Introduction
In cartilage, the extracellular matrix (ECM)
occupies 95% of the space which plays a dominant role in
chondrocyte functions. ECM is composed of a large number of
different molecules, which mediate cell-matrix and matrix-matrix
communications, thereby determining the histoarchitecture specific
to each organ, and provide cells with crucial information on
migration, adhesion and differentiation (1–4).
There are three classes of ECM molecules, which include filamentous
collagen molecules, proteoglycans and non-filamentous collagen
molecules (1–4). Matrilins are a family of four
filamentous-forming adapter oligomeric extracellular proteins that
are involved in the development and homeostasis of cartilage and
bone (1,5–7).
They can form homo- and hetero-oligomers through the assembly of
C-terminal coiled-coil structures (1,5–7).
As regards their distribution, it has been demonstrated that
matrilin-1 and -3 are restricted to cartilage and bone, whereas
matrilin-2 and -4 are present in a broader range of tissues
(1,2,8,9).
Matrilins are thought to play a role in the ECM by acting as
bridges which connect matrix components and the cell membrane in
the cartilage to form macromolecular networks (10,11). In cartilage, matrilins interact
with type II and type IV collagen, and aggrecan, as well as the
cartilage oligomer matrix protein (COMP) (1,10,11). Matrilins consist of one or two von
Willebrand factor, type A (vWFA) domains, a varying number of
epidermal growth factor (EGF) repeats, and a C-terminal coiled-coil
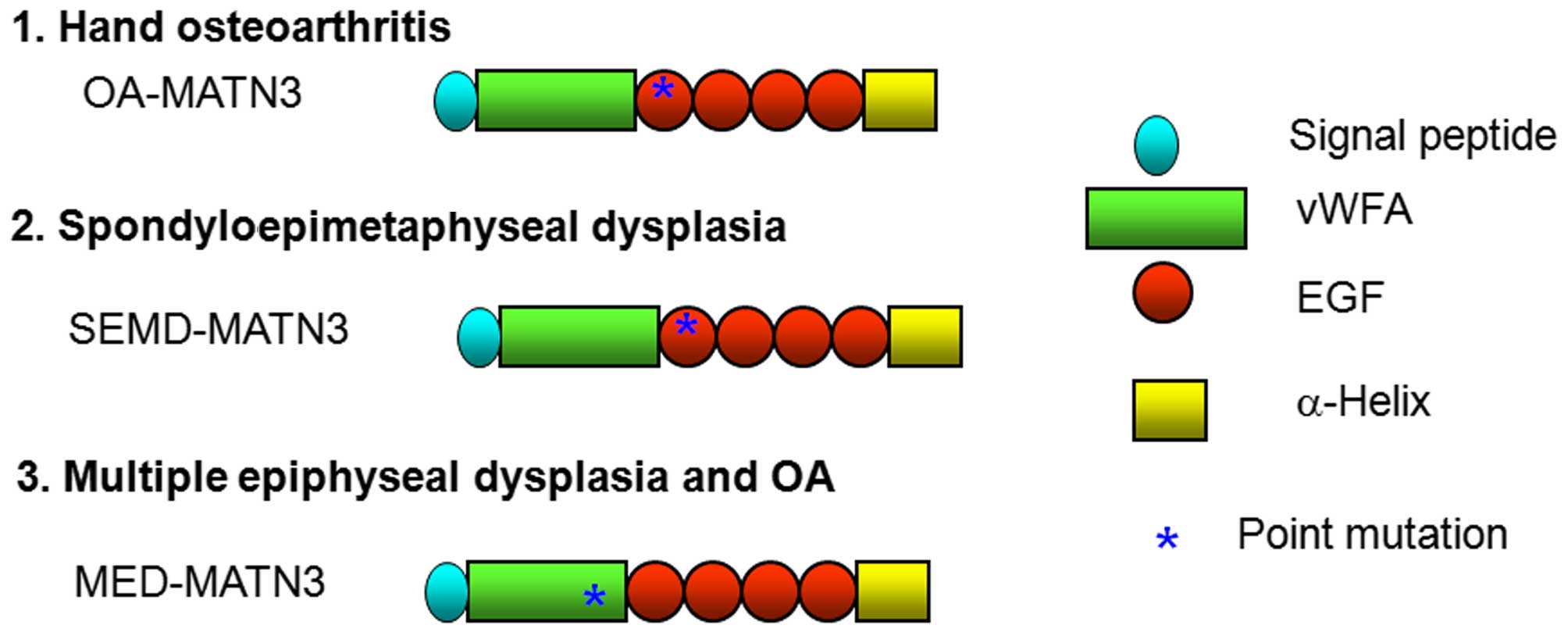
domain (1–3). The polypeptide of matrilin-3
predicted from the nucleotide sequence of this clone shared 83%
identity with matrilin-3 from mouse and 61% with that from chicken
(5,12,13). It was composed of 486 amino acid
residues that were arranged in 7 domains: a signal peptide, a vWFA
domain, four EGF repeats and an α-helical region. The gene for
human matrilin-3 was assigned to chromosome region 2. The
corresponding mRNA of 2.8 kb has been found to be expressed in
every type of cartilage investigated thus far (5,12,13).
The vWFA domain is found in various collagenous and
non-collagenous ECM proteins, and is composed of a central β-sheet
core flanked by α-helices (5,10,11). In humans, at least nine missense
point mutations in the matrilin-3 gene that affect the vWFA domain
(typically the β-sheets) have been found in patients with multiple
epiphyseal dysplasia (MED), which is characterized by delayed and
irregular ossification of the epiphyses and early onset
osteoarthritis (OA) (14–20). Mutations of matrilin-3 have also
been reported in other osteochondrodysplasias, including bilateral
hereditary microepiphyseal dysplasia, which is an MED-like disorder
characterized by small epiphyses in the hip and knee joints
(14–20).
EGF domain-containing molecules have also been
identified ubiquitously in the ECM, cell membrane and nucleus and
similar to vWFA domain-containing molecules, they participate in
cell adhesion, protein-protein interactions and the formation of
multiprotein complexes (21,22). Mutations in the first EGF domain
of matrilin-3 have also been reported in osteochondrodysplasias,
such as spondyloepimetaphyseal dysplasia (SEMD), which includes a
number of conditions associated with vertebral, epiphyseal and
metaphyseal anomalies (23) and
idiopathic hand osteoarthritis (HOA) (24).
Matrilin-3 has also been reported to be upregulated
in cartilage from patients with OA, and a strong correlation
between enhanced matrilin-3 gene and protein expression and the
extent of tissue damage has been noted (5,12,25). Matrilin-3 is also produced in
vitro by primary chondrocytes isolated from articular
cartilage. However, dedifferentiated chondrocytes of the third
passage do not express matrilin-3 at all (12). Therefore, it has been suggested
that matrilin-3 serves as a marker for the differentiation state of
chondrocytes (5,12,13,25). These findings suggest that the
tight regulation of matrilin-3 expression is essential for the
maintenance of the cartilage ECM microenvironment.
In mice, matrilin-3 is expressed exclusively in the
developing cartilage of the skeletal system, with its expression
first being detected on embryonic day 12.5 and remains unaltered
from embryonic day 15.5 until birth. In newborn mice, matrilin-3 is
widely expressed in the cartilage of the developing bones of the
eyes, nasal cavity, ribs, long bones, sternum and trachea, although
its expression is more restricted by 6 weeks of age (5,12,13). Although it has been shown that
matrilin-1 and -3 can form hetero-oligomers and are often
co-localized in tissue, clear differences in their spatial
distribution have been shown by double immunolabeling (5,12,13). In addition, although matrilin-1
expression continues to be present in tissues that remain
cartilaginous throughout life (such as in costal cartilage and in
the nasal septum), matrilin-3 expression ceases to be present after
birth in these tissues.
