Introduction
Drug-induced liver injury (DILI) is the most
frequent cause of acute liver failure (1); of various drugs, acetaminophen
(APAP) overdose accounts for approximately one-half of all cases of
acute liver failure in the USA and the UK (2). Acute liver failure suddenly affects
young, otherwise healthy individuals, and is associated with an
extremely high rate of mortality, and is the most frequent cause of
emergency liver transplantation (1). Drug-induced hepatotoxicity is
usually caused by the formation of drug metabolites, which are
often formed by the action of cytochrome P450 2E1 (CYP2E1; EC
1.14.13.n7), which transforms various chemicals into reactive
metabolites. For example, APAP is converted by CYP2E1 into
N-acetyl-p-benzoquinone imine (NAPQI), an electrophilic metabolite
that binds to cysteine groups in proteins (3), depletes glutathione (GSH), and
causes respiratory dysfunction and reactive oxidative stress
(4). Since finding a matching
donor for liver transplantation is not always easy, novel therapies
for treating DILI are urgently needed.
The proteasome is a multisubunit enzyme complex that
degrades ubiquitin-tagged proteins; it plays a critical role in the
regulation of proteins that control cell-cycle progression and
apoptosis. Consequently, it has become an important target for
anticancer therapy (5). In in
vitro experiments and in animal studies, the inhibition of the
proteasome, either alone or in combination with conventional
chemotherapeutic agents, demonstrated antitumour effects against
numerous tumour types (5). In
2008, bortezomib (VELCADE; formerly, PS-341, LDP-341, and MLN341)
was approved by the US Food and Drug Administration (FDA) as a
therapeutic agent for multiple myeloma (6). Recently, the therapeutic potential
of bortezomib has been re-evaluated, and it has been reported that
the compound is therapeutically effective for various diseases,
such as tumours (7), congenital
erythropoietic porphyria (8), and
graft-versus-host disease (9).
To investigate the role of the proteasome and the
effects of its inhibition on DILI, we examined the effects of two
proteasome inhibitors, bortezomib and MG132, on drug- and
chemical-induced hepatotoxicity. Interestingly, bortezomib
alleviated APAP-induced hepatotoxicity, whereas MG132 had the
opposite effect. In addition, bortezomib treatment decreased liver
damage induced by CCl4 or thioacetamide (TAA) and
significantly decreased hepatic CYP2E1 transcription, leading to
diminished enzyme activity. Results of the present study suggest
that clinical treatment with bortezomib may be useful for
alleviating DILI and possibly other forms of acute liver
disease.
Materials and methods
Materials
Bortezomib was purchased from Biovision (Mountain
View, CA, USA). APAP, CCl4, TAA, MG132, sodium
4-phenylbutyrate (4-PBA), and 2-aminoethyl diphenylborinate (2-APB)
were purchased from Sigma-Aldrich (St. Louis, MO, USA). The primary
antibodies used in this study were anti-CYP2E1 (AB1252; Millipore,
Bedford, MA, USA), anti-connexin 32 (CX32; 35-8900; Invitrogen,
Carlsbad, CA, USA), anti-glyceraldehyde-3-phosphate dehydrogenase
(GAPDH; MAB374) (Millipore, Billerica, MA, USA), anti-binding
immunoglobulin protein (BiP; 3177; Cell Signaling Technology,
Beverly, MA, USA), anti-CCAAT-enhancer-binding protein homologous
protein (CHOP; 5554; Cell Signaling Technology) and anti-β-actin
(A5316; Sigma-Aldrich).
Animals
Male C57BL/6J mice, which were 6–8 weeks of age,
were purchased from Orient Bio, Inc. (Seoul, Korea) and housed
under special pathogen-free conditions. All animals were treated in
accordance with the Animal Care Guidelines of Ewha Womans
University. To induce hepatotoxicity, mice were injected
intraperitoneally with the following: APAP (350 or 500 mg/kg), TAA
(200 mg/kg), or CCl4 (2 ml/kg), as described previously
(10). Before being injected,
mice were fasted overnight. To inhibit the proteasome, bortezomib
was injected twice: first at 12 h (1 mg/kg) and then at 1 h (1
mg/kg) prior to the injection of APAP, as previously described
(11). The inhibitor MG132 was
injected (5 mg/kg) twice: first at 12 h and then at 1 h prior to
the administration of APAP, as previously described (12,13). In some cases, different doses of
bortezomib (0-1 mg/kg) were injected twice: first at 12 h and then
at 1 h prior to the administration of APAP, and 1 mg/kg bortezomib
was injected at different times prior to the administration of
APAP. For the inhibition of gap junctions, 2-APB (20 mg/kg) was
injected 2 h prior to the administration of APAP. DMSO-treated mice
were used as the relevant control group. Each group consisted of
4–6 mice. For liver extraction, the mice were sacrificed at 0, 2, 4
6 h after the APAP injection and, at 24 h after the TAA or
CCl4 injection. The livers were then perfused with PBS
to remove the blood via portal vein.
Cell culture
Human hepatocarcinoma Hep3B cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
(v/v) heat-inactivated foetal bovine serum, penicillin, and
streptomycin (Gibco, Carlsbad, CA, USA). Cells were maintained at
37°C in a humidified atmosphere containing 5% CO2. Prior
to RNA extraction for expression studies or luciferase assays,
cells were treated with bortezomib (10-250 nM) (14,15) in serum-free medium for 15 h. To
reduce ER stress, 4-PBA (1–5 mM) was added 1 h before bortezomib
treatment.
Serum enzyme marker measurement
Serum aspartate aminotransferase (AST) and alanine
aminotransferase (ALT) levels were measured by the Korean Animal
Clinical Research Center (Guri, Korea) using a Hitachi 7020
automatic biochemical analyser (Hitachi, Tokyo, Japan).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total mRNA from the liver tissues was extracted
using an RNeasy Mini kit (Qiagen, Valencia, CA, USA), and cDNA was
prepared from the mRNA using a Verso cDNA synthesis kit (Thermo
Fisher Scientific, Waltham, MA, USA) according to the
manufacturer's instructions. Primer sets are described in Table I. Relative gene expression was
calculated as 2−ΔΔCt by quantitative PCR, using the SYBR
Green PCR Master Mix (Applied Biosystems, Warrington, UK) and an
ABI PRISM 7500 Sequence Detection system (Applied Biosystems)
(16,17).
 | Table IPrimers used for quantitative
PCR. |
Table I
Primers used for quantitative
PCR.
| Species | Gene | Primer sequences
(5′→3′) | Refs. |
|---|
| Mouse | CYP2E1 | F:
GTTGCCTTGCTTGTCTGGAT | |
| | R:
AGGAATTGGGAAAGGTCCTG | |
| BiP | F:
TCATCGGACGCACTTGGAA | (50,51) |
| | R:
CAACCACCTTGAATGGCAAGA | |
| CHOP | F:
GTCCCTAGCTTGGCTGACAGA | (50,51) |
| | R:
TGGAGAGCGAGGGCTTTG | |
| CX32 | F:
TGGTCCCTGCAGCTTATCTT | (10) |
| | R:
CCTCAAGCCGTAGCATTTTC | |
| CX43 | F:
ATCCAAAGACTGCGGATCTC | (10) |
| | R:
GACCAGCTTGTACCCAGGAG | |
| GAPDH | F:
CACTCTTCCACCTTCGATGC | |
| | R:
CCCTGTTGCTGTAGCCGTAT | |
Western blot analysis
Liver tissues were lysed in
radioimmu-noprecipitation assay buffer [50 mM Tris-HCl, pH 7.5, 150
mM NaCl, 1% (v/v) Nonidet P-40, 0.5% (w/v) sodium deoxycholate,
0.1% (w/v) SDS] containing 50 mM NaF, 2 mM
Na3VO4, and protease and phosphatase
inhibitors (Sigma-Aldrich). Protein concentration was quantified
using Bradford assay reagent (Bio-Rad Laboratories, Hercules, CA,
USA). Protein samples (40 µg) were subjected to
SDS-polyacrylamide gel [8% (w/v)] electrophoresis and then
transferred to nitrocellulose membranes. Membranes were incubated
with primary antibodies followed by peroxidase-conjugated secondary
antibodies (111-035-003 and 115-035-003; Jackson ImmunoResearch,
West Grove, PA, USA) after blocking with 5% (w/v) BSA in 20 mM
Tris-HCl at pH 7.5, 500 mM NaCl, and 0.1% (v/v) Tween-20. The
immunocomplexes were detected by chemiluminescence using
SuperSignal West Pico chemiluminescent substrate (Thermo Fisher
Scientific) and detected with a Bio-Imaging Analyzer (LAS-4000;
Fuji, Tokyo, Japan).
Hematoxylin and eosin (H&E)
staining
Liver tissues were fixed in 4% (w/v)
paraformaldehyde, embedded in paraffin, sectioned at 4-µm
thickness, and then stained with H&E using standard methods.
