovine serum albumin (BSA;
Sigma-Aldrich) for 1 h. The slides were incubated with a rabbit
polyclonal antibody to type II collagen (BS1071; Bioworld
Technology, Inc., St. Louis Park, MN, USA) overnight at 4°C. The
slides were then incubated with peroxidase-conjugated affinipure
goat anti-rabbit IgG (ZB-2301; Zhongshan GoldenBridge
Biotechnology, Co., Ltd., Beijing, China) for 30 min, followed by
incubation with diaminobenzidine (ZLI-9018 DAB kit; Zhongshan
GoldenBridge Biotechnology, Co., Ltd.) for 5 min. The slides were
finaly counterstained with hematoxylin (Sigma-Aldrich) and
dehydrated. Images were captured using a light microscope (BH2;
Olympus, Tokyo, Japan). The primary chondrocytes were termed
passage (P)0 and P2 chondrocytes at approximately 80% confluence
were used in our experiments.
Cell viability assay
The chondrocytes were seeded in 96-well culture
plates at a density of 5×104 cells/ml and incubated (100
µl/well) for 24 h. The chondrocytes were stimulated with 2
µg/ml TM (Sigma-Aldrich) as the model group. A series of
concentrations of BZD (50, 100, 200, 400, 800, 1200 µg/ml)
were also added in the presence of TM follwed by incubation for 24,
48 or 72 h. Following intervention, 100 µl 1% MTT
(Sigma-Aldrich) were added to each well followed by incubation at
37°C for 4 h, and the supernatant was then replaced with 150
µl/well of DMSO (Hengxing Chemical Preparation Co., Ltd.,
Tianjin, China) and shaken for 10 min. The absorbance was detected
using an ELISA reader (model ELX800; BioTek Instruments, Inc.,
Winooski, VT, USA) by measuring the optical denstiy (OD) value at
490 nm.
Experimental design
According to the assay, there were 4 experimental
groups as follows: the control group, which included P2
chondrocytes without treatment; the model group, which included
apoptotic chondrocytes stimulated with TM; the dosing group, which
included TM-stimulated chondrocytes treated with BZD; and the
positive control group, which included TM-stimulated chondrocytes
treated with 5 mM 4-phenylbutyric acid (4-PBA) (Sigma-Aldrich), as
previously described (20).
4′,6-Diamidino-2-phenylindole (DAPI)
staining
The chondrocytes were washed with PBS and fixed with
4% paraformaldehyde (Sigma-Aldrich) at 4°C for 15 min. The
chondrocytes were stained in 5 µg/ml DAPI for 5 min. Τhe
images of apoptotic chondrocyte morphology were captured using a
fluorescence microscope (Olympus).
Detection of apoptosis by flow cytometry
with Annexin V-FITC and JC-1 staining
The apoptotic rate of the chondrocytes was detected
with Annexin V/PI staining (Nanjing KeyGen Biotech, Nanjing, China)
using a fluorescence-activated cell sorting (FACS) machine
(FACSCalibur™; Becton-Dicskinson Biosciences, San Diego, CA, USA).
The changes in mitochondrial membrane potential (ΔΨm) were measured
using a JC-1 kit (Nanjing Keygen Biotech), and the processed cells
were also analyzed using the FACS machine. Both staining procedures
were carried out according to the relevant manufacturer's
instructions.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was extracted from the cells using TRIzol
reagent (Invitrogen Life Technologies, Grand Island, NY, USA). RNA
(1 µg) was reverse transcribed into cDNA using a reverse
transcription kit (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) according to the manufacturer's instructions. Subsequently,
the DNA bands were analyzed by gel electrophoresis (1.5% agarose)
using a gel documentation system (Bio-Rad, Hercules, CA, USA) and
normalized to those of β-actin. The PCR primers used were as
follows: β-actin forward, 5′-GAG AGG GAA ATC GTG CGT GAC-3′ and
reverse, 5′-CAT CTG CTG GAA GGT GGA CA-3′; Bip forward, 5′-ATC AAC
CCA GAT GAG GCT GTA GCA-3′ and reverse, 5′-AGA CCT TGA TTG TTA CGG
TGG GCT-3′; Xbp1 forward, 5′-AGC ATA GGC CTG TCT GCT TCA CTA-3′ and
reverse, 5′-TGG TAA AGT CCA GCA CTT GGG AGT-3′; Atf4 forward,
5′-AAT GGC TGG CTA TGG ATG GG-3′ and reverse, 5′-TGT CTG AGG GGG
CTC CTT ATT AG-3′; caspase-9 forward, 5′-GCC TCA TCATCA ACA ACG-3′
and reverse, 5′-CTG GTA TGG GAC AGC ATC T-3′; caspase-3 forward,
5′-GGA CCT GTG GAC CTG AAA-3′ and reverse, 5′-GGG TGC GGT AGA GTA
AGC-3′; Bcl-2 forward, 5′-TGG CAT CTT CTC CTT CCC-3′ and reverse,
5′-GGT ACA TCT CCC TGT TGA CG-3′; Chop forward, 5′-TTC ACT ACT CTT
GAC CCT GCG TC-3′ and reverse, 5′-CAC TGA CCA CTC TGT TTC CGT
TTC-3′; Bax forward, 5′-GGC GAT GAA CTG GAC AAC-3′ and reverse,
5′-TCC CGA AGT AGG AAA GGA G-3′.
Western blot analysis
Total protein was extracted from the cells using
radioimmunoprecipitation assay (RIPA) lysis buffer (Beyotime
Biotech, Shanghai, China) with 1 mM phenylmethanesulfonyl fluoride
(PMSF; Beyotime Biotech), and the bicinchoninic acid assay (BCA)
was used to measure the protein concentrations. Proteins were
separated by electrophoresis on SDS-PAGE gels (12%) and
electrotransferred onto PVDF membranes (Thermo Fisher Scientific,
Inc.). The membranes were blocked in 5% skimmed milk, then
incubated with primary antibodies against Bip (3183S), Xbp1
(12782), Atf4 (11815S), Chop (5554S), Bcl-2 (2876S), caspase-9
(9506S), caspase-3 (9662S) and Bax (2772S; all from Cell Signaling
Technology, Inc., Beverly, MA, USA) and β-actin (sc-47778; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA) at 4°C overnight,
followed by goat anti-rabbit horseradish peroxidase-conjugated
(HRP) secondary antibody IgG (ZB-2301; Zhongshan GoldenBridge
Biotechnology, Co., Ltd.) at room temperature for 1 h. Eventually,
the blots were detected by using a Bio-Rad ChemiDoc XRS+ imaging
system (Bio-Rad), and β-actin was used as the internal control.
