Introduction
The occurrence of angiostenosis resulting from
delayed reendothelialization and intimal hyperplasia (IH) following
endovascular interventions often leads to failure of operation and
remains a challenge to patients and surgeons (1). However, when the vascular integrity,
particularly the endangium, is disrupted, a delay in
reendothelialization and IH is inevitable. Therefore, insufficient
or decelerated reendothelialization and IH are considered the two
main elements that retard vascular wound healing, form atheromatous
plaque and increase in-stent restenosis rates (2). However, once restenosis and thrombus
have occurred, a few treatment options may be utilized and often,
the therapeutic effect of these approaches becomes weak. Although
the newly designed drug-eluting stents have reduced the incidence
of restenosis, they are inadequate to increase reendothelialization
(3,4). Thus, effective drug-eluting stents
with fewer side effects continue to be designed and tested for the
treatment of more complex and severe arterial disease. Although
vascular injury problems are the main focus of the present study,
only a few interventional measures or effective agents that can
promote reendothelialization and ameliorate IH after vascular
injury have been developed thus far.
Curcumin (Cur), the major active component of
curcuminoids, is a natural extract found in the rhizomes of
turmeric of the Zingiberaceae family. Cur has long been used in
China as a traditional medicine for various ailments (5). It possesses beneficial
pharmacological effects, including antioxidant, antiproliferative
(6,7), anti-inflammatory, anti-mutagenic and
anti-tumorigenic properties in in vitro and in vivo
models (8–10). Previous findings have shown that
Cur treatment following subarachnoid hemorrhage in rat
cerebrovascular disease models reduced the mortality rate and
improved functional and histologic outcomes. Mounting evidence has
demonstrated the potential therapeutic effect of Cur on acute
ischemic stroke, possibly through mitochondrial protection,
inhibition of the cell death pathway and improvement in regional
cerebral blood flow (10–12). Cur also acts as an inhibitor and
regulator of inflammation, restraining the
lipopolysaccharide-induced overexpression of inflammatory cytokines
in vascular smooth muscle cells by disrupting the TLR4-MAPK
pathways and intracellular reactive oxygen species (ROS) production
(13). In the peripheral vascular
system, Cur may induce angiogenesis and accelerate wound healing in
acute phase diabetic rats through the increased expression of
vascular endothelial growth factors (14). In the cardiovascular system, the
beneficial effects of Cur included reduced hypertension, improved
vascular dysfunction and alleviated oxidative stress (5). Accumulating evidence has
demonstrated that Cur regulates and ameliorates mechanical vascular
dysfunction. However, the protective effect of Cur following
vascular intimal injury, as well as the relevant mechanism
associated with the ECs autophagy improvement remains to be
investigated.
The abovementioned studies have demonstrated that
Cur protects the nervous and vascular systems. However, the effects
of Cur on accelerating reendothelialization and ameliorating IH
following arterial intimal injury and the underlying mechanisms
involved remain unknown. The aim of the present study was to
elaborate the intimate relationship between IH and autophagy.
Simultaneously, the application of Cur to an arterial intimal
injury model confirmed its effects on accelerating
reendothelialization and ameliorating IH by promoting EC
autophagy.
Materials and methods
Animals
Experiments were approved by the Ethics Committee of
the First Affiliated Hospital of Soochow University and were
performed in accordance with the guidelines of the National
Institutes of Health on the care and use of animals. Adult male
Sprague-Dawley (SD) rats weighing between 270 and 300 g were
purchased from the Animal Center of Chinese Academy of Sciences
(Shanghai, China). The rats were housed in temperature- and
humidity-controlled animal quarters with a 12-h light/dark cycle.
Animal body temperature was maintained at 37°C.
Rat carotid artery balloon injury
model
Subsequent to intraperitoneal anesthesia with
urethane (1,000 mg/kg), an operating microscope was used to
separate the carotid artery (CA). A right CA intima injury was
created according to an approach described in a previous
intervention project (15).
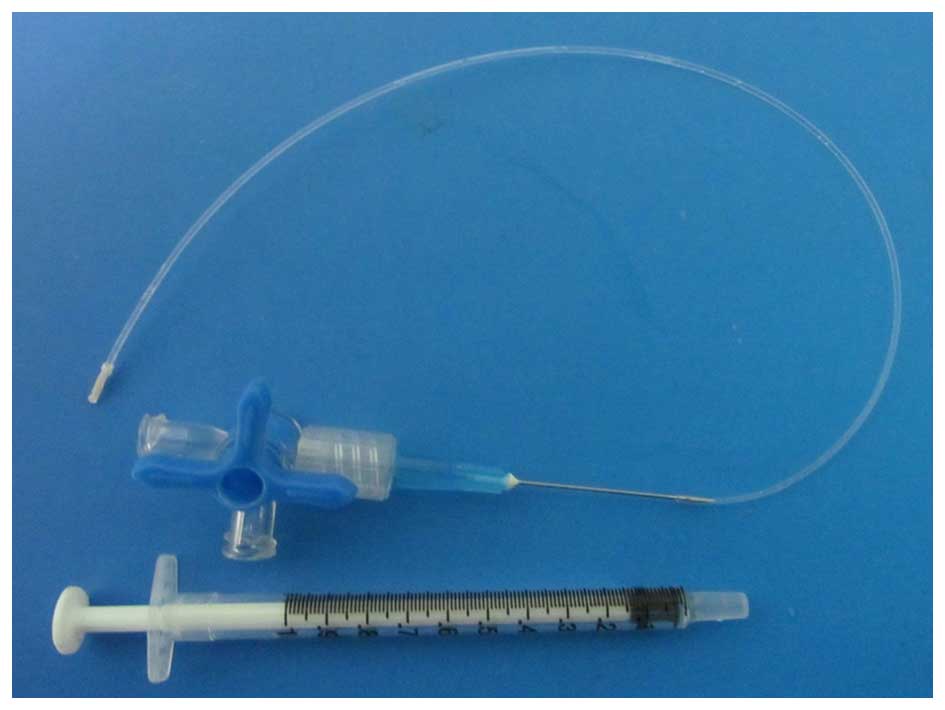
Specific equipment (stereotactic head frame) was designed and
manufactured by the experimental group members. In addition, a
catheter and balloon were purchased from Medtronic Inc. and
manufactured by the authors of this study. A T-branch pipe and 1-ml
syringe were used to modulate the size and pressure of the balloon.
The mode pattern is shown in Fig.