Briefly, following deparafinization, the sections were stained with
hematoxylin (Sigma-Aldrich) and then destained with acid ethanol (1
ml hydrochloric acid + 99 ml 70% ethanol). After washing with tap
water, sections were stained with eosin (Sigma-Aldrich).
Measurement of GSH levels
Levels of GSH in the fresh liver were measured using
a GSH assay kit (Biovision) according to the manufacturer's
instructions. Briefly, 40 mg of liver tissues were homogenized with
cold glutathione assay buffer, and 6 N perchloric acid was added.
The samples were then precipitated with 3 N potassium hydroxide,
and then centrifuged for 2 min at 13,000 x g. The neutralized
samples were incubated with the o-phthalaldehyde probe and
glutathione assay buffer for 40 min to detect reduced GSH.
Fluorescence was deteced using a microplate fluorescence reader
(BioTek Synergy H1; BioTek, Winooski, VT, USA).
Measurement of CYP2E1 enzyme
activity
In order to measure CYP2E1 activity, the rate of
transformation of p-nitrophenol to p-nitrocatechol
was analysed with isolated hepatic microsomes, as described
previously (18,19). Briefly, the liver microsomes were
incubated at 37°C in assay buffer containing potassium phosphate
buffer (50 mM, pH 7.4), NADPH (1 mM) and p-nitrophenol (0.1
mM), at the final concentrations indicated. After 20 min, the
reactions were terminated by the addition of 0.6 N perchloric acid,
and centrifuged at 10,000 × g for 5 min. Subsequently, 6 N sodium
hydroxide was added to the supernatant, and the absorbance was
measured at 546 nM using a microplate reader (BioTek Synergy H1;
BioTek).
Reporter plasmid construct containing
CYP2E1 gene promoter
In the present study, a 540-bp human CYP2E1 gene
promoter using 5′-TAGGTACCCAGAAGTGAGATTCCTGTTCT-3′ and
5′-CCCAAGCTTTGCCGATGGGGCTCCACTCT-3′ as primers was subcloned into
the corresponding restriction sites of the luciferase reporter
pGL3-basic vector (Promega, San Luis Obispo, CA, USA), as described
in a previous study (20). The
underlined letters denote the corresponding restriction sites,
KpnI and HindIII The subcloned sequences were
verified by DNA sequence analyses.
Luciferase assay
The luciferase assay was performed with the
Dual-Luciferase Reporter assay system (Promega) as previously
described (21). Reporter plasmid
containing the CYP2E1 promoter region was transfected into Hep3B
cells using Metafectene transfection reagent (Biontex, San Diego,
