Introduction
Hepatocellular carcinoma (HCC) is the fifth most
common type of cancer and the third most common cause of
cancer-related death globally, and it is characterized by high
malignancy and poor prognosis (1). However, lack of sensitivity to
therapy and multidrug resistance have resulted in poor efficacy of
these treatments (2). Cancer
immunotherapy has been a major research focus and has been
highlighted by gene transfer, enhancing T-cell receptor-recognizing
tumor antigens and the effect of antibody-dependent cellular
cytotoxicity (ADCC). However, cancer immunotherapy has also failed
previously, suggesting that cancer immunotherapy acts very
differently from immunotherapy against infectious diseases
(3). Finding more effective ways
of administering immunotherapy is thus a priority.
Macrophages play an important role in immune
defense. Activated macrophages suppress tumor cells through the
ADCC pathway via FcγRIII (CD16) and directly or indirectly by
cytokines including tumor necrosis factor (TNF)-α and interleukin
(IL)-1 or certain enzymes (4). As
part of the mono-nuclear phagocytic system, Kupffer cells (KCs) are
the main type of hepatic macrophages, and they play an important
role in the inhibition of liver tumors and their surface receptors
are the mediator in this pathway (5).
CD16 is important for specific and non-specific
immunity, and includes two forms, CD16a and CD16b: CD16a is the
functional form of CD16 while CD16b is ancillary (6). As the specific receptor of IgG Fc,
CD16a forms a bridge between cytoimmunity and humoral immunity, and
has an important relationship with the maturation of dendritic
cells (DCs) (7). Antibody-based
protein scaffolds, immunoglobulin Fc fragments with antigen binding
(Fcab) sites, have been shown to hold significant promise as
next-generation protein therapeutics; in particular, Fcab (CD16a)
molecules have been shown to mediate ADCC reactivity (8). Previous research has shown that the
correlation between CD16a binding affinity and ADCC potency is also
valid in Fcab proteins and that antigen specific Fcab molecules can
be further engineered for fine tuning of immuno-effector functions
(9).
CD16a functions via myeloid cells and natural killer
(NK) cells, and mediates the ADCC of the mononucleus macrophage
system, which is Mg2+-dependent, to kill target cells
including tumor cells (10).
However, the relationship between CD16a, KCs and HCC was not fully
understood. The aim of the present study was to investigate whether
CD16a is present in KCs in HCC. Furthermore, the function and
variance of CD16a and also the ADCC pathway were investigated using
cultured tumor cells for in vitro analysis.
Materials and methods
Patients and specimens
Samples of cancerous tissues, para-cancerous tissues
(<10 mm distance from the tumor) and adjacent normal hepatic
tissues (>10 mm distance from the tumor) (11) were obtained from 87 patients with
primary HCC, in which there was no macroscopic tumor thrombi or
satellite nodules and the diameter of the tumor was <10 cm. A
total of 39 normal liver samples were obtained from patients who
underwent hepatectomy as a result of injury in the Department of
Hepatobiliary Surgery, Second Clinical College, Chongqing
University of Medical Science (Yuzhong, China). The surgically
resected tissues were fixed in 10% formalin, embedded in paraffin,
cut into 5-mm sections and stained with hematoxylin and eosin.
Histopathological diagnosis and classification were performed by
the same pathologist. All specimens were handled and made anonymous
according to relevant ethical and legal standards. The experimental
protocols for this study were approved by the Human Research
Ethical Committee and the Animal Research Ethics Committee of
Chongqing Medical University, Chongqing, China. All patients
provided written informed consent prior to obtaining the
samples.
Experimental animals and cell line
Healthy adult male Kunming mice (20–25 g, n=48),
which were purchased from the Center of Experimental Animals,
Chongqing Medical University, were used in this study. The mice
were kept in individual ventilated cages and were allowed access to
food and water ad libitum. The mice were used accordingly
for the different experiments (for the isolation of the KCs and
hepatic cells, for the transplantatoin of H22 cells, and for the
harvesting of ordinary serum and immune serum; 12 mice were used
for each of these 4 procedures). All animals received humane care
in accordance with the National Institutes of Health Guidelines and
the legal requirements in China. The mouse ascitic H22 tumor cell
line (HCC line) was kindly provided by Dr Ma Fang, the Institute of
Ultrasonic Medical Engineering, Chongqing Medical University.
Reagents and antibodies
Collagenase IV, Percoll, special grade fetal calf
serum, gadolinium chloride (GaCl3) and DMEM were all purchased from
Sigma (St. Louis, MO, USA). Mouse anti-human monoclonal
antibody-CD68 (sc-20060), rabbit anti-mouse CD16a polyclonal
antibody (sc-20627) (both from Santa Cruz Biotechnology, Santa
Cruz, CA, USA), biotinylated anti-mouse TNF-α (BS1857) and mouse
anti-IgG Fc immune serum (BS9203P) (both from Bioworld, St. Louis
Park, MN, USA).
Immunohistochemical analysis
The SP method was used, and the primary antibodies
were mouse monoclonal anti-human CD68 (12) and CD16a (Santa Cruz Biotechnology,
dilution 1:200). The operation procedure was undertaken according
to the instructions of the SP kit (Santa Cruz Biotechnology). DAB
was used for coloration. The brown granules in the cytoplasm were
taken as CD68- and CD16a-positive signals. Negative mouse control
serum and PBS were used to replace the primary antibody as negative
control and blank control, respectively.
Identification and counting of positive
cells
Among the cells which were positive for the
anti-CD68 antibody, cells in the cancerous tissue or the sinusoids
of non-cancerous tissues with a stellate or spindle shape were
taken as KCs, as previously discussed (12). The cells which were positive for
the anti-CD16a antibody were brown, and the shape was either
stellate or spindle. Image Tool 3.0 software was employed to count
cells at ×200 magnification (CKX 41SF; Olympus, Tokyo, Japan) in
one image randomly selected for each specimen. The expression level
of CD16a was determined as the average optical density, which was
measured by the Beihang CM-2000B Biological Image Analysis System
(Beihang University, Beijing, China).
Preparation of KCs
KCs were isolated from mouse livers (6 mice were
sacrificed for this procedure) by collagenase digestion and
differential centrifugation using Percoll as previously described
by Liu et al (13).
Briefly, the livers were excised after perfusion via the portal
vein with Ca2+ and Mg2+-free Hanks' balanced
salt solution containing 0.05% collagenase IV (Sigma) at 37°C and
cut into small pieces in collagenase buffer. The suspension was
filtered through nylon gauze, and the filtrate was centrifuged at
500 × g for 10 min at 4°C. Cell pellets were resuspended in buffer,
parenchymal cells were removed by centrifugation at 50 × g for 3
min, and the non-parenchymal cells were centrifuged on a 70:30%
Percoll gradient (Sigma) at 800 × g at 4°C for 20 min. KCs
concentrated at the interface of the 30 and 70% were collected and
cultured at a density of 1×106 in 24-well culture plates
containing DMEM supplemented with 10% FBS and antibiotics (100 U/ml
of penicillin G and 100 mg/ml of streptomycin sulphate) at 37°C in
the presence of 5% CO2. Nonadherent cells were removed
after 1 h by replacing the buffer. All adherent cells phagocytized
latex beads, indicating that they were KCs. The viability of KCs,
as determined by trypan blue exclusion, was >90%.
Preparation of hepatic cells and H22
cells
Another 6 mice were sacrificed to obtain the hepatic
cells. According to the above-mentioned method, using a 200 mesh
stainless steel screen, the hepatic histiocyte suspension was
centrifuged at 50 × g at 4°C for 2 min, the pellet was centrifuged
at 50 × g at 4°C for 5 min 3 times. The cells were cultured at a
density of 1×106. After 60 min, the cells were washed
with PBS and centrifuged at 50 × g at 4°C, and the pellets
containing the liver cells were cultured for use.
The H22 cells were then preserved in vivo in
mouse ascites. The mice transplanted with H22 cells (0.2 ml,
5×106; 12 mice were used) for 2 weeks were sacrificed.
The ascites were obtained and diluted 10 times with PBS and then
centrifuged at 300 × g for 5 min 2–3 times. The supernatant was
discarded, and the pellet was then filtrated using a 200 mesh
stainless steel screen and cultured for use.
Preparation of mouse serum
Blood (2–3 ml) was obtained from mice transplanted
with H22 cells for 2 weeks after the mice had been decapitated and
blood collected and preserved in an incubator at 37°C for 1 h or in
a refrigerator at 4°C for 3–4 h. The serum was centrifuged at 1000
× g for 15 min, and then the supernatant was obtained using a 56°C
thermostatic water bath for 30 min, and 0.02% sodium azide was
applied as an antiseptic, as previously described (14). The sample was the immune serum
containing special IgG anti-H22 cell and was stored at 4°C for use.
Ordinary serum was obtained from normal mice (not transplanted with
H22 cells) by the same method mentioned above.