1. Briefly, the catheter was inserted through the external CA
and slipped into the common CA, where it was inflated to ~2 atm and
the inner surface of the common CA was rubbed back and forth three
times. The catheter was then removed and the impaired external CA
was carefully sutured by using a 12-0 proline under the operating
microscope (M651; Leica Microsystems) to prevent the impaired ECA
from developing a ligature. A 2-cm common CA region adjacent to the
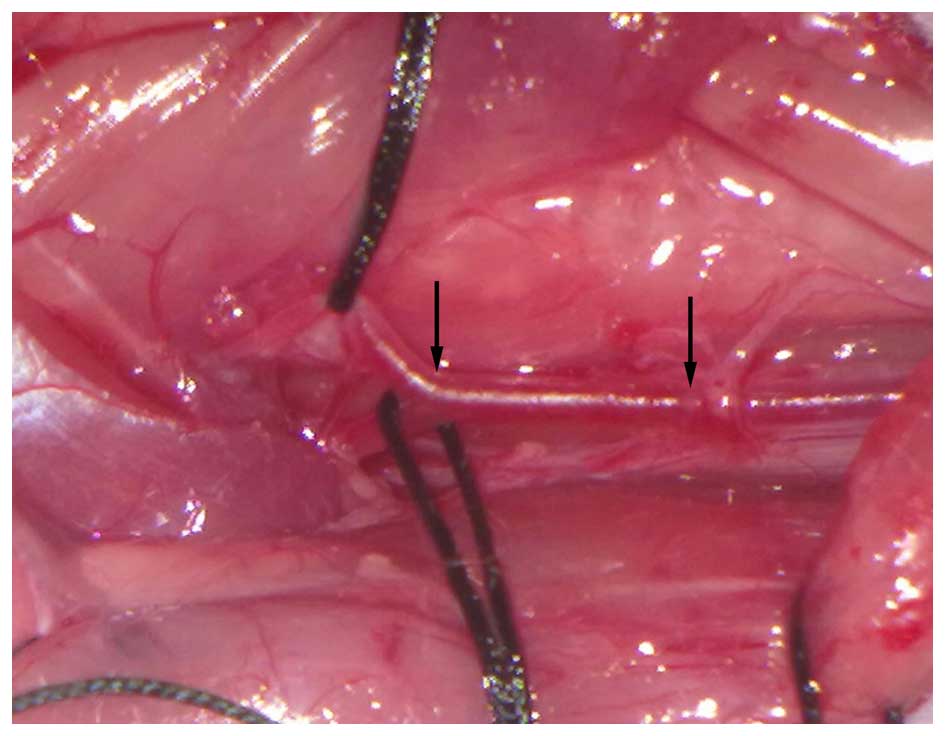
common CA intersection was then cut for this experiment. The vasal
site and obtained segment of the injured CA were marked by two
black arrows (Fig. 2).
Drug administration and animal experiment
design
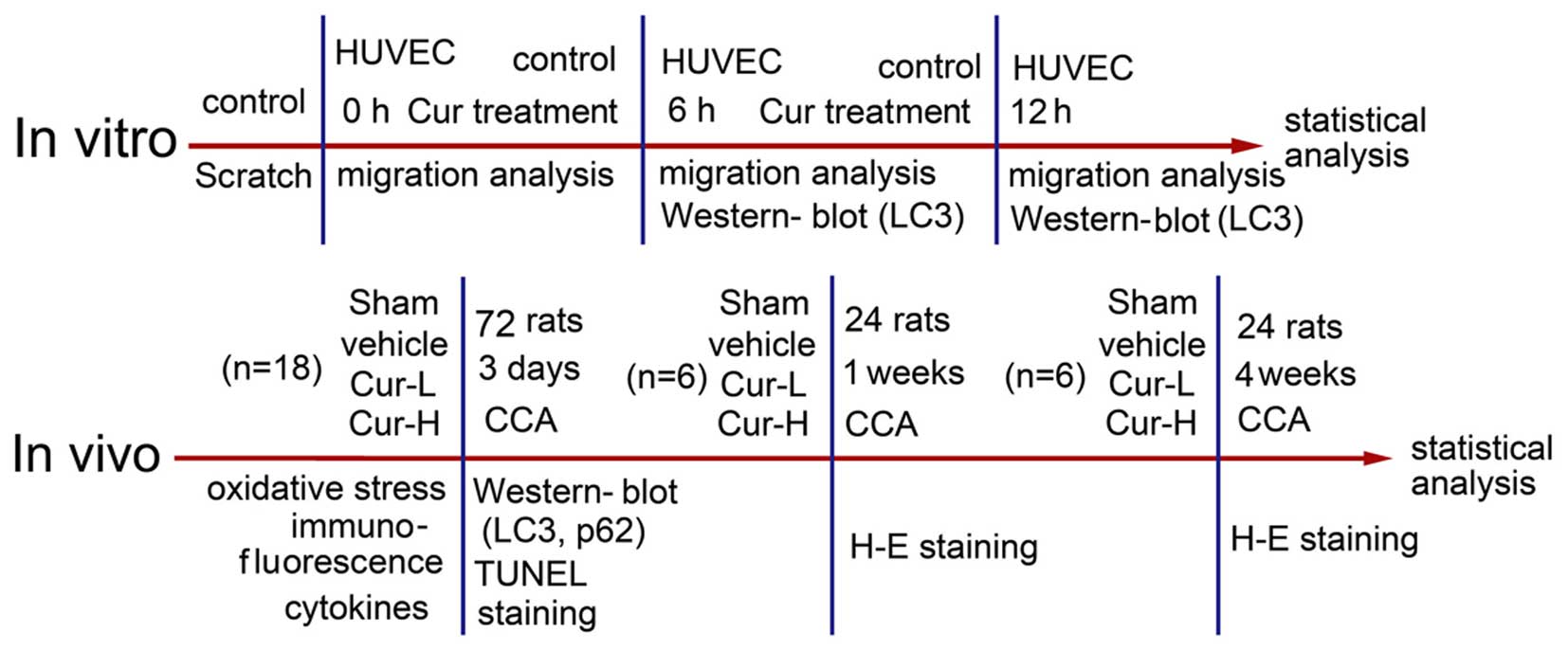
The animal experiment was divided into three
sections based on various time-points. The first section was used
to verify that Cur was able to promote EC autophagy. A total of 90
male SD rats were randomly divided into 4 groups of 18 rats each:
Sham group, injured + vehicle group, injured + Cur low-dose (25
mg/kg) group and injured + Cur high-dose (75 mg/kg) group. Cur or
an equal volume of a vehicle was orally administered at 6 h after
injury and then once daily for the subsequent 3 days. The dose of
Cur administered was based on a previous study (12), thus ensuring that the animals were
able to tolerate the doses without developing significant side
effects. The CAs of six rats from each group were collected and
homogenized in 50 mmol/l of ice-cold potassium phosphate buffer
(PBS, pH 6.8; P1022; Solarbio) and immediately used for the
measurement of oxidative stress indicators. The CAs collected from
another six rats from each group were frozen and homogenized in
RIPA lysis buffer (P0013; Beyotime Institute of Biotechnology,
Jiangsu, China) for western blot analysis. The CAs in the six
remaining rats from each group were collected for terminal
deoxynucleotidyl transferase-mediated dUTP nick end-labeling
(TUNEL) staining, hematoxylin and eosin (H&E) staining and
double immunofluoresence analysis. Twenty-four rats in the second
section that were housed for 2 weeks and another 24 rats housed for
4 weeks from the day of CA injury in the third section were,
respectively, divided into 4 groups and their respective CAs were
collected for histologic examination. The flow chart and the
experimental design are shown in Fig.
3.
Cell culture and treatment
Human umbilical vein endothelial cells (HUVECs) were
obtained and cultured as described previously (16,17). To evaluate the effect of Cur in
vitro, the cells were exposed to Cur at a concentration of 10
µM for 6 and 24 h prior to conducting any subsequent assays
(18). After the treatments,
total protein of the cells was collected and stored at −80°C until
analysis.
Cell migration assay
The EC migration assay was performed as previously
described (19). Briefly, the
cells were seeded in 24-well plates. When the cells reached a
post-confluent state, wounds of 1 mm width were created by scraping
the cell monolayers with a sterile pipette tip (P3361; Seebio). The
cells were treated with or without Cur, and incubated at 37°C for 6
and 12 h. Migration was documented by capturing images immediately
after scraping, as well as 6 and 12 h later at a magnification of
×100. Cell migration was quantified by measuring the recovered area
using the NIH Image Program.