Statistical analysis
Data were analyzed by one-way analysis of variance
(ANOVA) or the Student's t-test using SPSS 19.0 software and all
values are expressed as the means ± SD. A value of P<0.01 was
considered to indicate a statistically significant difference. A
value of P<0.05 was considered a trend.
Results
Morphology and identification of
chondrocytes
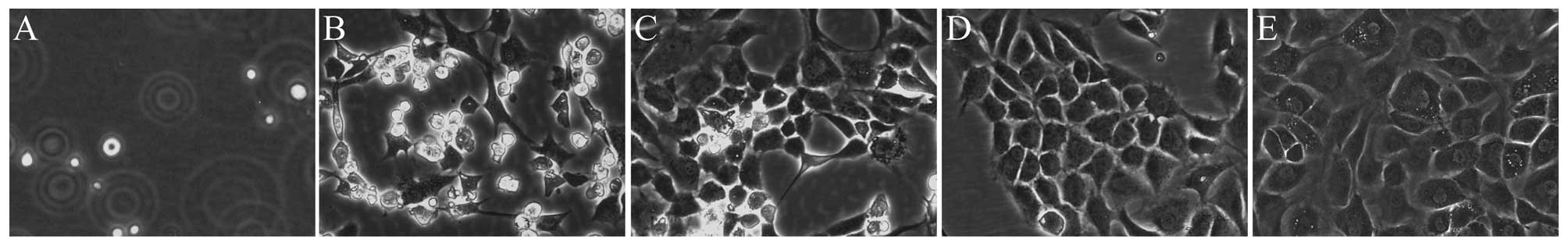
The chondrocytes in our study exhibited a morphology
which is considered typical of chondrocytes, with a spherical,
fusiform and slab-stone shape (Fig.
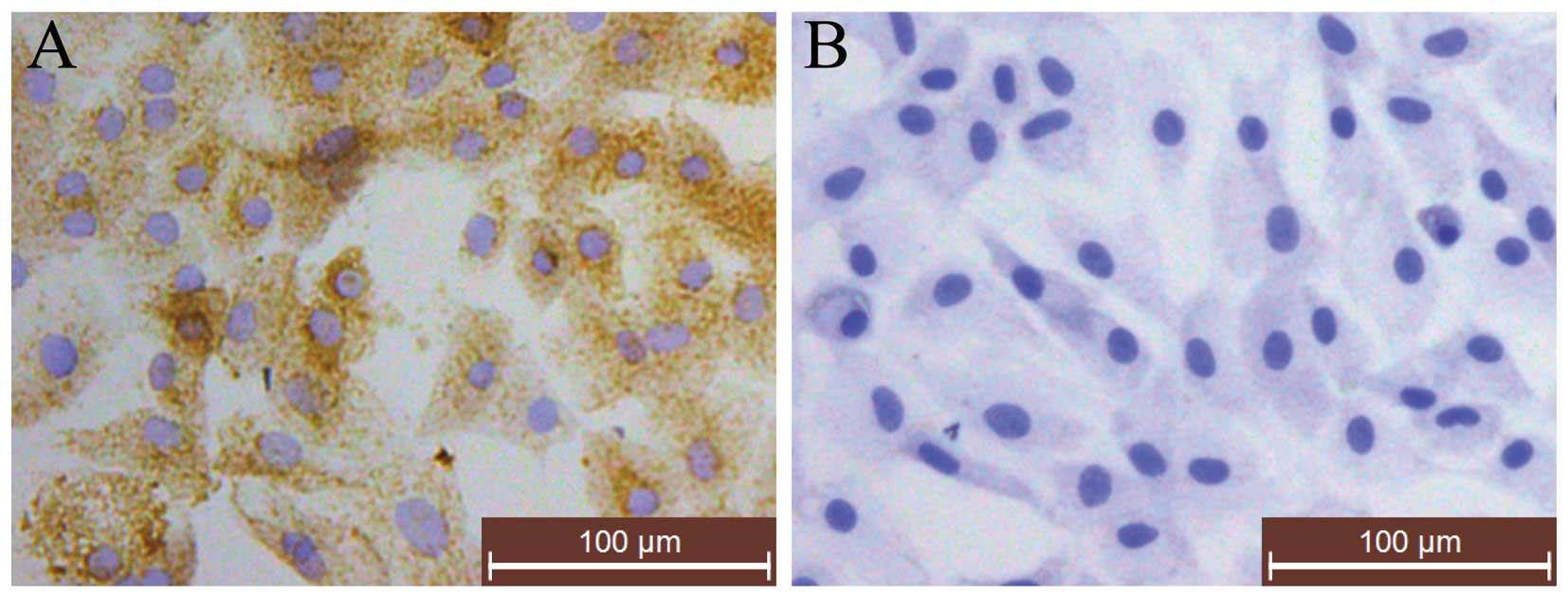
1), as described in previous studies (21,22). Immunohistochemical staining for
type II collagen indicated that the cytoplasm was stained brown,
which represented the positive expression of type II collagen,
while no brown staining was observed in the negative control cells
(Fig. 2). Following toluidine
blue staining, red/purple particles in the cytoplasm were observed
under a light microscope, which indicated the staining of
proteoglycans (23) (Fig. 2). Thus, P2 chondrocytes were used
in our experiments, as they were proven to be rich in ECM
components (type II collagen) and had the typical morphology of
chondrocytes.