Co-culture of KCs and H22 cells
KCs, H22 cells and hepatic cells were mixed to
culture, the cells were plated and counted and the cell count was
1×106 for each type of cell. GaCl3 (15), mouse ordinary serum, mouse immune
serum, and anti-IgG Fc immune serum were employed as intervention
factors. KCs and hepatic cells were set as group A; KCs and H22
cells were group B; KCs, H22 cells and 0.1 ml mouse ordinary serum
were group C; KCs, H22 cells and 0.1 ml mouse immune serum were
group D. GaCl3 (0.175 mg/ml) was added to the culture
flask to deplete macrophages or inhibit KC functions, and thus the
above groups were re-designated as group A1, B1, C1 and D1. When
0.1 ml anti-IgG Fc immune serum was added, the groups were group
A2, B2, C2 and D2. At 0, 2, 4, 8, 16 and 32 h, the samples were
obtained. KCs and H22 cells were separated from the feature of
adherence and the KCs were eluted with 0.25% trypase and washed
with PBS. The culture medium was centrifuged at 300 × g for 5 min,
and 0.5 ml supernatant was collected. The pellet was H22 cells,
which were then washed with PBS and diluted for the counting of
living cells using the Trypan blue kit (Beyotime, Haimen,
China).
Enzyme-linked immunosorbent assay (ELISA)
for TNF-α
ELISA was used to determine the levels of TNF-α. All
reagents, samples, and standards were prepared according to the
manufacturer's instructions (ELISA kit, Santa Cruz Biotechnology).
A 50 µl sample dilution was used. The reaction was performed
at 37°C for 120 min, plates were incubated with biotinylated
anti-mouse TNF-α (0.1 ml/well) at 37°C for 60 min. ABC solution
(0.1 ml) was added into each well, and the plate was incubated at
37°C for 30 min. TMB solution (0.1 ml) was further added into each
well, and the plate was incubated in the dark at 37°C for 20–25
min. TMB stop solution (0.1 ml) was added to each well. All ELISAs
were read on a microplate reader at 450 nm to determine the TNF-α
concentration.
RT-qPCR for TNF-α and CD16a mRNA
Total RNA was extracted from KCs using an SV Total
RNA isolation kit (Promega, Madison, WI, USA). Agarose gel
electrophoresis was employed to detect the three bands at 28S, 18S
and 5S for total RNA. RT-qPCR was employed to amplify the CD16a
mRNA and TNF-α mRNA from the mouse KCs. Specific primers
synthesized by Beijing SBS Genetech Co., Ltd. (Beijing, China) were
as follows: CD16a, 5′-AAG GCT GTG GTG AAA CTG GAC-3′ (sense) and
5′-GAG ATG GAG GAT GTA GTT GCT G-3′ (antisense); TNF-α, 5′-TCT ACT
GAA CTT CGG GGT GA-3′ (sense) and 5′-AGT AGA CCT GCC CGG ACT C-3′
(antisense). The products were 500 and 542 bp, respectively. The
primers for β-actin were 5′-CTT CTT TGC AGC TCC TTC GT-3′ (sense)
and 5′-TTC TCC ATG TCG TCC CAG T-3′ (anti-sense), and the product
was 300 bp. RT-qPCR was performed in a 40 µl volume
consisting of 1 µM of each primer, 2 µl of 0.5 mM
each dNTP, 16 µl RNase-free deionized water, 4 µl 10X
RNA PCR buffer, 8 µl 5 mM MgCl2, 1 µl Tfl
DNA polymerase, 2 µl 0.25 U/µl AMV reverse
transcriptase, 1 µl RNA enzyme inhibitor, and 2 µl
DNA template. The sample was heated to 94°C for 2 min in the first
chain of cDNA, followed by amplification for 40 cycles at 94°C for
30 sec, 55°C for 45 sec, and 72°C for 60 sec, and 1 cycle at 72°C
for 5 min, and 4°C for 8 min in the second chain. Subsequently, 5
µl of each product was analyzed by 1% agarose gel
(containing 0.5 µg/ml EB) electrophoresis. The relative
quantity of mRNA was described as the ratio of the CD16a or TNF-α
mRNA to β-actin mRNA.
Western blot analysis
The expression of CD16a protein in the KCs was
analyzed by western blot analysis. Protein extracts were obtained
according to the manufacturer's instructions. Protein concentration
was determined using the Bradford protein assay kit (Bio-Rad,
Hercules, CA, USA). Extracted protein was separated by 10% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and transferred
to polyvinylidene fluoride membranes (Dupont, Wilmington, DE, USA).
The membranes were washed with 0.1% Tween-20-phosphate-buffered
saline and incubated with 5% dry non-fat skimmed milk powder in
0.1% Tween-20-phosphate-buffered saline, at pH 7.4, for 1 h, and
subsequently with rabbit anti-mouse CD16a polyclonal antibody
(dilution, 1:200 and goat anti-rabbit IgG for 1 h. Finally, the
immune complexes were developed with an enhanced chemiluminescence
detection kit (Pierce, Waltham, MA, USA). β-actin was used as the
control. The GelGDoc2000 imaging system (Bio-Rad, Munich, Germany)
was employed to analyze the bands, and the protein level was
represented by the relative optical density (ROD) × areas
(mm2).
Statistical analysis
Data were analyzed using SPSS 10.0 software and are
expressed as the means ± SD. Statistical analysis was carried out
using the t-test and bivariant analysis. A P-value <0.05 was
considered to indicate a statistically significant difference.
Results
Distribution of KCs in human HCC
samples
CD68 is the specific marker of the macrophage, and
is a valid symbol of KCs (9). In
the present study, KCs were present in all 87 samples of HCC
tissues; these cells were puffy and dispersed in the liver
sinusoids, and the shapes were orbicular-ovate, astro or fusiform.
In the 39 normal liver tissue samples, the KCs were well
distributed. In cancerous tissues, especially in the poorly
differentiated tissue, the number of KCs was at a trace level
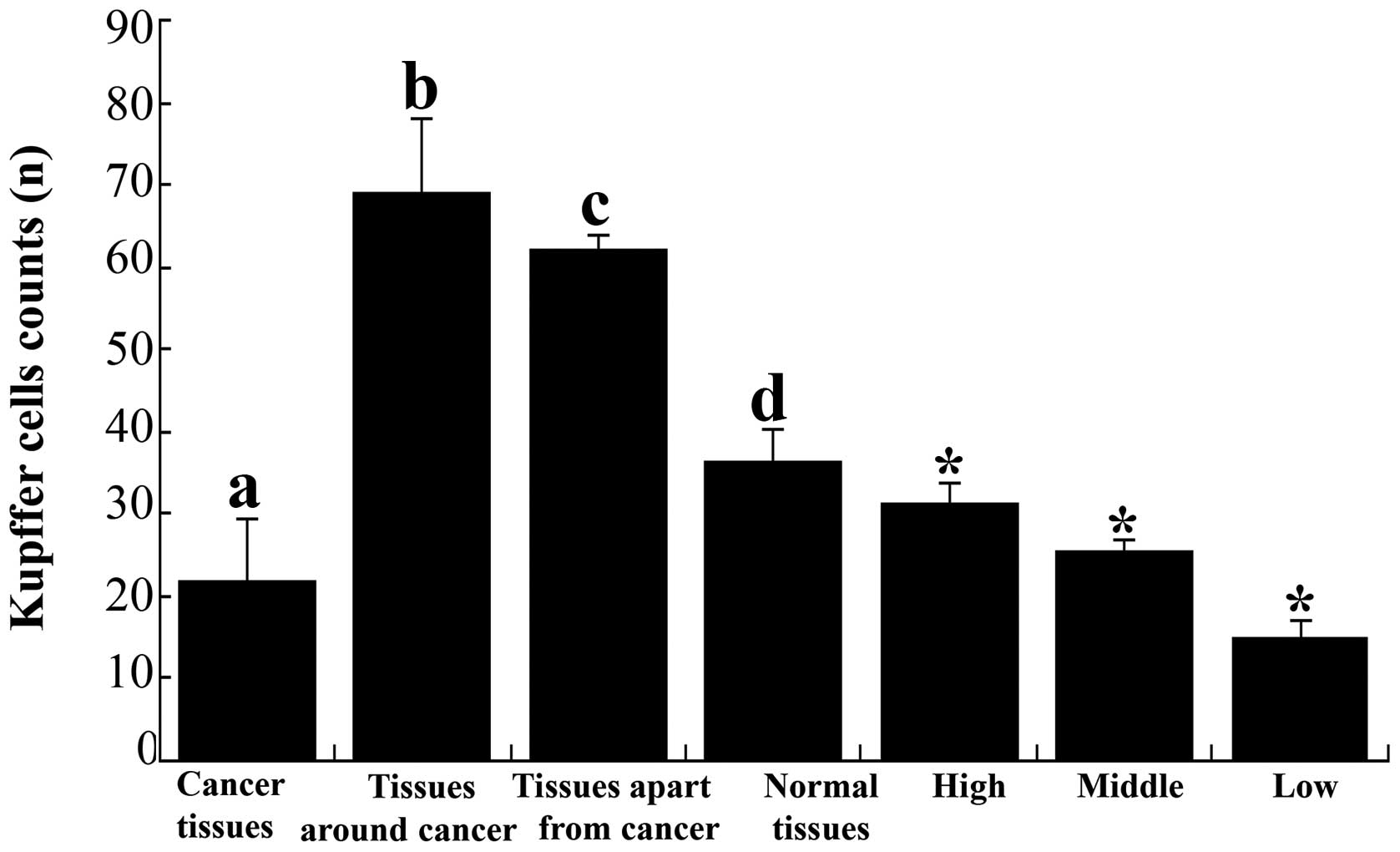
(Fig. 1). The quantities of KCs
were 21.6±7.8, 68.8±9.1, 62.0±1.9 and 36.2±4.1 in the HCC tissue
samples, tissues around the cancer, tissues apart from the cancer
and normal human hepatic tissue samples, respectively (Fig. 2). In cancerous tissues, the KCs
were fewer than in tissue around the cancer, apart from the cancer
and normal hepatic tissues (P<0.01). The level of KCs in tissues
around and apart from the cancer was higher than in normal hepatic
tissue samples (P<0.01), but there was no significant difference
between these two types of tissue (P>0.05) (Fig. 2). The quantities of KC in highly,
moderately and poorly differentiated tissue were 31.3±2.5, 25.3±1.5
and 14.8±2.2, respectively, and the difference between them was
significant (P<0.05) (Fig.
