Introduction
Environmental pollution emerged as a significant
threat to human health early in the 20th century, as industrial
societies became aware of an increased prevalence of a range of
chronic illnesses and conditions. Environmental pollution is fueled
by large-scale generation of synthetic chemicals that are normally
employed in necessary industrial processes (1,2).
Among these anthropogenic chemicals, perfluoroalkyl compounds
(PFCs) are organofluorines, which have fluorine instead of hydrogen
in their carbon chains. Strong bonds form between fluorine and
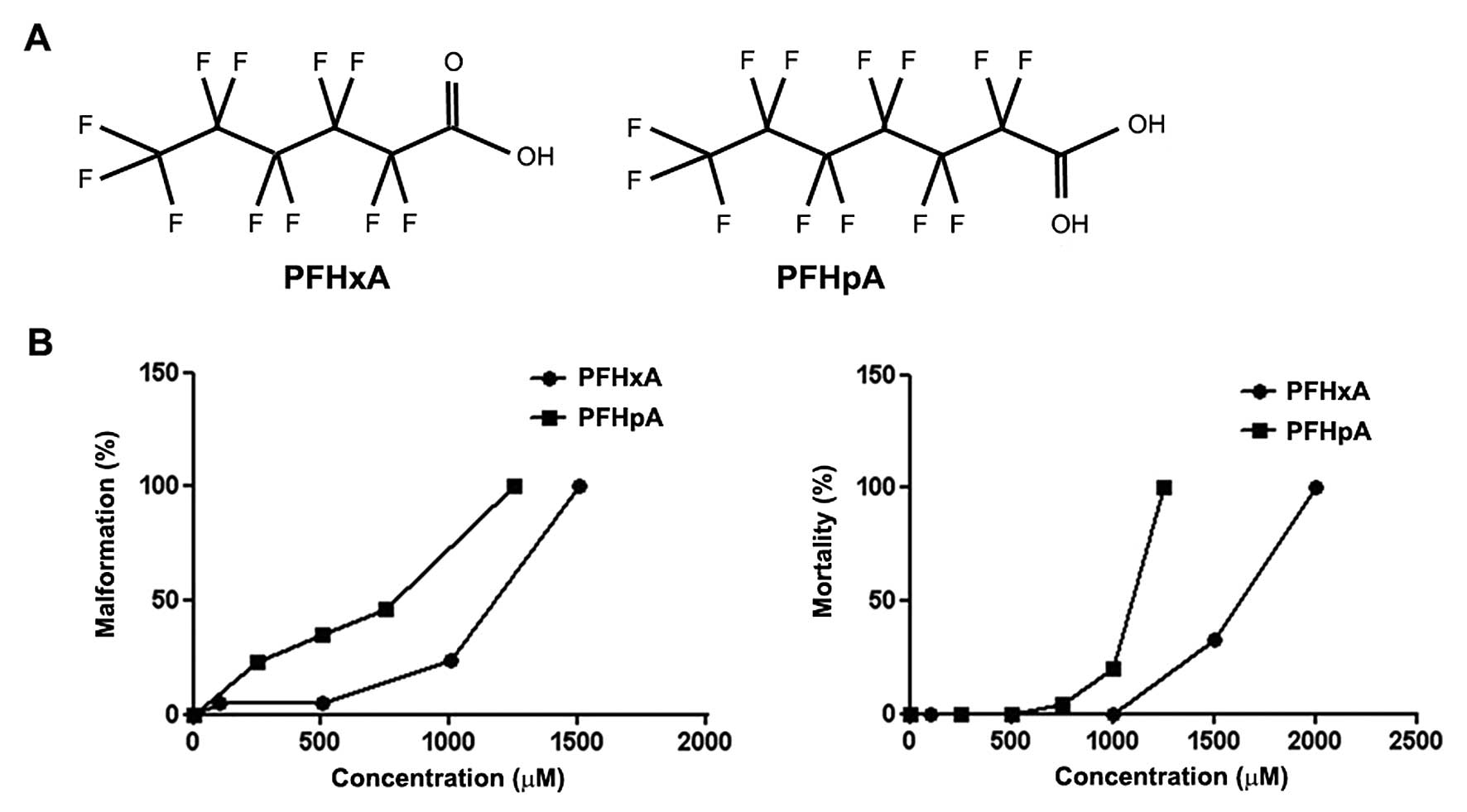
carbon (Fig. 1A). The
carbon-fluorine (C–F) bond is considered the strongest polar
covalent bond, with a high dissociation energy reaching 117
kcal/mol. The strong polarity and high dissociation energy of C-F
bonds make PFCs resistant to a range of biochemical processes such
as thermal, chemical, hydrolytic, and biological degradation
(2). Furthermore, the hydrophobic
fluorinated alkyl chain and the lipophilic functional group make
PFCs valuable in many industrial and commercial processes involved
in the production of fabrics, electronics, pesticides, emulsifiers,
paints and adhesives (3,4). This wide-ranging use of
perfluorohexanoic (PFHxA) and perfluoroheptanoic (PFHpA) acids has
resulted in their detection as pollutants in water, sediments, air,
wildlife and humans, and these compounds are not degraded by any
naturally occurring processes in the environment (5).
PFCs are present in atmospheric and aquatic systems
on a global level (6).
Environmental levels of PFCs were investigated in different world
regions, identifying high PFC levels reaching 100 ng/l (7–9).
The concentrations of various PFCs, which exert unpleasant effects
in humans, have been determined in human blood samples from
different world regions (10).
High levels of PFCs in serum and liver samples from both humans and
animals have also been detected (11). The levels of PFCs were elevated in
individuals with regular occupational exposure to PFCs (4.3 ppb),
as compared to the general population (12). Their unique physicochemical
properties make PFCs highly persistent, increasing their
bioaccumulation and toxicological effects (5,13).
In addition, it has been shown that PFCs exert toxic reproductive
and developmental effects on human and animal systems (12,14,15). Overexposure to these anthropogenic
chemicals causes severe fetal abnormalities in amphibians and
affects human development (12,16). The toxicity of PFCs is commonly
determined by their carbon chain length and by the functional
group.