Antibodies and reagents
Cur was extracted from Curcuma longa (Bailingwei
Technology Co., Ltd., Beijing, China, SB-431542). The rabbit
polyclonal antibody against LC3-A/B (ab58610) and the mouse
monoclonal antibody against p62 (610833) were purchased from BD
Biosciences (Palo Alto, CA, USA) and the mouse monoclonal antibody
against GAPDH (sc-365062) was from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). The FOX3-vWF rabbit polyclonal and mouse
monoclonal antibody-endothelial cell marker (ab6994 and ab68545)
were from Abcam (New Territories, Shatin, Hong Kong). Secondary
antibodies for western blot analysis, which included goat
anti-rabbit IgG-HRP (sc-2004) and goat anti-mouse IgG-HRP (sc-2005)
were purchased from Santa Cruz Biotechnology, Inc.. Normal rabbit
IgG (sc-2027) was from Santa Cruz Biotechnology, Inc. Secondary
antibodies for immunofluorescence, including Alexa Fluor-488 donkey
anti-rabbit IgG antibody (A21206) and Alexa Fluor-555 donkey
anti-mouse IgG antibody (A31570) were from Invitrogen-Life
Technologies (Carlsbad, CA, USA).
Western blot analysis
Western blot analysis was performed as described
previously (20,21). The CAs were ground in a cell lysis
buffer (P0013; Beyotime Institute of Biotechnology) using a mortar.
The whole lysate was centrifuged at 200 x g for 5 min at 4°C twice.
The supernatant was subsequently extracted and the protein
concentration was measured using an Enhanced BCA protein assay kit
(Beyotime Institute of Biotechnology). Protein samples (10
µg/lane) were loaded on a 12% sodium dodecyl
sulfate-polyacrylamide gel (P0012A; Beyotime Institute of
Biotechnology), separated and electrophoretically transferred to a
polyvinylidene difluoride (PVDF) membrane (Millipore Corporation,
Billerica, MA, USA), which was blocked with 5% bovine serum albumin
(BSA; BioSharp, Hefei, AH, China) for 1 h at room temperature. The
membrane was then incubated overnight at 4°C with primary
antibodies. The primary antibodies were applied at a dilution of
1:1,000. The membrane was incubated with the corresponding
HRP-conjugated secondary antibodies, which were diluted to 1:5,000
for 2 h at room temperature. The signal was developed using an
enhanced chemiluminescence (ECL) kit (Beyotime Institute of
Biotechnology) and exposed to X-ray film. The films were scanned
using an Epson Perfection 2480 scanner (Seiko Corp., Nagano,
Japan). The relative quantity of proteins was analyzed based on the
densitometric analysis using the Image J program and normalized to
that of the loading controls. The protein levels of GAPDH served as
loading controls. The densitometry ratio of the target protein to
the loading control was evaluated and analyzed as the relative
protein level of target protein.
Immunofluorescence microscopy
The common CA was carefully excised, fixed and
embedded in paraffin, cut into 4-µm sections and examined by
immunofluorescence staining. As previously described (20), we performed double-labeling of
microtubule-associated protein light chain 3 (LC-3) and P62/von
Willebrand Factor (vWF). The primary antibodies were applied at a
dilution of 1:100 and the secondary antibodies were diluted to
1:300. The normal and corresponding negative controls were also
included in the experiment. These methods have been previously
investigated and applied by our experimental group, the
fluorescence images were captured and the relative fluorescence
intensity of the hybridizations was analyzed as described in our
previous study (17).
Oxidative stress indicator assay
The concentrations and activitives of oxidative
stress indicators in the impaired CAs were measured by using
specific detection kits (Nanjing Jiancheng Bioengineering
Institute, Jiangsu, China), as per the manufacturer's instructions,
and as previously described (20).
TUNEL staining and quantification of
apoptotic ECs
TUNEL-positive cells exhibit DNA damage, indicating
apoptotic cell death (22).
Apoptosis of ECs in the intimal wall was detected by TUNEL
staining, following the manufacturer's protocol (DeadEnd
Fluorometric kit; Promega, Madison, WI, USA) and as described in a
previous study (20). To identify
vascular EC apoptosis, the sections were visualized on a
fluorescence microscope. To assess the extent of EC apoptosis, six
microscopic fields were examined and photographed parallel to
TUNEL-positive cell counting.
H&E staining and luminal area
measurement
The vessel tissue processing method was conducted
and the degree of neointima formation was evaluated by measuring
the luminal cross-sectional areas (23,24).
Statistical analyses
Data were presented as the means ± SEM SPSS 11.5
(SPSS Inc., Chicago, IL, USA) was used for statistical analysis.
Data were analyzed by one-way ANOVA (followed by Scheffé F-test for
post hoc analysis). P<0.05 was considered statistically
significant.
Results
Cur accelerated reendothelialization and
migration of HUVECs
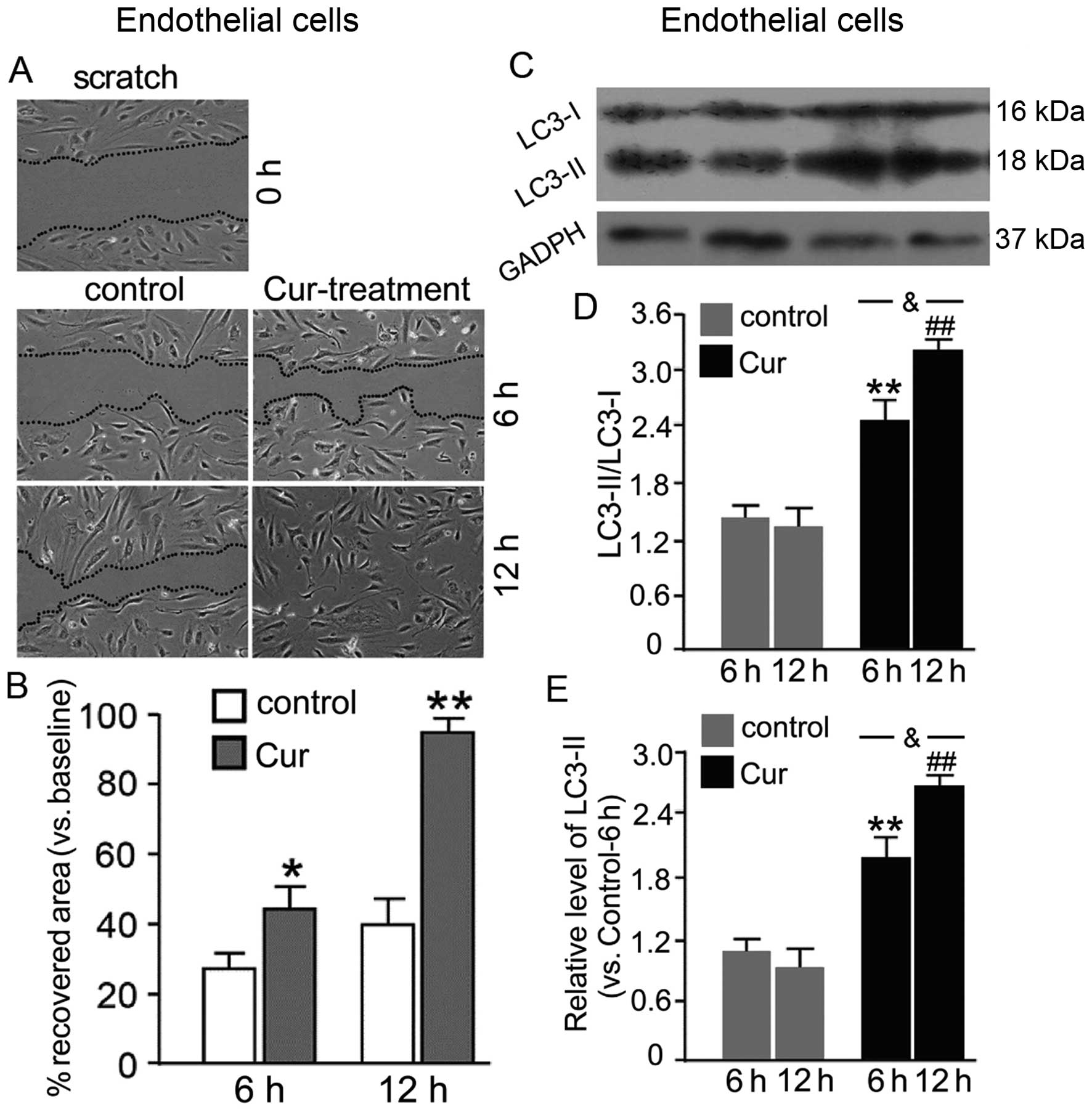
As shown in Fig. 4A
and B, a comparison of recovered areas showed that Cur
accelerated the closure of an artificial wound at 6 and 12 h after
experimental induction of the wound compared to the control
(P<0.05 and P<0.01, respectively). Simultaneously, as shown
in Fig. 4C–E, wherein treated
control-6 h was used as a standard, Cur increased the expression of
LC3-II and the ratio of LC3-II/LC3-I when HUVECs were exposed to
Cur, indicating that Cur induced autophagy in CAs at 6 and 12 h
(P<0.05 and P<0.01, respectively). Moreover, the expression
of LC3-II and the ratio of LC3-II/LC3-I were upregulated following
12 h compared to that observed in the 6 h group (P<0.05).