BZD enhances the viability of
chondrocytes stimulated with TM
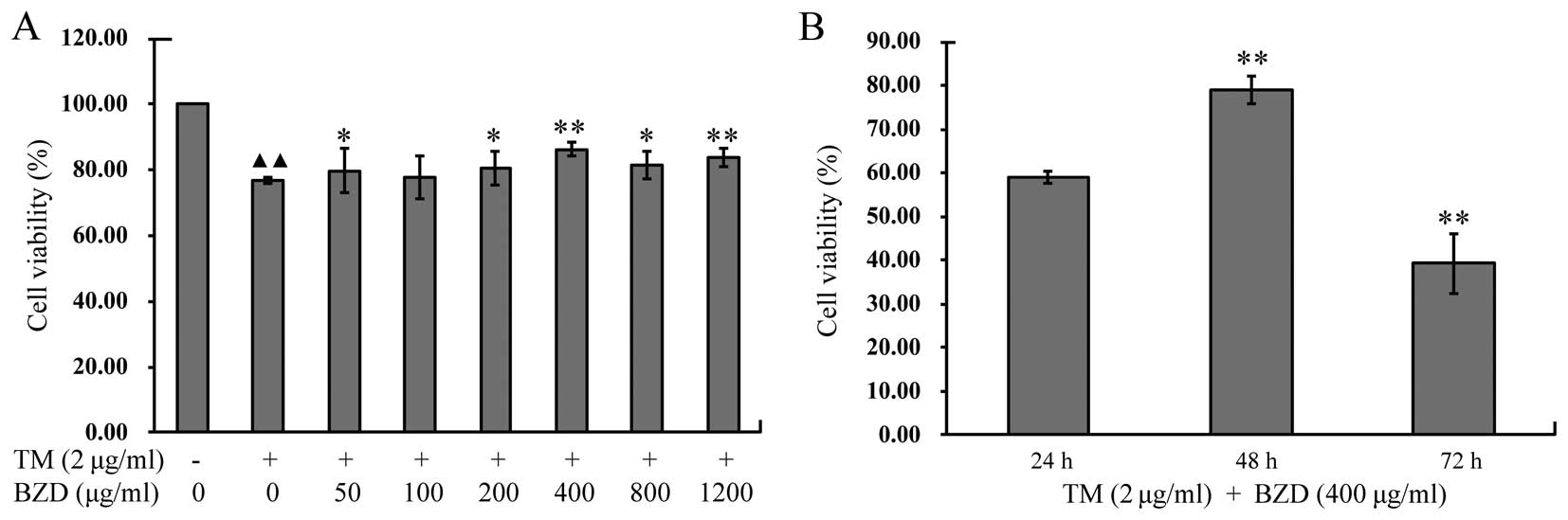
To examine the effects of BZD on chondrocyte
viability, the viability of the TM-stimulated chondrocytes treated
with BZD at various concentrations for different periods of time
was detected by MTT assay. The viability of the TM-stimulated
chondrocytes was markedly lower than that of the untreated cells in
the control group (P<0.01). The viability of the TM-stimulated
chondrocytes treated with various concentrations of BZD was
enhanced compared with the untreated TM-stimulated chondrocytes
(P<0.01 or P<0.05; Fig.
3A). The viability of the TM-stimulated chondrocytes treated
with 400 µg/ml BZD for 48 h was markedly enhanced compared
with that of the TM-stimulated chondrocytes treated with 400
µg/ml BZD for 24 or 72 h (P<0.01;Fig. 3B). These findings indicate that
BZD exerts a positive effect on TM-stimulated chondrocytes in a
dose- and time-dependent manner. Finally, the most suitable
concentration (400 µg/ml) of BZD for 48 h of treatment was
used in all the subsequent experiments.
Effects of BZD on the morphology of
TM-stimulated chondrocytes
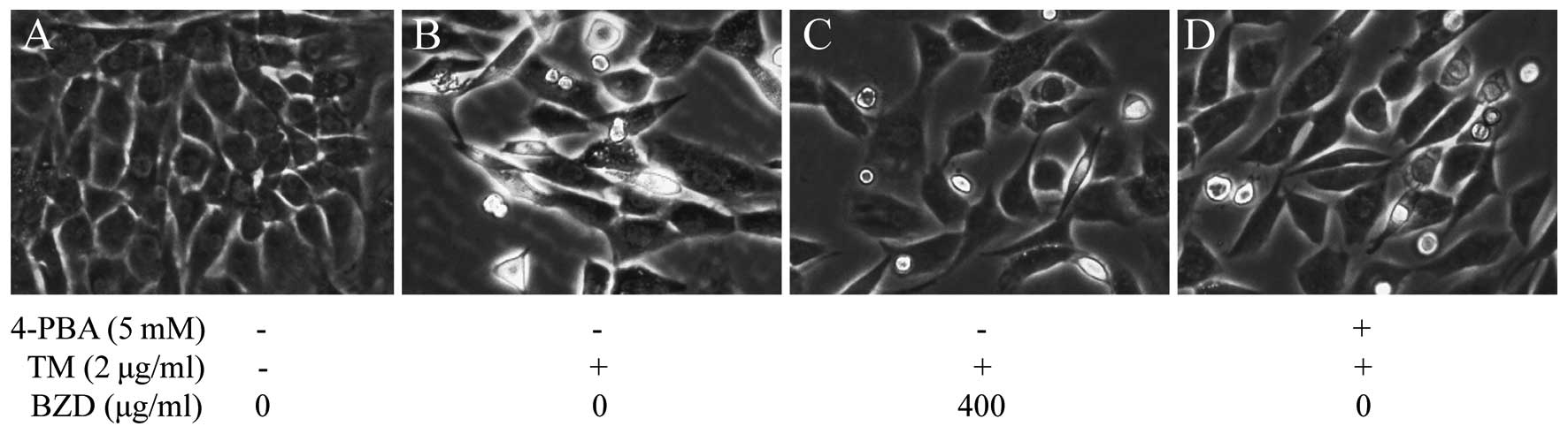
Following treatment, the morphological changes of
the TM-stimulated chondrocytes were observed under a phase contrast
micros cope. It was observed that the untreated cells in the
control group retained their normal growth state and exhibited no
significant changes in morphology (Fig. 4A). However, the TM-stimulated
chondrocytes became elongated and shrank in size, and were observed
floating in the medium, and were also detached from each other
(Fig. 4B). However, these changes
in cell morphology induced by stimulation with TM were not observed
or were less evident in the TM-stimulated chondrocytes treated with
BZD or 4-PBA than in the model group (Fig. 4C and D).
