Introduction
According to the Diabetes Atlas of the International
Diabetes Federation, 382 million individuals were affected by
diabetes worldwide in 2013 and its prevalence is expected to
increase to 592 million by the year 2035 (1). With the increasing prevalence of
diabetes in children and adolescents, male reproductive dysfunction
induced by diabetes has attracted worldwide attention (2–5).
Numerous studies have demonstrated that male
reproductive dysfunction induced by diabetes may be mediated
through hormonal alterations in the hypothalamic-pituitary-gonadal
axis or through direct effects on testes, sperm, epididymis and
Sertoli-blood testis barrier. Diabetes impairs spermatogenesis,
increases germ cell depletion, alters sperm parameters, induces
morphological alterations in the testes, alters glucose metabolism
in the Sertoli-blood testis barrier, reduces testosterone
production, leads to ejaculatory dysfunction and reduces libido
(6–15). However, the underlying mechanisms
of diabetes-related male reproductive dysfunction remain largely
unknown.
Increasing evidence has indicated that advanced
glycation end products (AGEs) play a causative role in the
progression of diabetes complications (16–18). AGEs are a heterogeneous class of
compounds formed by the non-enzymatic glycation of proteins, which
is accelerated in diabetes as a result of hyper-glycaemia and
oxidative stress. The receptor for AGEs (RAGE) exists in the
testes, epididymides and sperm (19,20). N′-carboxymethyl-lysine, a
prominent AGE, accumulates in the reproductive tract of patients
with diabetes (21), as well as
in animal models of both diabetes (22) and metabolic syndrome (23). Men suffering from diabetes have
poor sperm quality and functions, coinciding with the increase in
AGEs and RAGE (21,24,25). These studies show that AGEs play
important roles in male reproduction dysfunction induced by
diabetes.
To date, the mechanism underlying the low
testosterone levels associated with diabetes has not been fully
elucidated (26,27). As AGEs are important instigators
of diabetes complications, we hypothesized that these compounds
possibly play a contributory role in testosterone production by
affecting Leydig cells directly. Thus, the aim of this study, was
to determine whether AGEs exert inhibitory effects on testosterone
secretion by rat Leydig cells. In addition, the possible mechanisms
responsible for these effects were investigated.
Materials and methods
Animals
Male Sprague-Dawley rats (7–8 weeks old, weighing
250–300 g) were purchased from the Guangdong Medical Laboratory
Animal Center of China. The animals were kept in a
temperature-controlled room (20°C) with a 12 h-light/12 h-dark
photoperiod and were give access to food and water ad
libitum. All animal experiments were performed in accordance
with the guidelines of the Guangdong Ocean University Animal Use
Committee, and the protocols were approved by the Animal Ethics and
Welfare Committee of Guangdong Ocean University (Approval No.
2014031903).
Materials
Dulbecco's modified Eagle's medium/nutrient mixture
F-12 (DMEM/F12), fetal bovine serum (FBS), Hanks and
penicillin/streptomycin were purchased from Gibco Laboratories
(Grand Island, NY, USA). Bovine serum albumin (BSA),
tauroursodeoxycholic acid (TUDCA), N-acetyl-L-cysteine (NAC) and
collagenase were obtained from Sigma-Aldrich (St. Louis, MO, USA).
Percoll™ (Sterile) was obtianed from GE Healthcare (Pittsburgh, PA,
USA). The testosterone ELISA kit was purchased from Cayman Chemical
Co. (Ann Arbor, MI, USA). Anti-steroidogenic acute regulatory
protein (StAR; Cat. no. sc-25806), anti-cholesterol side-chain
cleavage enzyme (P450scc; Cat. no. sc-292456),
anti-3β-hydroxysteroid dehydrogenase (3β-HSD; Cat. no. sc-30820),
anti-glucose-regulated protein 78 (GRP78; Cat. no. sc-13968), and
anti-C/EBP homologous protein (CHOP; Cat. no. sc-166682) antibodies
were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA);
anti-actin (Cat. no. #4970) and anti-rabbit IgG (Cat. no. #7074)
were obtained from Cell Signaling Technology, Inc. (Danvers, MA,
USA). The endotoxin assay kit was purchased from Jinruisi Inc.
(Nanjing, China). The bicinchoninic acid (BCA) protein assay kit
was obtained from Shenenergy Inc. (Shanghai, China). TRIzol was
purchased from Invitrogen Life Technologies (Carlsbad, CA, USA).
M-MLV reverse transcriptase was obtained from Promega (Madison, WI,
USA). SYBR Premix Ex Taq™ was obtained from Takara Biot echnology
Co., Ltd. (Dalian, China). ECL chemiluminescent substrate was
obtained from Millipore (Billerica, MA, USA).
Preparation of AGEs
The AGEs were prepared as previously described
(28). Briefly, fatty acid-free
BSA was incubated with 50 mM D-glucose in phosphate-buffered saline
(PBS) solution in the dark and under ysterile conditions for 7
weeks at 37°C. Unincorporated glucose was removed by dialysis with
PBS. Control non-glycated BSA was incubated in the absence of
glucose under the same conditions. AGE-BSA solutions were tested
for endotoxin concentrations and confirmed to be endotoxin free
(<2.5 U/ml of endotoxin).