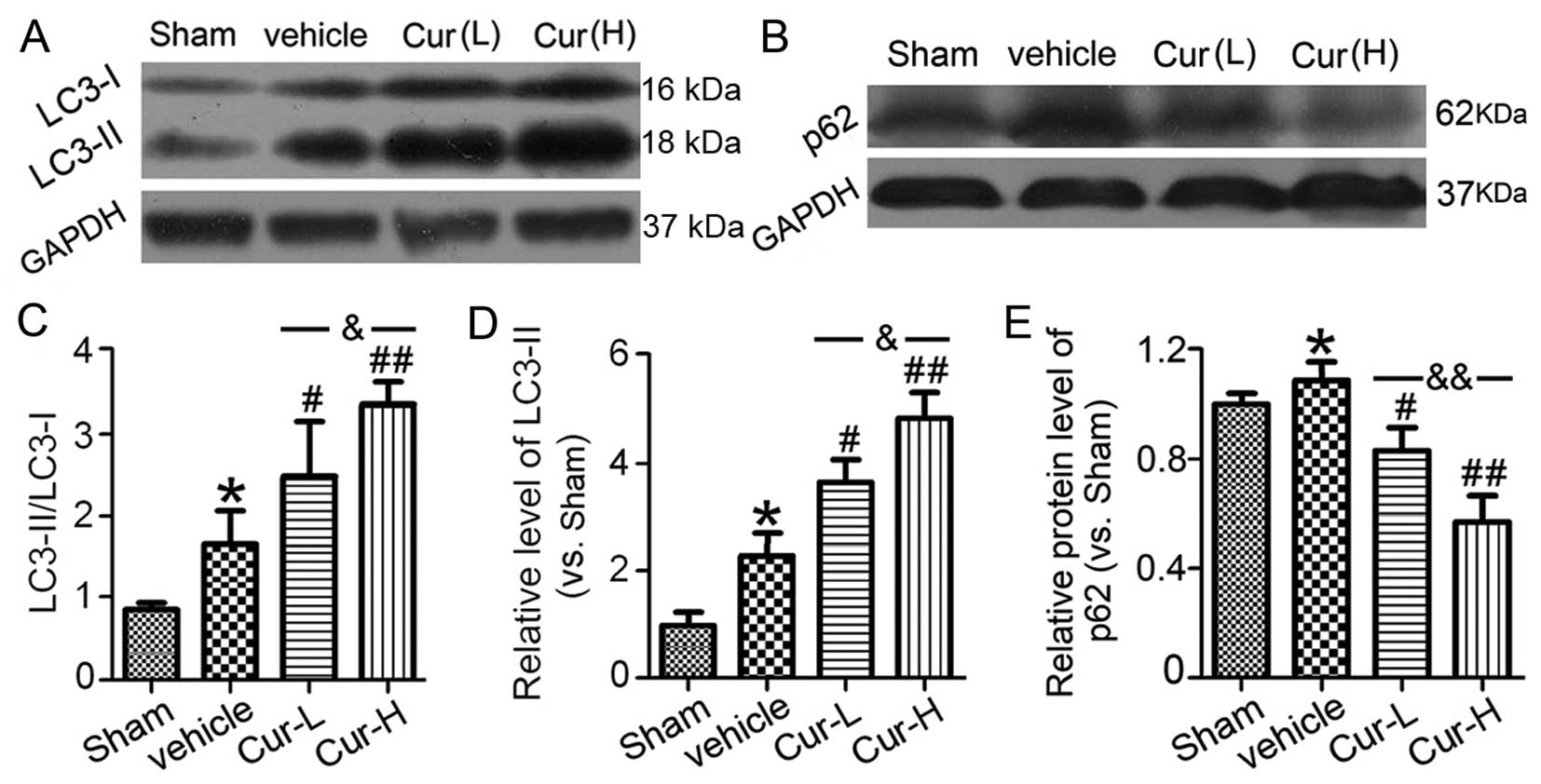
Cur treatments enhance autophagy in
vascular ECs in vitro and in vivo
To investigate the autophagy-promoting role of Cur,
the autophagic levels of HUVECs from various treatment groups were
evaluated by detecting LC3-II accumulation and calculating for the
ratio of LC-I/LC3-II in western blot analysis (Fig. 5). The LC3-II level and the ratio
of LC3-II/LC3-I slightly increased compared to that observed in the
sham group (P<0.05). Cur upregulated LC3-II and the ratio of
LC3-II/LC3-I, indicating that Cur induced autophagy in CAs
(P<0.05 and P<0.01, respectively). However, Cur treatment
also markedly suppressed p62 expression (P<0.05 and P<0.01,
respectively), whose level was negatively correlated with the
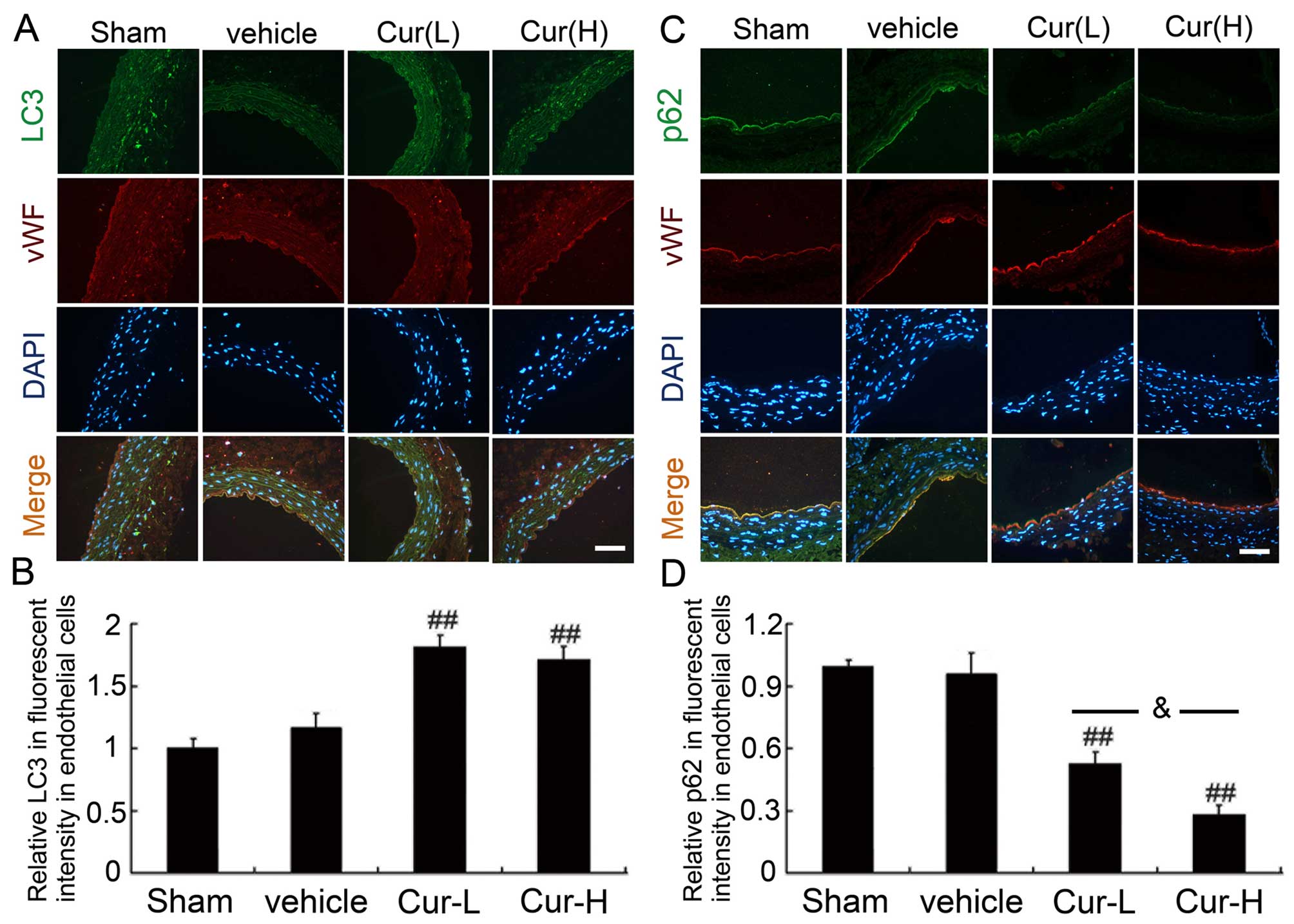
autophagic standard. Accordingly, to distinguish the cell identity
for LC3 and p62, double-immunofluorescence staining with vWF was
performed on vascular sections attached to different groups
(Fig. 6). A less diffuse cell
staining of LC3-I was observed in the sham group, whereas a more
punctate staining for LC3-II was observed in the injured group
compared to that observed in the sham group. However, Cur treatment
resulted in a significant increase in punctate staining compared to
that observed in the sham and injured groups (P<0.01). An
increase in p62 fluorescence intensity was detected after vascular
injury, which may have been restrained by Cur (P<0.01).