BZD decreases the apoptosis of
TM-stimulated chondrocytes
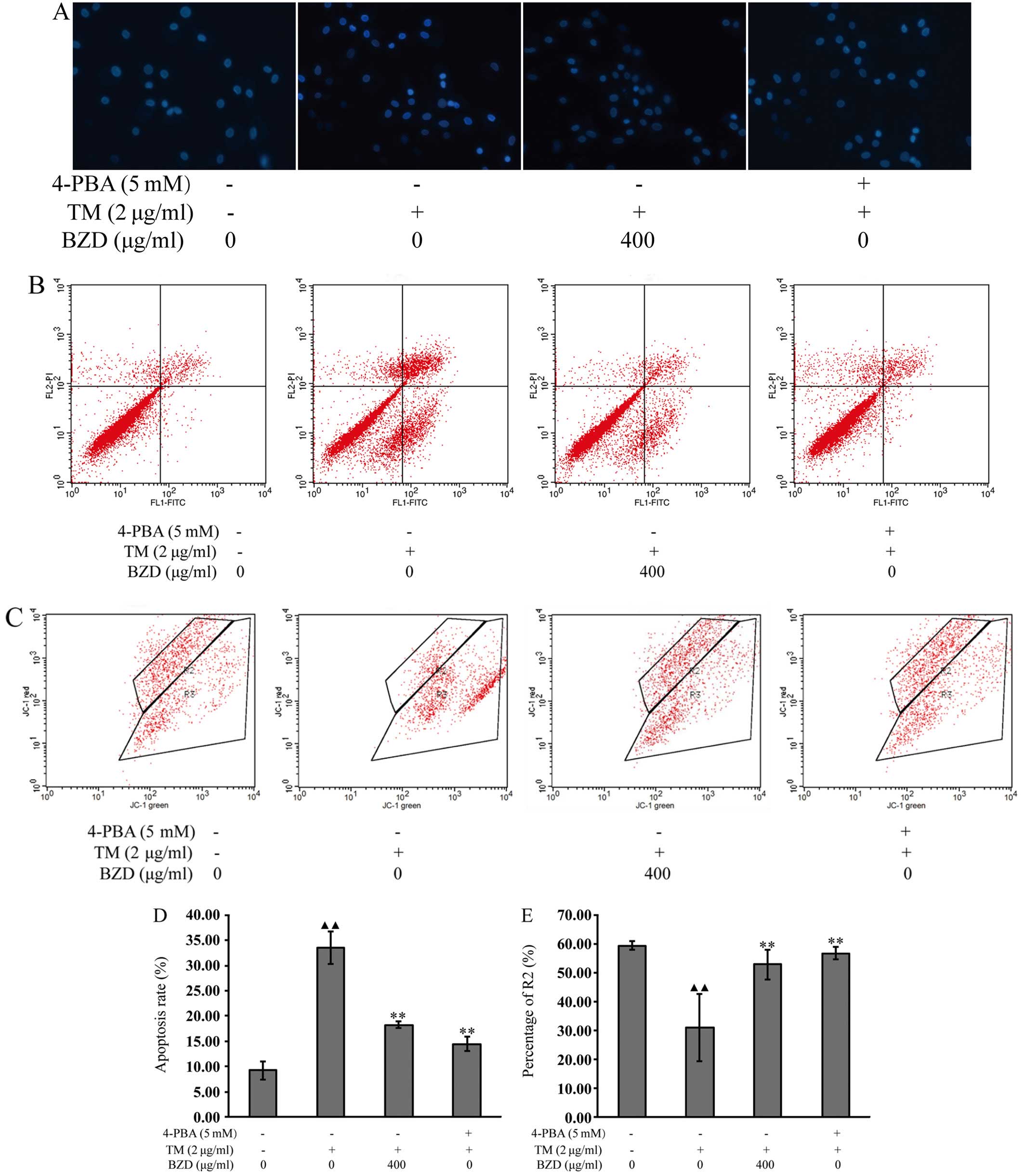
DAPI staining was used to monitor cell apoptosis, by
emitting a strong fluorescence signal when DAPI combined with DNA
in the nucleus. Normal nuclei were round in shape and light blue in
color. However, nuclei of the apoptotic cells were stained bright
blue and became deformed or fragmented due to the aggregation of
DNA in the TM-stimulated cells. In the dosing and positive control
groups, the apoptosis of the chondrocyte nuclei was still evident,
but to a lesser extent than the model group (Fig. 5A).
To further examine the effects of BZD on the
apoptosis of TM-stimulated chondrocytes, Annexin V-FITC binding
assay was used to measure chondrocyte apoptosis. As shown in
Fig. 5B and D, the number of
dead, apoptotic cells and the apoptotic rate of the TM-stimulated
chondrocytes treated with BZD were significantly lower than those
of the TM-stimulated cells not treated with BZD. The results
revealed that BZD exerted a positive effect by reducing or
inhibiting the apoptosis of TM-stimulated chondrocytes.
JC-1 is an indicator of the initiation of cell
apoptosis that is widely used to quantify ΔΨm by measuring the
fluorescence intensities. The effects of BZD on the changes in ΔΨm
were examined by JC-1 assay. As shown in Fig. 5C and E, a significant reduction in
ΔΨm in the TM-stimulated chondrocytes was observed compared with
the untreated cells. When compared with the TM-stimulated
chondrocytes, the ΔΨm in the TM-stimulated chondrocytes treated
with BZD or 4-PBA was markedly increased. This suggests that BZD