Culture of primary rat Leydig cells
Rat Leydig cells were isolated from the testes of
mature rats as previously described (29,30) with some modifications. Briefly,
male Sprague-Dawley rats were sacrificed by CO2
inhalation. The testes were quickly removed, decapsulated and
placed in a 50-ml plastic tube containing 3 ml of collagenase
solution (0.5 mg/ml) and incubated in an oscillating incubator (100
r/min, 34°C) for 30 min. The cell suspension was transferred to a
50-ml tube and kept on ice for 2 min to allow the tubules to
settle. The supernatant containing Leydig cells was filtered
through a 100-µm nylon cell strainer (BD Biosciences, San
Jose, CA, USA). The cells were centrifuged at 1,500 rpm for 10 min
at 4°C. The pellet was resuspended in 2 ml DMEM/F12 and loaded onto
the top of the discontinuous Percoll gradient (21, 26, 37 and 60%)
and centrifuged at 3,000 rpm for 30 min at 4°C. The cells in the
interphase between 37 and 60% were collected and maintained in
DMEM/F12 medium containing 10% FBS, 100 U/ml penicillin and 100
mg/ml streptomycin at 34°C with 5% CO2. The purity of
the Leydig cells was examined by 3β-hydroxysteroid staining and
>90% of the cells stained positive (data not shown).
Determination of cell viability
The effects of AGEs on Leydig cell viability were
evaluated by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT) assay. Rat Leydig cells were plated into 96-well
culture plates. Following 48 h of incubation with various
concentrations of AGEs (25, 50, 100 and 200 µg/ml), 100
µl MTT (5 mg/ml) were added to each well and the cells were
incubated for 2 h at 34°C. The medium was discarded and 150
µl DMSO was then added to each well. The absorbance was
measured at 490 nm using a microplate reader. The results were
expressed as the percentage of MTT reduction, assuming that the
absorbance of the control cells was 100%.
Measurement of testosterone
concentration
The rat Leydig cells cultured in 6-well plates were
pre-incubated with various concentrations of AGEs or BSA for 12 h
and the culture medium was replaced with fresh medium containing
human chorionic gonadotropin (hCG; 4 ng/ml) with or without the
same concentrations AGEs or BSA. Following treatment under
different conditions for 12 h, the medium was collected and
centrifugated 12,000 rpm for 5 min at 4°C, and the supernatant were
collected and assayed for testosterone using ELISA kit. The
sensitivity of the assay was 32 pg/ml. Intra- and inter-assay
variations were below 6.6 and 7.5%, respectively.
Measurement of intracellular reactive
oxygen species (ROS)
Intracellular ROS levels were determined by
measuring the probe, dichlorofluorescein diacetate (DCFH-DA).
Briefly, cells treated with or without AGEs for 12 h were washed
and incubated with fresh medium containing 10 µM DCFH-DA for
30 min in the dark. The medium was then removed and the cells were
washed with PBS. Fluorescence at excitation, 485 nm and emission,
535 nm was measured using a microplate reader (TriStar LB941;
Berthold Technologies, Oak Ridge, TN, USA).
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
The Leydig cells were treated under different
conditions and total RNA was then isolated using TRIzol reagent
following the manufacturer's instructions. cDNA was synthesized
from 1 µg of total RNA using M-MLV reverse transcriptase in
a total reaction volume of 20 µl. The PCR reaction mixtures
contained 10 µl SYBR® Premix Ex Taq™, 0.4
µl of each primer (10 µM), 2 µl template cDNA
and dH2O up to a final volume of 20 µl. The
cycling conditions were 94°C for 40 sec, followed by 40 cycles of
94°C for 15 sec, 64°C for 15 sec, and 72°C for 15 sec. The
following primers were used: StAR forward,
5′-ACCACATCTACCTGCACGCCAT-3′ and reverse,
5′-CCTCTCGTTGTCCTTGGCTGAA-3′; 3β-HSD forward,
5′-AGCAAAAAGATGGCCGAGAA-3′ and reverse,
5′-GGCACAAGTATGCAATGTGCC-3′; P450scc forward,
5′-TTCCCATGCTCAACATGCCTC-3′ and reverse,
5′-ACTGAAAATCACATCCCAGGCAG-3′; β-actin forward,
5′-GGAAATCGTGCGTGACATTAAAG-3′ and reverse,
5′-CGGCAGTGGCCATCTCTT-3′. The relative gene expression levels were
normalized to β-actin using the ∆∆Ct method, where Ct was the cycle
threshold.
Western blot analysis
After being subjected to the various treatments, the
cells were washed twice with PBS and then lysed for 20 min in lysis
buffer (50 mM Tris-HCl, pH 7.5, 250 mM NaCl, 2 mM EDTA, 10%
glycerol, 0.1% NP-40, 0.5 mM PMSF, 10 µg/ml aprotinin, 10
µg/ml leupeptin, 1 mM NaF, 0.1 mM
Na3VO4 and 1 mM dithiothreitol). The protein
concentration was measured by BCA protein assay. Protein (30
µg) was separated by SDS-polyacrylamide gel electrophoresis,
and transferred onto PVDF membranes. The membranes were blocked in
5% non-fat milk powder in Tris-buffered saline (TBS)/0.1% Tween-20
for 1 h at room temperature and incubated with specific antibodies
(3β-HSD, P450scc, StAR, GRP78, CHOP, β-actin) in 5% BSA in TBS at
4°C overnight. After washing, the membranes were incubated with
HRP-conjugated second antibody in TBS for 1 h at room temperature.
Finally, the labeled proteins were detected using the ECL kit.
Densitometric analyses of the bands were performed using ImageJ
software (obtained from the NIH websites, http://rsb.info.nih.gov/nih-image).
Statistical analysis
Statistical comparison was carried out using one-way
analysis of variance (ANOVA). The data represent the means ± SEM.
Values of P<0.05 were considered to indicate statistically
significant differences.