Materials and methods
Extraction of RNA from animal tissues,
reverse transcription-polymerase chain reaction (RT-PCR) and
preparation of wild-type and point mutation of matrilin-3 gene
constructs
A total of 3 newborn C57BL/6 mice (Charles River
Laboratories, Wilmington, MA, USA) were sacrificed by
CO2 asphyxiation and the rib cages were harvested. All
procedures were approved by the Ethics Committee of Qinghai
Provincial People's Hospital. Tissue was homogenized, and total RNA
was isolated using an RNeasy kit (Qiagen, Germantown, MD, USA).
First-strand cDNA was synthesized using a SuperScript II kit
(Invitrogen, Grand Island, NY, USA) according to the manufacturer's
instructions. Wild-type matrilin-3 cDNA was cloned from the newborn
C57BL/6 mouse rib library. Specific primers (Table I) introduced a 5′-terminal
BamH1 and a 3′-terminal HindIII restriction enzyme
site into the expression vector, pcDNA3.1/v5-His (Invitrogen).
Point mutations of R116W, T298M and C299S, according to the mouse
sequence, associated with MED, HOA and SEMD, respectively, were
linked by overlap extension PCR with the described primers
(Fig. 1 and Table I) from the wild-type matrilin-3
plasmid. Recombinant sequences were confirmed by DNA
sequencing.
 | Table IPrimers used in the present
study. |
Table I
Primers used in the present
study.
| Primers | Primer sequences
(5′→3′) | PCR purpose |
|---|
| 1 |
TAATACGACCTATAGGG | T7, amplifying
inserts from pCDNA3.1 |
| 2 |
CTAGAAGGCAACAGTCGAGG | BGH, amplifying
inserts from pCDNA3.1 |
| 3 |
CCCAAGCTTATGTTGCTCTCAGCCCCCTTA | Clone matrilin-3
with HindIII |
| 4 |
CCGGATCCTTAACGATGTACTTGTCCAT | Clone matrilin-3
with BamH1 |
| 5 |
ATGGGAAAATGTGTTCAGCCATTGATA | T298M mutation |
| 6 |
CAATGGCTGAACACATTTTCCCATCAGCGT | T298M mutation |
| 7 |
GGAAAACGTCTTCAGCCATTGATAAGT | C299S mutation |
| 8 |
CAATGGCTGAAGACGTTTTCCCATCAG | C299S mutation |
| 9 |
ATCTGCGTTAGAGCCACAACAAGCAGT | R116W mutation |
| 10 |
AAAGAACAACTTGGGTTCATT | R116W mutation |
Transfection of plasmid DNA
Plasmids containing wild-type, MED, SEMD, HOA and
both SEMD plus HOA-point mutations of matrilin-3 were transfected
into the COS-1 or MCT (chondrocytes) cells (kind gift from Dr Lei
Wei, Brown University, Rhode Island Hospital, Providence, RI, USA)
using Lipofectamine 2000 (Invitrogen Life Technologies, Carlsbad,
CA, USA) according to the manufacturer's instructions. Briefly, the
COS-1 or MCT cells were trypsinized and counted. Each 60-mm plate
was seeded with 6×105 cells overnight until the cells
reached 70% confluence in Dulbecco's modified Eagle's medium (DMEM)
supplemented with 10% fetal bovine serum (FBS; Invitrogen Life
Technologies). On the following day, the cells were rinsed with
DMEM 3 times, and then transfected with the plasmid/Lipofectamine
2000 mix, as previously described (5,13).
We also created a group of untransfected cells (not transfected
with any plasmids) and a group of mock-transfected cells
(transfected with pcDNA3.1 empty vector). The COS-1
cell-conditioned medium was collected for western blot analysis on
days 1 to 5, and the MCT cell-conditioned medium was collected for
western blot analysis on days 1 to 3.
Western blot analysis
Conditioned medium (5 µl) or cell lysates (20
µg) was mixed with 5 µl 2X SDS reducing gel loading
buffer containing 5% β-mercaptoethanol. After being boiled for 10
min, the samples were loaded onto 4–15% gradient gels (Bio-Rad
Laboratories, Hercules, CA, USA). Following electrophoresis, the
proteins were transferred onto immobilon-polyvinylidene difluoride
membranes (Merck Millipore, Billerica, MA, USA). The blots were
blocked with 5% non-fat milk (Bio-Rad Laboratories). Monoclonal
antibody against V5 epitope tag (Invitrogen; dilution 1:5,000) for
conditioned medium, rabbit polyclonal anti-growth arrest DNA
damage-inducible gene 153 (GADD153) antibody (Santa Cruz
Biotechnology, Santa Cruz, CA, USA; SC-575, dilution 1:200) and
monoclonal anti-β actin antibody (Sigma, St. Louis, MO, USA; A5441,
dilution 1:5,000) were used as the primary antibodies. The
secondary antibodies were horseradish peroxidase-conjugated goat
anti-mouse IgG (H+L) (Bio-Rad Laboratories; dilution 1:5,000) or
horseradish peroxidase-conjugated goat anti-rabbit IgG (H+L)
(Bio-Rad Laboratories; dilution 1:5,000). The visualization of
immunoreactive proteins was carried out using ECL western blotting
detection reagents (Amersham Pharmacia Biotech Inc., Piscataway,
NJ, USA) followed by exposure of the membranes to Kodak X-OMAT AR
film. The molecular weights of the immunoreactive proteins were
determined with two sets of protein markers, as previously
described (1,5,13).
Immunofluorescence staining
The COS-1 and MCT cells were seeded onto 8-well
chamber slides, and 1 µg wild-type matrilin-3 or the MED-,
SEMD- and HOA-associated mutations, or SEMD plus HOA-mutant
matrilin-3 cDNA were transfected into the cells in each well using
Lipofectamine 2000 (Invitrogen). Three days following transfection,
the cells in monolayer were fixed with 70% ethanol and 50 mM
glycine for 1 h. Immunofluorescence staining was performed by
incubating the cells with anti-V5 primary antibody (Invitrogen) at
1:1,000 for 2 h, followed by incubation with donkey anti-mouse
rhodamine secondary antibody (The Jackson Laboratory, Bar Harbor,
ME, USA) at 1:200 dilutions in the presence of Hoechst staining
solution (Sigma). The slides were mounted with coverslips in
Gel/Mount.
Electron microscopy
After the MCT cells were seeded onto 60-mm plates, 3
µg wild-type matrilin-3 or SEMD, or SEMD plus HOA-mutant
matrilin-3 cDNA were transfected into the cells using Lipofectamine
2000 (Invitrogen). Three days following transfection, monolayer
cells were trypsinized and centrifuged. The cell pellets were
washed in DMEM 3 times and then fixed in 1.5% glutaraldehyde/1.5%
paraformaldehyde containing 0.05% tannic acid, followed by 1%
OsO4, then rinsed, dehydrated and embedded in Spurrs
epoxy. Ultrathin sections were cut at 60–80 nm, mounted onto
formvar-coated 1×2-mm slot grids, stained with uranyl acetate and
lead citrate, and imaged at 120 kV using a Hitachi H800
transmission electron microscope.
Measurement of expression levels of
matrilin-3 in knee joints of Dunkin-Hartley guinea pigs by
immunohistochemistry
Six 9-month-old Dunkin-Hartley guinea pigs (Charles
River Laboratories) were anesthetized, and the right knee joints
were harvested and decalcified with 10% EDTA solution. All
procedures were approved by the Ethics Committee of Qinghai
Provincial People's Hospital. These were routinely processed and
embedded in paraffin. Longitudinal serial sections of bone at
thicknesses of 5 µm were prepared and collected on
commercially available, positively charged glass slides (Superfrost
Plus; Fisher Scientific, Shanghai, China). The sections were dried
on a hot plate to increase adherence to the slides.