exerts a positive effect on TM-stimulated chondrocytes as it
inhibits the reduction of ΔΨm in these cells.
BZD inhibits ER stress-mediated
chondrocyte apoptosis
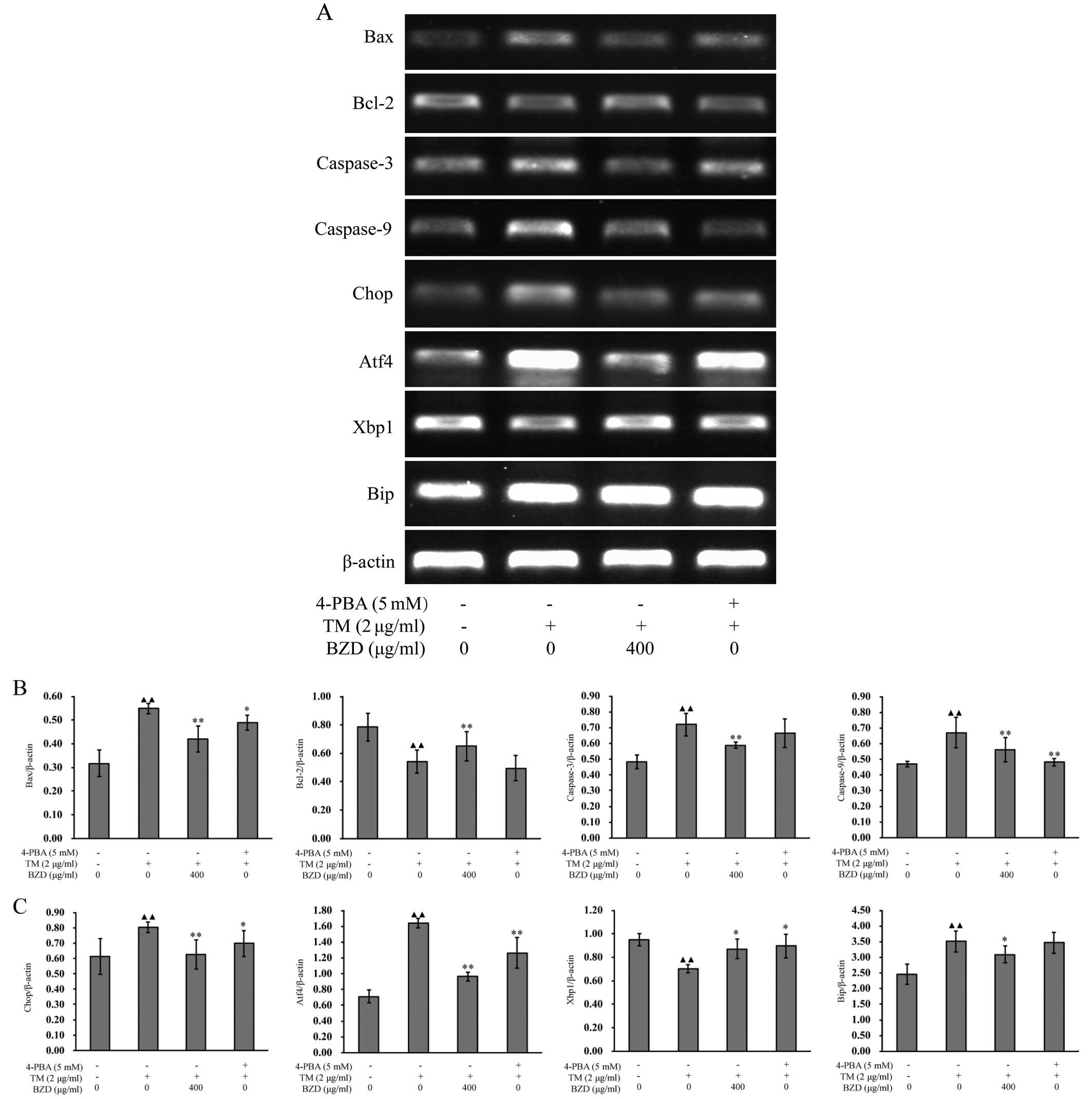
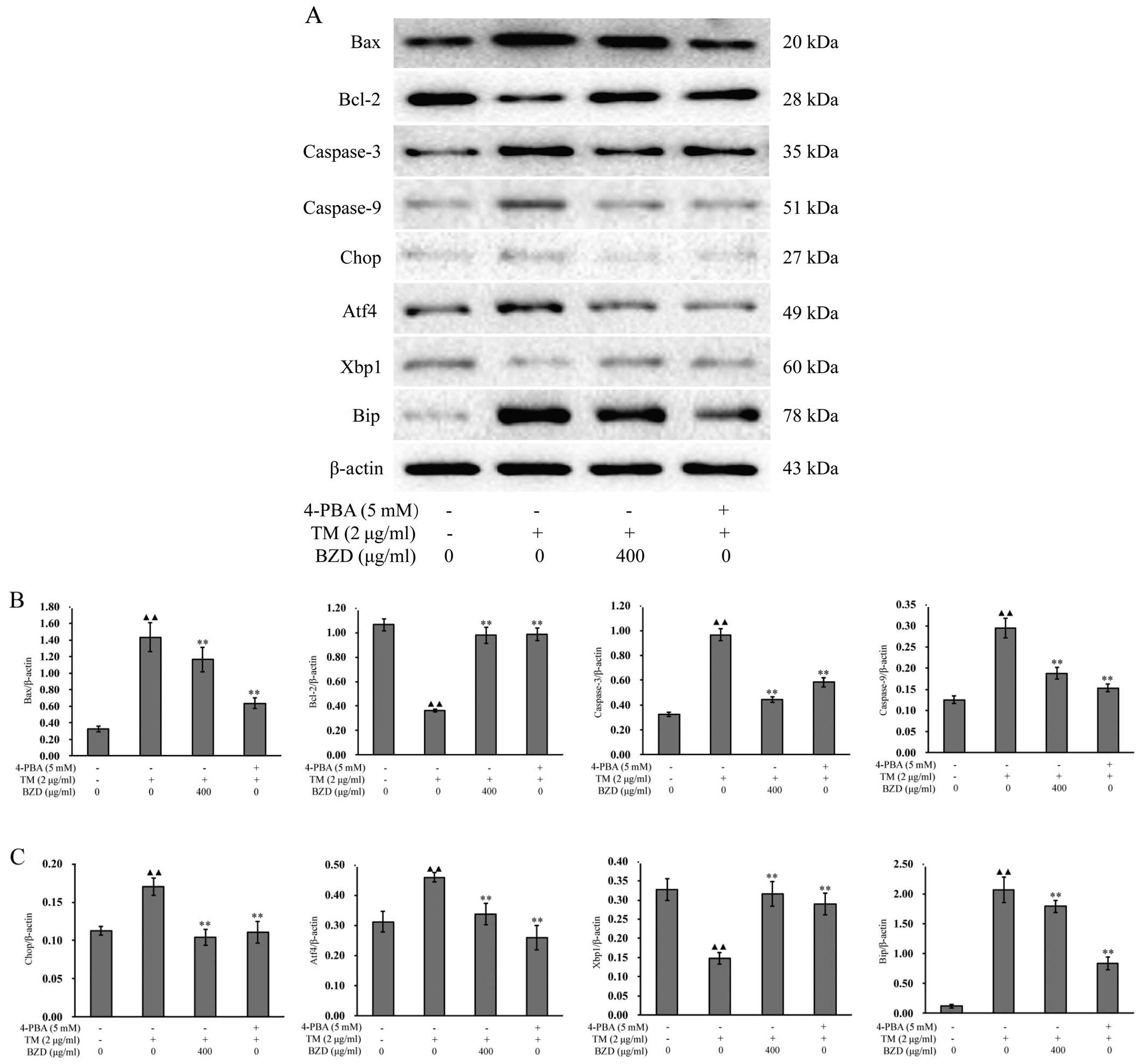
To confirm the inhibitory effects of BZD on ER
stress-mediated chondrocyte apoptosis, we measured the expression
levels of established the ER stress-associated markers (24), Bip, Xbp1, Chop and Atf4. When ER
stress is unresolved, the cell is functionally compromised and may
undergo apoptosis (25). Some
pathways have been directly implicated in ER stress-induced
apoptosis, such as those involving caspases,the Bcl-2 family and
the pro-apoptotic member, Bax (26,27). In order to further eludicate the
mechanisms responsible for the inhibitory effects of BZD on the
apoptosis of TM-stimulated chondrocytes, the mRNA and protein
expression levels of apoptosis-associated markers were measured by
RT-PCR and western blot analysis, respectively. As shown by the
results of RT-PCR, when compared with the model group, the mRNA
expression of Bip, Atf4, Chop, Bax, caspase-3 and caspase-9 in the
dosing group was downregulated, whereas the expression of Xbp1 and
Bcl-2 was upregulated (Fig. 6).
Furthermore, the changes in the expression levels of all the
aforementioned markers in the dosing group showed no significant
differences compared to those observed in the positive control
group (P<0.01, P<0.05; Fig.
6). The results obtained for the protein levels were very
similar (P<0.01, P<0.05; Fig.
7). This suggests that BZD inhibits the apoptosis of
TM-stimulated chondrocytes by regulating the ER stress pathway.