Nonetheless, the high dose of Cur apparently added an extra effect
to low-dose Cur (P<0.05).
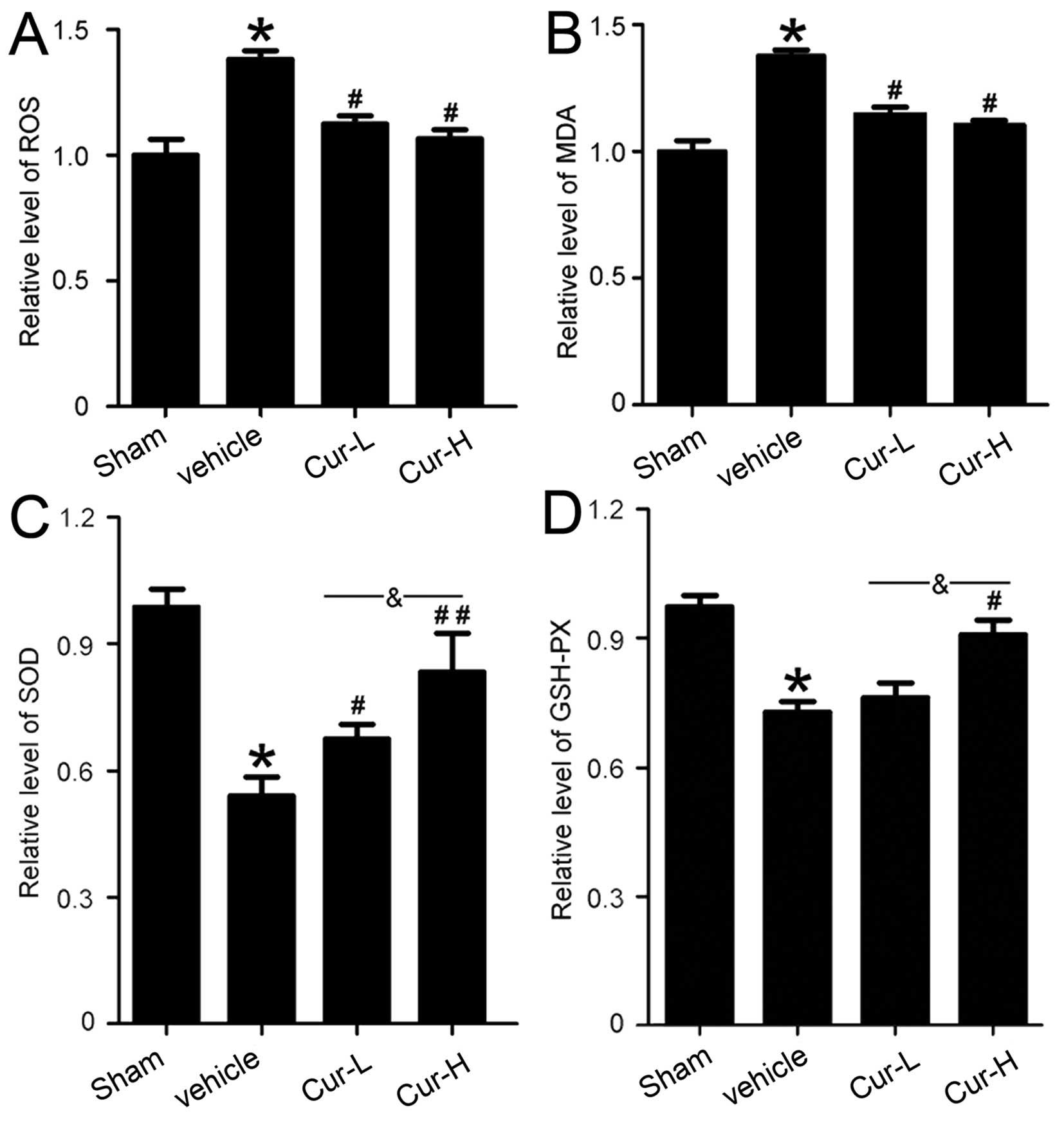
Inhibitory effects of Cur on balloon
injury-induced oxidative stress
To assess the effect of Cur on oxidative stress
following CA intimal injury, we detected the levels of ROS and MDA
and the activities of GSH-Px and SOD in rat CA tissues. As shown in
Fig. 7, compared to the sham
group, the levels of ROS (P<0.05) and MDA (P<0.05) were
significantly elevated, whereas that of GSH-Px (P<0.01) and SOD
(P<0.05) decreased. However, Cur treatment restrained the
elevation of the levels of ROS (P<0.05) and MDA (P<0.05) and
reversed the reduction of SOD (P<0.05 and P<0.01,
respectively) and GSH-Px (P<0.05) activities following balloon
injury. These findings showed that high-dose Cur suppressed the
downregulation of SOD (P<0.05) and GSH-Px (P<0.05)
activities. These results are suggestive of the antioxidant action
of Cur.