Results
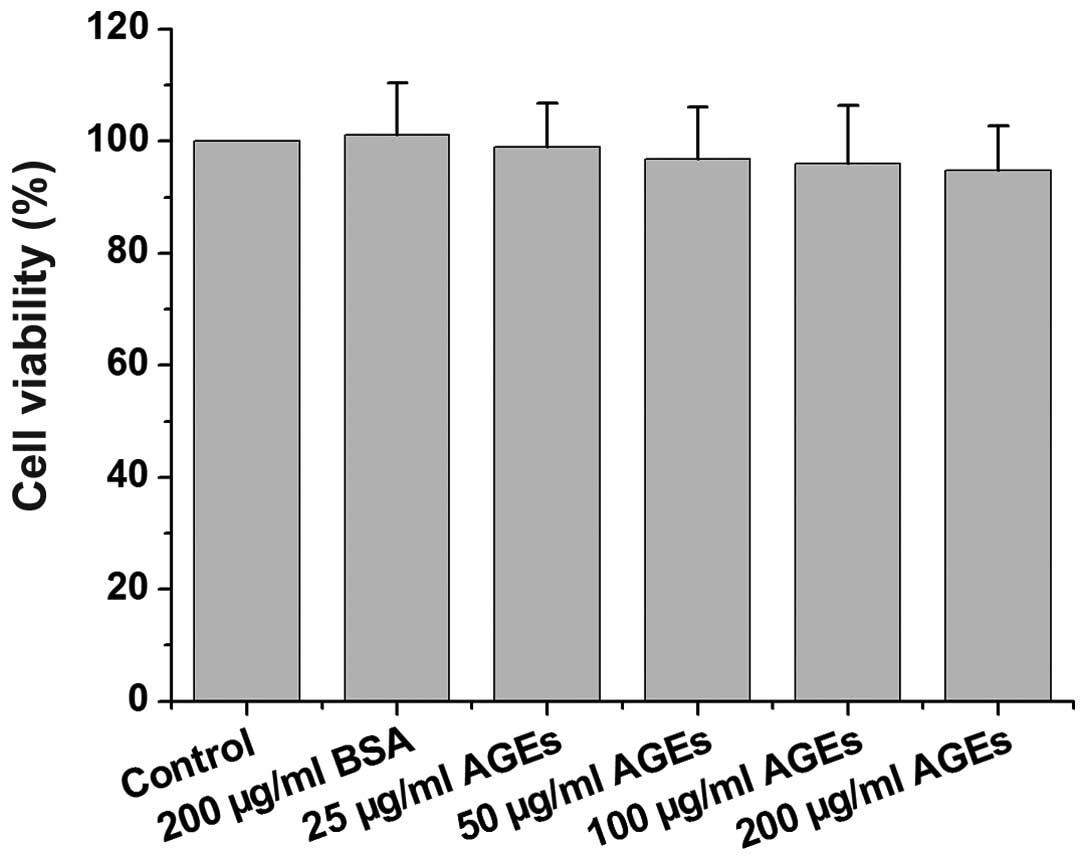
Effect of AGEs on the viability of rat
primary Leydig cells
To assess the effect of AGEs on the viability of rat
primary Leydig cells, the cells were treated with AGEs or BSA for
48 h and MTT assay was then performed. The viability of the cells
treated with AGEs is shown in Fig.
1. The viability of the cells treated with 200 µg/ml
BSA, or with 25, 50, 100 and 200 µg/ml AGEs was 101.1±9.2,
99.0±7.7, 96.8±9.3, 96.0±10.3 and 94.7±8.0% of the control value,
respectively. These data indicated that the viability of the Leydig
cells treated with AGEs (concentrations ≤200 µg/ml) for 48 h