The frog embryo teratogenicity assay-Xenopus
(FETAX) utilizing anuran Xenopus embryos is a
well-characterized method used to measure the developmental
toxicity of chemicals. The FETAX assay is performed on fertilized
Xenopus embryos in the middle of the blastula stage, at 96 h
(17). Teratogenic effects are
assessed by determining their influences on mortality,
malformation, and growth inhibition, effects that clearly indicate
developmental toxicity. This assay has been used to investigate
teratogens and developmental toxicants that negatively affect human
health and the ecosystem (18,19). FETAX is appropriate for the
screening of chemicals (including nanomaterials), evaluating
biotrans-formation, and can serve as a model of limb development.
It is used to inform risk assessments of the effects of
environmental hazards on amphibian and vertebrate populations. In
addition to FETAX, reverse transcriptase-polymerase chain reaction
(RT-PCR) and whole-mount in situ hybridization are useful in
the investigation of the developmental toxicity of these compounds,
as well as mechanisms underlying any effects on Xenopus
embryos.
The toxicological potentials of PFHxA and PFHpA need
to be investigated to determine the adverse effects of PFCs with
different lengths. In the present study, we evaluated the
developmental toxicity and teratogenicity of PFHxA and PFHpA using
different biological assays. The results obtained from the in
vivo model system showed that PFHpA is a more potent
developmental toxicant and teratogen than PFHxA. In particular,
PFHxA and PFHpA affected mortality and the growth rates of embryos,
and caused severe developmental liver and heart defects. These
results suggested that although PFHxA and PFHpA are developmental
toxicants and teratogens, PFHpA has more severe effects on liver
and heart development via the induction of extracellular
signal-regulated kinase (ERK) and c-Jun N-terminal kinase (JNK)
phosphorylation.
Materials and methods
Ethics statement
We obtained a specific waiver from the Institutional
Review Board of Kyungpook National University for the experimental
use of amphibians or reptiles in Korea (no. KNU-2013-222). All the
members of our research group attended educational and training
courses on the appropriate care and usage of experimental animals.
Adult Xenopus laevis were entrained in 12-h light/dark
cycles at 18°C in containers provided by the Institutional Review
Board of Kyungpook National University constructed to
specifications for laboratory animal maintenance. There were no
unexpected deaths of adult Xenopus during the study.
Chemicals and reagents
PFHxA and PFHpA were purchased from Sigma-Aldrich
(St. Louis, MO, USA) and dissolved in dimethyl sulfoxide (DMSO) to
produce 1 M stock solutions. The FETAX medium used in the
experiment contained 10.7 mM NaCl, 1.14 mM NaHCO3, 0.4
mM KCl, 0.1 mM CaCl2, 0.35 mM
CaSO4.2H2O, and 0.3 mM MgSO4 and
the levels of DMSO did not exceed 0.15%, which was below the
permissible level for this assay. PFC stocks were freshly made and
diluted in FETAX medium.
In vitro fertilization
Experiments on Xenopus laevis were conducted
in accordance with documented standards of the Animal Care and Use
Committee, in agreement with international laws and policies
[National Institutes of Health (NIH) Guide for the Care and Use of
Laboratory Animals, NIH publication no. 85–23, 1985]. Adult
Xenopus were purchased from Nasco (Fort Atkinson, WI, USA)
and kept in climate-controlled clear plastic aquariums with
dechlorinated tap water at 18±2°C, with 12-h light/dark cycles. The
housed frogs were fed only three times a week. For the induction of
ovulation, human chorionic gonadotropin (1,000 IU) (Sigma-Aldrich)
was injected under the skin of a female Xenopus in the
evening. The following day, the females were made to lay eggs in
60-mm plastic dishes. The eggs were immediately fertilized in 0.1X
modified Barth's saline (MBS) containing 88 mM NaCl, 5 mM Hepes,
2.5 mM NaHCO3, 1 mM KCl, 1 mM MgSO4, and 0.7
mM CaCl2 (pH 7.8) after washing three times with 0.1X
MBS. The frogs were sacrificed as follows: The anesthetic of choice
for male frogs, tricaine methanesulfonate (MS222) was dissolved in
dechlorinated water in an induction tank at a dose of 500 mg of
MS222/l of water. Xenopus testes were obtained from the
sacrificed males and minced in 1 ml of cold 1X MBS. Following
successful fertilization, the jelly coat was removed from the
embryos by gentle swirling in a 2% L-cysteine solution. The embryos
were then transferred to 1X MBS containing 3% Ficoll 400 (GE
Healthcare, Little Chalfont, UK). Unfertilized eggs and dead
embryos were removed and the remaining embryos were maintained at
22±0.5°C until they reached blastula stage 8.5.
FETAX assay
FETAX assays were conducted to assess the
developmental toxicity and teratogenic effects of the PFCs based on
the American Society of Testing Material (ASTM) guide (ASTM
E1439-98). Finely cleaved embryos in the blastula stage 8.5 were
selected and used to exclude the effects of spontaneous embryonic
developmental problems. Embryos were impartially allocated to
100-mm petri dishes (20–25 embryos/dish) and exposed to different
concentrations of PFHxA (0.1, 0.5, 1, 1.5 and 2 mM) and PFHpA
(0.25, 0.5, 0.75, 1 and 1.25 mM). DMSO (0.1%) and FETAX medium
alone were used as the controls. Embryos were incubated at 23°C
until the end of the assay, and the media were changed daily and
dead embryos were removed. At the end of the experiment, embryo
mortality was recorded and the surviving embryos were fixed in 4%
formaldehyde to determine whether there was any malformation.
Head-tail lengths and malformations were evaluated under a light
microscope and images were analyzed using Axiovision software
version 4.8 (Carl Zeiss, Munich, Germany) to measure growth
inhibition.
Whole-mount in situ hybridization
Xenopus embryos were prepared for whole-mount
in situ hybridization using a previously described method
(20), with minor modification.
Whole-mount in situ hybridization was conducted at the
tadpole stage (stage 34–36) using digoxigenin-labeled RNA probes
and BM purple staining. The probes were prepared in the following
manner: RT-PCR products of phosphotyrosine-binding protein (xPTB; a
liver marker) and NKX2.5 (heart marker) were sub-cloned into a
T-easy vector (Promega, Fitchburg, WI, USA), linearized with
ApaI and NcoI, respectively, and transcribed using a
SP6 RNA polymerase kit (Invitrogen, Carlsbad, CA, USA).