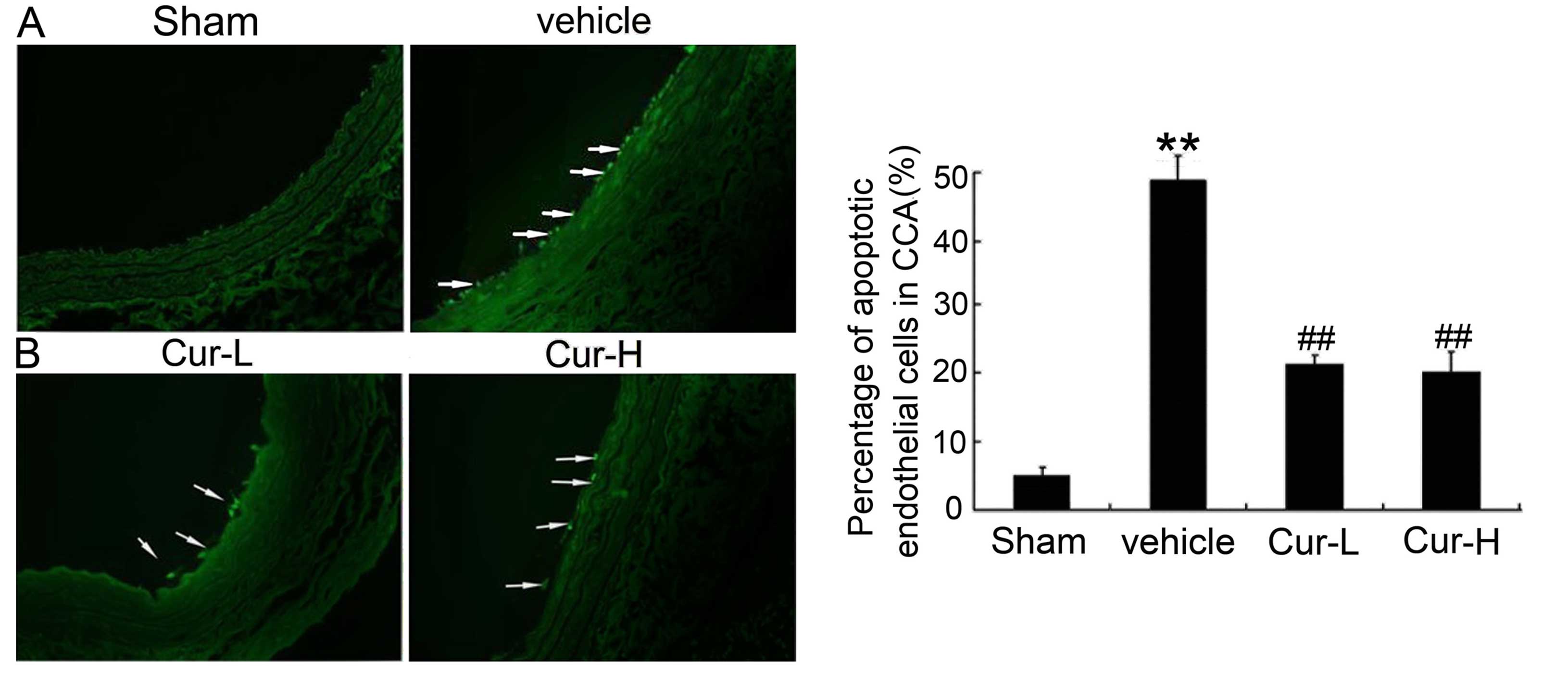
Cur inhibits intimal injury-induced
vascular EC apoptosis
As shown in Fig.
8A, only a few TUNEL-positive apoptotic cells were identified
in the endothelium of CA in the sham group, whereas the apoptotic
index was significantly higher in the injured group (P<0.01).
Compared to the injured group, low-dosage Cur or high-dosage Cur
imparted a obvious protection effect on vascular EC apoptosis
(P<0.01).
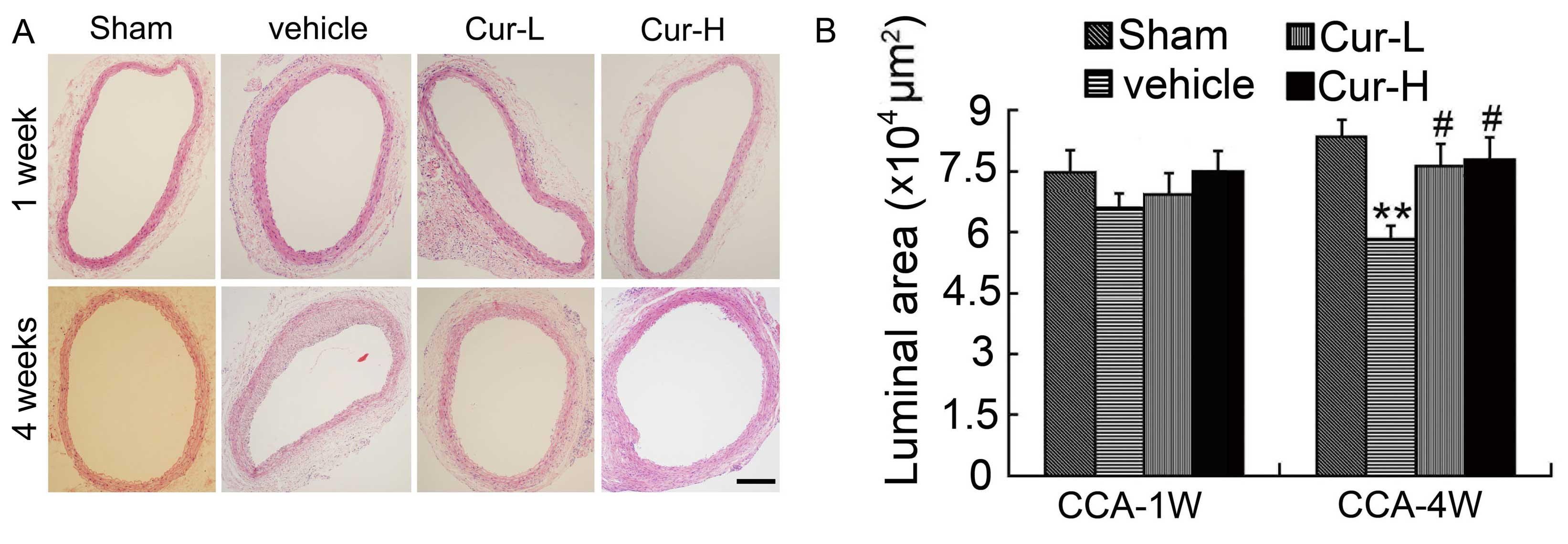
Oral administration of Cur reverses
established IH
A significant hemadostenosis or plaque was not
detected in the injured and treatment groups at 1 week after
injury, wherein the neointima remained fairly stable in size
(Fig. 9A). The rats subjected to
balloon injury of the CA after 4 weeks developed a reproducible IH
response (Fig. 9A). A significant
difference in the luminal area between the injured and Cur
treatment groups was observed (Fig.
9B). In rats that were treated with daily Cur treatments for 4
weeks, the luminal area of CA was significantly reduced compared to
that observed in the sham vessels (P<0.01). Cur-treated CA
showed a significant suppression of IH formation compared to that
observed in the injured group (P<0.05). However, no amelioration
of neointima size and thickness was observed in the high-dose Cur
group compared to that observed in the low-dose Cur group.
Discussion
Delayed reendothelialization and IH inevitably
occurred on account of arterial endothelium denudation after
intimal injury and played an important role in preventing in-stent
thrombosis and vessel restenosis (1,2,4).
The results of the present study have shown that the oral
administration of Cur accelerated EC migration and ameliorated IH
in the injured CA, which involved an increase in EC autophagy. The
present results also showed that i) Cur promotes EC migration by
enhancing the autophagy effect, which was confirmed by the
upregulation of LC3-II and the downregulation of p62. ii) Cur
markedly ameliorates IH on the injured CA after 4 weeks of
induction by balloon compression, as indicated in the comparison of
the extent of hemadostenosis and the expression of LC3 and p62 by
western blot analysis and immunofluorescence staining.
EC dysfunction is an initial event in arterial
hypertension and atherosclerosis and plays a crucial role in the
regulation of CA disease (25,26). In addition, the endothelium
produces nitric oxide (NO) for vasorelaxation and controls
leukocyte adherence, which in turn regulates vascular permeability.