was not significantly altered.
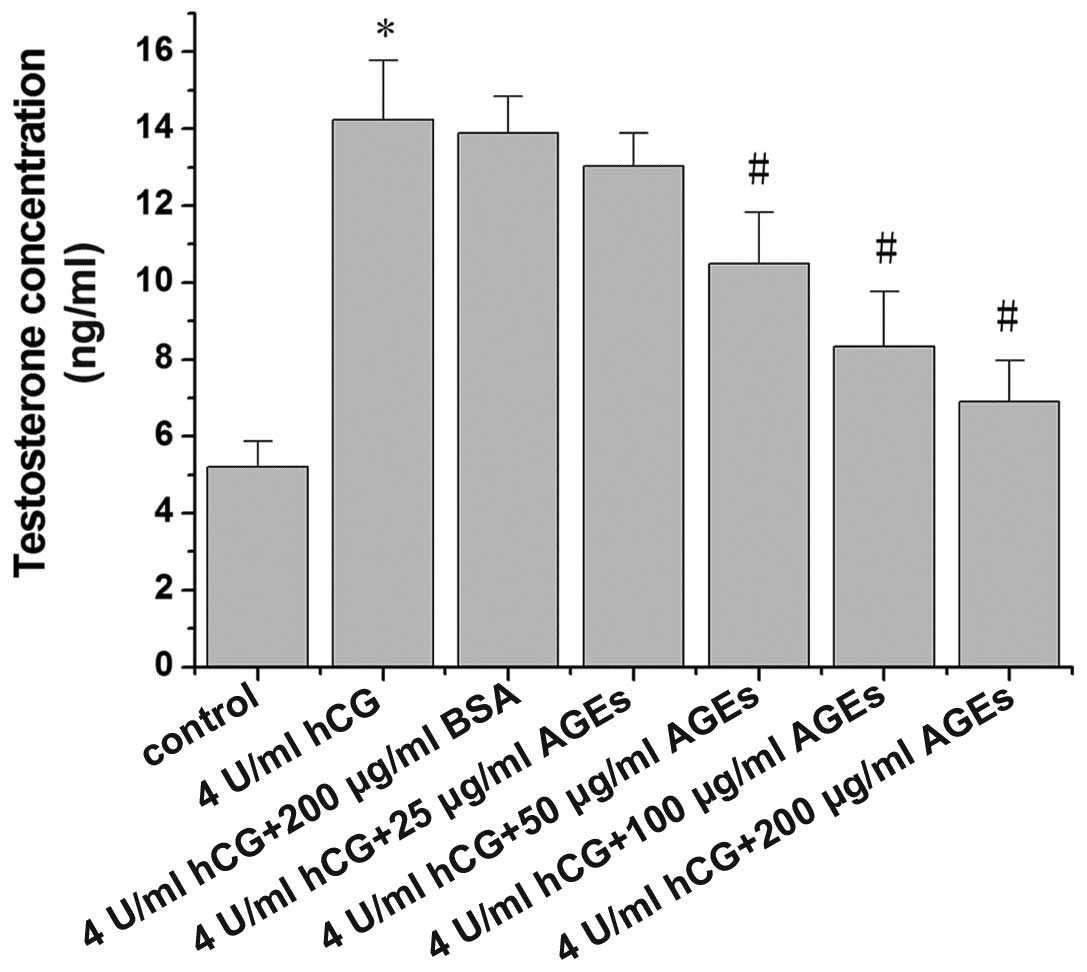
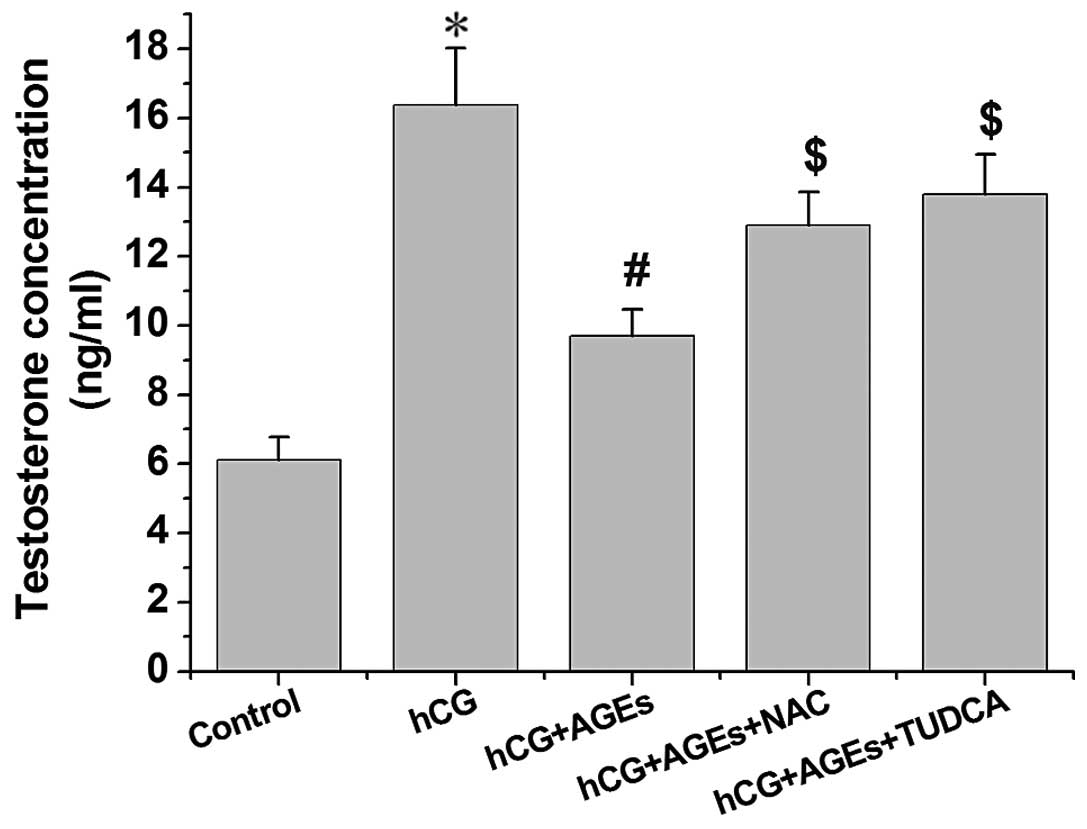
AGEs inhibits testosterone secretion by
hCG-treated Leydig cells
The influence of AGEs on hCG-stimulated testosterone
production in Leydig cells is illustrated in Fig. 2. Exposure to hCG at 4 U/ml induced
a significant increase in testosterone secretion by rat Leydig
cells (P<0.01). Following treatment with AGEs for 24 h,
testosterone secretion by the rat Leydig cells was reduced in a
dose-dependent manner, with significant decreases being observed
from the concentration of 50 µg/ml AGEs.