RT-PCR
RNA was isolated from stage 34–46 embryos using
TRIzol reagent, according to the manufacturer's instructions
(Invitrogen), and quantified using an Optizen 320 UV spectrometer
(Mecacys, Deajeon, Korea). An aliquot of RNA (1 g) was reverse
transcribed using a PrimeScript First-Strand cDNA Synthesis kit,
according to the manufacturer's instructions (Takara, Shiga,
Japan). Reactions containing template RNA without reverse
transcriptase were prepared as the negative controls (no-RT) and
carrier was used as the loading control. cDNA was amplified using
specific PCR primer sets (Table
I) and 2X Emerald Amp PCR Master mix (Takara). The PCR products
were separated in a 1–1.2% agarose gel and visualized on a
transilluminator (Vilber Lourmat, Marne-la-Vallée, France)
following ethidium bromide staining. The data were representative
of at least three experiments. The Primer3 program was used to
design primers and ODC was used as an internal control.
 | Table IPrimers for RT-PCR. |
Table I
Primers for RT-PCR.
| Gene name | GeneBank accession
no. | RT-PCR primer
sequences (5′→3′) | Product size
(bp) | Annealing
temperature (°C) | Cycles |
|---|
| xPTB | AY183756 | F:
ATGGATGCACTCAAGTCTGC | 933 | 58 | 28 |
| | R:
TGGTAGCTTCAGAAGTGTCAG | | | |
| NKX2.5 | NM_001172192 | F:
GTCAGCATCAACCCCTACAG | 491 | 53 | 28 |
| | R:
TGCCACCCAGTCCTTTAATC | | | |
| Cyl18 | AY188285 | F:
AGGCCGTAGCACCTGTGGGT | 462 | 55 | 21 |
| | R:
GGTCCCACCAGCTCGGATCA | | | |
| ODC | NM_001086698 | F:
GTGATGGGCTGGATCGTATC | 386 | 55 | 21 |
| | R:
TCCATTGAATGTCGAGGCTG | | | |
Extraction and analysis of the liver
Twenty-five embryos from each group were exposed to
PFHxA and PFHpA, respectively, for 10 days. The surviving tadpoles
were fixed in 4% formaldehyde for 2 h at room temperature. Tadpoles
(3–5) were randomly selected from each group
and their livers removed under a light microscope (Zeiss Stemi
2000-c; Carl Zeiss). Images were captured and analyzed to monitor
changes in the liver size.
Statistical analysis
Data for the three endpoints from each FETAX assay
were analyzed using GraphPad Prism software (GraphPad Software,
Inc., La Jolla, CA, USA). Images from the RT-PCR and whole-mount
in situ hybridization assays were analyzed by ImageJ
software (NIH). Statistical analyses were conducted using SPSS 13.0
software (version 18; SPSS, Inc., Chicago, IL, USA).
Results
Developmental toxicity and teratogenicity
assessment of PFCs by FETAX assay
FETAX assays were performed to determine the
potential teratogenicity and developmental toxicity of PFCs. FETAX
assay results were expressed relatively in terms of the teratogenic
index (TI)=LC50/EC50, where LC50 was the concentration that at
which 50% of the embryos were killed and EC50 was the concentration
at which 50% of the embryos were malformed. The minimum
concentration that inhibits growth (MCIG) was also determined.
Compounds were considered toxic when TI was ≥1.2 (17).
We performed an initial FETAX assay to determine
whether PFHxA and PFHpA had embryonic lethal or terato-genic
effects. This result confirmed that PFHxA and PFHpA were potential
teratogens and developmental toxicants. The LC50 value for PFHxA in
Xenopus embryos was 1523.5 µM, while the EC50 value
was 1046.5 µM, and the MCIG value was 412.3 µM, which
produced a TI value of 1.43, indicating that this compound was a
potential teratogen and developmental toxicant. The LC50 value for
PFHpA was 942.4 µM in the Xenopus embryos, with an
EC50 of 754.9 µM and an MCIG value of 297.4 µM,
producing a TI value of 1.25. Thus, this compound was also
considered a teratogen and developmental toxicant (Table II). PFHxA had a significantly
higher TI value (TI=1.43) than PFHpA (TI=1.25). Furthermore, the
LC50 and EC50 values were higher for PFHxA, which had a shorter
fluorinated carbon chain than PFHpA.
| Table IITeratogenic index for PFHxA and
PFHpA. |
Table II
Teratogenic index for PFHxA and
PFHpA.
| Acid | LC50 (µM)
lethality | EC50 (µM)
malformation | MCIG
(µM) | TI=LC50/EC50 |
|---|
| PFHxA | 1523.5 | 1046.5 | 412.3 | 1.43 |
| PFHpA | 942.4 | 754.9 | 297.4 | 1.25 |
Effects of PFHxA and PFHpA on Xenopus
mortality and malformation rates
Our FETAX assay results confirmed that PFHxA and
PFHpA are potential teratogens and developmental toxicants. We then
investigated the effects of different concentrations of PFHxA and
PFHpA on mortality and malformation in developing embryos. As shown
in Fig. 1B, exposure to PFHxA or
PFHpA greatly increased the mortality and general malformation
rates in the developing embryos. These concentration-dependent
effects on mortality and malformation were more severe as the
number of carbons increased. PFHpA significantly affected mortality
and malformation rates within a narrow concentration range
(Fig. 1B).