2).
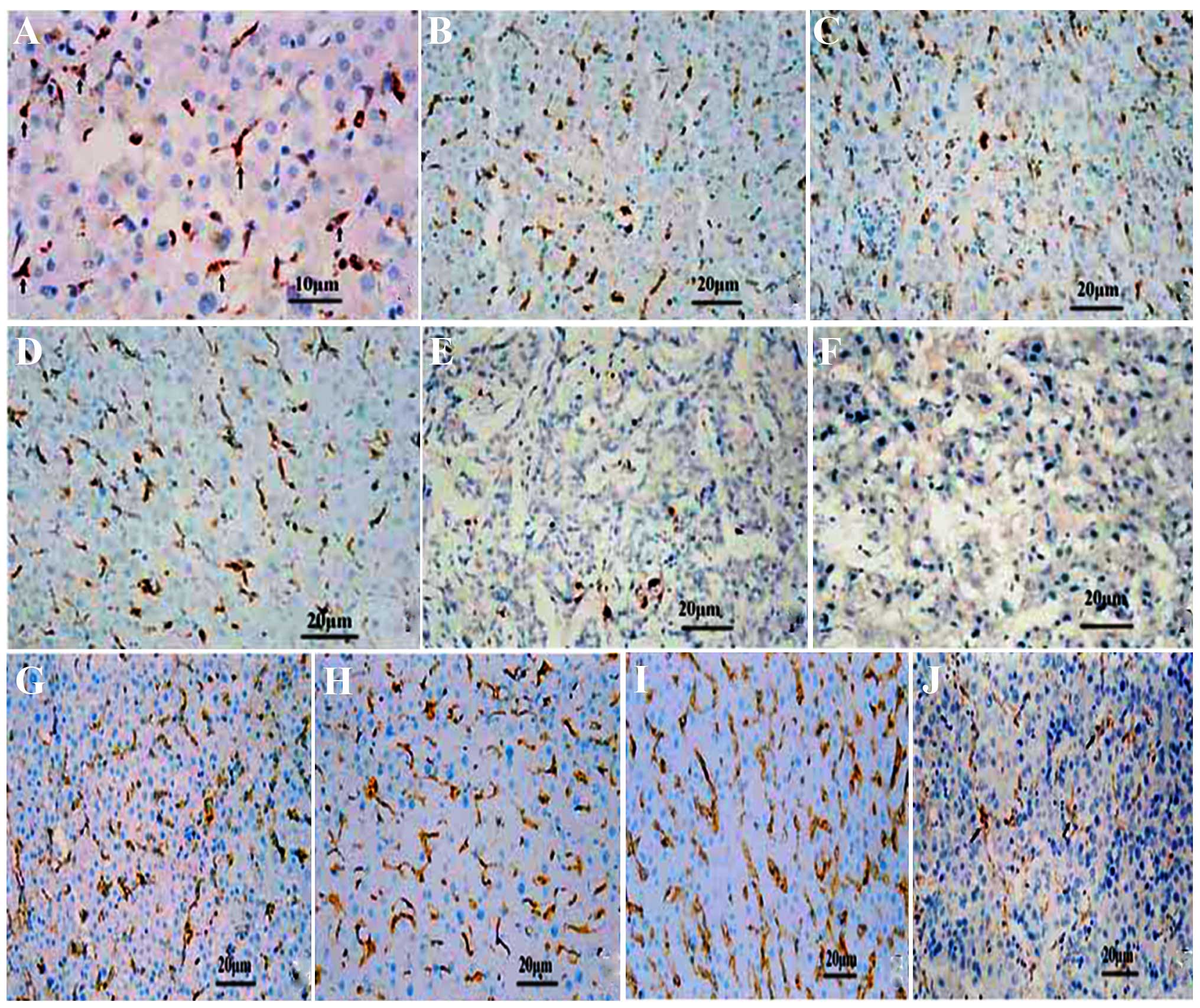 | Figure 1The distribution and expression of
Kupffer cells (KCs) and CD16a in various differentiated cancer
tissues studied by immunohistochemical analysis (SP method). (A)
The KCs were puffy, of orbicular-ovate and of astro or fusiform
morphology in the sinusoid at ×400 magnification. (B) KCs in human
hepatic tissue around the cancer, (C) normal human hepatic tissue,
(D) human hepatic tissue apart from the cancer, (E) human liver
cancer, and (F) poorly differentiated human liver cancer,
respectively. CD16a in the human liver detected by
immunohistochemical analysis in (G) the normal liver. (H) Hepatic
tissue around cancer. (I) Hepatic tissue apart from cancer. (J) In
liver cancer tissues, the CD16a-positive cells were puffy, and
exhibited orbicular-ovate, astro or fusiform morphology in the
sinusoid. Original magnification, ×200. |
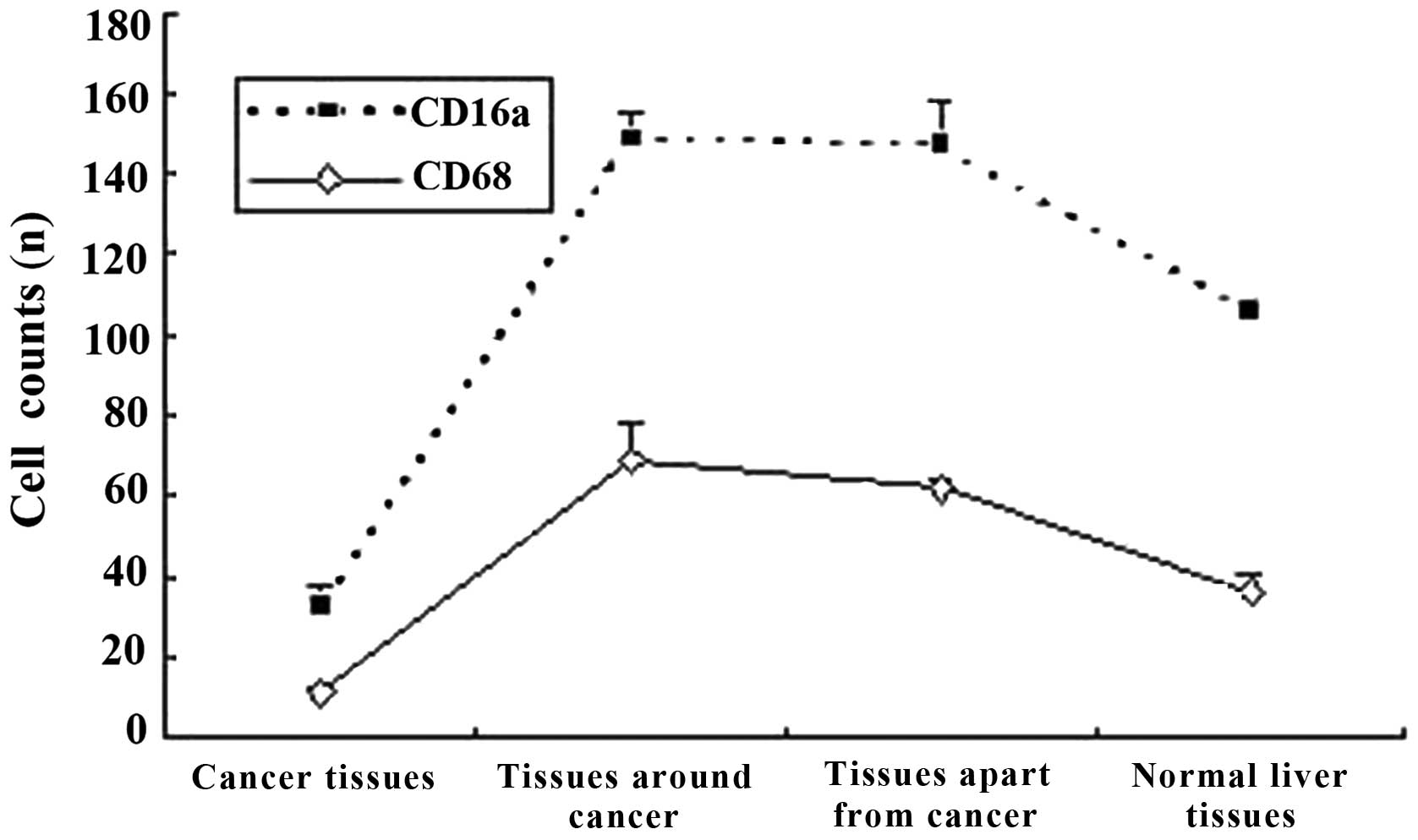
Expression of KCs CD16a in human HCC
The CD16a-positive cells were puffy and
well-distributed. In the tissues around and apart from the cancer,
the number (Table I) and the
distribution of CD16a-positive cells rose (Fig. 3). In the HCC tissue samples,
positive cells were also observed, but the number decreased and
distribution was correlated to the degree of differentiation
(Table I). The CD16a-positive
cells were present in the sinusoids, with a shape similar to the
KCs (Fig. 1).