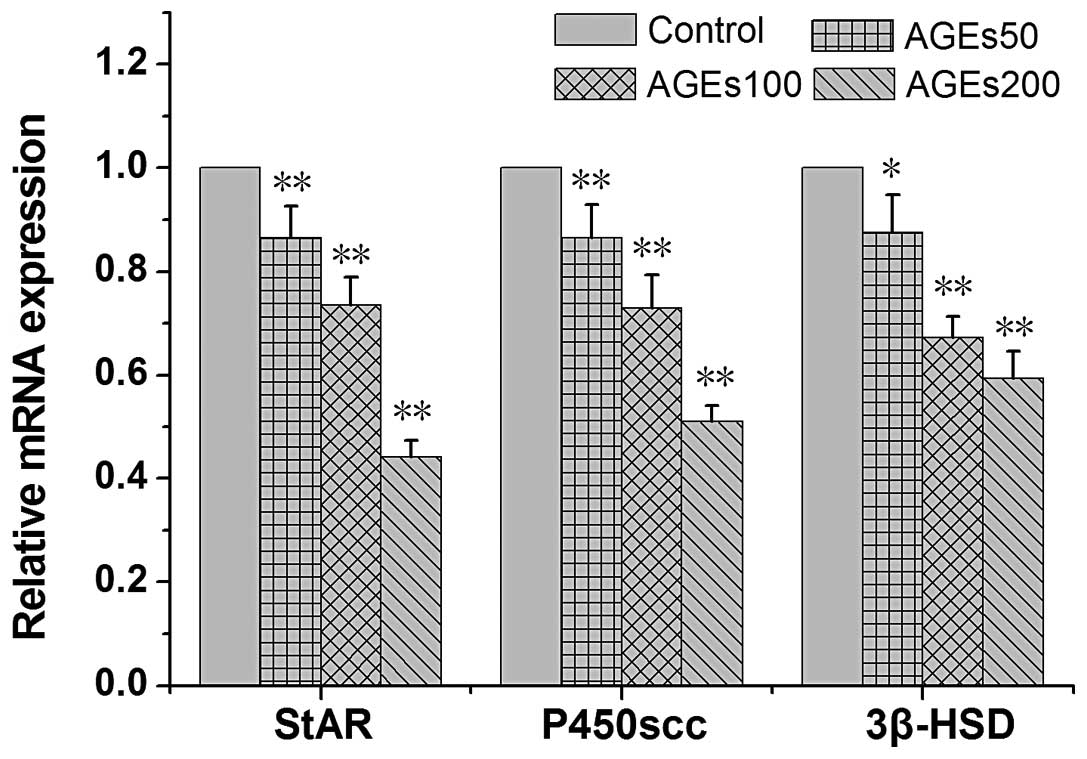
Effects of AGEs on the expression levels
of StAR, P450scc and 3β-HSD
In order to examine the influence of AGEs on the
transcriptional levels of genes related to the testosterone
synthetic pathway in rat Leydig cells, the mRNA levels of StAR,
P450scc and 3β-HSD in rat Leydig cells treated with AGEs for 24 h
were measured. Generally, treatment with AGEs led to a significant
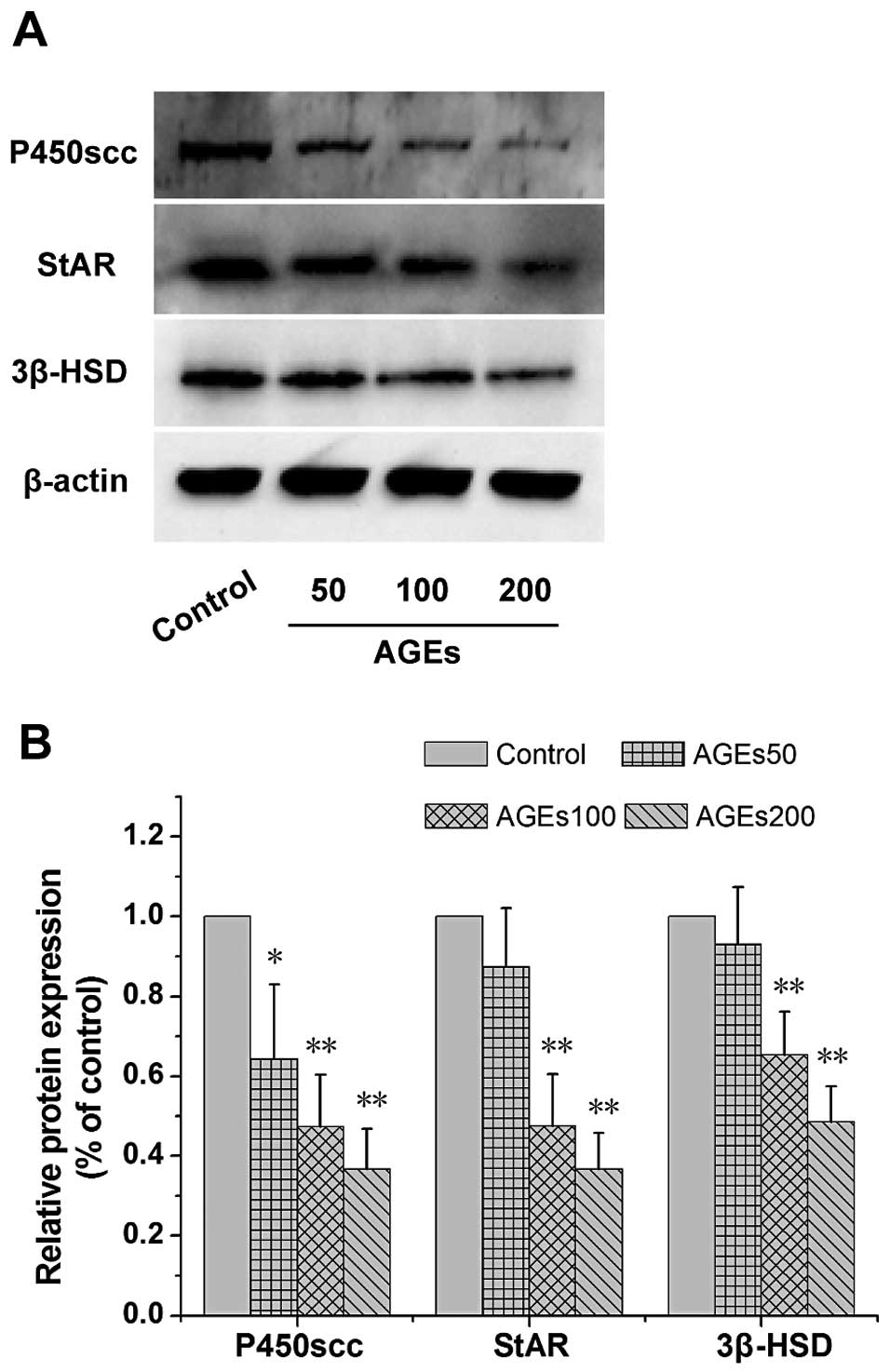
decrease in the mRNA levels of StAR, P450scc and 3β-HSD (Fig. 3). Furthermore, the protein levels
of StAR, P450scc and 3β-HSD in the cells treated with AGEs for 24 h
were investigated. The results revealed that AGEs decreased the
StAR, P450scc and 3β-HSD protein levels in a dose-dependent manner
(Fig. 4).