CA, USA USA), and pRL-cytomegalovirus promoter (Renilla
luciferase vector; Promega) was co-transfected to normalise the
transfection efficiency. If necessary, pcDNA5/FRT/HNF-1α, obtained
from Addgene (Addgene plasmid 31104), or its control vector
pcDNA5/FRT/TO was transfected. After 24 h, each well was treated
with bortezomib or 4-PBA, as indicated in the figure legends. Cells
were lysed using Promega lysis buffer (Promega). Firefly and
Renilla luciferase activities were measured sequentially in
the same sample using a GloMax™ 20/20 Luminometer (Promega).
Luciferase activities were normalised by dividing the firefly
luciferase activity by the Renilla luciferase activity.
Statistical analysis
Values are expressed as the means ± SEM. Statistical
significance was determined using the Student's t-test, and a
p-value <0.05 was considered to indicate a statistically
significant difference.
Results
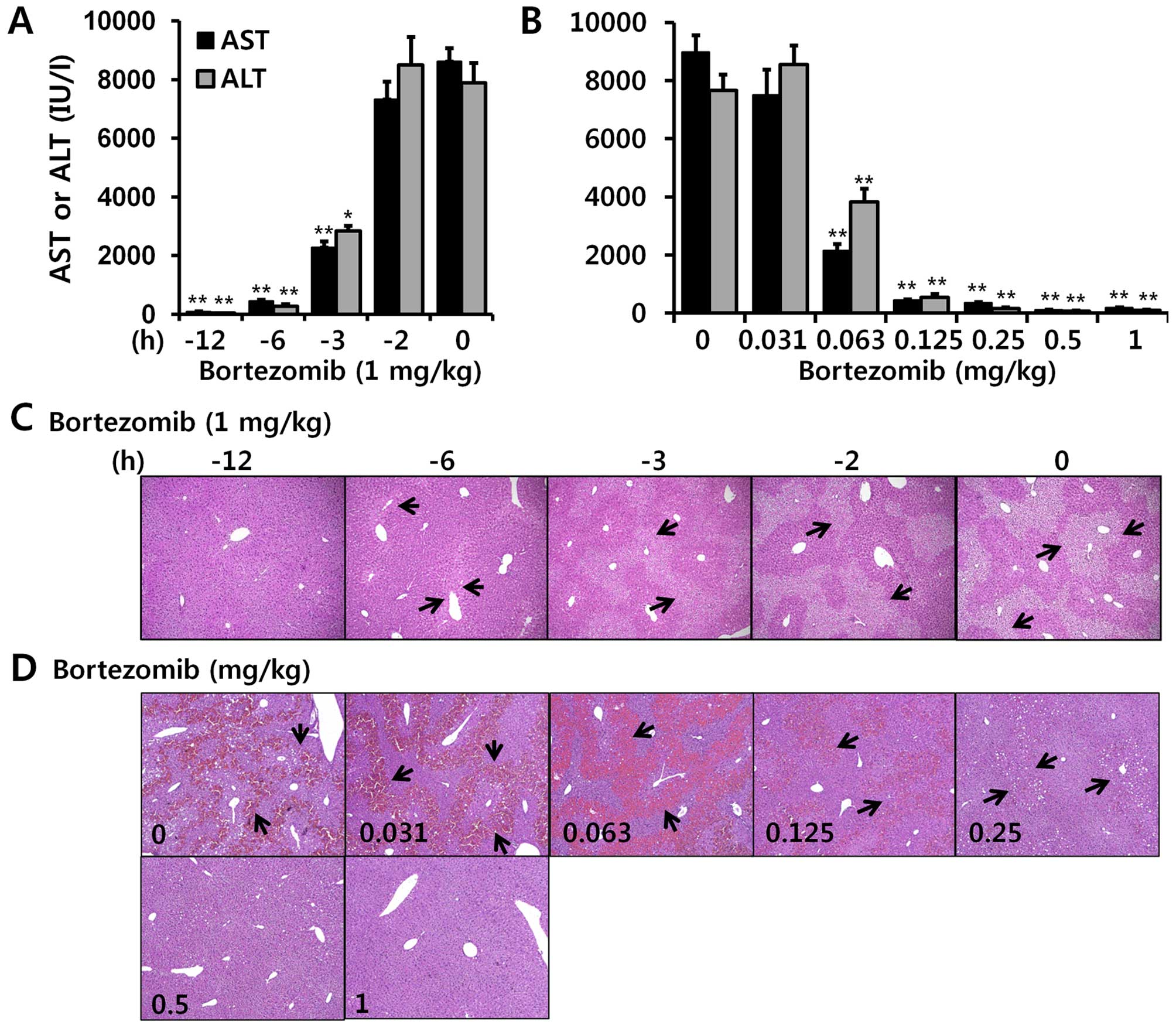
Bortezomib treatment alleviates
APAP-induced hepatotoxicity in a time- and dose-dependent
manner
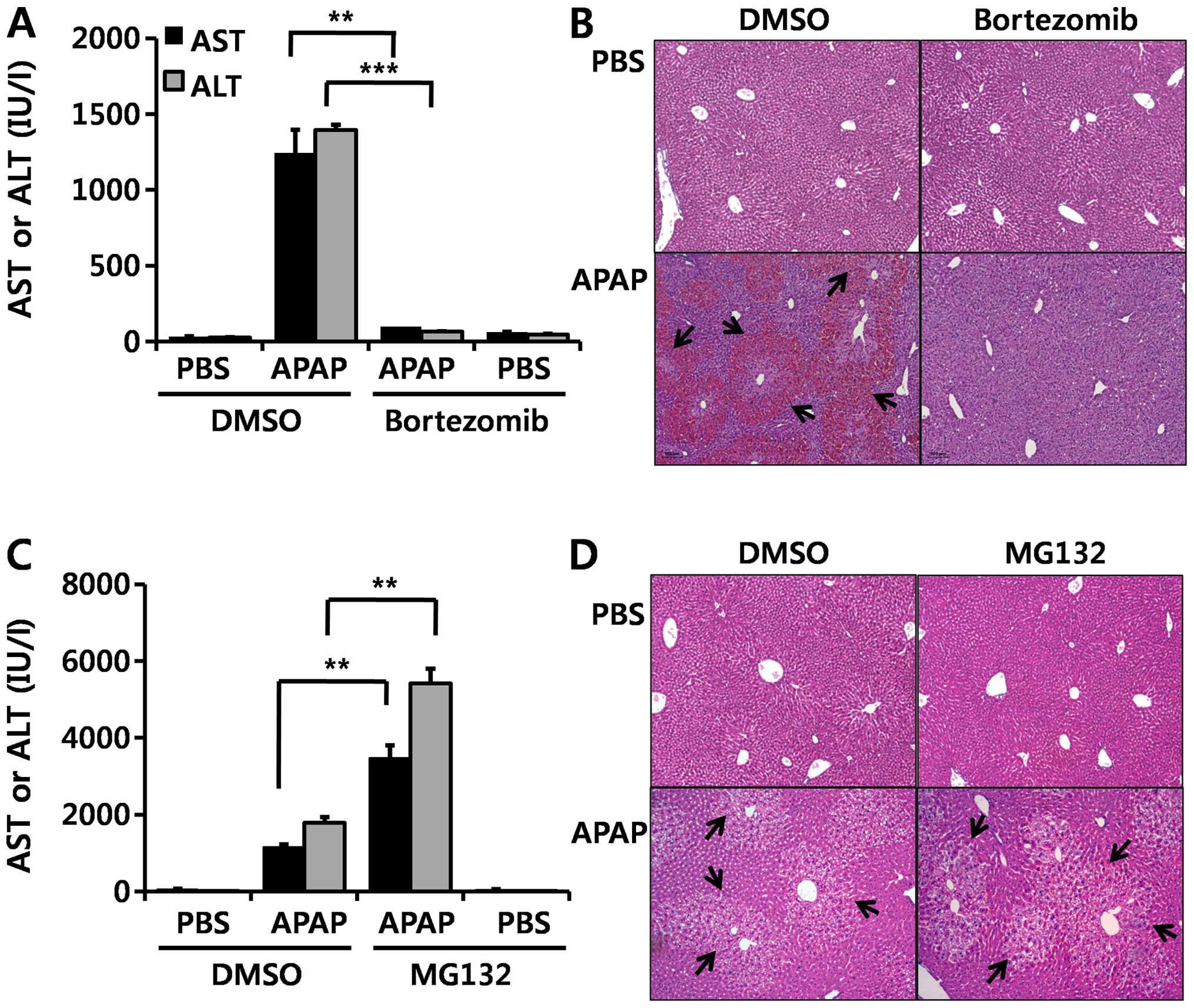
In order to determine the effect of bortezomib on
DILI induced by APAP, we injected bortezomib intraperitoneally
twice: first at different time points (1 mg/kg) (as indicated in
Fig. 1A) and then at 1 h (1
mg/kg) prior to the administration of APAP. Injecting 500 mg/kg
APAP markedly elevated serum AST and ALT (Fig. 1). When bortezomib was injected 2 h
before APAP administration, serum AST and ALT levels were greatly
elevated, similar to the DMSO-treated group (Fig. 1A). Treatment with bortezomib for
more than 3 h was needed to diminish the APAP-induced
hepatotoxicity, as indicated by decreased serum AST and ALT levels
and H&E staining (Fig. 1A and
C). In addition, bortezomib decreased APAP-induced
hepatotoxicity in a dose-dependent manner, and this protective
effect was noted at concentrations greater than 0.063 mg
bortezomib/kg (Fig. 1B and
D).
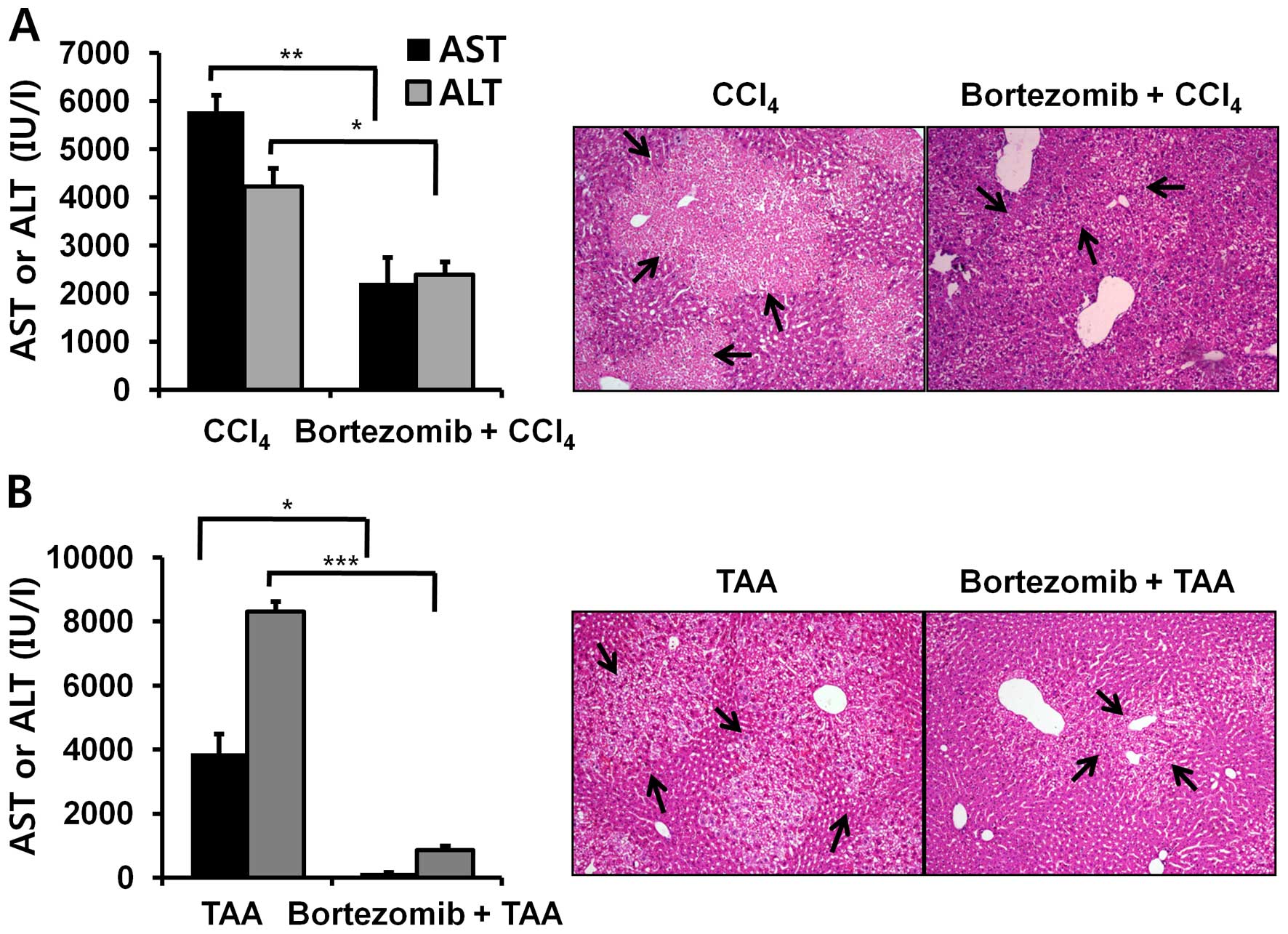
Bortezomib treatment diminishes
hepatotoxicity induced by other chemicals
Since the toxic mechanism differs between various
drugs and chemicals, we investigated whether bortezomib treatment
also protects against liver damage caused by other chemicals.
Surprisingly, bortezomib treatment also alleviated CCl4-
and TAA-induced hepatotoxicity (Fig.
2). Thus, we posit that the common toxic mechanism of all three
chemicals involves CYP2E1 (22–24) or gap junction function (25).