The Cur effects of reduced hypertension, improved vascular
dysfunction and alleviated oxidative stress in mice may be
attributable to the increase in NO bioavailability and the powerful
antioxidant activity of Cur (27,28). EC autophagy plays a pivotal role
in regulating vascular homeostasis, and OxyLDL leads to apoptotic
cell death, which is dependent on LOX-1 overexpression and the
subsequent attenuation of the protective autophagic response
(29).
Autophagy is associated with nutrient deprivation
and a cellular mechanism of autophagy, in which proteins and
organelles are encased in specialized intracellular vesicles and
then broken down by lysosomal proteases for recycling. During times
of stress such as nutrient starvation or treatment with certain
toxic agents, the rate of autophagy is increased (30,31). LC3 is a cytoplasmic protein that
is rapidly cleaved to produce LC3-I and is activated by lipidation
with phosphatidylethanolamine to form LC3-II by a ubiquitin-like
ligation pathway. LC3-II is incorporated into the vesicle membrane
and promotes membrane elongation, a variation of LC3-II that may
actually be an indication of the stimulation of autophagy (32–34). P62, as a multidomain protein that
contains different kinds of interaction domains such as the
LC3-interacting region, is involved in the degradation of protein
aggregates and cytoplasmic bodies via selective autophagy. Thus,
p62 was used to detect autophagy flow (35). In addition, various methods have
demonstrated the existence of autophagy in human atherosclerotic
plaques, which may in turn contribute to the development of
pharmacological approaches to stabilize vulnerable and
rupture-prone lesions (36). The
role of autophagy in neointima remodeling following CA mechanical
injury has been previously investigated (15). However, the potential correlation
between vascular ECs autophagy was not discussed intensively,
although we have demonstrated the vital effect of ECs autophagy on
increasing EC migration and proliferation, which coincided with the
in vivo experimental results that the application of Cur, as
a type of antioxidant and antiproliferative agent, accelerated
reendothelialization and suppressed IH. Thus, EC autophagy may be
important in the development of neointimal lesions, where rapid
cell proliferation may lead to extensive nutrient depletion.
In terms of effective intervening measures and
agents, there is currently an increasing number of studies that
have explored the inhibitory effect for IH. For instance, the
periadventitial application of rapamycin-loaded nanoparticles
resulted in the sustained inhibition of vascular restenosis, which
may be used as an approach for the development of a safer, more
efficacious drug delivery system for the treatment of IH (37). Local application of insulin
directly onto the CA decreases neointimal thickness without
affecting glycemia and apparently reduces smooth muscle cell
migration after arterial injury. These results provide evidence of
an insulin-coated stent that may offer protection against
restenosis (38). Furthermore,
delayed intermittent and repetitive inhaled carbon monoxide
resulted in a significant reduction in neointima size through the
inhibition of intimal and medial smooth muscle cell proliferation
(15). Although Cur was equipped
with various protective benefits involving the regulation of
proinflammatory cytokines, growth factors, factors involved in
proliferation and apoptosis, and adhesion molecules, it was applied
to different animal models, including the model of atherosclerosis
and cerebrovascular disease. However, relevant studies on the
application of Cur in an intimal injury model have not investigated
this effect (39–41).
The present study has a number of limitations.
Firstly, the concrete autophagy signaling pathways remain to be
adequately elucidated. Future studies should focus on the
underlying formation mechanism behind the effect of Cur and
analogous drugs on relieving vascular injury caused by a balloon.
Secondly, use of a special inhibitor and stimulator of autophagy
may facilitate in understanding the complex formation mechanism and
in the development of a novel drug for the treatment of vascular
intimal injury. Consequently, some drugs that target EC autophagy
potentially have great therapeutic prospects for the treatment of
vascular intimal injury after interventional surgery.
In conclusion, the application of Cur imparts
beneficial effects by increasing the proliferation and migration of
ECs by enhancing its autophagy level. Simultaneously, Cur
suppresses balloon-induced IH by promoting the autophagy effect.
Cur may be used as an anti-inflammatory and pro-autophagy agent and
therefore serves as a novel treatment for vascular intimal injury
induced by interventional therapies.
Acknowledgments
The present study was supported by grants from the
National Natural Science Foundation of China (nos. 81371279,
81422013, 81471196 and 81400949), the Jiangsu Province's
Outstanding Medical Academic Leader program (no. LJ201139), the
Scientific Department of Jiangsu Province (no. BL2014045) and the
Suzhou Government (nos. LCZX201301, SZS201413 and SYS201332) and a
Project Funded by the Priority Academic Program Development of
Jiangsu Higher Education Institutions.
References
|
1
|
Seedial SM, Ghosh S, Saunders RS,
Suwanabol PA, Shi X, Liu B and Kent KC: Local drug delivery to
prevent restenosis. J Vasc Surg. 57:1403–1414. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Acharjee S and Cannon CP: Duration of dual
antiplatelet therapy following percutaneous coronary intervention
with drug-eluting stents: A review of recent evidence. Crit Pathw
Cardiol. 9:203–206. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stettler C, Wandel S, Allemann S, Kastrati
A, Morice MC, Schömig A, Pfisterer ME, Stone GW, Leon MB, de Lezo
JS, et al: Outcomes associated with drug-eluting and bare-metal
stents: A collaborative network meta-analysis. Lancet. 370:937–948.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Buonamici P, Marcucci R, Migliorini A,
Gensini GF, Santini A, Paniccia R, Moschi G, Gori AM, Abbate R and
Antoniucci D: Impact of platelet reactivity after clopidogrel
administration on drug-eluting stent thrombosis. J Am Coll Cardiol.
49:2312–2317. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kukongviriyapan U, Pannangpetch P,
Kukongviriyapan V, Donpunha W, Sompamit K and Surawattanawan P:
Curcumin protects against cadmium-induced vascular dysfunction,
hypertension and tissue cadmium accumulation in mice. Nutrients.
6:1194–1208. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dutta S, Padhye S, Priyadarsini KI and
Newton C: Antioxidant and antiproliferative activity of curcumin
semicarbazone. Bioorg Med Chem Lett. 15:2738–2744. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Weber WM, Hunsaker LA, Abcouwer SF, Deck
LM and Vander Jagt DL: Anti-oxidant activities of curcumin and
related enones. Bioorg Med Chem. 13:3811–3820. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lim CS, Jin DQ, Mok H, Oh SJ, Lee JU,
Hwang JK, Ha I and Han JS: Antioxidant and antiinflammatory
activities of xanthorrhizol in hippocampal neurons and primary
cultured microglia. J Neurosci Res. 82:831–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Biswas SK, McClure D, Jimenez LA, Megson
IL and Rahman I: Curcumin induces glutathione biosynthesis and
inhibits NF-kappaB activation and interleukin-8 release in alveolar
epithelial cells: Mechanism of free radical scavenging activity.
Antioxid Redox Signal. 7:32–41. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Z, Dabrosin C, Yin X, Fuster MM,
Arreola A, Rathmell WK, Generali D, Nagaraju GP, El-Rayes B,
Ribatti D, et al: Broad targeting of angiogenesis for cancer
prevention and therapy. Semin Cancer Biol. S1044-579X(15)00002-4.
2015. View Article : Google Scholar
|
|
11
|
Kuo CP, Lu CH, Wen LL, Cherng CH, Wong CS,
Borel CO, Ju DT, Chen CM and Wu CT: Neuroprotective effect of
curcumin in an experimental rat model of subarachnoid hemorrhage.