Discussion
Increasing evidence indicates that there is an
association between cartilage degradation and chondrocyte apoptosis
(28,29). Therefore, the inhibition of
chondrocyte apoptosis is essential and may be a potential
therapeutic strategy for OA. In the present study, we investigated
the effects of BZD on TM-induced chondrocyte apoptosis mediated by
ER stress. Our results demonstrated that BZD inhibited the
apoptosis of TM-stimulated chondrocytes by regulating the ER stress
signaling pathway, as evidenced by the upregulation of the
expression of Xbp1 and Bcl-2 and the downregulation of the
expression of Bip, Atf4, Chop, Bax, caspase-3 and caspase-9.
Currently, pharmacological approaches to the treatment of OA
include analgesics, anti-inflammatory agents, intraarticular
corticosteroids and hyaluronic acid (30). However, several adverse effects
have been associated with these drugs (31–33). Thus, there has been a growing
interest in Chinese herbal medicines as they have the same
benefits, and are less costly and have fewer adverse effects
(34–36). BZD has a good structure and
exhibits diverse physicochemical properties. Moreover, it exerts
potentially drug-like and multi-target effects (37,38). There is evidence that BZD is
effective in the treatment of OA in both clinical and in in
vitro studies (16,17).
ER stress activates a set of signaling pathways
collectively known as the unfolded protein response (UPR) (39). TM is a bacterial toxin that has
been shown to inhibit the N-linked glycosylation of nascent
proteins and to lead to the activation of UPR in mammalian cells
(40). It is generally used as an
ER stress inducer. Thus, TM-stimulated chondrocytes were used as a
cellular model of apoptosis in the present study. It was observed
that the morphological changes of the TM-stimulated chondrocytes
were identical to those accompanying apoptosis, which indicated
that the model of apoptosis was successfully established.
Additionally, 4-PBA is a chemical chaperone whose beneficial
effects have been associated with the suppressed expression of ER
stress markers (41). Hence, the
TM-stimulated chondrocytes treated with 4-PBA were used as the
positive control.
MTT assay is used to monitor cytotoxicity,
proliferation and activation (42). In our study, the viability of both
untreated and treated cells was measured by MTT assay. According to
the results, the viability of the TM-stimulated chondrocytes was
evidently enhanced by BZD in a dose- and time-dependent manner. To
further examine the effects of BZD on the apoptosis of
TM-stimulated chondrocytes, DAPI, Annexin V-FITC binding assay and
JC-1 assay were performed. Annexins, the Ca2+-dependent
and phospholipid-binding proteins, specifically bind to
phosphatidylserine (PS) in the plasma membrane (43). When apoptosis occurs, PS is
translocated to the extracellular layer from the cytosolic layer of
the plasma membrane, and Annexin V-TITC binding assay is used to
detect PS exposure on the membranes of apoptotic cells (44). JC-1 assay detects the loss of ΔΨm.
Alterations in ΔΨm are indicative of changes in the mitochondrial
metabolic activity leading to cell cycle arrest and apoptosis;
therefore, ΔΨm is an indicator of cell integrity (45). The results of the Annexin V and
JC-1 assays further established the occurrence of apoptosis and
demonstrated that BZD reduced the loss of ΔΨm, thereby inhibiting
chondrocyte apoptosis.
ER stress is caused by the accumulation of unfolded
proteins. Bip maintains the non-activated state of signal
transduction factors while combining the end of PERK, Atf6 and
IRE1. Xbp1 mRNA, a transcription factor, is frame switched by the
removal of a small intron (46).
It has been found that ER stress is induced during chondrocyte
differentiation and activates the IRE1-Xbp1 pathway (47). ER membrane-associated initiator
caspases, such as caspase-3, are also activated by IRE1α (12). PERK makes a crucial contribution
to enhancing the expression of Chop in the UPR, subsequently
inducing the translational upregulation of Atf4 by transmitting
signals phosphorylating eIF-2a (48,49).
Although apoptosis can be induced by several
different triggers, it is characteristically associated downstream
with the sequential activation of caspases (50). The caspase family of enzymes is a
key factor in cell apoptosis. Caspase-9 is a member of the caspase
family of cysteine proteases, which has been implicated in
apoptosis and cytokine processing (51). Caspase-3 is one of the effector
caspases that mediates the final stage of cell death by apoptosis
(52). It was previously found
that the expression of caspase-3 was ~2-fold higher in OA-affected
cartilage and correlated with chondrocyte apoptosis, activated by
caspase-9 (53). Bax
(pro-apoptotic molecule) and Bcl-2 (anti-apoptotic molecule) are
two biomarkers which are associated with the mechanisms of cell
death by apoptosis in OA. It has been demonstrated that Bax is
upregulated in OA cartilage and that Bcl-2 is downregulated
(54). Bcl-2 is a cytoplasmic
protein that is involved in promoting cell survival and preventing
apoptosis (55). The
transcription of Bcl-2 is regulated by Chop; thus, the upregulation
of Bcl-2 may occur during cartilage repair in OA. In our study, the
changes in the mRNA and protein levels of these markers were
detected by RT-PCR and western blot analysis, respectively. On the
one hand, treatment with BZD led to the downregulation of the
expression of Bip, Atf4, Chop, Bax, caspase-3 and caspase-9 in the
dosing group, thus decreasing chondrocyte apoptosis. On the other
hand, BZD upregulated the expression levels of Xbp1 and Bcl-2.
Taken together, our findings demonstrated that BZD inhibited the
apoptosis of TM-stimulated chondrocytes by regulating the ER stress
pathway.
In conclusion, our data demonstrate that BZD
effectively exerted its anti-apoptotic effects on chondrocytes
stimulated with TM. The inhibition or prevention of chondrocyte
apoptosis was mainly the result of the suppression of the ER stress
pathway. The findings of the present study indicate that BZD is a
potent therapeutic agent for the treatment of OA. However, due to
the limitations of in vitro experiments, the exact
mechanisms responsible for the inhibition of ER stress-mediated
chondrocyte apoptosis by BZD were not completely elucidated. Thus,
further studies are warranted to examine the effects of BZD on OA
both in vitro and in vivo.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant no. 81373818), the Key
Project of Fujian Provincial Department of Science and Technology
Department (grant no. 2014Y0064), the Natural Science Foundation of
Fujian Province (grant no. 2014J01357), the Special Research Fund
for Doctor Discipline in College (grant no. 20123519110001) and the
Developmental Fund of Chen Keji Integrative Medicine (grant no. CKJ
2015010).
References
|
1
|
Chu CR, Millis MB and Olson SA:
Osteoarthritis: from palliation to prevention: AOA critical issues.