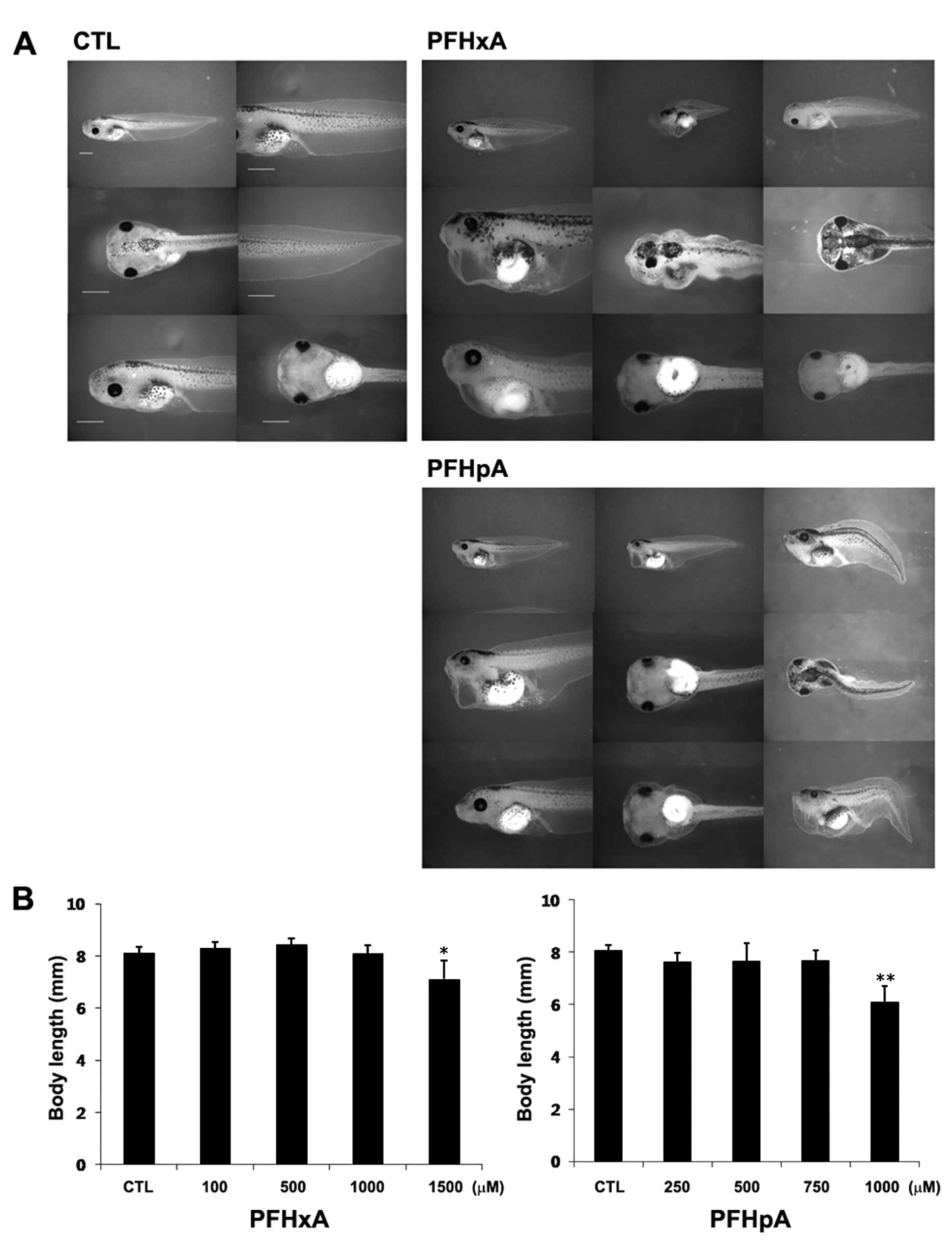
To investigate the mechanism underlying this
toxicity, we assessed the phenotypes of embryos exposed to PFHxA
and PFHpA. PFHxA and PFHpA were not highly toxic until early stage
26, as no significant differences were observed in the external
phenotypes. PFC exposure induced certain embryo nic abnormalities
that were observed at the end of the study, such as pericardial
edema, dorsal fin blisters, a curved body axis, lack of eye
pigment, and disturbed facial cells (Fig. 2A). PFHxA-treated embryos developed
mild and general malformations including gut mis-coiling and
improper eye shape, whereas PFHpA induced more severe and diverse
malformations (Fig. 2A). In
agreement with this, the whole body length was reduced by 14 and
24% in tadpoles treated with 1,500 µM of PFHxA or 1,000
µM of PFHpA, respectively (Fig. 2B). These results indicated that
embryos exposed to PFHxA or PFHpA show a range of malformations and
a reduced whole body length, which is a clear indicator of toxicity
while PFHpA has slightly more severe defects compared to PFHxA.
PFHpA modulates liver and heart
development during embryogenesis
We examined stage 36 tadpoles that had been
continuously exposed to 130 µM PFHxA or PFHpA using
whole-mount in situ hybridization. The liver is a major
metabolizing organ that detoxifies many compounds and is therefore
vulnerable to the toxic effects of PFCs. We also measured a
liver-specific marker, xPTB, which is expressed in the diverticulum
region of the embryonic liver and plays an important role in
low-density lipoprotein (LDL) receptor endocytosis in the liver
(21). Livers of embryos treated
with PFHxA developed abnormally and were unusually large (Fig. 3A). We also observed enlarged
livers in tadpoles exposed to PFHxA although these effects were
more pronounced in the presence of PFHpA (Fig. 2A). These results indicated that
PFHxA and PFHpA induced liver defects and should be regarded as
developmental toxicants and teratogens.
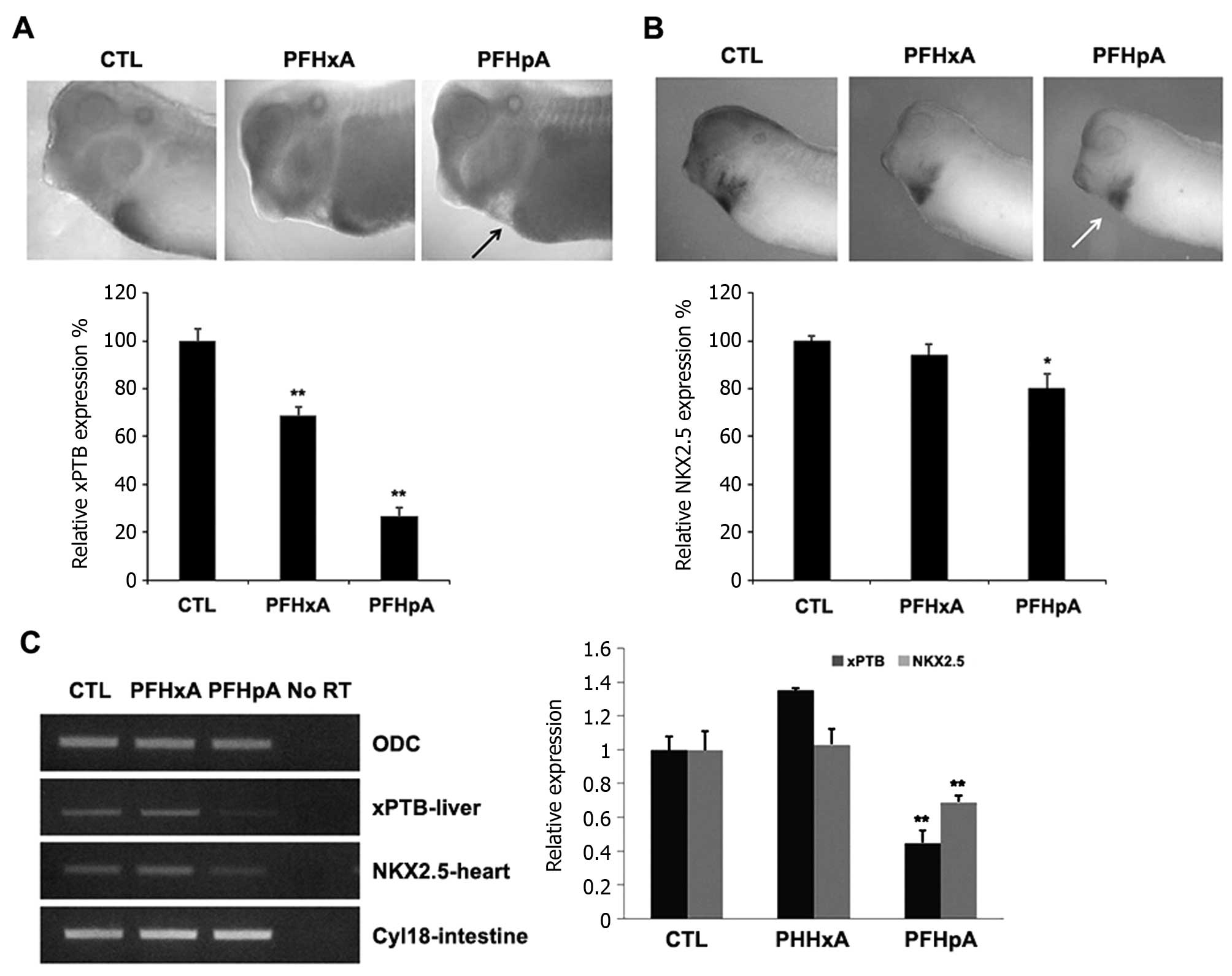 | Figure 3PFHpA severely affects liver and
heart development. (A) PFHxA and PFHpA exposure altered the
expression of tissue-specific mRNAs. Whole-mount in situ
hybridization of stage 36 embryos exposed to 130 µM PFHxA
and PFHpA. xPTB expression levels were reduced in the liver and
heart of exposed embryos (black arrow). The histogram shows the
relative area and intensity of xPTB in the liver, expressed as a
percentage of the control. Data are the mean of three independent
experiments. (B) PFHxA and PFHpA exposure altered the expression of
tissue-specific mRNAs. Whole-mount in situ hybridization of
stage 36 embryos exposed to 130 µM PFHxA and PFHpA. NKX2.5
expression levels were reduced in the heart of exposed embryos
(white arrow). The histogram shows the relative area and intensity
of NKX2.5 mRNA expression in the heart, expressed as a percentage
of the control. Data are the mean of three independent experiments.
(C) The levels of xPTB, NKX2.5, and Cyl18 mRNA expression were
determined by RT-PCR. PCR was conducted without reverse
transcriptase (No-RT) as a negative control and the carrier was
used as the loading control. The histogram shows the relative
expression levels of xPTB, NKX2.5, expressed as a percentage of the
control ODC. Mean ± SD; **P<0.05,