 | Table INumber of CD16a-positive cells and
the average optical density. |
Table I
Number of CD16a-positive cells and
the average optical density.
| Cancer
tissues
(n=87) | Tissues around
cancer
(n=87) | Tissues apart from
cancer
(n=87) | Normal liver
tissues
(n=39) | Degree of
differentiation
|
|---|
High
(n=27) | Moderate
(n=41) | Poor
(n=19) |
|---|
| Positive cell count
(means ± SD) | 21.6±5.4 | 79.7±6.9 | 85.0±10.7 | 47.9±7.6 |
26.3±3.6c |
19.3±3.0d |
14.5±1.3e |
| ta | – | 21.724 | 10.693 | 6.342 |
4.427c,d |
3.307c,d | – |
| Pa | – | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | – |
| tb | – | 3.681 | 3.544 | – | – | – | – |
| Pb | – | <0.05 | <0.05 | – | – | – | – |
| Average
optical | | | | | | | |
| density | 0.78±0.1 | 0.44±0.07 | 0.35±0.02 | 0.24±0.03 |
0.59±0.05f |
0.7±0.04g |
0.86±0.03h |
| ta | – | 8.0456 | 12.38 | 15.588 |
4.282f,g |
6.025g,h | – |
| Pa | – | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | – |
| tb | – | – | 7.973 | – | – | – | – |
| Pb | – | – | <0.01 | – | – | – | – |
The quantities of the CD16a-positive cells were
21.6±5.4, 79.7±6.9, 85.0±10.7 and 47.9±7.6 in the HCC tissue
samples, tissues around the cancer, tissue apart from the cancer
and normal human hepatic tissue samples, respectively. The
quantities in the tissue samples around and apart from the cancer
increased markedly compared with the cancerous tissue (P<0.01)
and normal hepatic tissue samples (P<0.05). The average optical
density of the cancerous tissue increased markedly compared with
the tissue around the cancer, apart from the cancer and the normal
hepatic tissue (P<0.01) (Table
I). The number of CD16a-positive cells and the average optical
density were significantly different in the highly, moderately and
poorly differentiated cancerous tissue (P<0.05) (Table I). The distributive tendency of
the CD16a-positive cells was coincident with the KCs from the
associativity analysis (correlation coefficient was 0.984, P=0.016)
(Fig. 3).
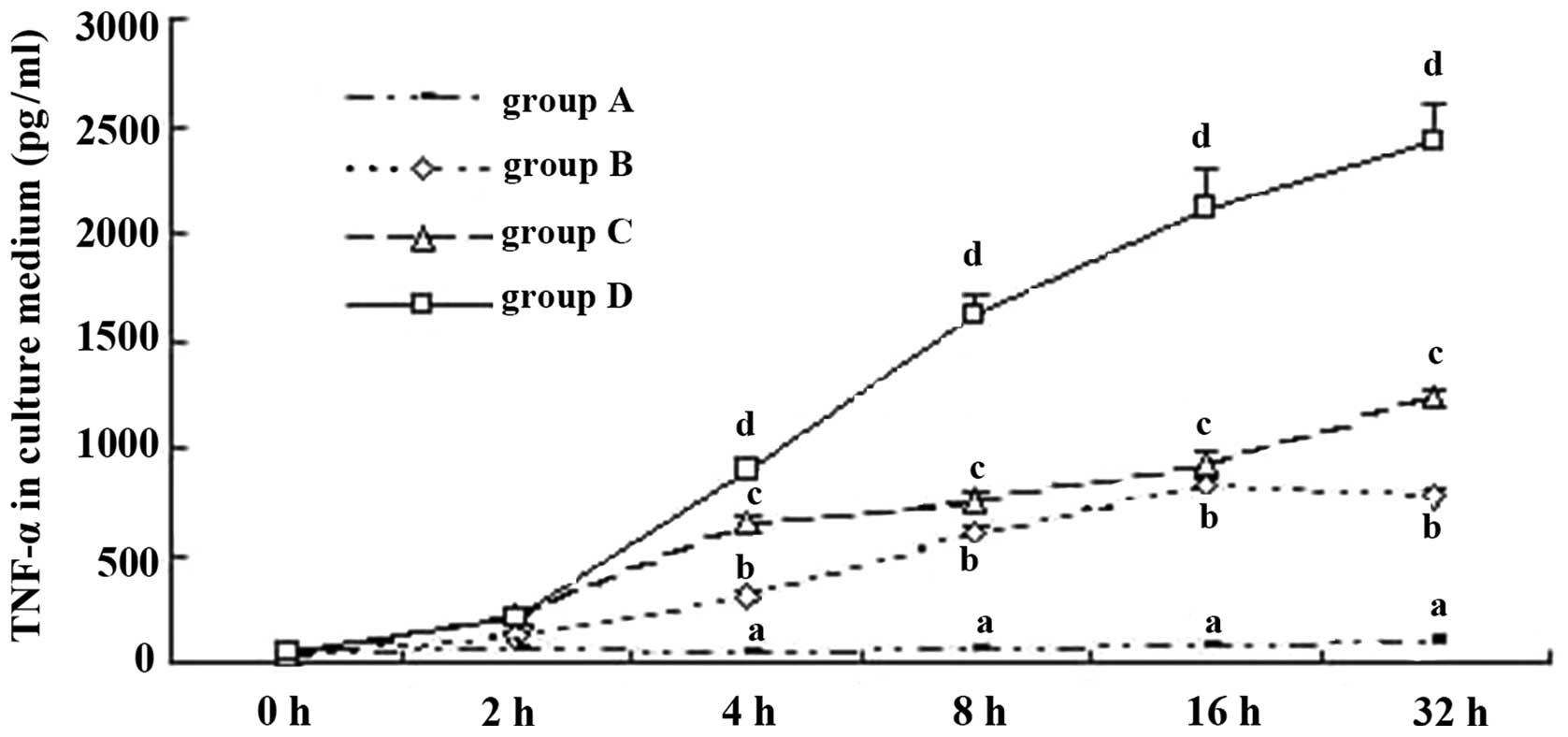
TNF-α expression in KCs from mice
TNF-α is one of the most important indicators for
the activation of KCs (16). In
the present study, ELISA and RT-qPCR were employed to determine the
TNF-α expression level in the culture supernatant and KCs, in order
to analyze the activation of mouse KCs under different culture
conditions. When the KCs were cultured with hepatic cells, the
TNF-α level was stable in the supernatant (group A) and increased
with the addition of H22 cells (group B), indicating that H22 cells
activated KCs. When ordinary serum (group C) and immune serum
(group D) were added to group B, the TNF-α level increased
markedly, especially in group D (Fig.
4). Hence, we suggest that H22 cells directly activate KCs, and
the ordinary and immune serum, which contained ordinary or special
IgG, exerted a promotive effect. This phenomenon was noticeable
especially when special IgG was added. In relation to the TNF-α
level, in groups A–C the difference between cell groups was
insignificant (P>0.05). Serum containing IgG activated KCs, the
linkage of CD16a in KCs, and Fc in IgG was probably the key. Hence,
anti-IgG Fc serum was employed to test this possibility in the
present study. As expected, anti-IgG Fc effectively served as an
obstacle to the functioning of the immune serum (Figs. 5 and 6).