J Bone Joint Surg Am. 96:e1302014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Loeser RF: Aging and osteoarthritis: the
role of chondrocyte senescence and aging changes in the cartilage
matrix. Osteoarthritis Cartilage. 17:971–979. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Del Carlo M Jr and Loeser RF: Cell death
in osteoarthritis. Curr Rheumatol Rep. 10:37–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim HA, Lee YJ, Seong SC, Choe KW and Song
YW: Apoptotic chondrocyte death in human osteoarthritis. J
Rheumatol. 27:455–462. 2000.PubMed/NCBI
|
|
5
|
Thomas CM, Fuller CJ, Whittles CE and
Sharif M: Chondrocyte death by apoptosis is associated with the
initiation and severity of articular cartilage degradation. Int J
Rheum Dis. 14:191–198. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hashimoto S, Ochs RL, Komiya S and Lotz M:
Linkage of chondrocyte apoptosis and cartilage degradation in human
osteoarthritis. Arthritis Rheum. 41:1632–1638. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Almonte-Becerril M, Navarro-Garcia F,
Gonzalez-Robles A, Vega-Lopez MA, Lavalle C and Kouri JB: Cell
death of chondrocytes is a combination between apoptosis and
autophagy during the pathogenesis of Osteoarthritis within an
experimental model. Apoptosis. 15:631–638. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Han X, Zhang P, Jiang R, Xia F, Li M and
Guo FJ: Explore on the effect of ATF6 on cell growth and apoptosis
in cartilage development. Histochem Cell Biol. 142:497–509. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Uehara Y, Hirose J, Yamabe S, Okamoto N,
Okada T, Oyadomari S and Mizuta H: Endoplasmic reticulum
stress-induced apoptosis contributes to articular cartilage
degeneration via C/EBP homologous protein. Osteoarthritis
Cartilage. 22:1007–1017. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ron D and Walter P: Signal integration in
the endoplasmic reticulum unfolded protein response. Nat Rev Mol
Cell Biol. 8:519–529. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang L, Xie H and Liu H: Endoplasmic
reticulum stress, diabetes mellitus, and tissue injury. Curr
Protein Pept Sci. 15:812–818. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Oyadomari S and Mori M: Roles of
CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ.
11:381–389. 2004. View Article : Google Scholar
|
|
13
|
Urano F, Wang X, Bertolotti A, Zhang Y,
Chung P, Harding HP and Ron D: Coupling of stress in the ER to
activation of JNK protein kinases by transmembrane protein kinase
IRE1. Science. 287:664–666. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Matsuo M, Nishida K, Yoshida A, Murakami T
and Inoue H: Expression of caspase-3 and -9 relevant to cartilage
destruction and chondrocyte apoptosis in human osteoarthritic
cartilage. Acta Med Okayama. 55:333–340. 2001.
|
|
15
|
Takada K, Hirose J, Senba K, Yamabe S,
Oike Y, Gotoh T and Mizuta H: Enhanced apoptotic and reduced
protective response in chondrocytes following endoplasmic reticulum
stress in osteo-arthritic cartilage. Int J Exp Pathol. 92:232–242.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou GS, Li XF and Guan GH: Effects of
Bushen Zhuangjin Decoction containing serum on the apoptosis of
chondrocytes induced by mechanics stimulus. Zhongguo Zhong Xi Yi
Jie He Za Zhi. 32:789–792. 2012.In Chinese. PubMed/NCBI
|
|
17
|
Li X, Chen J, Liang W, Li H, Liu F, Weng
X, Lin P, Chen W, Zheng C, Xu H, et al: Bushen Zhuangjin Decoction
promotes chondrocyte proliferation by stimulating cell cycle
progression. Exp Ther Med. 9:839–844. 2015.PubMed/NCBI
|
|
18
|
Li X, Liang W, Dang C, et al: Empirical
study on Bushen Zhuangjin Decoction inhibiting inflammatory
cytokine expression experiments to delay the degeneration of
articular cartilage. Feng Shi Bing Yu Guan Jie Yan. 3:20–25.
2014.In Chinese.
|
|
19
|
Li H, Li X, Liu G, Chen J, Weng X, Liu F,
Xu H, Liu X and Ye H: Bauhinia championi (Benth.) Benth.
polysaccharides upregulate Wnt/β-catenin signaling in chondrocytes.
Int J Mol Med. 32:1329–1336. 2013.PubMed/NCBI
|
|
20
|
Zeng W, Guo YH, Qi W, Chen JG, Yang LL,
Luo ZF, Mu J and Feng B: 4-Phenylbutyric acid suppresses
inflammation through regulation of endoplasmic reticulum stress of
endothelial cells stimulated by uremic serum. Life Sci. 103:15–24.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Weng X, Lin P, Liu F, Chen J, Li H, Huang
L, Zhen C, Xu H, Liu X, Ye H and Li X: Achyranthes bidentata
polysaccharides activate the Wnt/β-catenin signaling pathway to
promote chon-drocyte proliferation. Int J Mol Med. 34:1045–1050.
2014.PubMed/NCBI
|
|
22
|
Yu F, Li X, Cai L, Li H, Chen J, Wong X,
Xu H, Zheng C, Liu X and Ye H: Achyranthes bidentata
polysaccharides induce chon-drocyte proliferation via the promotion
of the G1/S cell cycle transition. Mol Med Rep. 7:935–940.
2013.PubMed/NCBI
|
|
23
|
Ishizeki K, Kagiya T, Fujiwara N, Otsu K
and Harada H: Expression of osteogenic proteins during the
intrasplenic transplantation of Meckel's chondrocytes: a
histochemical and immunohistochemical study. Arch Histol Cytol.
72:1–12. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Araki E, Oyadomari S and Mori M:
Endoplasmic reticulum stress and diabetes mellitus. Intern Med.
42:7–14. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rojas C, Pan-Castillo B, Valls C, Pujadas
G, Garcia-Vallve S, Arola L and Mulero M: Resveratrol enhances
palmitate-induced ER stress and apoptosis in cancer cells. PLoS
One. 9:e1139292014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Riedl SJ and Shi Y: Molecular mechanisms
of caspase regulation during apoptosis. Nat Rev Mol Cell Biol.
5:897–907. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
McCullough KD, Martindale JL, Klotz LO, Aw
TY and Holbrook NJ: Gadd153 sensitizes cells to endoplasmic
reticulum stress by down-regulating Bcl2 and perturbing the
cellular redox state. Mol Cell Biol. 21:1249–1259. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim HT, Lo MY and Pillarisetty R:
Chondrocyte apoptosis following intraarticular fracture in humans.