Immunohistochemistry was carried out using the avidin-biotin
complex (ABC) method (Vector Laboratories, Shanghai, China).
Representative sections were de-paraffined and re-hydrated through
conventional methods. Endogenous peroxidase was blocked by treating
the sections with 3% hydrogen peroxide in methanol for 30 min. The
sections were then digested with bovine testicular hyaluronidase,
as previously described (14,15) (4,000 U/ml in PBS; Sigma) for 30
min at 37°C. Non-specific protein binding was blocked by incubation
with 10% normal goat serum. The sections were incubated in a
polyclonal rabbit antibody against our matrilin-3 antibody (1:200)
at 4°C overnight. Thereafter, the sections were treated
sequentially with biotinylated goat anti-rabbit IgG (Vectastain
Elite; Vector Laboratories) and avidin-enzyme complex (Vectastain
Elite; Vector Laboratories). These treatments were followed by
standardized development in 3,3′-diaminobenzidine (DAB; Zymed
Laboratories Inc., South San Francisco, CA, USA). The sections were
counterstained with modified Harris hematoxylin (Fisher
Scientific). In order to investigate the specificity of our peptide
antibody, pre-immunized serum from the same rabbit and 0.01 M PBS
service as a negative control.
Results
Osteochondrodysplasia (C299S)-related
matrilin-3 point mutations lead to secretion insufficiency and
protein trafficking into the cytoplasm of chondrocytes
Proteins are basic functional units of the body.
Matrilin-3 is a non-collagenous extracellular protein. The
secretion of matrilin-3 is dependent on primary structure and
folding (10,11,15). In the present study, the secretion
of matrilin-3 was determined by measuring the amount of protein
expressed in the conditioned medium of the cells at different time
points. From days 1–5, we transfected the cells (with wild-type
matrilin-3 or the MED-, SEMD- and HOA-associated mutations, or SEMD
plus HOA-mutant matrilin-3 cDNA) and conditioned medium from the
cells was then collected on days 1–5 for the COS-1 cells and on
days 1–3 for the MCT cells. The amount of protein was determined by
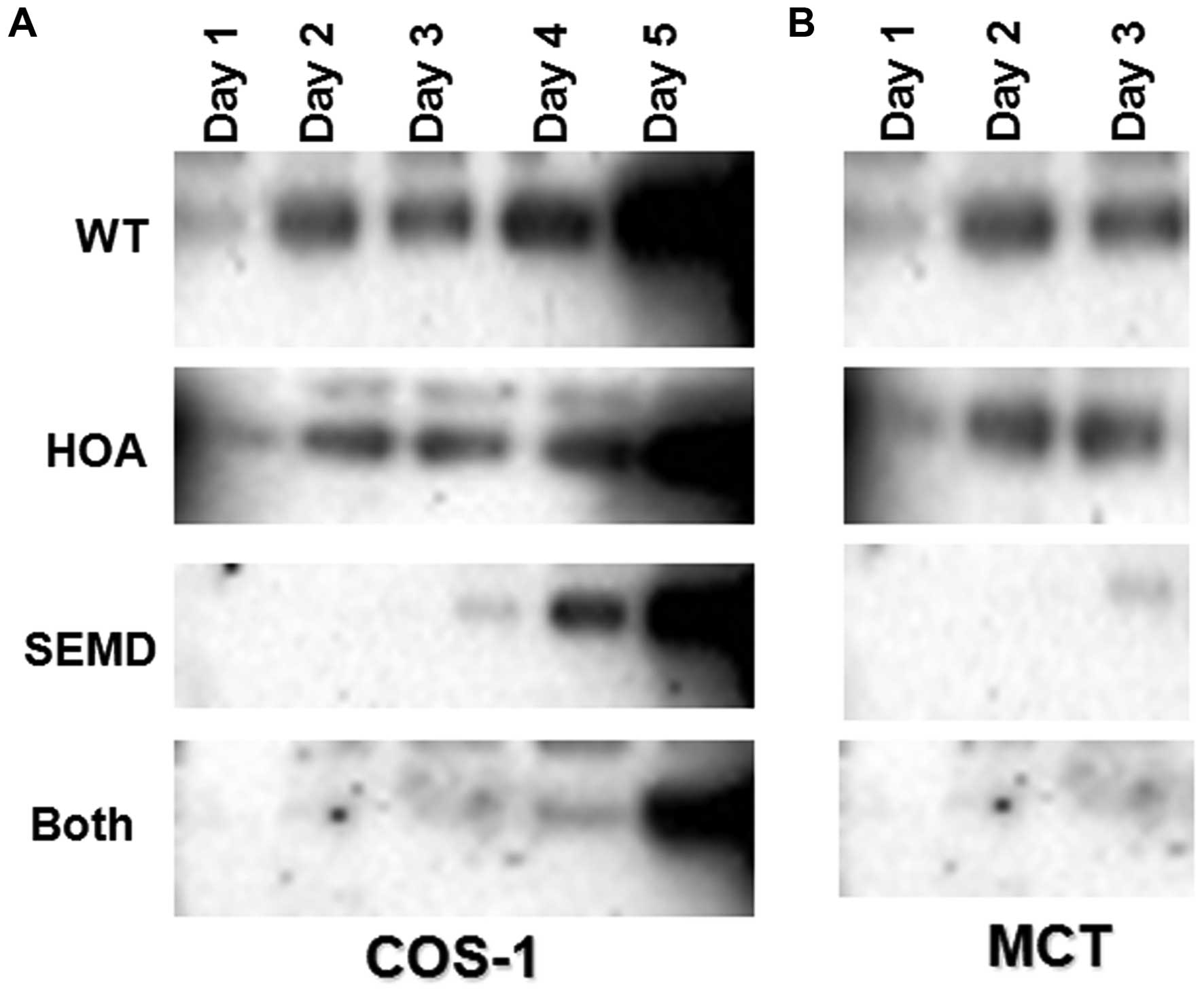
western blot analysis with V5 tag antibody. We found that the
transfection of the COS-1 cells with the SEMD-associated mutation
or SEMD plus HOA-mutant matrilin-3 cDNA (Both) decreased the amount
of protein being secreted. However, transfection with wild-type
matrilin-3 (WT) or HOA-associated mutation did not prevent or
decrease protein secretion into the cell medium (Fig. 2A). We also transfected the same
plasmids into the MCT (chondrocytes) cells, and collected the
medium on days 1 to 3. We found that transfection of the MCT cells
with SEMD-associated mutation or SEMD plus HOA-mutant matrilin-3
cDNA decreased protein secretion; however, transfection with
wild-type matrilin-3 (WT) or HOA-associated mutation did not
prevent protein secretion (Fig.