Anesthesiology. 115:1229–1238. 2011.PubMed/NCBI
|
|
12
|
Li S, Wu C, Zhu L, Gao J, Fang J, Li D, Fu
M, Liang R, Wang L, Cheng M, et al: By improving regional cortical
blood flow, attenuating mitochondrial dysfunction and sequential
apoptosis galangin acts as a potential neuroprotective agent after
acute ischemic stroke. Molecules. 17:13403–13423. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meng Z, Yan C, Deng Q, Gao DF and Niu XL:
Curcumin inhibits LPS-induced inflammation in rat vascular smooth
muscle cells in vitro via ROS-relative TLR4-MAPK/NF-κB pathways.
Acta Pharmacol Sin. 34:901–911. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kant V, Gopal A and Kumar D, Pathak NN,
Ram M, Jangir BL, Tandan SK and Kumar D: Curcumin-induced
angiogenesis hastens wound healing in diabetic rats. J Surg Res.
193:978–988. 2015. View Article : Google Scholar
|
|
15
|
Madigan M, Entabi F, Zuckerbraun B,
Loughran P and Tzeng E: Delayed inhaled carbon monoxide mediates
the regression of established neointimal lesions. J Vasc Surg.
61:1026–1033. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pacifici M and Peruzzi F: Isolation and
culture of rat embryonic neural cells: A quick protocol. J Vis Exp.
(63): e39652012.PubMed/NCBI
|
|
17
|
Li H, Huang S, Wang S, Zhao J, Su L, Zhao
B, Zhang Y, Zhang S and Miao J: Targeting annexin A7 by a small
molecule suppressed the activity of phosphatidylcholine-specific
phospholipase C in vascular endothelial cells and inhibited
atherosclerosis in apoli-poprotein E−/− mice. Cell Death
Dis. 4:e8062013. View Article : Google Scholar
|
|
18
|
Wang YF, Gu YT, Qin GH, Zhong L and Meng
YN: Curcumin ameliorates the permeability of the blood-brain
barrier during hypoxia by upregulating heme oxygenase-1 expression
in brain microvascular endothelial cells. J Mol Neurosci.
51:344–351. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dong Z, Cheng Y, Zhao J, Su L, Zhao B,
Zhang Y, Zhang S and Miao J: Discovery of a benzoxazine derivative
promoting angiogenesis in vitro and in vivo. J Cell Physiol.
223:202–208. 2010.PubMed/NCBI
|
|
20
|
Wang Y, Gao A, Xu X, Dang B, You W, Li H,
Yu Z and Chen G: The Neuroprotection of Lysosomotropic Agents in
Experimental Subarachnoid Hemorrhage Probably Involving the
Apoptosis Pathway Triggering by Cathepsins via Chelating
Intralysosomal Iron. Mol Neurobiol (Aug). 12:2014.
|
|
21
|
Li H, Gao A, Feng D, Wang Y, Zhang L, Cui
Y, Li B, Wang Z and Chen G: Evaluation of the protective potential
of brain microvascular endothelial cell autophagy on blood-brain
barrier integrity during experimental cerebral ischemia-reperfusion
injury. Transl Stroke Res. 5:618–626. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schlunk F, Schulz E, Lauer A, Yigitkanli
K, Pfeilschifter W, Steinmetz H, Lo EH and Foerch C: Warfarin
pretreatment reduces cell death and MMP-9 activity in experimental
intrace-rebral hemorrhage. Transl Stroke Res. 6:133–139. 2015.
View Article : Google Scholar
|
|
23
|
Wang Z, Chen G, Zhu WW, Bian JY, Shen XO
and Zhou D: Influence of simvastatin on microthrombosis in the
brain after subarachnoid hemorrhage in rats: A preliminary study.
Ann Clin Lab Sci. 40:32–42. 2010.PubMed/NCBI
|
|
24
|
Zhang JM, Wang Y, Miao YJ, Zhang Y, Wu YN,
Jia LX, Qi YF and Du J: Knockout of CD8 delays reendothelialization
and accelerates neointima formation in injured arteries of mouse
via TNF-α inhibiting the endothelial cells migration. PLoS One.
8:e620012013. View Article : Google Scholar
|
|
25
|
Sitia S, Tomasoni L, Atzeni F, Ambrosio G,
Cordiano C, Catapano A, Tramontana S, Perticone F, Naccarato P,
Camici P, et al: From endothelial dysfunction to atherosclerosis.
Autoimmun Rev. 9:830–834. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Andoh J, Sawyer B, Szewczyk K, Nortley M,
Rossetti T, Loftus IM, Yáñez-Muñoz RJ and Hainsworth AH: Transgene
delivery to endothelial cultures derived from porcine carotid
artery ex vivo. Transl Stroke Res. 4:507–514. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tesfamariam B and DeFelice AF: Endothelial
injury in the initiation and progression of vascular disorders.
Vascul Pharmacol. 46:229–237. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lindner V, Fingerle J and Reidy MA: Mouse
model of arterial injury. Circ Res. 73:792–796. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mollace V, Gliozzi M, Musolino V, Carresi
C, Muscoli S, M1-145 ollace R, Tavernese A, Gratteri S, Palma E,
Morabito C, et al: Oxidized LDL attenuates protective autophagy and
induces apoptotic cell death of endothelial cells: Role of
oxidative stress and LOX-1 receptor expression. Int J Cardiol.
184:152–158. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lalaoui N, Lindqvist LM, Sandow JJ and
Ekert PG: The molecular relationships between apoptosis, autophagy
and necroptosis. Semin Cell Dev Biol. 39:63–69. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luo T, Park Y, Sun X, Liu C and Hu B:
Protein misfolding, aggregation, and autophagy after brain
ischemia. Transl Stroke Res. 4:581–588. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Matsui Y, Takagi H, Qu X, Abdellatif M,
Sakoda H, Asano T, Levine B and Sadoshima J: Distinct roles of
autophagy in the heart during ischemia and reperfusion: Roles of
AMP-activated protein kinase and Beclin 1 in mediating autophagy.
Circ Res. 100:914–922. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mizushima N and Yoshimori T: How to
interpret LC3 immuno-blotting. Autophagy. 3:542–545. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Martinet W and De Meyer GR: Autophagy in
atherosclerosis: A cell survival and death phenomenon with
therapeutic potential. Circ Res. 104:304–317. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lin X, Li S, Zhao Y, Ma X, Zhang K, He X
and Wang Z: Interaction domains of p62: A bridge between p62 and
selective autophagy. DNA Cell Biol. 32:220–227. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu H, Cao Y, Tong T, Shi J, Zhang Y, Yang
Y and Liu C: Autophagy in atherosclerosis: A phenomenon found in
human carotid atherosclerotic plaques. Chin Med J (Engl).
128:69–74. 2015. View Article : Google Scholar
|
|
37
|
Shi X, Chen G, Guo LW, Si Y, Zhu M, Pilla
S, Liu B, Gong S and Kent KC: Periadventitial application of
rapamycin-loaded nanoparticles produces sustained inhibition of
vascular restenosis. PLoS One. 9:e892272014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chiang S, Breen DM, Guo J, Mori Y and
Giacca A: Local insulin application on the carotid artery inhibits
neointima formation. Can J Physiol Pharmacol. 91:1086–1094. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Masuda T, Hidaka K, Shinohara A, Maekawa
T, Takeda Y and Yamaguchi H: Chemical studies on antioxidant
mechanism of curcuminoid: Analysis of radical reaction products
from curcumin. J Agric Food Chem. 47:71–77. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sikora E, Scapagnini G and Barbagallo M:
Curcumin, inflammation, ageing and age-related diseases. Immun
Ageing. 7(1)2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lapchak PA and McKim JM Jr: CeeTox™
Analysis of CNB-001 a Novel Curcumin-Based
Neurotrophic/Neuroprotective Lead Compound to Treat Stroke:
Comparison with NXY-059 and Radicut. Transl Stroke Res. 2:51–59.
2011. View Article : Google Scholar : PubMed/NCBI
|