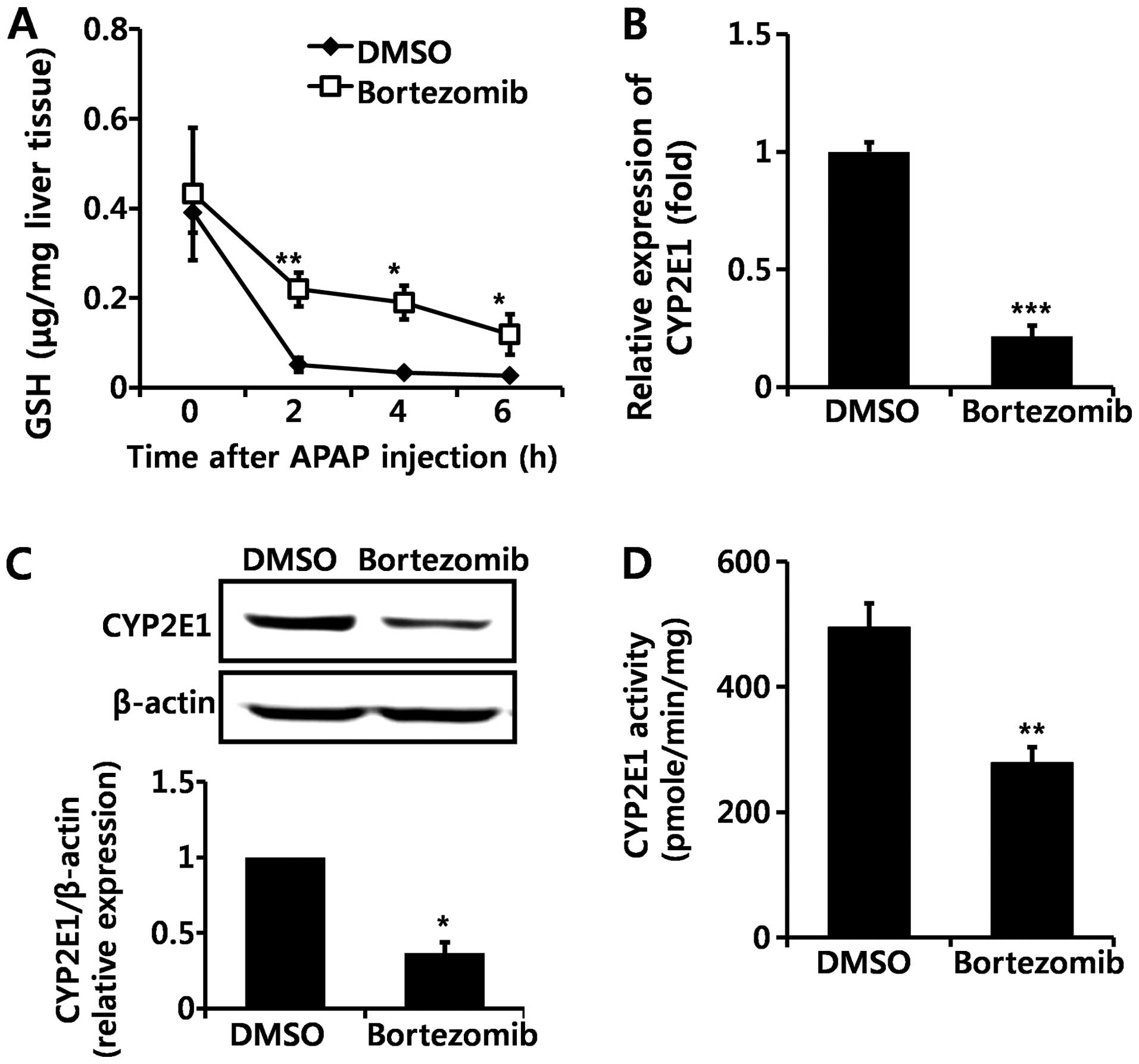
Bortezomib decreases CYP2E1 enzyme
expression and activity
GSH is an important antioxidant that detoxifies
NAPQI by conjugation, and GSH depletion leads to toxic cascades of
NAPQI (3,4). Basal levels of GSH did not differ
significantly between DMSO- and bortezomib-treated mice (Fig. 3A). Although GSH was markedly
decreased in both DMSO- and bortezomib-treated groups following
APAP administration, GSH levels remained higher at all time-points
in bortezomib-versus DMSO-treated animals (Fig. 3A). Lower GSH depletion in
bortezomib-treated mice implies the decreased formation of NAPQI.
Since NAPQI is transformed by CYP2E1, we subsequently measured
CYP2E1 expression by both RT-qPCR (Fig. 3B) and western blot analysis
(Fig. 3C). Bortezomib treatment
significantly decreased CYP2E1 transcription (Fig. 3B), which resulted in lower CYP2E1
protein levels (Fig. 3C). In
addition, CYP2E1 enzymatic activity was significantly lower in the
livers of bortezomib-treated versus control mice (Fig. 3D). Therefore, we conclude that the
decreased CYP2E1 enzyme activity caused by bortezomib
administration alleviates liver damage induced by APAP,
CCl4, and TAA exposure. This is in agreement with
results of previous studies, in which a CYP2E1 deficiency prevented
hepatotoxicity induced by these three chemicals (22–24).
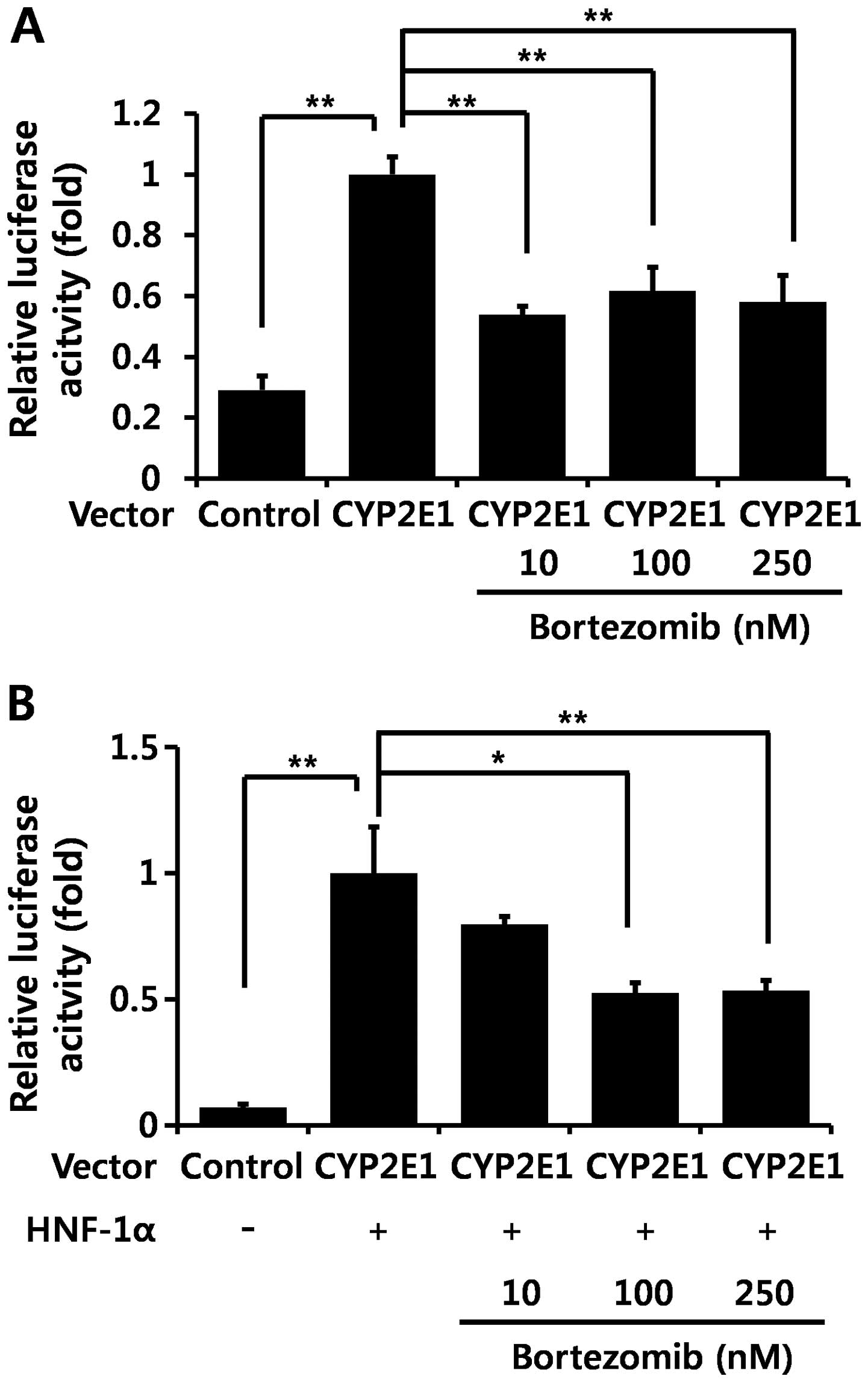
Bortezomib diminishes hepatocyte nuclear
factor (HNF)-1α-induced CYP2E1 transcription
In order to confirm whether bortezomib regulates
CYP2E1 transcription, we transfected a hepatocyte cell line (Hep3B
cells) with the reporter plasmid containing the CYP2E1 gene
promoter region, and performed luciferase assays. Bortezomib
treatment significantly decreased CYP2E1 promoter activity by
itself (Fig. 4A) as well as
HNF-1α-induced CYP2E1 promoter activity (Fig. 4B). HNF-1α is the main
transcription factor which positively regulates CYP2E1 gene
transcription (26–28).