2B). This result is in line with that obtained with the COS-1
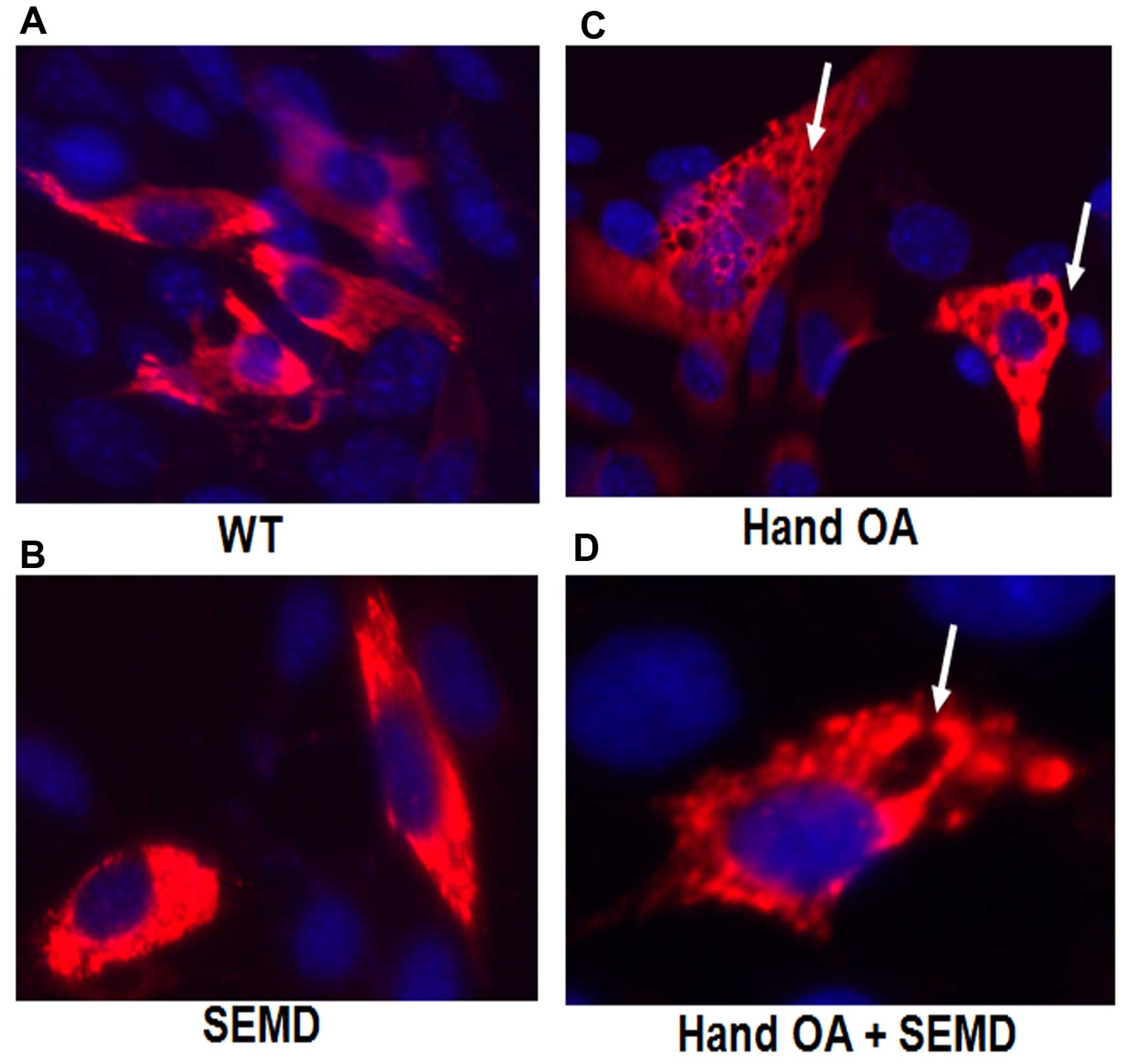
cells. In order to determine the morphological alterations
following transfection with the matrilin-3 plasmids with the
different point mutations, immunofluorescence staining was carried
out. We found that there were numerous vesicles that contained the
SEMD-associated mutation and the SEMD plus HOA-mutant matrilin-3,
whereas by contrast, there were only a few vesicles in the cells
transfected with the wild-type (WT) or HOA-associated mutation
(Fig. 3). The cytoplasm of the
cells transfected with mutant matrilin-3 was greatly enlarged, with
multiple vacuoles. Thus, a point mutation in the cells transfected
with the SEMD-associated mutation or SEMD plus HOA-mutant
matrilin-3 cDNA led to a deficiency in matrilin-3 secretion,
intracellular retention of the mutant protein, and an altered
cytoplasm.
Transfection with mutant matrilin-3 leads
to endoplasmic reticulum (ER) stress
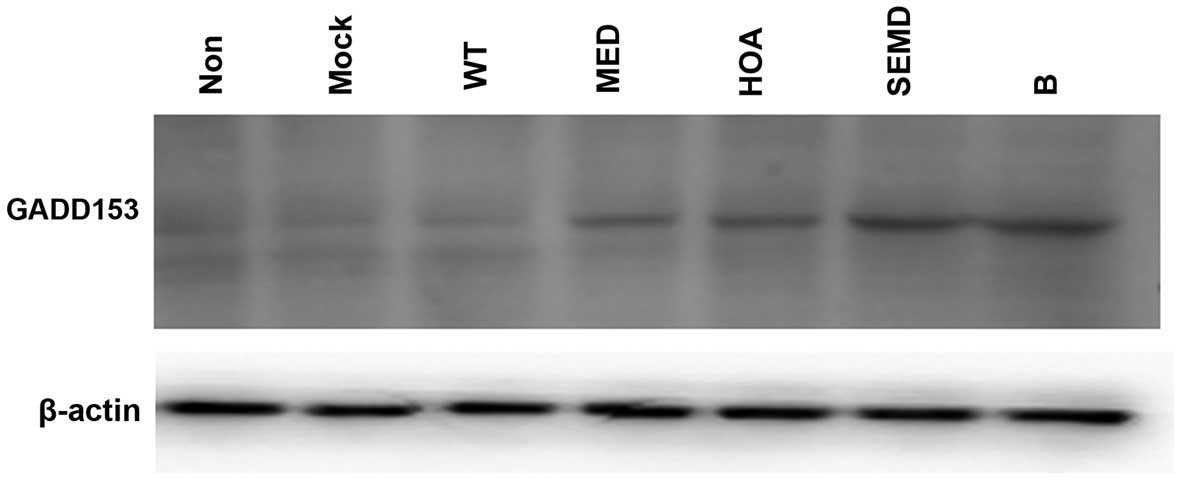
To determine the association between the first
EGF-like domain of matrilin-3 and secretion insufficiencies,
protein trafficking into the cytoplasm and ER stress, the expession
levels of GADD153 (a key molecule in the ER stress pathway) were
measured by western blot analysis. The results revealed that there
was a high level of GADD153 expression in the cells transfected
with the plasmids carrying all of the matrilin-3 point mutations
(MED-, HOA- and SEMD-assoicated mutations or SEMD plus HOA-mutant
matrilin-3 cDNA); however, the expression level of GADD153 was very
low in the untransfected, mock-transfected and wild-type matrilin
3-transfected cells (Fig. 4). It
is important to note that there was a high level of GADD153
expression in the cells transfected with the HOA-assoicated
mutation, which was not indicative of secretion insufficiencies or
protein trafficking into the cytoplasm, as shown by our
above-mentioned experiments (Figs.
2 and 3). This result
indicate although there are some inconsistencies, a higher level of
GADD153 expression may be an earlier marker for bone degenerative
diseases.