*P<0.05 compared with the control cells. PFHxA,
perfluorohexanoic acid; PFHpA, perfluoroheptanoic acid; SD,
standard deviation. |
The heart-specific marker, NKX2.5, was used to
examine the cardiovascular effects of PFC exposure. The amphibian
heart is composed of one ventricle, surrounded by atria on both
sides, separated by a septum (22). Embryos treated with PFHxA and
PFHpA showed enlarged atria and a loss of the atrial septum
(Fig. 3B). Hearts from embryos
exposed to either PFHxA or PFHpA were smaller and contained no
trabeculae. Additionally, the pericardiac cavity was expanded and
the atrial and ventricular walls were thinner than those observed
in the control embryo hearts, which contained prominent trabeculae
and the atria and ventricle were separated (Fig. 3B). This effect was more prominent
in embryos exposed to PFHpA, which reduced trabeculation of the
ventricle as well as the thickness of the atrial and ventricular
walls (Fig. 3B). The results
showed that PFHxA and PFHpA treatment caused abnormal heart
development.
PFC exposure alters tissue-specific mRNA
expression in vivo
To investigate the effects of PFC exposure on the
liver and heart further, the mRNA expression levels of three liver,
heart, and intestine markers were determined by RT-PCR analysis.
Previously, PFCs were shown to be tissue-specific toxicants
(14,23,24). For this study, we exposed stage 36
embryos to 130 µM PFHxA or PFHpA, a concentration that
induced the development of various malformations, rather than
embryonic lethality. As shown in Fig.
2C, mRNA expression of xPTB was considerably reduced in
PFHpA-treated embryos, as compared to the control or PFHxA-treated
embryos (Fig. 3A and C). Previous
findings have also shown reduced xPTB expression following exposure
to various PFCs (16,25,26). Furthermore, the heart-specific
marker, NKX2.5, was used to examine the effects of PFCs on the
heart. As shown in Fig. 2B and C,
mRNA expression of the heart-specific marker, NKX2.5, was reduced
in the PFHpA-treated embryos (Fig. 3B
and C). NKX2.5 is critical for vasculogenesis and angiogenesis
(27). We also assessed the mRNA
levels of an intestinal marker, Cyl18, which is mainly expressed in
the presumptive small intestine of embryos at the tailbud stage. As
shown in Fig. 2C, we observed no
significant effects of either PFHxA or PFHpA exposure on Cy118
expression. These results suggested that PFHpA negatively affected
liver and heart development (Fig.
3C).
PFCs may induce liver toxicity by
modifying ERK and JNK phosphorylation
The roles of peroxisome proliferator-activated
receptor-γ (PPARγ), ERK, and JNK pathways in mediating the
developmental toxicity associated with exposure to PFCs remain to
be elucidated. To investigate the effects of PFCs on PPARγ, ERK,
and JNK pathways, stage 36 embryos were exposed to 130 µM
PFHxA or PFHpA and then analyzed by western blotting with
anti-PPARγ, phosphorylated-ERK, ERK, p-JNK, JNK, and β-actin
antibodies. PFHpA strongly induced ERK and JNK phosphorylation
while PPARγ expression remained the same, which in turn activated
signaling that caused liver toxicity, known as hepatocarcinogenesis
(Fig. 4). These results clearly
demonstrated that the observed developmental defects in the liver
were associated with the PFHpA-mediated activation of ERK/JNK
pathways.