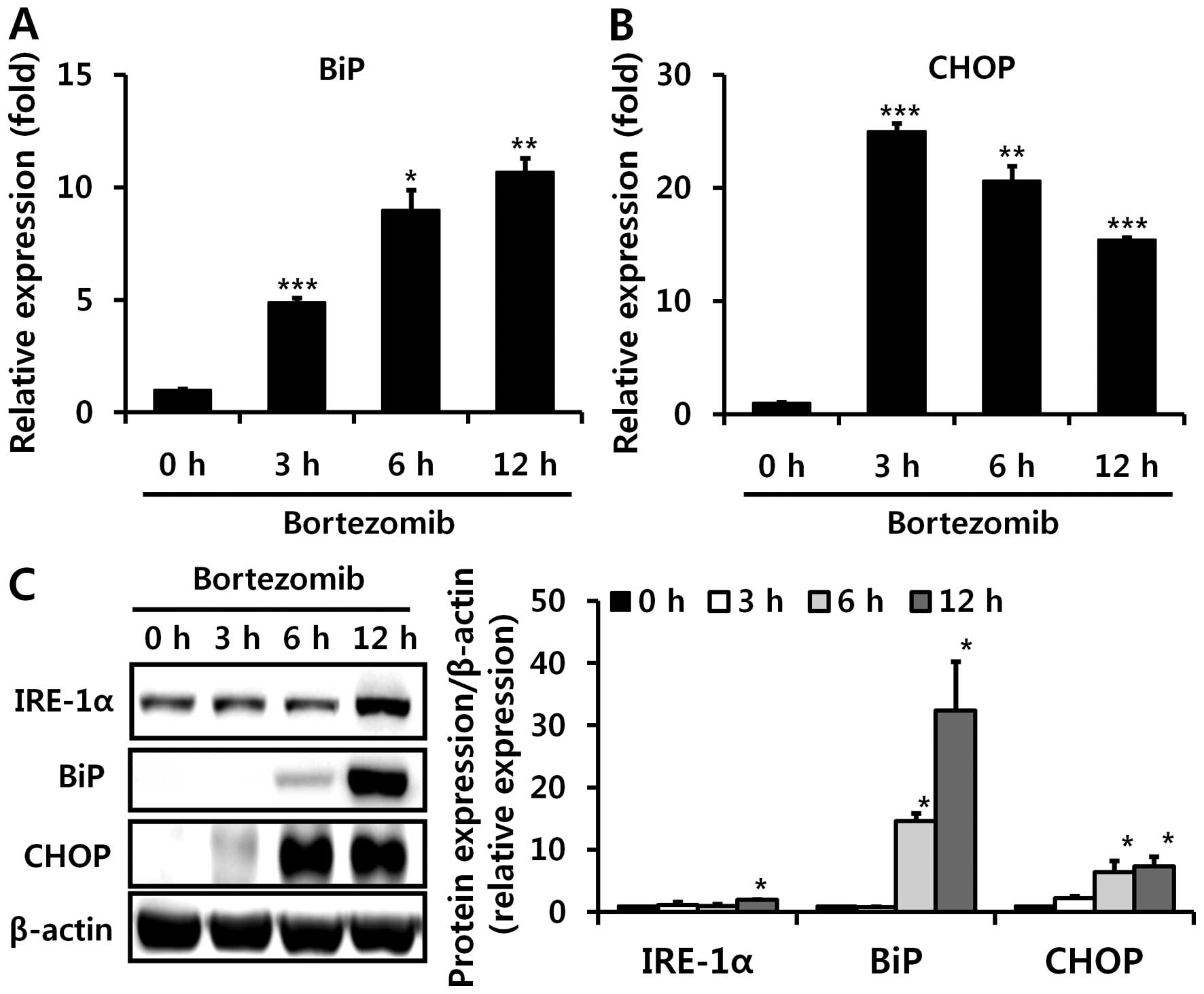
Bortezomib induces endoplasmic reticulum
(ER) stress in the liver, which partially contributes to decreased
CYP2E1 levels
Since bortezomib can lead to ER stress in
vitro (29,30), which can induce decreased CYP2E1
mRNA levels (31), we next
investigated whether bortezomib administration also causes ER
stress in the liver in vivo. We examined altered levels of
several proteins that were reported to be induced during ER stress.
BiP, also referred to as glucose-regulated protein 78, is a central
regulator of ER stress (32), and
CHOP is one of the most highly induced transcription factors during
ER stress (33). Bortezomib
injections markedly elevated hepatic mRNA and protein levels of BiP
and CHOP, indicating the in vivo induction of ER stress
(Fig. 5). IRE-1α protein
expression was also increased 12 h after bortezomib treatment
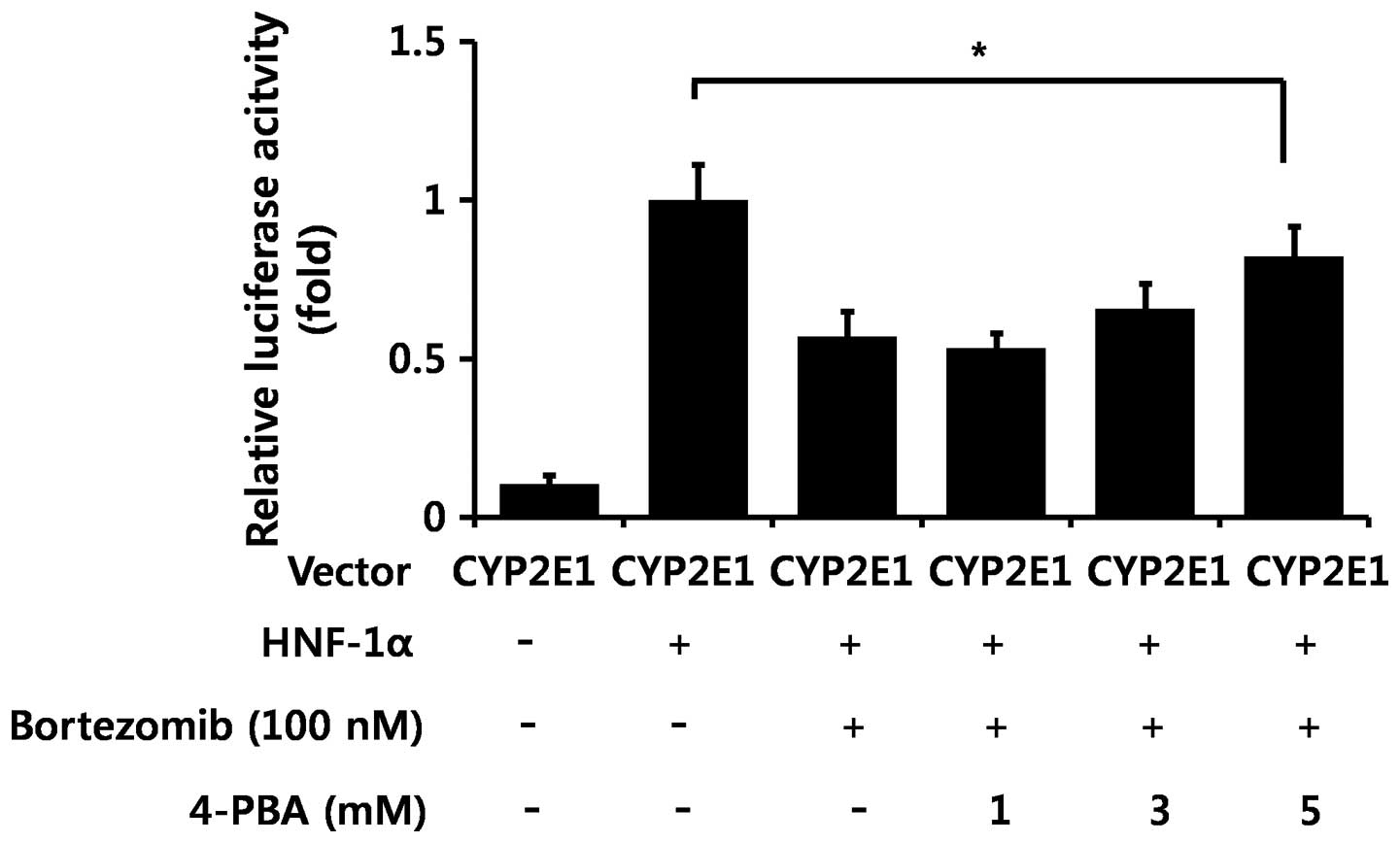
(Fig. 5C). Subsequently, we used
4-PBA to confirm whether ER stress is directly involved in
bortezomib-induced CYP2E1 decrement. Treatment with 4-PBA (5 mM),
which is reported to decrease ER stress (34), partially recovered CYP2E1
luciferase activity decreased by bortezomib treatment (Fig. 6). Therefore, bortezomib-induced ER
stress partially contributes to decreased CYP2E1 levels.
Protective effect of bortezomib on
APAP-induced hepatotoxicity is not derived from direct inhibition
of the proteasome
In the present study, in order to investigate
whether this effect of bortezomib is due to proteasome inhibition,
we injected MG132, another well-known proteasome inhibitor, before
APAP administration. By contrast to bortezomib treatment (Fig. 7A and B), MG132 aggravated
APAP-induced hepatotoxicity, as demonstrated by elevated serum AST
and ALT levels (Fig. 7C) and by
enhanced H&E staining (Fig.
7D). These results suggest that the protective effect of
bortezomib on APAP-induced hepatotoxicity is not derived from
direct inhibition of the proteasome.