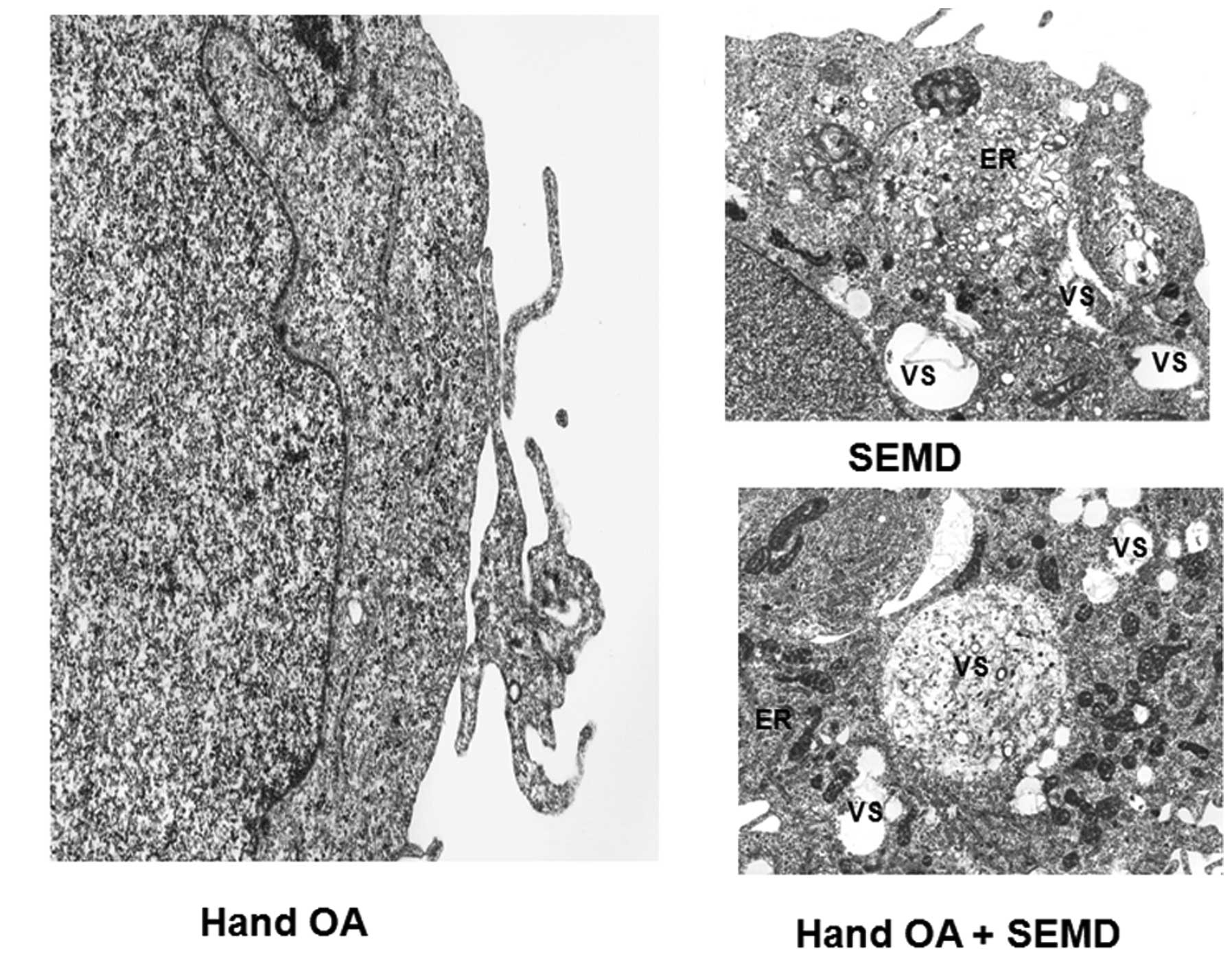
Ultrastructural alterations in cells
transfected with the plasmids carrying matrilin-3 point
mutations
In order to examine the morphology of the COS-1
cells transfected with the plasmids carrying matrilin-3 point
mutations, an electron microscope was employed. There was an
abnormal cytoplasm in the cells transtected with the MED- and
SEMD-associated mutations. The ultrastructural abnormalities
included an expanded ER and numerous vesicles. However, the cells
transfected with the HOA-associated mutation did not exhibit these
abnormalities (Fig. 5).
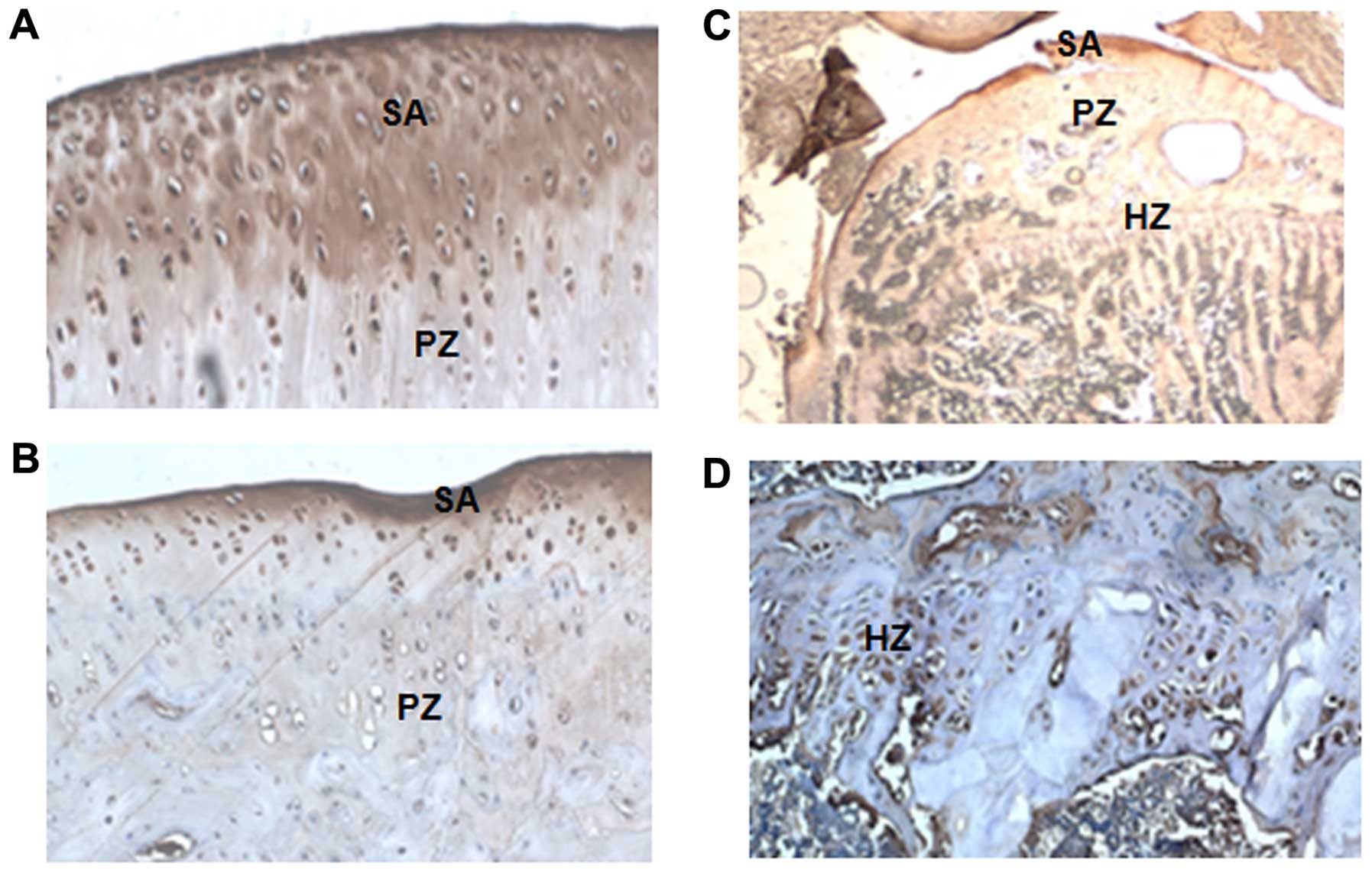
High levels of matrilin-3 expression in
developmental model of OA using Dunkin-Hartley guinea pigs
OA is one of the most common and disabling diseases
affecting the elderly, and almost 80% of individuals of the age of
75 are affected (26,27). OA develops due to wear and tear,
combined with the loss of articular cartilage. It has been
suggested that the degeneration of articular cartilage and
insufficient self-repair are the primary causes of OA (25–27). In the present study, a
developmental animal model of OA with Dunkin-Hartley guinea pigs
was used to investigate matrilin-3 expression. The matrilin-3
expression levels were measured in the knee joints of the guinea
pigs by immunohistochemistry. We found that matrilin-3 was strongly
expressed in the articular cartilage, including the surface area,
and proliferating and hypertrophic areas (Fig. 6).