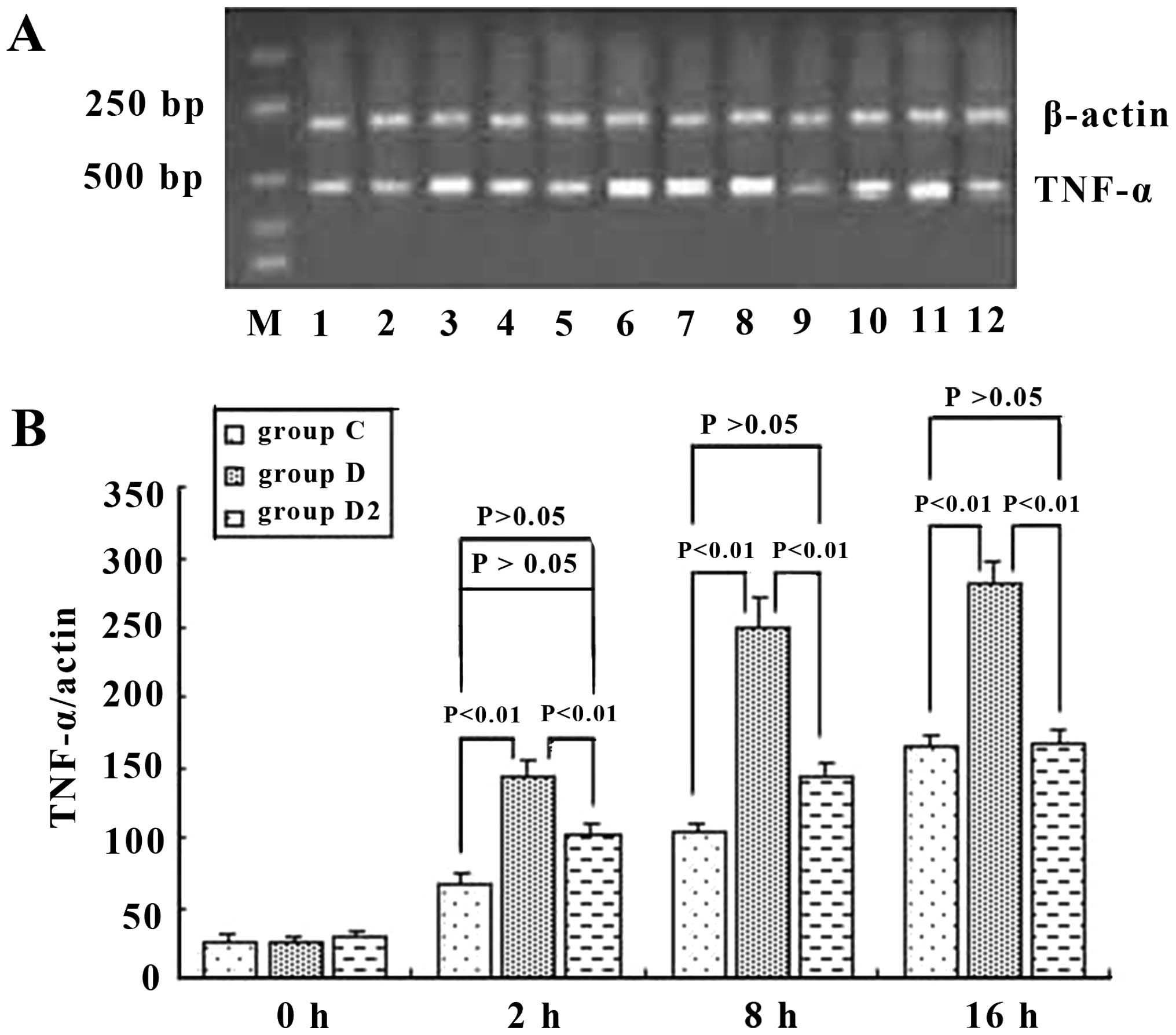 | Figure 6Tumor necrosis factor (TNF)-α mRNA of
mouse Kupffer cells (KCs) (A) in group C, D, and D2, and
semi-quantitative analysis (B) at different time points. It was
shown that ordinary serum and immune serum increased the expression
level of TNF-α mRNA, but the immune serum increased it more: 2.45-,
2.39- and 1.72-fold greater than that of the ordinary serum at 2, 8
and 16 h, respectively. Anti-IgG Fc blocked this effect of immune
serum and reduced the mRNA expression level of TNF-α in KCs by
26.4, 42.2 and 44.1% at 2,8 and 16 h, respectively. P-values
represent the difference between two indicated groups, and groups
C,D and D2 are as described in the Materials and
methods. |
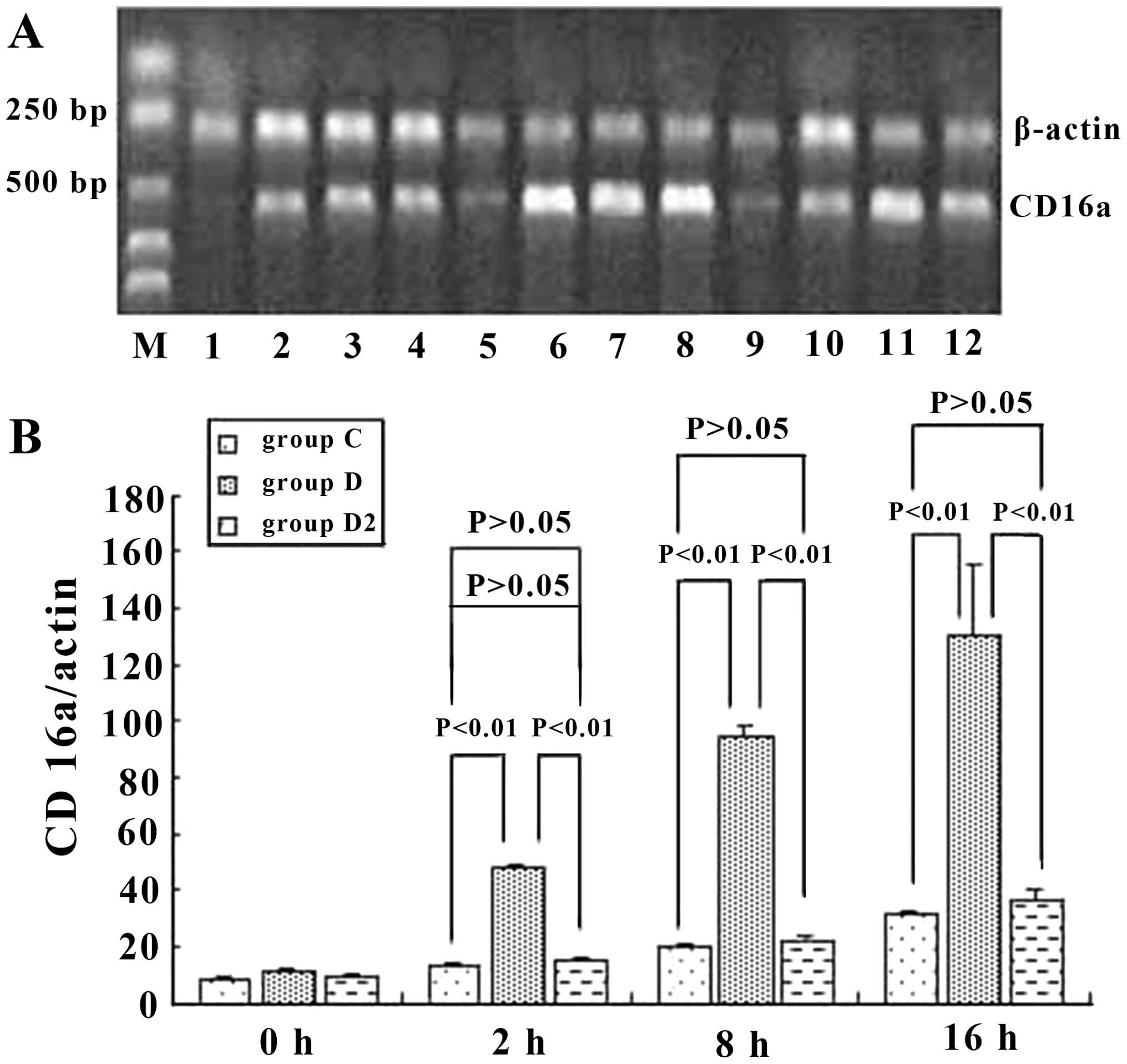
CD16a expression in KCs
To assess the role of CD16a in KCs, we examined
whether the activation of KCs was accompanied by an increase in the
mRNA and protein expression of CD16a and whether this increase
would be prevented by inhibition with IgG Fc. The results
demonstrated that the mRNA level of CD16a in KCs was increased in
ordinary serum and immune serum groups. However, the
transcriptional level of CD16a in the immune serum group increased
in a more marked manner and was 3.9-, 4.9- and 3.9-fold greatern
than that of the ordinary serum group at 2, 8 and 16 h,
respectively. Anti-IgG Fc blocked this effect of the immune serum
and decreased the expression of CD16a in KCs by 59.6, 73.5 and
71.9% at 2, 8 and 16 h, respectively, which was at the same level
as the ordinary serum (Fig. 7).