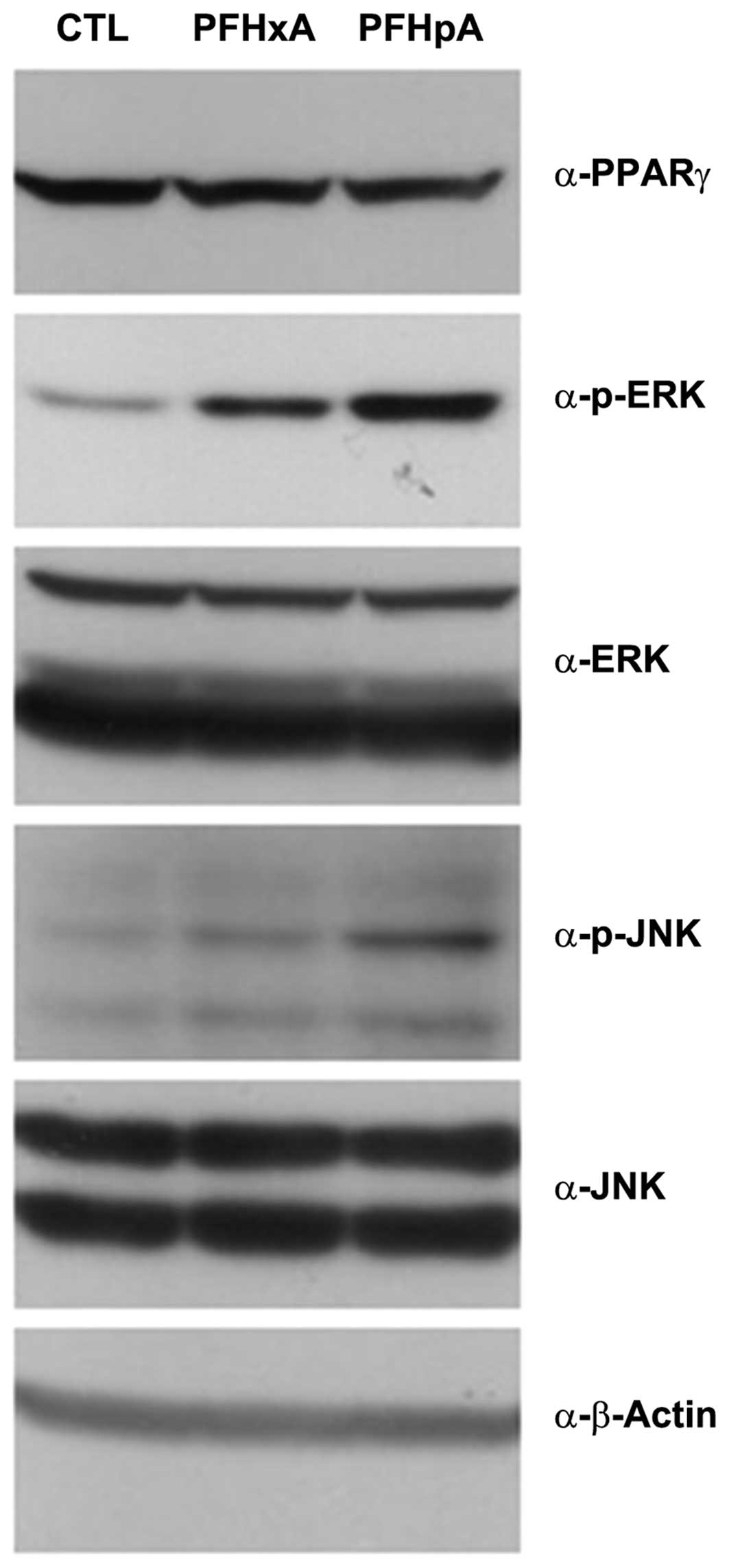 | Figure 4PFCs may induce liver toxicity by
modifying ERK and JNK phosphorylation. Xenopus embryos were
exposed to 130 µM PFHxA or PFHpA. Total protein was isolated
and analyzed by western blotting with antibodies raised against
PPARγ, p-ERK, ERK, p-JNK, JNK, or β-actin. PFHxA and PFHpA
treatment activated PPARγ and ERK/JNK phosphorylation. PFCs,
perfluoroalkyl compounds; ERK, extracellular signal-regulated
kinase; JNK, c-Jun N-terminal kinase; PFHxA, perfluorohexanoic
acid; PFHpA, perfluoroheptanoic acid; PPARγ, peroxisome
proliferator-activated receptor-γ. |
PFC treatment induces hepatomegaly
Hepatomegaly is often associated with chronic
inflammatory liver disease. The liver is the major metabolizing
organ and it is targeted following PFC exposure (28). Previous findings have shown that
exposure to PFCs caused various chronic abnormalities, including
hepato-cellular hypertrophy and hepatomegaly (29,30). To investigate the effects of PFHxA
and PFHpA on the liver, livers from 10-day old tadpoles that had
been continuously exposed to 130 µM PFHxA or PFHpA exhibited
hepatomegaly (Fig. 5). These
results suggested that high levels of exposure to PFHpA or PFHxA
may induce hepatic disorders.
Discussion
PFCs have emerged as persistent organic pollutants
in the ecosystem and human exposure to these may increase the risk
of developmental abnormalities (31). Several studies have examined the
toxicity of different PFC in many aquatic and other animals
(24,32,33). These studies have confirmed that
exposure during particular developmental stages or under certain
nutritional conditions produced adverse effects on neonates
(34,35). Amphibians are more sensitive to
environmental exposures during the fetal stage. The results of that
study showed that the PFCs tested induced malformations in
Xenopus embryos, including pericardial edema and hepatic
defects. The industrial production of PFCs and their derivatives
has been banned because of their toxicity and persistence in the
environment. Given these conditions, developmental toxicity studies
of PFCs remain in their infancy and further large-scale
investigations are required.
Xenopus laevis is an amphibian species
considered to be a suitable model for studying developmental
defects induced by toxicants. In the present study, we conducted a
FETAX assay using Xenopus laevis to examine the
developmental toxicities of PFHxA and PFHpA. The FETAX assay
provides a classical approach to detecting teratogenic effects of
chemicals on fertilized Xenopus embryos in the middle of the
blastula stage (17). When
Xenopus embryos were exposed to C6–7 PFCs, various
malformations and toxicity were observed, such as abnormal
development, bent and uneven body axis deformation, multiple
edemas, growth inhibition, and intestinal miscoiling (Fig. 2A). The PFCs tested produced
different TI values, showing that the intensity of their effects on
development were different, although they may operate via identical
mechanisms.