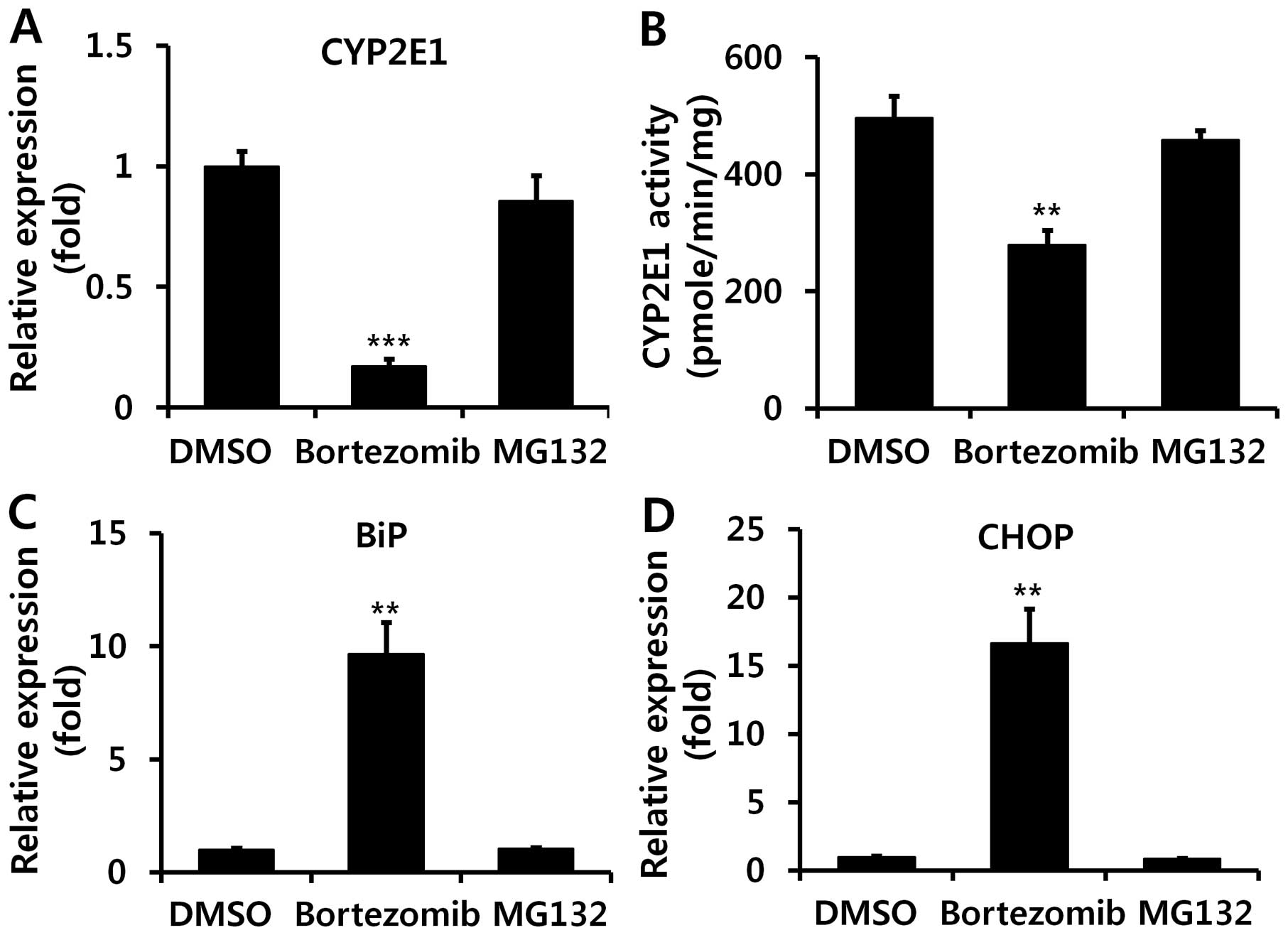
Proteasome inhibitor MG132 does not
induce hepatic ER stress
Since we noted that MG132, a second proteasome
inhibitor, aggravated APAP-induced liver injury, by contrast with
bortezomib (Fig. 1), we
subsequently examined the effect of MG132 on CYP2E1 expression and
ER stress. Unlike treatment with bortezomib, hepatic mRNA levels of
CYP2E1 were not diminished upon MG132 administration (Fig. 8A), and we also noted relatively
unaltered CYP2E1 enzyme activity upon administration of MG132
(Fig. 8B). In addition, it was
demonstrated that MG132 administration did not induce ER stress, as
indicated by unaltered mRNA levels of BiP and CHOP (Fig. 8C and D).
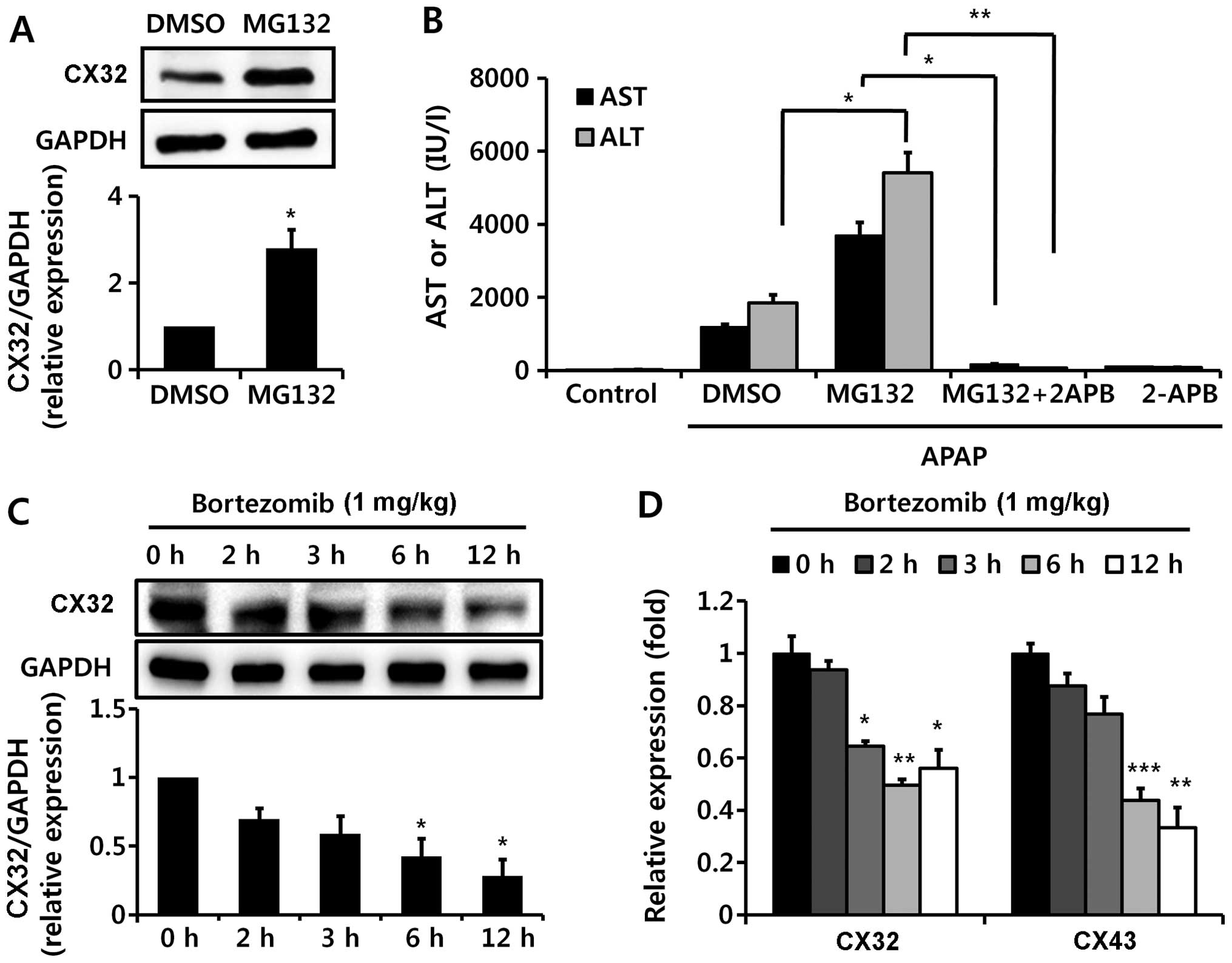
MG132, but not bortezomib, increases CX32
levels, which increases hepatotoxicity
In addition to CYP2E1 enzyme activity, it has
previously been suggested that the common toxic mechanism of all
three chemicals (APAP, TAA, and CCl4) may also involve
gap junctions (10,25,35,36), since gap junctions have been
reported to play a critical role in the propagation of
hepatotoxicity of all three chemicals used in the present study;
ablation of CX32 (a key protein in hepatic gap junctions)
significantly protects against DILI (10,25,35,36). Thus, we examined CX32 levels.
Notably, MG132 administration significantly increased CX32 protein
expression (Fig. 9A), possibly by
inhibiting its degradation, and 2-APB, a gap junction inhibitor,
abolished MG132-aggravated APAP-induced liver damage (Fig. 9B). Unexpectedly, bortezomib
decreased CX32 expression, as well as CX43 mRNA levels (Fig. 9C and D). Therefore, the opposite
effects of bortezomib and MG132 on CX32 levels may contribute to
the different impacts of bortezomib and MG132 on DILI.