Western blot analysis was employed to detect the production of
CD16a. When immune serum was added, CD16a protein expression was
increased significantly. However, the expression of CD16a protein
was decreased by 77.2, 76.4 and 82.1% at 2, 8 and 16 h,
respectively, when anti-IgG Fc was applied (P<0.05) (Fig. 8). Hence, at 2, 8 and 16 h, the
expression of CD16a and TNF-α in KCs rose and the trend was
consistent (P<0.05) when ordinary and immune serum were applied
(Fig. 6 and 7), but when anti-IgG Fc was added, the
expression of CD16a and TNF-α decreased (P>0.05).
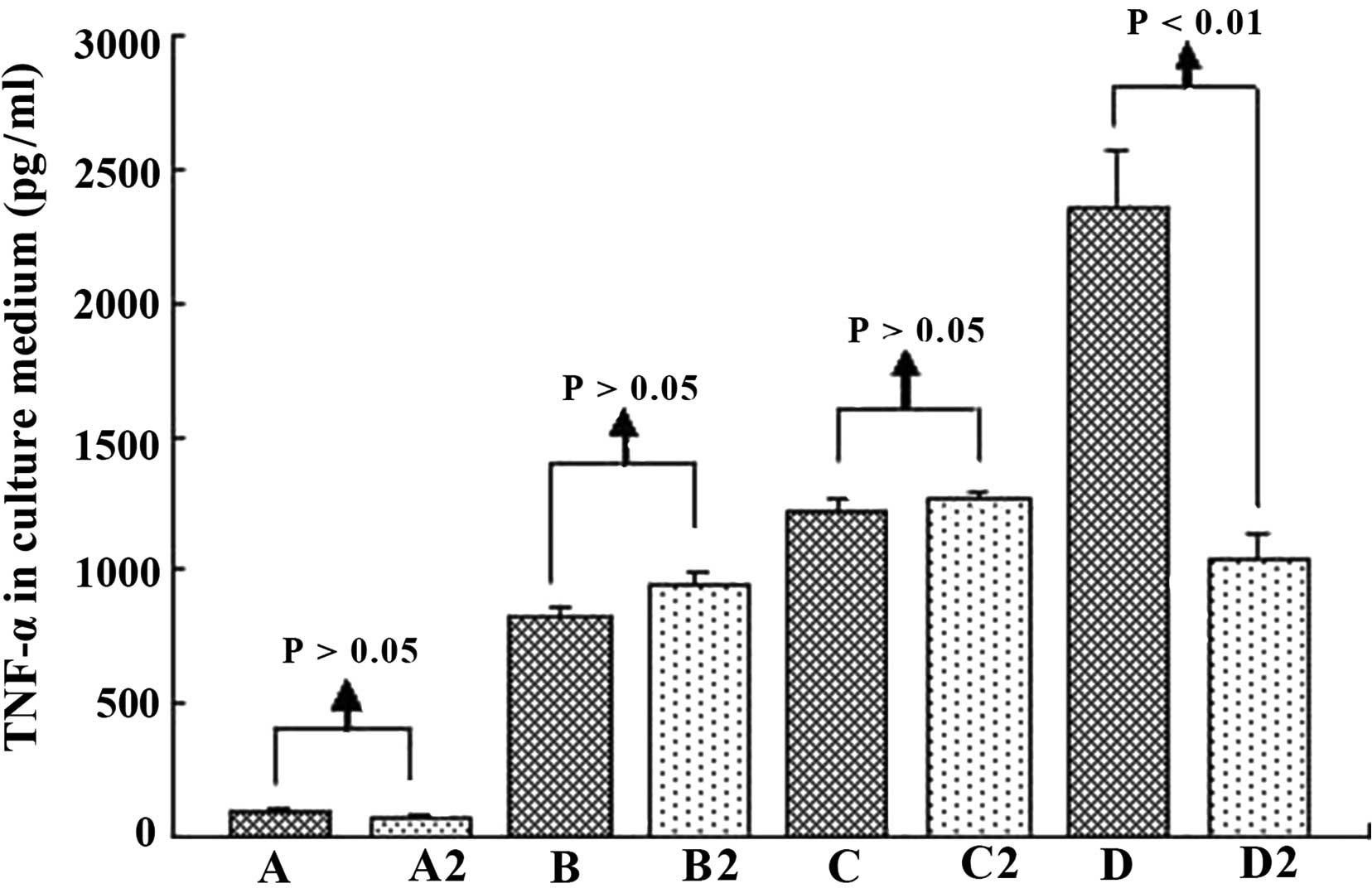
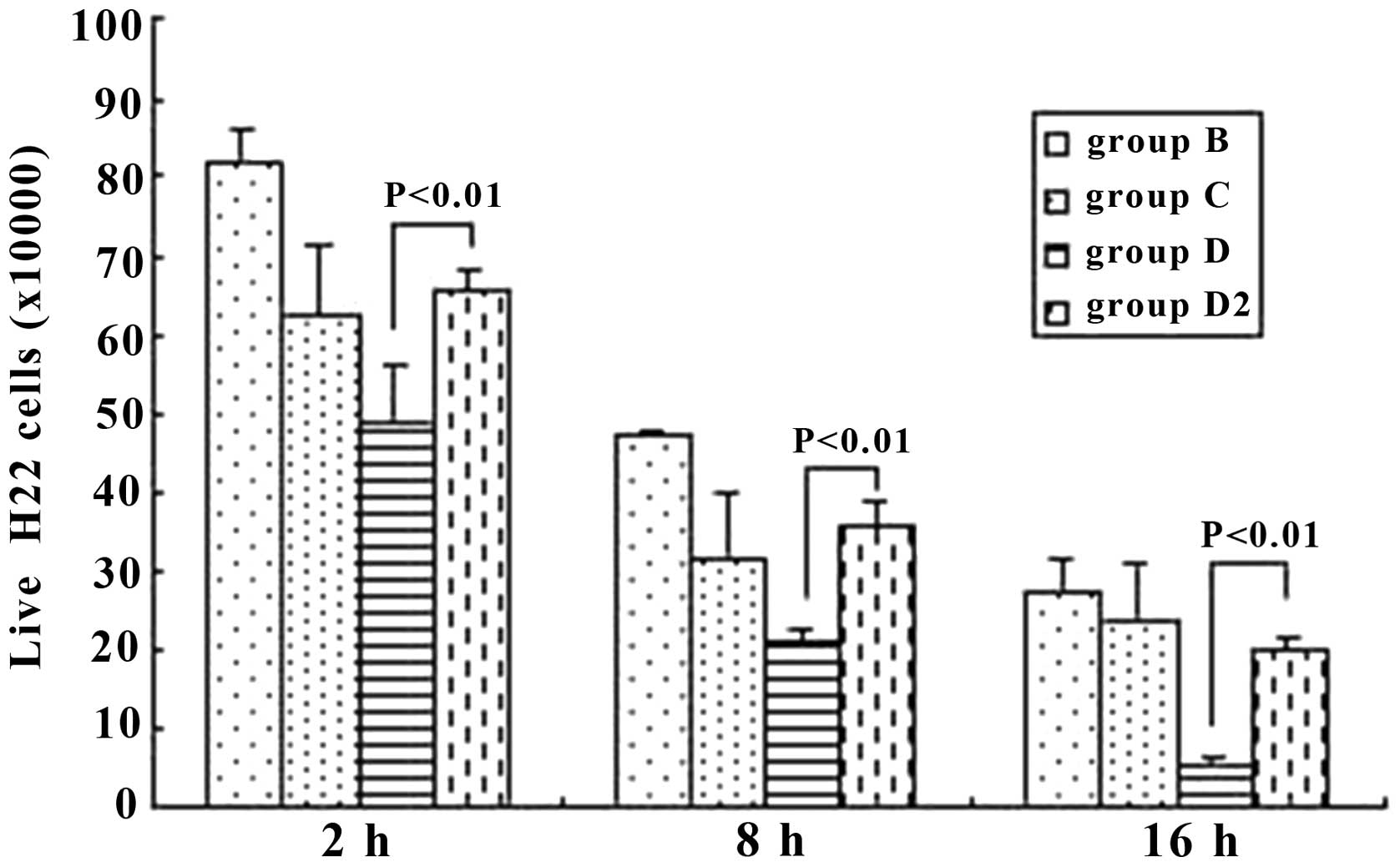
Living H22 cells
The living H22 cells were Trypan-blue resistant.