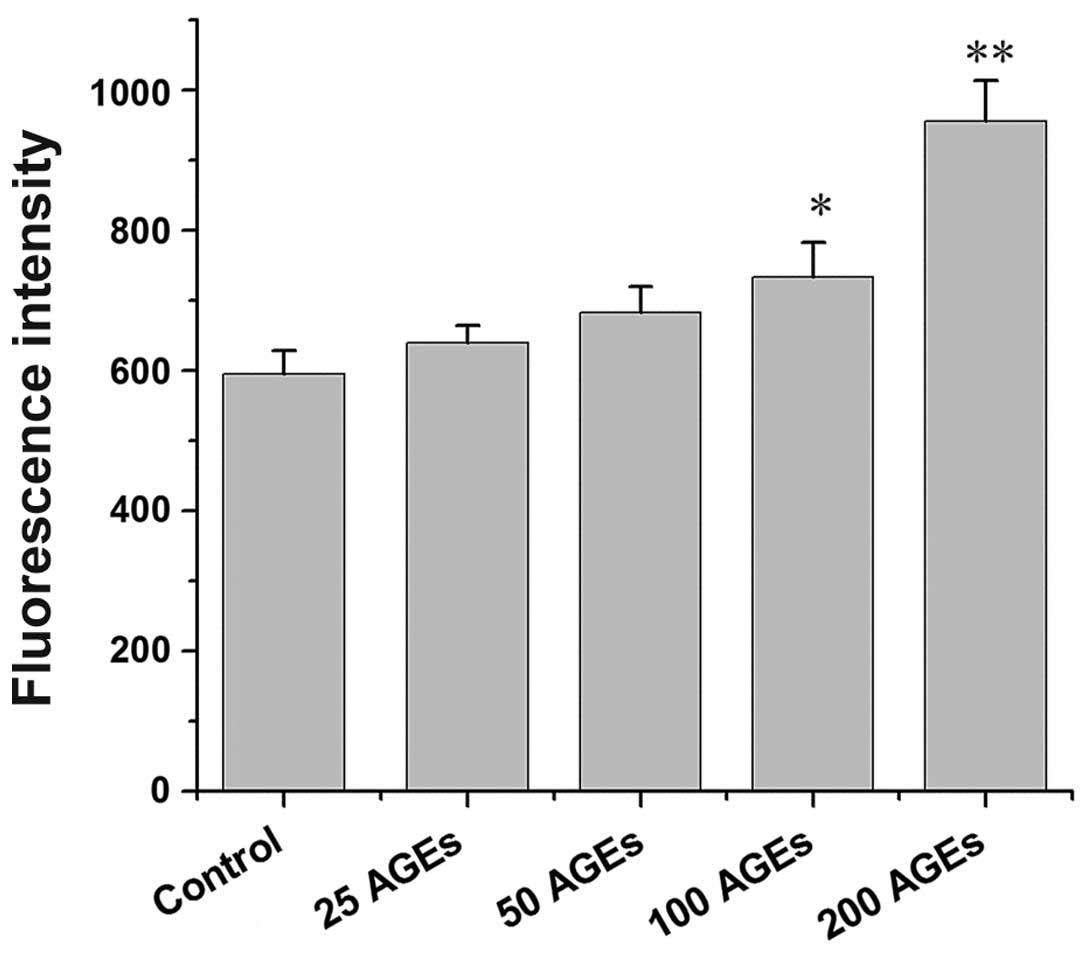
Oxidative stress is involved in the
AGE-induced inhibition of testosterone secretion
Since oxidative stress plays an important role in
AGE-induced dysfunction in other tissues or cells, the role of
oxidative stress in the inhibition of testosterone secretion by
Leydig cells induced by AGEs was investigated. The results revealed
that the AGEs significantly increased the levels of ROS in the rat
Leydig cells in a concentration-dependent manner (Fig. 5). Following pre-treatment with
NAC, an antioxidant agent, the inhibitory effects on testosterone
secretion induced by AGEs were significantly reversed (Fig. 6).
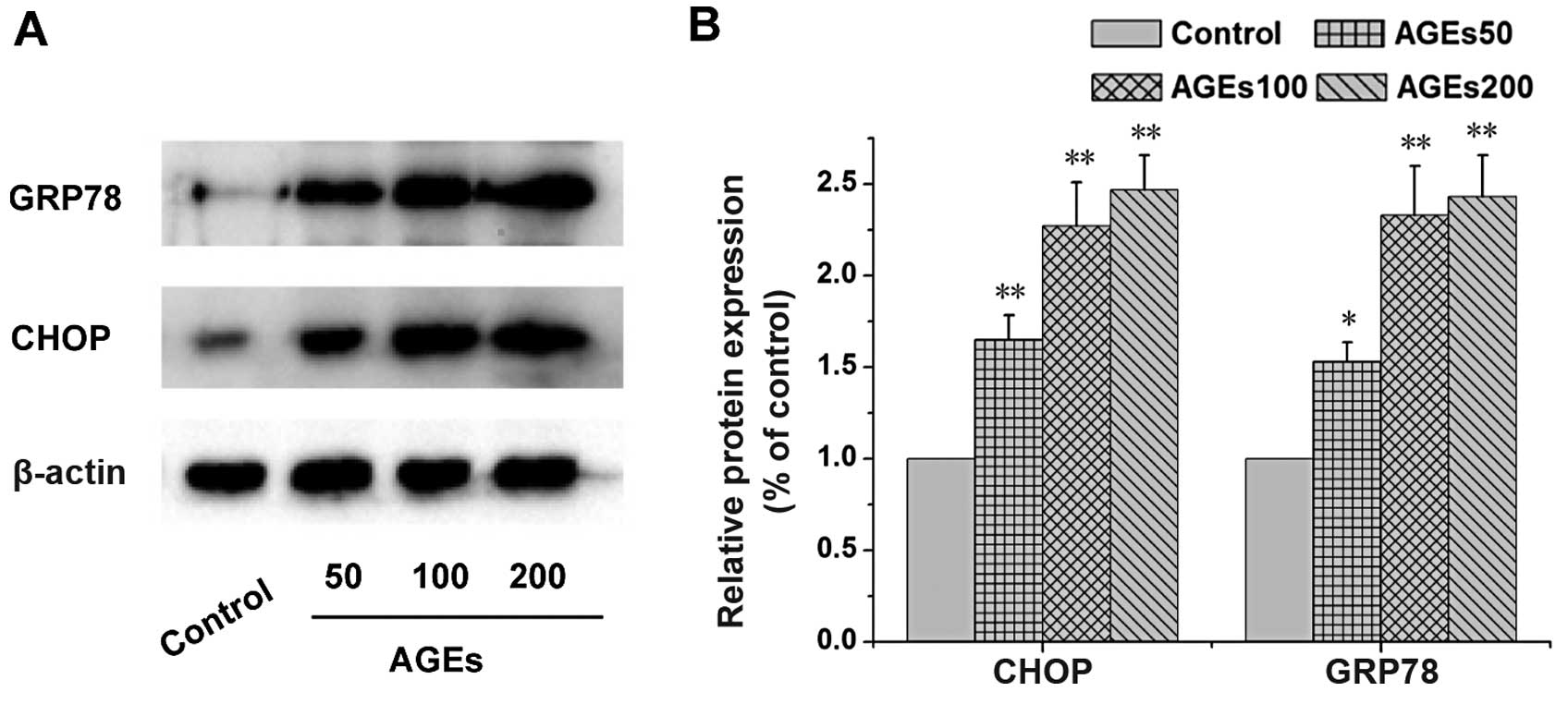
AGEs inhibit testosterone secretion
through ER stress
The Leydig cells were pre-treated with TUDCA, an
endoplasmic reticulum stress inhibitor, and the levels of
testosterone were measured. As shown in Fig. 6, we found that TUDCA significantly
inhibited the AGE-induced decrease in the secretion of testosterone
(P<0.01). The results of western blot analysis revealed that the
expression levels of endoplasmic reticulum stress-related proteins
(CHOP and GRP78) were increased (Fig.
7). In particular, following treatment with 200 µg/ml
AGEs, the expression levels of CHOP and GRP78 were 2.40- and
2.51-fold of those of the control, respectively (P<0.01). These
results indicated that AGEs inhibited the synthesis of testosterone
partly by inducing endoplasmic reticulum stress.
Discussion
It has been demonstrated that in diabetic men and
animal models, diabetes causes reduced testosterone synthesis and
secretion (26,27). Several clinical and animal studies
have focused on the molecular mechanisms responsible for the
alterations induced by diabetes in male reproductive potential,
such as endocrine disorders, neuropathy and increased oxidative
stress (14). A growing body of
evidence has indicated that AGEs are influential instigators,
mediators and/or contributors to male reproductive dysfunction
(19,21,24,25). However, little is known of the
role ofAGEs in Leydig cell function.
In this study, we found that AGEs decreased
testosterone synthesis in rat Leydig cells induced by hCG in a
dose-dependent manner. Testosterone is synthesized from cholesterol
in Leydig cells. Cholesterol, synthesized de novo in the