Discussion
Protein metabolism, including both synthesis and
degradation, is crucial for cellular homeostasis. Eukaryotic cells
possess three different systems which are necessary for protein
degradation: mitochondrial proteases, which degrade the majority of
mitochondrial proteins; lysosomal proteases, which degrade membrane
and endocytosed proteins; and the ubiquitin-proteasome system,
which degrades the vast majority (80–90%) of intracellular proteins
(37). The proteasome system has
emerged as a master regulator of diverse cellular processes,
including cell cycle, survival, and apoptosis, and plays a critical
role in many diseases, such as cancer (38) and neurodegenerative (39) and cardiovascular diseases
(37). In the present study, we
demonstrated that the FDA-approved proteasome inhibitor bortezomib
is effective at preventing drug- and chemical-induced acute liver
injury via the regulation of CYP2E1 expression.
Hepatic CYP2E1 levels were markedly decreased upon
bortezomib treatment. Since CYP2E1 degradation involves the
ubiquitin-proteasome pathway (40,41), the protective effect of bortezomib
on drug- and chemical-induced liver injury seems attributable to a
different mechanism than proteasome inhibition. Although bortezomib
was originally developed as a proteasome inhibitor, other distinct
mechanisms of bortezomib have been uncovered. One of these is ER
stress (29,30), which, in turn, induces
IRE-1α-mediated degradation of CYP2E1 mRNA (31). In the present study, ER stress
induced by bortezomib treatment also played a role in decreased
CYP2E1 transcription, which may alleviate DILI. The main governing
factor of CYP2E1 transcriptional regulation is HNF-1α (26–28), and we noted that bortezomib
significantly decreased HNF-1α-induced promoter activation of
CYP2E1, and 4-PBA treatment partially recovered bortezomib's
inhibition of CYP2E1 promoter activity. Therefore, ER stress plays
a role in bortezomib-induced CYP2E1 decrement. Although MG132 was
reported to induce ER stress in vitro (42), its administration in vivo
did not cause ER stress. Although questions such as whether the
relatively long half-life of bortezomib (43) contributes to the different
phenotype in vivo compared with MG132, or, rather, a
distinct mechanism of bortezomib other than proteasome inhibition
may exist, require further study, the protective effect of
bortezomib on DILI is noteworthy, considering its current clinical
use.
Interestingly, the effects of two well-known
proteasome inhibitors, bortezomib and MG132, exerted opposite
effects on APAP-induced liver injury. Since proteasome inhibition
potentiates CYP2E1-mediated toxicity in HepG2 cells by elevating
CYP2E1 levels (44), the
aggravating effects of MG132 on APAP-induced liver damage are
predictable. However, since no marked change was observed in CYP2E1
enzyme activity, the aggravating effect of MG132 on APAP-induced
liver damage appears to be due to some other mechanism, such as gap
junction elevation; this was confirmed by elevation of CX32 levels.
Gap junctions have recently been implicated as important players in
amplifying DILI (25,36). Gap junction channels are composed
of CX proteins and play an important role in intercellular
communication and in the propagation of liver toxicity and
inflammation (10,25,36). Ablation or decreased levels of
CX32, a major hepatic gap junction protein, protected against DILI
which was induced by the same three hepatotoxic agents used in the
present study (10,25,36); thus, diminished CX32 levels caused
by bortezomib treatment protect against DILI, and, on the contrary,
the increased CX32 levels caused by MG132 are expected to exert
opposite effects on DILI. Gap junction proteins are degraded by the
proteasome system; thus, proteasome inhibition usually increases CX
levels (45–47). On the other hand, ER stress
decreases CX43 expression at both the protein and mRNA levels by
inhibiting CX43 promoter activity and accelerating the degradation
of CX43 (48). Therefore, we
suggest that the balance between proteasome inhibition and
induction of ER stress determines the effect of bortezomib on CX
levels. Decreased bortezomib-induced CX32 levels also contribute to
the protective effect on APAP-induced liver injury in addition to
decreasing levels of CYP2E1 enzymatic activity.
Since bortezomib is currently being used in clinical
settings, the protective effect of bortezomib on APAP-induced liver
injury has clinical importance despite its unclear mechanism.
Unfortunately, our study shows that bortezomib in mice was only
effective when treatments longer than 3 h were used before APAP
administration. Considering that the half-life of CYP2E1 protein is
6–7 h (41), the time needed for
bortezomib to transcriptionally diminish CYP2E1 expression makes
sense, but this finding weakens the practical usefulness of
bortezomib as a treatment for DILI, since many patients need
medical support when liver damage is already present (49). Still, it is possible for
bortezomib to be used to prevent DILI, as the use of high doses of
some drugs known to be metabolised by CYP2E1 is clinically
inevitable. In addition, bortezomib may help to increase the range
of drug dosage, which may otherwise be limited due to DILI.
Proteasome inhibition has recently emerged as an
effective therapeutic target in several human diseases. The present
study suggests that proteasome inhibition has different effects
with respect to DILI, depending on the specific drug employed.
Bortezomib, but not MG132, was effectively used for alleviating
drug- and chemical-induced liver injury in mice. Since ER stress,
induced by proteasome inhibition, has distinct effects, each
proteasome inhibitor should be individually scrutinised for its
therapeutic applicability in future studies.
Acknowledgments
This study was supported by a Gachon University
Research Grant from 2014 (GCU 2014-5101) and awards from the
National Research Foundation of Korea funded by the Korean
government (Ministry of Education, Science and Technology)
(NRF-2013R1A1A1057912).
Abbreviations:
|
DILI
|
drug-induced liver injury
|
|
ALT
|
alanine aminotransferase
|
|
APAP
|
acetaminophen
|
|
2-APB
|
2-aminoethyl diphenylborinate
|
|
AST
|
aspartate aminotransferase
|
|
BiP
|
binding immunoglobulin protein
|
|
CHOP
|
CCAAT-enhancer-binding protein
homologous protein
|
|
CX
|
connexin
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
ER
|
endoplasmic reticulum
|
|
FDA
|
US Food and Drug Administration
|
|
GAPDH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
|
GSH
|
glutathione
|
|
HNF
|
hepatocyte nuclear factor
|
|
NAPQI
|
N-acetyl-p-benzoquinone imine
|
|
4-PBA
|
sodium 4-phenyl-butyrate
|
|
TAA
|
thioacetamide
|
References
|
1
|
Bernal W, Auzinger G, Dhawan A and Wendon
J: Acute liver failure. Lancet. 376:190–201. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hinson JA, Roberts DW and James LP:
Mechanisms of acetaminophen-induced liver necrosis. Handb Exp
Pharmacol. 369–405. 2010. View Article : Google Scholar :
|
|
3
|
James LP, Alonso EM, Hynan LS, Hinson JA,
Davern TJ, Lee WM, Squires RH, Pediatric Acute, Liver Failure and
Study Group: Detection of acetaminophen protein adducts in children
with acute liver failure of indeterminate cause. Pediatrics.
118:e676–e681. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Burcham PC and Harman AW: Acetaminophen
toxicity results in site-specific mitochondrial damage in isolated
mouse hepatocytes. J Biol Chem. 266:5049–5054. 1991.PubMed/NCBI
|
|
5
|
Adams J: The proteasome: structure,
function, and role in the cell. Cancer Treat Rev. 29(Suppl 1): 3–9.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Richardson PG, Mitsiades C, Hideshima T
and Anderson KC: Bortezomib: proteasome inhibition as an effective
anticancer therapy. Annu Rev Med. 57:33–47. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Deming DA, Ninan J, Bailey HH, Kolesar JM,
Eickhoff J, Reid JM, Ames MM, McGovern RM, Alberti D, Marnocha R,
et al: A Phase I study of intermittently dosed vorinostat in
combination with bortezomib in patients with advanced solid tumors.
Invest New Drugs. 32:323–329. 2014. View Article : Google Scholar :
|
|
8
|
Blouin JM, Duchartre Y, Costet P, Lalanne
M, Ged C, Lain A, Millet O, de Verneuil H and Richard E:
Therapeutic potential of proteasome inhibitors in congenital
erythropoietic porphyria. Proc Natl Acad Sci USA. 110:18238–18243.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Z, Wu Q, Yan Z, Li D, Lu G, Mou W, Wu
S, Pan X, Lu Q and Xu K: The protection and therapy effects of
bortezomib in murine acute graft-versus-host disease. Transplant
Proc. 45:2527–2535. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Park WJ, Park JW, Erez-Roman R,
Kogot-Levin A, Bame JR, Tirosh B, Saada A, Merrill AH Jr,
Pewzner-Jung Y and Futerman AH: Protection of a ceramide synthase 2
null mouse from drug-induced liver injury: role of gap junction
dysfunction and connexin 32 mislocalization. J Biol Chem.
288:30904–30916. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wagner-Ballon O, Pisani DF, Gastinne T,
Tulliez M, Chaligné R, Lacout C, Auradé F, Villeval JL, Gonin P,
Vainchenker W and Giraudier S: Proteasome inhibitor bortezomib
impairs both myelofibrosis and osteosclerosis induced by high
thrombopoietin levels in mice. Blood. 110:345–353. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Carvalho AN, Marques C, Rodrigues E,
Henderson CJ, Wolf CR, Pereira P and Gama MJ: Ubiquitin-proteasome
system impairment and MPTP-induced oxidative stress in the brain of
C57BL/6 wild-type and GSTP knockout mice. Mol Neurobiol.