From 2 h to 16 h, we demonstrated that the number of living H22
cells decreased in every group (P<0.05). However, it should be
noted that at the same time points (2, 8 or 16 h), when immune
serum (group D) was applied and the anti-IgG Fc was also added
(group D2), the difference was significant between these two groups
(P<0.01), with group D2 markedly increasing (Fig. 9).
Discussion
KCs are important defensive cells in the liver; the
number and function of KCs changes markedly when the liver is
diseased (17). CD68 is a
specific surface marker of monocytes and macrophages, including
KCs, and thus it can be used as a marker for KCs in the liver of
mice. In this study, the results indicated that the number of KCs
in cancerous tissue was markedly lower than that in cancer-adjacent
tissue and non-cancerous hepatic tissue samples, and numbers
decreased with decreasing degrees of histological grade
differentiation. These results are in agreement with previous
findings (18). It is likely that
the irregular arrangement of liver cancer cells led to the
disorganization of the hepatic lobule, and the normal sinus
hepaticus on which the KCs were dependent then disappeared
(19). This also explains why the
quantity of KCs in poorly differentiated cancer tissue samples was
less. If the cancer tissue differentiated more poorly, the
heteromorphosis of the tissue was more obvious and the normal sinus
hepaticus was ruined more. However, other factors likely influenced
the distribution of KCs, such as immunoreactivity, the migration of
KCs from outside into the liver, and splitting and activation of
KCs.
Macrophages that infiltrate the tumor stroma are
often referred to as tumor-associated macrophages (TAMs).
Experimental models using mice have shown that TAMs facilitate
tumor progression by promoting inflammation, stimulating
angiogenesis and suppressing antitumor immunity (20,21). However, macrophage-activating
agents suppress tumor growth and diminish tumor metastasis
(22). AngII acts upstream of a
potent macrophage amplification program and it has been shown that
tumors remotely exploit the hormone pathway to stimulate
cancer-promoting immunity (23).
In the present study, the number of KCs in the para-cancerous
hepatic tissue was markedly increased, indicating that the KCs were
important for defending against hepatoma carcinoma cells.
ADCC has been recognized as one of several molecular
mechanisms by which monoclonal antibodies of the IgG isotype class
mediate anti-tumor effects (24,25). ADCC reactions are dependent on the
simultaneous binding of IgG1 molecules to tumor cells and to Fc
receptors such as CD16a expressed on a variety of immuno-effector
cells, such as NK and monocytic cells (26,27). This indicates that the strength of
antibody interaction with CD16a correlates with ADCC potencies.
As the bridge between macrophages and IgG Fc, CD16a
is the key molecule of ADCC. If a promoting factor is applied
between tumor cells and CD16a, further activation of macrophages
and the promotion of tumor cell phagocytosis are expected to occur
(7). In this study, it was shown
that KCs were activated in the liver following cancer, the number
of CD16a positive cells was greater than in the normal liver tissue
samples, and decreased with the decrease in the degree of
differentiation. CD16a-positive cells and KCs were distributed in
the hepatic sinusoid and mainly had a similar shape. The expression
of CD16a mRNA and protein in KCs was confirmed in this study. These
results suggest the existence of CD16a in KCs and that the
activation of KCs, which mainly resulted in CD16a expression,
mediated ADCC reactions to secrete TNF-α.
The average optical density of CD16a in KCs in
cancer tissue was more than in other hepatic tissue samples,
implying that CD16a was important in the ADCC of KCs to hepatoma
cells. Activated KCs kill target cells directly by swallowing and
releasing lyosomal enzyme, NO and peroxydase. They also cooperate
to resist tumor cells by secreting cytokines including TNF-α, IL-1,
IL-6 and granulocyte-macrophage colony-stimulating factor (GM-CSF)
(5). In our present study, mouse
KCs were cultured with H22 tumor cells. The TNF-α level was
increased in the groups treated with the ordinary and the immune
serum, and we also noted that the number of H22 tumor cells
decreased. After the KCs were treated with GaCl3, the TNF-α level
and the number of H22 cell did not exhibit marked changes. These
indicated that KCs likely killed tumor cells directly by
phagocytosis, in a complementary way, and cytokines.
KCs are resident macrophages and play an important
role in the defense against invading particles via phagocytosis
(28). KCs can be activated by
foreign stimuli, such as SiO2 nanoparticles, and release a variety
of bioactive mediators, such as reactive oxygen species, TNF-α and
NO (29). These findings indicate
that certain other factors play an important role in the activation
of KCs. In the present study, KCs and H22 cells were co-cultured.
Subsequently, we noted that the number of H22 cells was reduced,
demonstrating that tumor cells that contain tumor antigen were one
of the activating factors of KCs.
The antibody effect is an important way of humoral
immunity, while ADCC and the activation of the complement system
are the main pathways (30). The
ordinary mouse serum was added to the culture medium of KCs and H22
cells, and the results at 2, 8 and 16 h indicated that ordinary
serum did not effectively inhibit the growth of tumor cells. On the
contrary, however, Ichiyama et al found that immunoglobulin
injected into the veins downregulated the immune function by
inhibiting the activation of NF-κB, which was induced by TNF-α in
macrophages in a dose-dependent manner, and also by blocking the
surface receptor CD16a on mononuclear macrophages (31). However, we noted that the mouse
serum containing tumor specific IgG boosted the clearance of H22
cells markedly, showing that the tumor-specific antibody exerted a
marked effect on the immune function of the KCs. When the anti-IgG
Fc immune serum was added, the expression of CD16a mRNA and protein
in KCs, as well as the number of living H22 cells, decreased
notably, demonstrating that the suppression of tumor cells by KCs
was inhibited when the Fc fragments of IgG were blocked by the
anti-IgG Fc immune serum. However, the expression of CD16a and the
function of the KCs were not completely consistent, and the
ordinary serum made the KCs secrete TNF-α also. These results
indicated that certain other important pathways contributed to the
inhibition of tumor cells, and the blockade of the Fc fragment of
IgG likely caused other changes (31) and led to the functional diversity
of the KCs. However, the antibodies in the anti-H22 cells immune
serum were not specifically identified, which is a shortcoming of
this study.
Several variants which enhanced CD16a binding
affinity have been identified and have been demonstrated to enhance
ADCC activity and increase phagocytosis (32). Furthermore, the gene polymorphism
of CD16a is relevant to the occurrence and prognosis of many
diseases, including tumor-related diseases (33,34). A previous work has shown that both
glyco- and protein-engineering lead to a higher binding affinity to
CD16a. From a mechanistic point of view, these data suggest that
this gain in affinity is achieved by different mechanisms (35), and it has also been shown that
CD16a takes effect through several mechansisms (36). In the present study, we proved
that CD16a was expressed in KCs, and we also investigated a
profound action mechanism of ADCC in KCs for further. As a
low-affinity receptor of IgG, CD16a may be potentially used for the
development of novel bioimmunotherapy for liver cancer.
Acknowledgments
This study was supported by grants from the
Chongqing Health and Family Planning Commission (2015ZDXM012), the
Natural Science Foundation of Hubei Provincial Department of
Education (B2015468), the Natural Science Foundation of Hubei
Province of China (2015CFB615) and the National Natural Scientific
Foundation of China (no. 81272570).
References
|
1
|
Lafaro KJ, Demirjian AN and Pawlik TM:
Epidemiology of hepatocellular carcinoma. Surg Oncol Clin N Am.
24:1–17. 2015. View Article : Google Scholar
|
|
2
|
Liu ZM, Tseng HY, Tsai HW, Su FC and Huang
HS: Transforming growth factor β-interacting factor-induced
malignant progression of hepatocellular carcinoma cells depends on
superoxide production from Nox4. Free Radic Biol Med. 84:54–64.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kaneda Y: Therapeutic strategies for
controlling the metastasis and recurrence of cancers: contribution
of drug delivery technologies. Adv Drug Deliv Rev. 64:707–709.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Leong WS, Thomas KA, Chan CH and Stevenson
GT: A standardized conversion of IgG antibody to bispecific form
with inversely altered affinities for Fcγ-receptors II and III. Mol
Immunol. 48:760–768. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ehling J and Tacke F: Role of chemokine
pathways in hepatobiliary cancer. Cancer Lett. pii:
S0304-3835(15)00411-5. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Flaherty MM, MacLachlan TK, Troutt M,
Magee T, Tuaillon N, Johnson S, Stein KE, Bonvini E, Garman R and
Andrews L: Nonclinical evaluation of GMA161 - an antihuman CD16
(FcγRIII) monoclonal antibody for treatment of autoimmune disorders
in CD16 transgenic mice. Toxicol Sci. 125:299–309. 2012. View Article : Google Scholar
|
|
7
|
Herter S, Birk MC, Klein C, Gerdes C,
Umana P and Bacac M: Glycoengineering of therapeutic antibodies
enhances monocyte/macrophage-mediated phagocytosis and
cytotoxicity. J Immunol. 192:2252–2260. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kobayashi E, Motoi S, Sugiura M, Kajikawa
M, Kojima S, Kohroki J and Masuho Y: Antibody-dependent cellular
cytotoxicity and cytokine/chemokine secretion by KHYG-1 cells
stably expressing FcγRIIIA. Immunol Lett. 161:59–64. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kainer M, Antes B, Wiederkum S,
Wozniak-Knopp G, Bauer A, Rüker F and Woisetschläger M: Correlation
between CD16a binding and immuno effector functionality of an
antigen specific immunoglobulin Fc fragment (Fcab). Arch Biochem
Biophys. 526:154–158. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chaigne-Delalande B, Li FY, O'Connor GM,
Lukacs MJ, Jiang P, Zheng L, Shatzer A, Biancalana M, Pittaluga S,
Matthews HF, et al: Mg2+ regulates cytotoxic functions
of NK and CD8 T cells in chronic EBV infection through NKG2D.