Osteoarthritis Cartilage. 10:747–749. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
D'Lima DD, Hashimoto S, Chen PC, Colwell
CW Jr and Lotz MK: Human chondrocyte apoptosis in response to
mechanical injury. Osteoarthritis Cartilage. 9:712–719. 2001.
View Article : Google Scholar
|
|
30
|
Tsai CC, Chou YY, Chen YM, Tang YJ, Ho HC
and Chen DY: Effect of the herbal drug guilu erxian jiao on muscle
strength, articular pain, and disability in elderly men with knee
osteoarthritis. Evid Based Complement Alternat Med.
2014:2974582014.PubMed/NCBI
|
|
31
|
Gloth FM III: Pharmacological management
of persistent pain in older persons: focus on opioids and
nonopioids. J Pain. 12(3 Suppl 1): S14–S20. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schnitzer TJ, Hochberg MC, Marrero CE,
Duquesroix B, Frayssinet H and Beekman M: Efficacy and safety of
naproxcinod in patients with osteoarthritis of the knee: a 53-week
prospective randomized multicenter study. Semin Arthritis Rheum.
40:285–297. 2011. View Article : Google Scholar
|
|
33
|
Dostrovsky NR, Towheed TE, Hudson RW and
Anastassiades TP: The effect of glucosamine on glucose metabolism
in humans: a systematic reviewof the literature. Osteoarthritis
Cartilage. 19:375–380. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lai JN, Chen HJ, Chen CC, Lin JH, Hwang JS
and Wang JD: Duhuo jisheng tang for treating osteoarthritis of the
knee: a prospective clinical observation. Chin Med. 2:42007.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen FP, Chang CM, Hwang SJ, Chen YC and
Chen FJ: Chinese herbal prescriptions for osteoarthritis in Taiwan:
analysis of National Health Insurance dataset. BMC Complement
Altern Med. 14:912014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Moudgil KD and Berman BM: Traditional
Chinese medicine: potential for clinical treatment of rheumatoid
arthritis. Expert Rev Clin Immunol. 10:819–822. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zheng CS, Ye HZ, Li XH, Xu HF, Wu GW and
Liu XX: Study on the active components groups of BUSHENZHUANGJIN
decoction for osteoarthritis by molecular docking method. J Tradit
Chin Orthop Traumatol. 24:8–10. 2012.In Chinese.
|
|
38
|
Guo FJ, Xiong Z, Lu X, Ye M, Han X and
Jiang R: ATF6 upregulates XBP1S and inhibits ER stress-mediated
apoptosis in osteoarthritis cartilage. Cell Signal. 26:332–342.
2014. View Article : Google Scholar
|
|
39
|
Bull VH and Thiede B: Proteome analysis of
tunicamycin-induced ER stress. Electrophoresis. 33:1814–1823. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Koyama M, Furuhashi M, Ishimura S, Mita T,
Fuseya T, Okazaki Y, Yoshida H, Tsuchihashi K and Miura T:
Reduction of endoplasmic reticulum stress by 4-phenylbutyric acid
prevents the development of hypoxia-induced pulmonary arterial
hypertension. Am J Physiol Heart Circ Physiol. 306:H1314–H1323.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mosmann T: Rapid colorimetric assay for
cellular growth and survival: application to proliferation and
cytotoxicity assays. J Immunol Methods. 65:55–63. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang G, Gurtu V, Kain SR and Yan G: Early
detection of apoptosis using a fluorescent conjugate of Annexin V.
Biotechniques. 23:525–531. 1997.PubMed/NCBI
|
|
43
|
Dapat E, Jacinto S and Efferth T: A
phenolic ester from Aglaia loheri leaves reveals cytotoxicity
towards sensitive and multidrug-resistant cancer cells. BMC
Complement Altern Med. 13:2862013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Acton BM, Jurisicova A, Jurisica I and
Casper RF: Alterations in mitochondrial membrane potential during
preimplantation stages of mouse and human embryo development. Mol
Hum Reprod. 10:23–32. 2004. View Article : Google Scholar
|
|
45
|
Zheng C, Ye Z, Li X, Xu H, Wu G and Liu X:
Study on diversity and drug-like property of compounds in Bushen
Zhuangjin decoction based on computer simulation. Fujian Zhongyi
Xueyuan Xuebao. 22:22–25. 2012.In Chinese.
|
|
46
|
Chevet E, Hetz C and Samali A: Endoplasmic
reticulum stress-activated cell reprogramming in oncogenesis.
Cancer Discov. 5:586–597. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Puthalakath H, O'Reilly LA, Gunn P, Lee L,
Kelly PN, Huntington ND, Hughes PD, Michalak EM, McKimm-Breschkin
J, Motoyama N, et al: ER stress triggers apoptosis by activating
BH3-only protein Bim. Cell. 129:1337–1349. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fu HY, Okada K, Liao Y, Tsukamoto O,
Isomura T, Asai M, Sawada T, Okuda K, Asano Y, Sanada S, et al:
Ablation of C/EBP homologous protein attenuates endoplasmic
reticulum-mediated apoptosis and cardiac dysfunction induced by
pressure overload. Circulation. 122:361–369. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
B'chir W, Maurin AC, Carraro V, Averous J,
Jousse C, Muranishi Y, Parry L, Stepien G, Fafournoux P and Bruhat
A: The eIF2α/ATF4 pathway is essential for stress-induced autophagy
gene expression. Nucleic Acids Res. 41:7683–7699. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Creagh EM: Caspase crosstalk: integration
of apoptotic and innate immune signalling pathways. Trends Immunol.
35:631–640. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Allan LA and Clarke PR: Apoptosis and
autophagy: Regulation of caspase-9 by phosphorylation. FEBS J.
276:6063–6073. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jänicke RU, Sprengart ML, Wati MR and
Porter AG: Caspase-3 is required for DNA fragmentation and
morphological changes associated with apoptosis. J Biol Chem.
273:9357–9360. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Sharif M, Whitehouse A, Sharman P, Perry M
and Adams M: Increased apoptosis in human osteoarthritic cartilage
corresponds to reduced cell density and expression of caspase-3.
Arthritis Rheum. 50:507–515. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu F, Liu G, Liang W, Ye H, Weng X, Lin
P, Li H, Chen J, Liu X and Li X: Duhuo Jisheng decoction treatment
inhibits the sodium nitroprussiate-induced apoptosis of
chondrocytes through the mitochondrial-dependent signaling pathway.
Int J Mol Med. 34:1573–1580. 2014.PubMed/NCBI
|
|
55
|
Hildeman DA, Zhu Y, Mitchell TC, Kappler J
and Marrack P: Molecular mechanisms of activated T cell death in
vivo. Curr Opin Immunol. 14:354–359. 2002. View Article : Google Scholar : PubMed/NCBI
|