47:662–672. 2013. View Article : Google Scholar
|
|
13
|
Sun H, Kosaras B, Klein PM and Jensen FE:
Mammalian target of rapamycin complex 1 activation negatively
regulates Polo-like kinase 2-mediated homeostatic compensation
following neonatal seizures. Proc Natl Acad Sci USA. 110:5199–5204.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Oerlemans R, Franke NE, Assaraf YG, Cloos
J, van Zantwijk I, Berkers CR, Scheffer GL, Debipersad K, Vojtekova
K, Lemos C, et al: Molecular basis of bortezomib resistance:
proteasome subunit beta5 (PSMB5) gene mutation and overexpression
of PSMB5 protein. Blood. 112:2489–2499. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Markovina S, Callander NS, O'Connor SL,
Kim J, Werndli JE, Raschko M, Leith CP, Kahl BS, Kim K and Miyamoto
S: Bortezomib-resistant nuclear factor-kappaB activity in multiple
myeloma cells. Mol Cancer Res. 6:1356–1364. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Park JW, Park ES, Choi EN, Park HY and
Jung SC: Altered brain gene expression profiles associated with the
pathogenesis of phenylketonuria in a mouse model. Clin Chim Acta.
401:90–99. 2009. View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
18
|
Phillips IR and Shephard EA: Cytochrome
P450 Protocols. Humana Press; Totowa, NJ: 2006
|
|
19
|
Reinke LA and Moyer MJ: p-Nitrophenol
hydroxylation. A microsomal oxidation which is highly inducible by
ethanol. Drug Metab Dispos. 13:548–552. 1985.PubMed/NCBI
|
|
20
|
Matsunaga N, Ikeda M, Takiguchi T,
Koyanagi S and Ohdo S: The molecular mechanism regulating 24-hour
rhythm of CYP2E1 expression in the mouse liver. Hepatology.
48:240–251. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park JW, Lee MH, Choi JO, Park HY and Jung
SC: Tissue-specific activation of mitogen-activated protein kinases
for expression of transthyretin by phenylalanine and its
metabolite, phenylpyruvic acid. Exp Mol Med. 42:105–115. 2010.
View Article : Google Scholar :
|
|
22
|
Wong FW, Chan WY and Lee SS: Resistance to
carbon tetrachloride-induced hepatotoxicity in mice which lack
CYP2E1 expression. Toxicol Appl Pharmacol. 153:109–118. 1998.
View Article : Google Scholar
|
|
23
|
Kang JS, Wanibuchi H, Morimura K,
Wongpoomchai R, Chusiri Y, Gonzalez FJ and Fukushima S: Role of
CYP2E1 in thioacetamide-induced mouse hepatotoxicity. Toxicol Appl
Pharmacol. 228:295–300. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee SS, Buters JT, Pineau T,
Fernandez-Salguero P and Gonzalez FJ: Role of CYP2E1 in the
hepatotoxicity of acetaminophen. J Biol Chem. 271:12063–12067.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Patel SJ, Milwid JM, King KR, Bohr S,
Iracheta-Vellve A, Li M, Vitalo A, Parekkadan B, Jindal R and
Yarmush ML: Gap junction inhibition prevents drug-induced liver
toxicity and fulminant hepatic failure. Nat Biotechnol. 30:179–183.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ueno T and Gonzalez FJ: Transcriptional
control of the rat hepatic CYP2E1 gene. Mol Cell Biol.
10:4495–4505. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu SY and Gonzalez FJ: Role of the
liver-enriched transcription factor HNF-1 alpha in expression of
the CYP2E1 gene. DNA Cell Biol. 14:285–293. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cheung C, Akiyama TE, Kudo G and Gonzalez
FJ: Hepatic expression of cytochrome P450s in hepatocyte nuclear
factor 1-alpha (HNF1alpha)-deficient mice. Biochem Pharmacol.
66:2011–2020. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nawrocki ST, Carew JS, Dunner K Jr, Boise
LH, Chiao PJ, Huang P, Abbruzzese JL and McConkey DJ: Bortezomib
inhibits PKR-like endoplasmic reticulum (ER) kinase and induces
apoptosis via ER stress in human pancreatic cancer cells. Cancer
Res. 65:11510–11519. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Armstrong JL, Flockhart R, Veal GJ, Lovat
PE and Redfern CP: Regulation of endoplasmic reticulum
stress-induced cell death by ATF4 in neuroectodermal tumor cells. J
Biol Chem. 285:6091–6100. 2010. View Article : Google Scholar :
|
|
31
|
Hur KY, So JS, Ruda V, Frank-Kamenetsky M,
Fitzgerald K, Koteliansky V, Iwawaki T, Glimcher LH and Lee AH:
IRE1α activation protects mice against acetaminophen-induced
hepatotoxicity. J Exp Med. 209:307–318. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee AS: The ER chaperone and signaling
regulator GRP78/BiP as a monitor of endoplasmic reticulum stress.
Methods. 35:373–381. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Oyadomari S and Mori M: Roles of
CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ.
11:381–389. 2004. View Article : Google Scholar
|
|
34
|
Basseri S, Lhoták S, Sharma AM and Austin
RC: The chemical chaperone 4-phenylbutyrate inhibits adipogenesis
by modulating the unfolded protein response. J Lipid Res.
50:2486–2501. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tong X, Dong S, Yu M, Wang Q and Tao L:
Role of heteromeric gap junctions in the cytotoxicity of cisplatin.
Toxicology. 310:53–60. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Asamoto M, Hokaiwado N, Murasaki T and
Shirai T: Connexin 32 dominant-negative mutant transgenic rats are
resistant to hepatic damage by chemicals. Hepatology. 40:205–210.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Herrmann J, Ciechanover A, Lerman LO and
Lerman A: The ubiquitin-proteasome system in cardiovascular
diseases - a hypothesis extended. Cardiovasc Res. 61:11–21. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mani A and Gelmann EP: The
ubiquitin-proteasome pathway and its role in cancer. J Clin Oncol.
23:4776–4789. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ciechanover A and Brundin P: The ubiquitin
proteasome system in neurodegenerative diseases: sometimes the
chicken, sometimes the egg. Neuron. 40:427–446. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Goasduff T and Cederbaum AI:
NADPH-dependent microsomal electron transfer increases degradation
of CYP2E1 by the proteasome complex: role of reactive oxygen
species. Arch Biochem Biophys. 370:258–270. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Roberts BJ: Evidence of
proteasome-mediated cytochrome P-450 degradation. J Biol Chem.
272:9771–9778. 1997.PubMed/NCBI
|
|
42
|
Park HS, Jun Y, Han CR, Woo HJ and Kim YH:
Proteasome inhibitor MG132-induced apoptosis via ER stress-mediated
apoptotic pathway and its potentiation by protein tyrosine kinase
p56lck in human Jurkat T cells. Biochem Pharmacol. 82:1110–1125.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Levêque D, Carvalho MC and Maloisel F:
Review. Clinical pharmacokinetics of bortezomib. In Vivo.
21:273–278. 2007.PubMed/NCBI
|
|
44
|
Pérez MJ and Cederbaum AI: Proteasome
inhibition potentiates CYP2E1-mediated toxicity in HepG2 cells.
Hepatology. 37:1395–1404. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fernandes R, Girão H and Pereira P: High
glucose down-regulates intercellular communication in retinal
endothelial cells by enhancing degradation of connexin 43 by a
proteasome-dependent mechanism. J Biol Chem. 279:27219–27224. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Minogue PJ, Beyer EC and Berthoud VM: A
connexin50 mutant, CX50fs, that causes cataracts is unstable, but
is rescued by a proteasomal inhibitor. J Biol Chem.
288:20427–20434. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Laing JG, Tadros PN, Westphale EM and
Beyer EC: Degradation of connexin43 gap junctions involves both the
proteasome and the lysosome. Exp Cell Res. 236:482–492. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang T, Wan Y, Zhu Y, Fang X, Hiramatsu
N, Hayakawa K, Paton AW, Paton JC, Kitamura M and Yao J:
Downregulation of gap junction expression and function by
endoplasmic reticulum stress. J Cell Biochem. 107:973–983. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Larson AM: Acetaminophen hepatotoxicity.
Clin Liver Dis. 11:525–548. vi2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Farfel-Becker T, Vitner E, Dekel H, Leshem
N, Enquist IB, Karlsson S and Futerman AH: No evidence for
activation of the unfolded protein response in neuronopathic models
of Gaucher disease. Hum Mol Genet. 18:1482–1488. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hetz C, Lee AH, Gonzalez-Romero D, Thielen
P, Castilla J, Soto C and Glimcher LH: Unfolded protein response
transcription factor XBP-1 does not influence prion replication or
pathogenesis. Proc Natl Acad Sci USA. 105:757–762. 2008. View Article : Google Scholar : PubMed/NCBI
|