Science. 341:186–191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou XP, Yang GS, Lu JH, Zhang HB, Li QG,
Han LZ and Zong M: Prospective study of liver parenchyma volume in
hepatectomy of primary liver cancer. Zhonghua Wai Ke Za Zhi.
43:1370–1374. 2005.In Chinese. PubMed/NCBI
|
|
12
|
Stănculeţ N, Grigoraş A, Avădanei R,
Floarea-Strat A, Amălinei C and Căruntu ID: Relationship between
Kuppfer cells, inflammation, and fibrosis in chronic hepatitis B
and C. Rev Med Chir Soc Med Nat Iasi. 117:880–889. 2013.
|
|
13
|
Liu ZJ, Yan LN, Li XH, Xu FL, Chen XF, You
HB and Gong JP: Up-regulation of IRAK-M is essential for endotoxin
tolerance induced by a low dose of lipopolysaccharide in Kupffer
cells. J Surg Res. 150:34–39. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qu YH, Li Y, Wu YF, Fang JP, Huang SL,
Huang Y and Wei J: Influence of FcγRIIIa polymorphism on
rituximab-dependent NK cell-mediated cytotoxicity to Raji cells.
Zhongguo Shi Yan Xue Ye Xue Za Zhi. 18:1269–1274. 2010.In Chinese.
PubMed/NCBI
|
|
15
|
Liu C, Yang Z, Wang L, Lu Y, Tang B, Miao
H, Xu Q and Chen X: Combination of sorafenib and gadolinium
chloride (GdCl3) attenuates dimethylnitrosamine (DMN)-induced liver
fibrosis in rats. BMC Gastroenterol. 15:1592015. View Article : Google Scholar
|
|
16
|
Yang P, Zhou W, Li C, Zhang M, Jiang Y,
Jiang R, Ba H, Li C, Wang J, Yin B, et al: Kupffer-cell-expressed
trans-membrane TNF-α is a major contributor to lipopolysaccharide
and D-galactosamine-induced liver injury. Cell Tissue Res.
363:371–383. 2016. View Article : Google Scholar
|
|
17
|
Xu FL, You HB, Li XH, Chen XF, Liu ZJ and
Gong JP: Glycine attenuates endotoxin-induced liver injury by
downregulating TLR4 signaling in Kupffer cells. Am J Surg.
196:139–148. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu K, He X, Lei XZ, Zhao LS, Tang H, Liu
L and Lei BJ: Pathomorphological study on location and distribution
of Kupffer cells in hepatocellular carcinoma. World J
Gastroenterol. 9:1946–1949. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tanaka M, Nakashima O, Wada Y, Kage M and
Kojiro M: Pathomorphological study of Kupffer cells in
hepatocellular carcinoma and hyperplastic nodular lesions in the
liver. Hepatology. 24:807–12. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qian BZ and Pollard JW: Macrophage
diversity enhances tumor progression and metastasis. Cell.
141:39–51. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fang W, Ye L, Shen L, Cai J, Huang F, Wei
Q, Fei X, Chen X, Guan H, Wang W, et al: Tumor-associated
macrophages promote the metastatic potential of thyroid papillary
cancer by releasing CXCL8. Carcinogenesis. 35:1780–1787. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Eljaszewicz A, Wiese M, Helmin-Basa A,
Jankowski M, Gackowska L, Kubiszewska I, Kaszewski W, Michalkiewicz
J and Zegarski W: Collaborating with the enemy: function of
macrophages in the development of neoplastic disease. Mediators
Inflamm. 2013:8313872013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cortez-Retamozo V, Etzrodt M, Newton A,
Ryan R, Pucci F, Sio SW, Kuswanto W, Rauch PJ, Chudnovskiy A,
Iwamoto Y, et al: Angiotensin II drives the production of
tumor-promoting macrophages. Immunity. 38:296–308. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Westphal S, Brinkmann H, Kalupa M, Wilke
A, Seitz-Merwald I and Penack O: Anti-tumor effects of anti-T-cell
globulin. Exp Hematol. 42:875–882. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu Z, Leng EC, Gunasekaran K, Pentony M,
Shen M, Howard M, Stoops J, Manchulenko K, Razinkov V, Liu H, et
al: A novel antibody engineering strategy for making monovalent
bispecific heterodimeric IgG antibodies by electrostatic steering
mechanism. J Biol Chem. 290:7535–7562. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Poggi A, Boero S, Musso A and Zocchi MR:
Selective role of mevalonate pathway in regulating perforin but not
FasL and TNFalpha release in human Natural Killer cells. PLoS One.
8:e629322013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nimmerjahn F and Ravetch JV: Fcgamma
receptors as regulators of immune responses. Nat Rev Immunol.
8:34–47. 2008. View
Article : Google Scholar
|
|
28
|
Tseng MT, Lu X, Duan X, Hardas SS, Sultana
R, Wu P, Unrine JM, Graham U, Butterfield DA, Grulke E and Yokel
RA: Alteration of hepatic structure and oxidative stress induced by
intravenous nanoceria. Toxicol Appl Pharmacol. 260:173–182. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen Q, Xue Y and Sun J: Kupffer
cell-mediated hepatic injury induced by silica nanoparticles in
vitro and in vivo. Int J Nanomedicine. 8:1129–1140. 2013.PubMed/NCBI
|
|
30
|
Cassard L, Cohen-Solal JF, Fournier EM,
Camilleri-Broët S, Spatz A, Chouaïb S, Badoual C, Varin A, Fisson
S, Duvillard P, et al: Selective expression of inhibitory Fcgamma
receptor by metastatic melanoma impairs tumor susceptibility to
IgG-dependent cellular response. Int J Cancer. 123:2832–2839. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ichiyama T, Ueno Y, Hasegawa M, Niimi A,
Matsubara T and Furukawa S: Intravenous immunoglobulin inhibits
NF-kappaB activation and affects Fcgamma receptor expression in
monocytes/macrophages. Naunyn Schmiedebergs Arch Pharmacol.
369:428–433. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Romain G, Senyukov V, Rey-Villamizar N,
Merouane A, Kelton W, Liadi I, Mahendra A, Charab W, Georgiou G,
Roysam B, et al: Antibody Fc engineering improves frequency and
promotes kinetic boosting of serial killing mediated by NK cells.
Blood. 124:3241–3249. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Frankenberger M, Ekici AB, Angstwurm MW,
Hoffmann H, Hofer TP, Heimbeck I, Meyer P, Lohse P, Wjst M,
Häus-singer K, et al: A defect of CD16-positive monocytes can occur
without disease. Immunobiology. 218:169–174. 2013. View Article : Google Scholar
|
|
34
|
Ramírez J, Fernández-Sueiro JL,
López-Mejías R, Montilla C, Arias M, Moll C, Alsina M, Sanmarti R,
Lozano F and Cañete JD: FCGR2A/CD32A and FCGR3A/CD16A variants and
EULAR response to tumor necrosis factor-α blockers in psoriatic
arthritis: a longitudinal study with 6 months of followup. J
Rheumatol. 39:1035–1041. 2012. View Article : Google Scholar
|
|
35
|
Liu Z, Gunasekaran K, Wang W, Razinkov V,
Sekirov L, Leng E, Sweet H, Foltz I, Howard M, Rousseau AM, et al:
Asymmetrical Fc engineering greatly enhances antibody-dependent
cellular cytotoxicity (ADCC) effector function and stability of the
modified antibodies. J Biol Chem. 289:3571–3590. 2014. View Article : Google Scholar :
|
|
36
|
Conry SJ, Meng Q, Hardy G, Yonkers NL,
Sugalski JM, Hirsch A, Davitkov P, Compan A, Falck-Ytter Y, Blanton
RE, et al: Genetically associated CD16(+)56(−) natural killer cell
interferon (IFN)-αR expression regulates signaling and is
implicated in IFN-α-induced hepatitis C virus decline. J Infect
Dis. 205:1131–1141. 2012. View Article : Google Scholar : PubMed/NCBI
|