Discussion
Matrilin-3 is the least complex member of the
matrilin family, and it consists of only one vWFA domain, four EGF
domains, and a C-terminal coiled-coil domain. The vWFA domain is
one of the most widely distributed domains involved in cell
adhesion and the formation of multiprotein complexes (10,11). The property of the vWFA domain in
cell adhesion and protein-protein interaction is mediated, in many
cases, by the metalion dependent adhesion site (MIDAS) located
within the domain (5,13). Mutations at the vWFA domain have
been reported to be related to MED. MED is an
osteochondrodysplasia, primarily characterized by delayed and
irregular ossification of the epiphyses and early-onset OA
(12). Several different
recessive mutations in the exon encoding the vWFA domain of
matrilin-3 cause the EDM5 form of MED (14,16). These point mutations in the vWFA
domain result in single amino acid changes of T120M, R121W, E134K,
I192N, V194D, A219D or E252K. Notably, all of these MED-causing
mutations are located in the β strands in the center of the vWFA
domain, which are important for the folding of the protein
structure (17). Subsequent
genetic analysis has indicated that the R121W mutation is recurrent
in multiple families with common or different ancestries (17). In the present study, we
demonstrated that a single point mutation in the vWFA domain of
mouse matrilin-3 (R116W), equivalent to the MED mutation (R121W) in
human matrilin-3, leads to a deficiency in matrilin secretion and
in the in vitro retention of matrilin-3 within the ER of
primary chondrocytes, which is consistent with the findings of
previous studies (14,28). This point mutation also leads to a
high level of GADD153 expression, which is a key marker of the ER
stress pathway (29–31).
In addition, we cloned the point mutations at the
first EGF domain which are associated with HOA (T298M) and SEMD
(C299S). Our in vitro experiments revealed that there was a
decrease in the amount of mutant protein being secreted into the
medium of the cells transfected with the plasmid carrying the
SEMD-associated mutation (C299S); however, this was not observed in
the cells transfected with the plasmid carrying the HOA-associated
mutation (T298M; secretion was only delayed for 24 h) (Fig. 2). In addition, excessive amounts
of mutant protein had accumulated intracellularly in the cells
transfected with the plasmid carrying the SEMD-associated mutation
(Fig. 3). These observations
indicate that the intracellular retention of mutant protein is
responsible for the deficiency of protein secretion, in terms of
both quantity and speed. Consistent with this hypothesis, we
observed a marked increase in the number of intracellular vesicles
that contained mutant matrilin-3 in the cells transfected with the
plasmid carrying the SEMD-associated mutation, as shown by
immunofluorescence staining and electron microscopy (Figs. 3 and 5).
The EGF domain is composed of approximately 30–40
amino-acid residues and has been found in a large number of mostly
animal proteins; it which is an evolutionary conserved protein
domain, which derives its name from EGF, where it was first
described (21,22). Most EGF-like domain proteins are
found in the extracellular domain of membrane-bound proteins or in
proteins known to be secreted. An exception to this is the
prostaglandin-endoperoxide synthase. The EGF-like domain includes
six cysteine residues which in the epidermal growth factor EGF have
been shown to form three disulfide bonds. The structures of
4-disulfide EGF-domains have been solved from the laminin and
integrin proteins (32,33). In the present study, although the
molecular mechanism of the cysteine mutation of C299S has yet to be
investigated, our results suggest that abnormal protein folding
contributes to the secretion deficiency of the mutant protein. This
abnormal protein folding leads to secretion deficiency, due to
intracellular retention of the mutant proteins which may share a
common mechanism of matrilin-3-associated MED, COMP-associated MED
or related pseudoachondroplasia (34). This retention in turn results in
excessive accumulation of the proteins that are associated with
COMP, such as collagen type IX, whose mutation also leads to
similar clinical manifestations (35). Our observation that cells
expressing mutant matrilin-3 exhibit an expanded cytoplasm with
multiple vacuoles by immunofluorescence staining and electron
microscopy, and our results showing a high level of GADD153 by
western blot analysis, which is similar to the phenotype of mutant
COMP-expressing cells (34,35), suggests that mutated matrilin-3 or
COMP leads to a common cellular phenotype. In light of the
discovery that COMP interacts with matrilin-1, -3 and -4 (36), our findings lend support to the
hypothesis that mutations in any of these interacting proteins
including matrilin, COMP, or collagenIX, result in a secretion
defect, which manifests in common chondrodysplasia pathological
phenotypes. It should also be noted that a portion of the mutant
protein is secreted into the medium. However, we do not know
whether the mutant protein is defective in its adhesion to matrix
ligands or subject to extracellular proteolysis. These
possibilities remain to be determined in future studies.
OA is one of the most common and disabling diseases
in the elderly, affecting nearly 80% of individuals over the age of
75 (25–27). OA develops due to the wear and
tear and the loss of articular cartilage (25–27). It has been previously suggested
that cartilage degeneration and insufficient self-repair are the
primary cause of OA (25–27). In this study, an developmental
animal model of OA with Dunkin-Hartley guinea pigs was used to
measure the matrilin-3 expression levels in the knee joints of the
guinea pigs by immunohistochemistry. The results revealed that
matrilin-3 was strongly expressed in articular cartilage, including
the surface area, and proliferating and hypertrophic areas, and its
expression level was increased in the articular knee joint
cartilage. The results of our examination of the cartilage of the
knee joints indicated heterogeneity. A genome-wide linkage analysis
of patients with idiopathic HOA who were carefully phenotyped for
the involvement of either or both the distal interphalangeal (DIP)
joints and the first carpometacarpal (CMC1) joints has previously
been performed (24). The
missense mutation co-segregates with HOA in several families. The
mutation frequency is slightly >2% in patients with HOA in the
Icelandic population and has a relative risk of 2.1 (24). Previous analysis of recombinant
full-length matrilin-3 revealed that the T298M mutation associated
with HOA does not influence the oligomerization of matrilin-3 or
its proteolytic processing by ADAM metallopeptidase with
thrombospondin type 1 motif (ADAMTS)-4 and -5. Nevertheless,
structural analyses indicate local conformational changes. These
changes do not affect the affinity for collagens II, IX, XI or
COMP, but have a major impact on the in vitro
fibrillogenesis of collagen II/IX/XI heterofibrils (14,28). In the present study, we found the
the T298M HOA -associated mutation does not significantly influence
protein secretion, but leads to high expression levels of GADD153
(Fig. 4). GADD153, which is a
CCAAT/enhancer binding protein (C/EBP), is a leucine zipper
transcription factor that is present in low levels under normal
conditions but is robustly expressed in response to stress
(29–31). GADD153 was originally identified
based on its induction following treatment of cells with growth
arresting and DNA-damaging agents, although the induced expression
of the gene has also been strongly linked to the perturbation of
homeostasis in the ER. It has been proven to be involved in the
pathogenesis of various diseases, including brain ischemia
(37), Alzheimer's disease
(38) and neurodegenerative
disease (37,38). The ER stress pathway is activated
by GADD153 and is involved in apoptosis. The T298M HOA-associated
mutation may be related to fibrillogenesis of collagen II/IX/XI
heterofibrils.