testes is transported from the outer to the inner mitochondrial
membrane by StAR. Transport across the mitochondrial membrane is
the rate limiting step of testosterone biosynthesis (31). P450scc mediates the conversion of
cholesterol to pregnenolone on the inner mitochondrial membrane,
while the conversion of pregnenolone to progesterone is carried out
by 3β-HSD (32). In this study,,
expression levels of StAR, P450scc and 3β-HSD were investigated. We
observed that StAR, P450scc, 3β-HSD expression was significantly
downregulated in the Leydig cells treated with AGEs both at the
mRNA and protein level. These results indicated that the
AGE-induced decrease in testosterone secretion was in part due to
the inhibition of StAR, P450scc and 3β-HSD expression.
Little is known about the mechanisms through which
AGEs inhibit testosterone synthesis and downregulate the expression
of StAR, P450scc and 3β-HSD. A growing body of evidence has
incicatd that AGEs exacerbate disease progression through two
general mechanisms, cross-linking intracellular, as well as
extracellular matrix proteins, and through binding to their cell
surface receptor, RAGE (33–35). The extracellular effects of AGE
include the modification of structural proteins, usually connective
tissue components, leading to the alteration of structure and
function. On the other hand, AGEs bind RAGE and activate multiple
signaling pathways, such as ROS, p21ras, ERK1/2 (p44/p42)
mitogen-activated protein kinases, p38 and SAPK/JNK
mitogen-activated protein kinases, phosphoinositol-3 kinase and the
JAK/STAT pathways, with important downstream inflammatory
consequences, such as the activation of nuclear factor-κB (NF-κB)
(16,33).
The enhancement of oxidative stress via the
activation of RAGE plays a pivotal role in the pathogenesis of
diabetes complications (24,36,37). In this study, the effect of AGEs
on oxidative stress in Leydig cells was examined. We found that
AGEs significantly increased the generation of ROS in rat Leydig
cells in a concentration-dependent manner. Pre-treatment with the
antioxidant agent, NAC, reversed the inhibitory effects of AGEs on
the synthesis of testosterone (P<0.01). These results
demonstrated that AGEs inhibited the generation of testosterone
partly by inducing oxidative stress.