Our results have demonstrated that PFHxA and PFHpA
acted as developmental toxicants and teratogens, thereby producing
lethal effects in Xenopus embryos. The difference between
their effects may reflect the variation in the number of
fluorinated carbons present in these compounds. In addition, PFHxA
and PFHpA considerably modulated the expression levels of various
tissue-specific genes that were relevant to the observed external
and internal phenotypes. In particular, xPTB was used as a
liver-specific marker and the protein encoded by this gene
complexes with the LDL receptor to mediate hepatic LDL removal by
endocytosis (35,36). Consistent with other reports, the
xPTB mRNA levels were considerably modulated following exposure to
either PFHxA or PFHpA (16). This
specific reduction in the xPTB mRNA level may cause increased LDL
accumulation in the liver, which can in turn lead to chronic
cardiac disease, such as atherosclerosis and myocardial infarction
(37). Consistent with this,
individuals with a high concentration of PFCs in their blood are
more susceptible to LDL-associated adverse effects (5).
The amphibian heart develops from the mesoderm and
is considered an essential organ for embryonic survival. During
embryogenesis, NKX2.5 acts as a key transcription factor,
regulating the transcription of downstream targets that are
essential for heart formation (38,39). Studies have shown that NKX2.5 has
an important role in the formation of the septum, which separates
the ventricle from the atrium (15,38,39). Severe heart abnormalities such as
atrial septal defects have been reported in association with the
NKX2.5 mutation and downregulation. PFHxA and PFHpA treatments
reduced the expression of NKX2.5 mRNA, which is critical for
vasculogenesis and angiogenesis (32). Thus, PFCs may cause improper
septum formation and impair cardiac function by downregulating the
expression of NKX2.5. By contrast, no major change was observed in
the intestine-specific marker, Cy118, following treatment with
PFHxA and PFHpA.
Compelling evidence for PFC toxicity has emerged
from pre-clinical and clinical studies (12,18). Activation of JNK and ERK pathway
causes toxic effects that include hepatocarcino-genesis. Livers
from PFHxA- and PFHpA-treated embryos exhibited hepatomegaly. This
developmental and teratogenic effect was more pronounced in the
presence of PFHpA, which may be due to its longer fluorinated
carbon chain.
In conclusion, our results confirm that PFHxA and
PFHpA are developmental toxicants and teratogens in Xenopus.
In addition, PFHpA induced more severe pericardial edema, blisters
on the dorsal fin, a curved body axis, lack of eye pigmentation,
and disturbed facial cells. Furthermore, PFHpA produced more severe
effects on liver and heart development through the induction of ERK
and JNK phosphorylation, and exposure to elevated levels of PFHpA
may therefore cause developmental toxicity and teratogenicity,
inducing hepatic and cardiac disorders.
Acknowledgments
The present study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF), funded by the Ministry of Education, Science and Technology
(no. NRF-2012R1A1A2042334), and by a grant from the National
Fisheries Research and Development Institute (NFRDI; contribution
no. RP-2015-AQ-072), Republic of Korea.
References
|
1
|
Andersen ME, Butenhoff JL, Chang SC,
Farrar DG, Kennedy GL Jr, Lau C, Olsen GW, Seed J and Wallace KB:
Perfluoroalkyl acids and related chemistries - toxicokinetics and
modes of action. Toxicol Sci. 102:3–14. 2008. View Article : Google Scholar
|
|
2
|
Houde M, Martin JW, Letcher RJ, Solomon KR
and Muir DC: Biological monitoring of polyfluoroalkyl substances: A
review. Environ Sci Technol. 40:3463–3473. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Calafat AM, Wong LY, Kuklenyik Z, Reidy JA
and Needham LL: Polyfluoroalkyl chemicals in the U.S. population:
Data from the National Health and Nutrition Examination Survey
(NHANES) 2003–2004 and comparisons with NHANES 1999–2000. Environ
Health Perspect. 115:1596–1602. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lindstrom AB, Strynar MJ and Libelo EL:
Polyfluorinated compounds: Past, present, and future. Environ Sci
Technol. 45:7954–7961. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nelson JW, Hatch EE and Webster TF:
Exposure to polyfluoroalkyl chemicals and cholesterol, body weight,
and insulin resistance in the general U.S. population. Environ
Health Perspect. 118:197–202. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yamashita N, Kannan K, Taniyasu S, Horii
Y, Petrick G and Gamo T: A global survey of perfluorinated acids in
oceans. Mar Pollut Bull. 51:658–668. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McLachlan MS, Holmstrom KE, Reth M and
Berger U: Riverine discharge of perfluorinated carboxylates from
the European continent. Environ Sci Technol. 41:7260–7265. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jin YH, Liu W, Sato I, Nakayama SF, Sasaki
K, Saito N and Tsuda S: PFOS and PFOA in environmental and tap
water in China. Chemosphere. 77:605–611. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakayama S, Strynar MJ, Helfant L, Egeghy
P, Ye X and Lindstrom AB: Perfluorinated compounds in the Cape Fear
Drainage Basin in North Carolina. Environ Sci Technol.
41:5271–5276. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Domingo JL: Health risks of dietary
exposure to perfluorinated compounds. Environ Int. 40:187–195.
2012. View Article : Google Scholar
|
|
11
|
Moody CA, Martin JW, Kwan WC, Muir DC and
Mabury SA: Monitoring perfluorinated surfactants in biota and
surface water samples following an accidental release of
fire-fighting foam into Etobicoke Creek. Environ Sci Technol.
36:545–551. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Olsen GW, Butenhoff JL and Zobel LR:
Perfluoroalkyl chemicals and human fetal development: An
epidemiologic review with clinical and toxicological perspectives.
Reprod Toxicol. 27:212–230. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dreyer A, Weinberg I, Temme C and
Ebinghaus R: Polyfluorinated compounds in the atmosphere of the
Atlantic and Southern Oceans: Evidence for a global distribution.