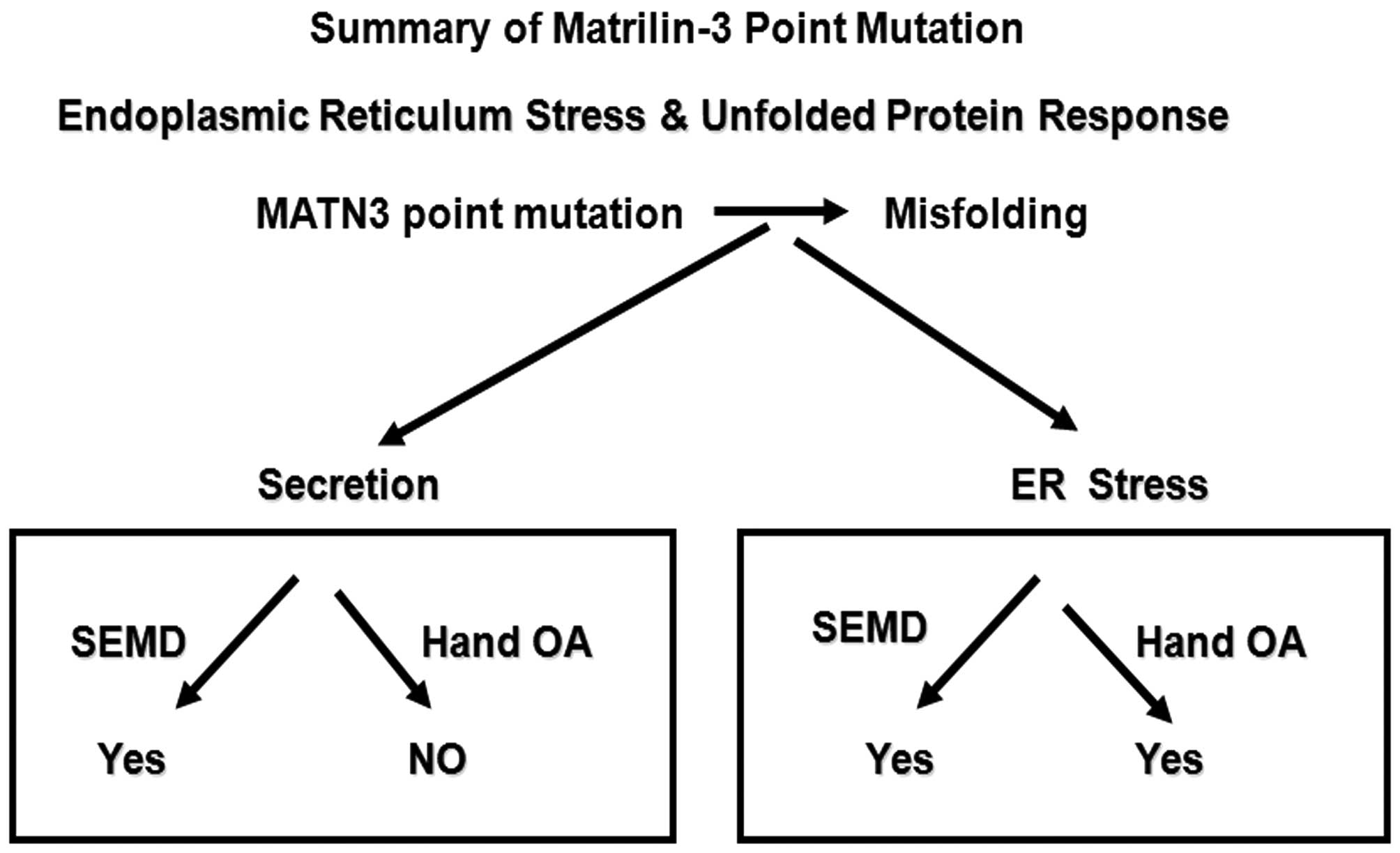
In conclusion, the findings of the present study
suggest that different point mutations at the first EGF-like domain
in matrilin-3 lead to distinct pathological mechanisms due to the
multiple functions of this EGF-like domain. This hypothesis would
explain how different mutations within matrilin-3 lead to the
development of a variety of diseases that affect cartilage
(Fig. 7).
Acknowledgments
The present study was funded partially through the
Chinese Natural Grant 81460022 and the New Faculty Foundation of
Qinghai Provincial People's Hospital (J.-M.L.) and partially
through the Department of Science and Technology of Hunan Province,
no. 2013FJ3126, and the Project of Scientific research platform of
The Affiliated Cancer Hospital of Xiangya Medical School, Central
South University, no. PT2013-09 (Y.-C.W.).
Abbreviations:
|
PAGE
|
polyacrylamide gel electrophoresis
|
|
RT-PCR
|
reverse transcription-polymerase chain
reaction
|
|
MED
|
multiple epiphyseal dysplasia
|
|
OA
|
osteoarthritis
|
|
HOA
|
hand osteo arthritis
|
|
SEMD
|
spondyloepimetaphyseal dysplasia
|
|
ER
|
endoplasmic reticulum
|
|
COMP
|
cartilage oligomeric matrix
protein
|
|
EGF
|
epidermal growth factor
|
References
|
1
|
Li L, Zhang L, Shao Y, Wang G, Gong R,
Wang Z, Peng J, Wang S, Genochio D, Zhao B and Luo J: Distinct
roles of two alternative splice variants of matrilin-2 in protein
oligomerization and proteolysis. Mol Med Rep. 6:1204–1210.
2012.PubMed/NCBI
|
|
2
|
Krieg T and LeRoy EC: Diseases of the
extracellular matrix. J Mol Med Berl. 76:224–225. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mateos J, De la Fuente A,
Lesende-Rodriguez I, Fernández-Pernas P, Arufe MC and Blanco FJ:
Lamin A deregulation in human mesenchymal stem cells promotes an
impairment in their chondrogenic potential and imbalance in their
response to oxidative stress. Stem Cell Res (Amst). 11:1137–1148.
2013. View Article : Google Scholar
|
|
4
|
Ankam S, Teo BK, Kukumberg M and Yim EK:
High throughput screening to investigate the interaction of stem
cells with their extracellular microenvironment. Organogenesis.
9:128–142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen Q, Zhang Y, Johnson DM and Goetinck
PF: Assembly of a novel cartilage matrix protein filamentous
network: molecular basis of differential requirement of von
Willebrand factor A domains. Mol Biol Cell. 10:2149–2162. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Frank S, Schulthess T, Landwehr R, Lustig
A, Mini T, Jenö P, Engel J and Kammerer RA: Characterization of the
matrilin coiled-coil domains reveals seven novel isoforms. J Biol
Chem. 277:19071–19079. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Klatt AR, Becker AK, Neacsu CD, Paulsson M
and Wagener R: The matrilins: modulators of extracellular matrix
assembly. Int J Biochem Cell Biol. 43:320–330. 2011. View Article : Google Scholar
|
|
8
|
Paulsson M, Piecha D, Segat D, Smyth N and
Wagener R: The matrilins: a growing family of A-domain-containing
proteins. Biochem Soc Trans. 27:824–826. 1999. View Article : Google Scholar
|
|
9
|
Piecha D, Muratoglu S, Mörgelin M, Hauser
N, Studer D, Kiss I, Paulsson M and Deák F: Matrilin-2, a large,
oligomeric matrix protein, is expressed by a great variety of cells
and forms fibrillar networks. J Biol Chem. 274:13353–13361. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Whittaker CA and Hynes RO: Distribution
and evolution of von Willebrand/integrin A domains: widely
dispersed domains with roles in cell adhesion and elsewhere. Mol
Biol Cell. 13:3369–3387. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luo J and Wan Y: Tightly regulated
distribution of family members of proteins is related to social
property in the open body system (Review). Int J Mol Med.
17:411–418. 2006.PubMed/NCBI
|
|
12
|
Belluoccio D, Schenker T, Baici A and
Trueb B: Characterization of human matrilin-3 (MATN3). Genomics.
53:391–394. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Y and Chen Q: Changes of matrilin
forms during endochondral ossification. Molecular basis of
oligomeric assembly. J Biol Chem. 275:32628–32634. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
van der Weyden L, Wei L, Luo J, Yang X,
Birk DE, Adams DJ, Bradley A and Chen Q: Functional knockout of the
matrilin-3 gene causes premature chondrocyte maturation to
hypertrophy and increases bone mineral density and osteoarthritis.
Am J Pathol. 169:515–527. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Y, Wang ZK, Luo JM, Kanbe K and Chen
Q: Multiple functions of the von Willebrand Factor A domain in
matrilins: secretion, assembly, and proteolysis. J Orthop Surg.
3:212008. View Article : Google Scholar :
|
|
16
|
Chapman KL, Mortier GR, Chapman K,
Loughlin J, Grant ME and Briggs MD: Mutations in the region
encoding the von Willebrand factor A domain of matrilin-3 are
associated with multiple epiphyseal dysplasia. Nat Genet.
28:393–396. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jackson GC, Barker FS, Jakkula E,
Czarny-Ratajczak M, Mäkitie O, Cole WG, Wright MJ, Smithson SF,
Suri M, Rogala P, et al: Missense mutations in the beta strands of
the single A-domain of matrilin-3 result in multiple epiphyseal
dysplasia. J Med Genet. 41:52–59. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mäkitie O, Mortier GR, Czarny-Ratajczak M,
Wright MJ, Suri M, Rogala P, Freund M, Jackson GC, Jakkula E,
Ala-Kokko L, et al: Clinical and radiographic findings in multiple
epiphyseal dysplasia caused by MATN3 mutations: description of 12
patients. Am J Med Genet A. 125A:278–284. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mabuchi A, Haga N, Maeda K, Nakashima E,
Manabe N, Hiraoka H, Kitoh H, Kosaki R, Nishimura G, Ohashi H and
Ikegawa S: Novel and recurrent mutations clustered in the von
Willebrand factor A domain of MATN3 in multiple epiphyseal
dysplasia. Hum Mutat. 24:439–440. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cotterill SL, Jackson GC, Leighton MP,
Wagener R, Mäkitie O, Cole WG and Briggs MD: Multiple epiphyseal
dysplasia mutations in MATN3 cause misfolding of the A-domain and
prevent secretion of mutant matrilin-3. Hum Mutat. 26:557–565.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wouters MA, Rigoutsos I, Chu CK, Feng LL,
Sparrow DB and Dunwoodie SL: Evolution of distinct EGF domains with
specific functions. Protein Sci. 14:1091–1103. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rao Z, Handford P, Mayhew M, Knott V,
Brownlee GG and Stuart D: The structure of a
Ca2+-binding epidermal growth factor-like domain: its
role in protein-protein interactions. Cell. 82:131–141. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Borochowitz ZU, Scheffer D, Adir V,
Dagoneau N, Munnich A and Cormier-Daire V: Spondylo-epi-metaphyseal
dysplasia (SEMD) matrilin 3 type: homozygote matrilin 3 mutation in
a novel form of SEMD. J Med Genet. 41:366–372. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Stefánsson SE, Jónsson H, Ingvarsson T,
Manolescu I, Jónsson HH, Olafsdóttir G, Pálsdóttir E, Stefánsdóttir
G, Sveinbjörnsdóttir G, Frigge ML, et al: Genomewide scan for hand
osteoarthritis: a novel mutation in matrilin-3. Am J Hum Genet.
72:1448–1459. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang S, Peng J, Guo Y, Javidiparsijani S,
Wang G, Wang Y, Liu H, Liu J and Luo J: Matrilin-2 is a widely
distributed extracellular matrix protein and a potential biomarker
in the early stage of osteoarthritis in articular cartilage. Biomed
Res Int. 2014:9861272014.PubMed/NCBI
|
|
26
|
Wei F, Moore DC, Wei L, Li Y, Zhang G, Wei
X, Lee JK and Chen Q: Attenuation of osteoarthritis via blockade of
the SDF-1/CXCR4 signaling pathway. Arthritis Res Ther. 14:R1772012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lawrence RC, Felson DT, Helmick CG, Arnold
LM, Choi H, Deyo RA, Gabriel S, Hirsch R, Hochberg MC, Hunder GG,
et al National Arthritis Data Workgroup: Estimates of the
prevalence of arthritis and other rheumatic conditions in the
United States. Part II. Arthritis Rheum. 58:26–35. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Otten C, Wagener R, Paulsson M and Zaucke
F: Matrilin-3 mutations that cause chondrodysplasias interfere with
protein trafficking while a mutation associated with hand
osteoarthritis does not. J Med Genet. 42:774–779. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu H, Qian J, Wang F, Sun X, Xu X, Xu W
and Zhang X and Zhang X: Expression of two endoplasmic reticulum
stress markers, GRP78 and GADD153, in rat retinal detachment model
and its implication. Eye (Lond). 24:137–144. 2010. View Article : Google Scholar
|
|
30
|
Feng LJ, Jiang TC, Zhou CY, Yu CL, Shen
YJ, Li J and Shen YX: Activated macrophage-like synoviocytes are
resistant to endoplasmic reticulum stress-induced apoptosis in
antigen-induced arthritis. Inflamm Res. 63:335–346. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Carlisle RE, Brimble E, Werner KE, Cruz
GL, Ask K, Ingram AJ and Dickhout JG: 4-Phenylbutyrate inhibits
tunicamycin-induced acute kidney injury via CHOP/GADD153
repression. PLoS One. 9:e846632014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Downing AK, Knott V, Werner JM, Cardy CM,
Campbell ID and Handford PA: Solution structure of a pair of
calcium-binding epidermal growth factor-like domains: implications
for the Marfan syndrome and other genetic disorders. Cell.
85:597–605. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bork P, Downing AK, Kieffer B and Campbell
ID: Structure and distribution of modules in extracellular
proteins. Q Rev Biophys. 29:119–167. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Vranka J, Mokashi A, Keene DR, Tufa S,
Corson G, Sussman M, Horton WA, Maddox K, Sakai L and Bächinger HP:
Selective intracellular retention of extracellular matrix proteins
and chaperones associated with pseudoachondroplasia. Matrix Biol.
20:439–450. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hecht JT, Makitie O, Hayes E, Haynes R,
Susic M, Montufar-Solis D, Duke PJ and Cole WG: Chondrocyte cell
death and intracellular distribution of COMP and type IX collagen
in the pseudoachondroplasia growth plate. J Orthop Res. 22:759–767.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mann HH, Ozbek S, Engel J, Paulsson M and
Wagener R: Interactions between the cartilage oligomeric matrix
protein and matrilins. Implications for matrix assembly and the
pathogenesis of chondrodysplasias. J Biol Chem. 279:25294–25298.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Oyadomari S and Mori M: Roles of
CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ.
11:381–389. 2004. View Article : Google Scholar
|
|
38
|
Prasanthi JRP, Larson T, Schommer J and
Ghribi O: Silencing GADD153/CHOP gene expression protects against
Alzheimer's disease-like pathology induced by 27-hydroxycholesterol
in rabbit hippocampus. PLoS One. 6:e264202011. View Article : Google Scholar : PubMed/NCBI
|