Environ Sci Technol. 43:6507–6514. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lau C, Anitole K, Hodes C, Lai D,
Pfahles-Hutchens A and Seed J: Perfluoroalkyl acids: A review of
monitoring and toxicological findings. Toxicol Sci. 99:366–394.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kato K, Wong LY, Jia LT, Kuklenyik Z and
Calafat AM: Trends in exposure to polyfluoroalkyl chemicals in the
U.S. Population: 1999–2008. Environ Sci Technol. 45:8037–8045.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim M, Son J, Park MS, Ji Y, Chae S, Jun
C, Bae JS, Kwon TK, Choo YS, Yoon H, et al: In vivo evaluation and
comparison of developmental toxicity and teratogenicity of
perfluoroalkyl compounds using Xenopus embryos. Chemosphere.
93:1153–1160. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mouche I, Malesic L and Gillardeaux O:
FETAX assay for evaluation of developmental toxicity. Methods Mol
Biol. 691:257–269. 2011. View Article : Google Scholar
|
|
18
|
Sharma B and Patiño R: Exposure of Xenopus
laevis tadpoles to cadmium reveals concentration-dependent bimodal
effects on growth and monotonic effects on development and thyroid
gland activity. Toxicol Sci. 105:51–58. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bacchetta R, Santo N, Fascio U, Moschini
E, Freddi S, Chirico G, Camatini M and Mantecca P: Nano-sized CuO,
TiO2 and ZnO affect Xenopus laevis development.
Nanotoxicology. 6:381–398. 2012. View Article : Google Scholar
|
|
20
|
Vize PD, McCoy KE and Zhou X: Multichannel
wholemount fluorescent and fluorescent/chromogenic in situ
hybridization in Xenopus embryos. Nat Protoc. 4:975–983. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou Y, Zhang J and King ML: Xenopus
autosomal recessive hypercholesterolemia protein couples
lipoprotein receptors with the AP-2 complex in oocytes and embryos
and is required for vitellogenesis. J Biol Chem. 278:44584–44592.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jahr M and Männer J: Development of the
venous pole of the heart in the frog Xenopus laevis: A
morphological study with special focus on the development of the
venoatrial connections. Dev Dyn. 240:1518–1527. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ruch RJ: Intercellular communication,
homeostasis, and toxicology. Toxicol Sci. 68:265–266. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cheng Y, Cui Y, Chen HM and Xie WP:
Thyroid disruption effects of environmental level perfluorooctane
sulfonates (PFOS) in Xenopus laevis. Ecotoxicology. 20:2069–2078.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ny A, Koch M, Schneider M, Neven E, Tong
RT, Maity S, Fischer C, Plaisance S, Lambrechts D, Héligon C, et
al: A genetic Xenopus laevis tadpole model to study
lymphangiogenesis. Nat Med. 11:998–1004. 2005.PubMed/NCBI
|
|
26
|
Kälin RE, Bänziger-Tobler NE, Detmar M and
Brändli AW: An in vivo chemical library screen in Xenopus tadpoles
reveals novel pathways involved in angiogenesis and
lymphangiogenesis. Blood. 114:1110–1122. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Raffin M, Leong LM, Rones MS, Sparrow D,
Mohun T and Mercola M: Subdivision of the cardiac Nkx2.5 expression
domain into myogenic and nonmyogenic compartments. Dev Biol.
218:326–340. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Son HY, Lee S, Tak EN, Cho HS, Shin HI,
Kim SH and Yang JH: Perfluorooctanoic acid alters T lymphocyte
phenotypes and cytokine expression in mice. Environ Toxicol.
24:580–588. 2009. View Article : Google Scholar
|
|
29
|
Van den Heuvel LP, Van den Born J, Jalanko
H, Schröder CH, Veerkamp JH, Assmann KJ, Berden JH, Holmberg C,
Rapola J and Monnens LA: The glycosaminoglycan content of renal
basement membranes in the congenital nephrotic syndrome of the
Finnish type. Pediatr Nephrol. 6:10–15. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yeung LW, Yamashita N, Taniyasu S, Lam PK,
Sinha RK, Borole DV and Kannan K: A survey of perfluorinated
compounds in surface water and biota including dolphins from the
Ganges River and in other waterbodies in India. Chemosphere.
76:55–62. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shi X, Du Y, Lam PK, Wu RS and Zhou B:
Developmental toxicity and alteration of gene expression in
zebrafish embryos exposed to PFOS. Toxicol Appl Pharmacol.
230:23–32. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Abbott BD, Wolf CJ, Schmid JE, Das KP,
Zehr RD, Helfant L, Nakayama S, Lindstrom AB, Strynar MJ and Lau C:
Perfluorooctanoic acid induced developmental toxicity in the mouse
is dependent on expression of peroxisome proliferator activated
receptor-alpha. Toxicol Sci. 98:571–581. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
O'Brien JM, Crump D, Mundy LJ, Chu S,
McLaren KK, Vongphachan V, Letcher RJ and Kennedy SW: Pipping
success and liver mRNA expression in chicken embryos exposed in ovo
to C8 and C11 perfluorinated carboxylic acids and C10
perfluorinated sulfonate. Toxicol Lett. 190:134–139. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Verreault J, Houde M, Gabrielsen GW,
Berger U, Haukås M, Letcher RJ and Muir DC: Perfluorinated alkyl
substances in plasma, liver, brain, and eggs of glaucous gulls
(Larus hyper-boreus) from the Norwegian arctic. Environ Sci
Technol. 39:7439–7445. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Conder JM, Hoke RA, De Wolf W, Russell MH
and Buck RC: Are PFCAs bioaccumulative? A critical review and
comparison with regulatory criteria and persistent lipophilic
compounds. Environ Sci Technol. 42:995–1003. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Garuti R, Jones C, Li WP, Michaely P, Herz
J, Gerard RD, Cohen JC and Hobbs HH: The modular adaptor protein
autosomal recessive hypercholesterolemia (ARH) promotes low density
lipoprotein receptor clustering into clathrin-coated pits. J Biol
Chem. 280:40996–41004. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Goldstein JL and Brown MS: Molecular
medicine. The cholesterol quartet. Science. 292:1310–1312. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang Q, Fang C, Wu X, Fan J and Dong S:
Perfluorooctane sulfonate impairs the cardiac development of a
marine medaka (Oryzias melastigma). Aquat Toxicol. 105:71–77. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jiang Q, Lust RM, Strynar MJ, Dagnino S
and DeWitt JC: Perflurooctanoic acid induces developmental
cardiotoxicity in chicken embryos and hatchlings. Toxicology.
293:97–106. 2012. View Article : Google Scholar : PubMed/NCBI
|