Furthermore, recent studies have indicated that AGEs
can also adversely affect endoplasmic reticulum function, leading
to pathogenic endoplasmic reticulum stress (38–41). Inhibitors of advanced glycation
acting as potent endoplasmic reticulum stress modulators with
beneficial effects in restoring endoplasmic reticulum homeostasis
and adjusting the physiological unfolded protein response level,
present an emerging therapeutic approach with significant
applications, particularly in the context of metabolic dysfunction
(42). In this study, rat Leydig
cells were pre-treated with TUDCA, an endoplasmic reticulum stress
inhibitor, and the levels of testosterone were measured. The result
revealed that TUDCA significantly inhibited the AGE-induced
decrease in testosterone secretion (P<0.01). To further
investigate the role of endoplasmic reticulum stress in the
AGE-induced inhibition of testosterone secretion by Leydig cells,
the expression levels of endoplasmic reticulum stress-related
protein, endoplasmic reticulum chaperone GRP78 and the
transcription factor CHOP were examined by western blot analysis.
The results revealed that the expression of endoplasmic reticulum
stress-related protein CHOP and GRP78 was markedly upregulated
following treatmetn with AGEs. It can thus be inferred that AGEs
inhibited the generation of testosterone partly by inducing
endoplasmic reticulum stress.
In conclusion, the present study demonstrates that
AGEs inhibit testosterone secretion by rat Leydig cells in
dose-dependent manner. The decrease in testosterone production
induced by AGEs possibly occurs through the induction of oxidative
stress and endoplasmic reticulum stress, and by a decrease in the
levels of StAR, 3β-HSD and P450scc in Leydig cells. This study
reveals the mechanisms underlying the inhibitory effects of AGEs on
testosterone secretion by rat Leydig cells and provides the basis
for further investigation of male reproductive disorders caused by
diabetes.
Acknowledgments
This study was supported by the Project of Enhancing
School with Innovation of Guangdong Ocean University
(GDOU2015050222) and grants from the Natural Science Foundation of
Guangdong Ocean University (nos. 0812270, and 1212340).
References
|
1
|
Guariguata L, Whiting DR, Hambleton I,
Beagley J, Linnenkamp U and Shaw JE: Global estimates of diabetes
prevalence for 2013 and projections for 2035. Diabetes Res Clin
Pract. 103:137–149. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pinhas-Hamiel O, Dolan LM, Daniels SR,
Standiford D, Khoury PR and Zeitler P: Increased incidence of
non-insulin-dependent diabetes mellitus among adolescents. J
Pediatr. 128:608–615. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pinhas-Hamiel O and Zeitler P: The global
spread of type 2 diabetes mellitus in children and adolescents. J
Pediatr. 146:693–700. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nadeau K and Dabelea D: Epidemiology of
type 2 diabetes in children and adolescents. Endocr Res. 33:35–58.
2008. View Article : Google Scholar
|
|
5
|
Urakami T, Kubota S, Nitadori Y, Harada K,
Owada M and Kitagawa T: Annual incidence and clinical
characteristics of type 2 diabetes in children as detected by urine
glucose screening in the Tokyo metropolitan area. Diabetes Care.
28:1876–1881. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jangir RN and Jain GC: Diabetes mellitus
induced impairment of male reproductive functions: a review. Curr
Diabetes Rev. 10:147–157. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sexton WJ and Jarow JP: Effect of diabetes
mellitus upon male reproductive function. Urology. 49:508–513.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baccetti B, La Marca A, Piomboni P,
Capitani S, Bruni E, Petraglia F and De Leo V: Insulin-dependent
diabetes in men is associated with hypothalamo-pituitary
derangement and with impairment in semen quality. Hum Reprod.
17:2673–2677. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schoeller EL, Schon S and Moley KH: The
effects of type 1 diabetes on the hypothalamic, pituitary and
testes axis. Cell Tissue Res. 349:839–847. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Scarano WR, Messias AG, Oliva SU,
Klinefelter GR and Kempinas WG: Sexual behaviour, sperm quantity
and quality after short-term streptozotocin-induced hyperglycaemia
in rats. Int J Androl. 29:482–488. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Agbaje IM, Rogers DA, McVicar CM, McClure
N, Atkinson AB, Mallidis C and Lewis SE: Insulin dependant diabetes
mellitus: implications for male reproductive function. Hum Reprod.
22:1871–1877. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ricci G, Catizone A, Esposito R, Pisanti
FA, Vietri MT and Galdieri M: Diabetic rat testes: morphological
and functional alterations. Andrologia. 41:361–368. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mallidis C, Agbaje I, McClure N and
Kliesch S: The influence of diabetes mellitus on male reproductive
function: a poorly investigated aspect of male infertility. Urologe
A. 50:33–37. 2011.In German. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
La Vignera S, Condorelli R, Vicari E,
D'Agata R and Calogero AE: Diabetes mellitus and sperm parameters.
J Androl. 33:145–153. 2012. View Article : Google Scholar
|
|
15
|
Alves MG, Martins AD, Cavaco JE, Socorro S
and Oliveira PF: Diabetes, insulin-mediated glucose metabolism and
Sertoli/blood-testis barrier function. Tissue Barriers.
1:e239922013. View Article : Google Scholar
|
|
16
|
Vlassara H and Uribarri J: Advanced
glycation end products (AGE) and diabetes: cause, effect, or both?
Curr Diab Rep. 14:4532014. View Article : Google Scholar :
|
|
17
|
Vlassara H and Striker GE: Advanced
glycation endproducts in diabetes and diabetic complications.
Endocrinol Metab Clin North Am. 42:697–719. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vlassara H and Striker GE: AGE restriction
in diabetes mellitus: a paradigm shift. Nat Rev Endocrinol.
7:526–539. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mallidis C, Agbaje I, Rogers D, Glenn J,
McCullough S, Atkinson AB, Steger K, Stitt A and McClure N:
Distribution of the receptor for advanced glycation end products in
the human male reproductive tract: prevalence in men with diabetes
mellitus. Hum Reprod. 22:2169–2177. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Moschonas DP, Piperi C, Korkolopoulou P,
Levidou G, Kavantzas N, Trigka EA, Vlachos I, Arapostathi C, Perrea
D, Mitropoulos D, et al: Impact of diet-induced obesity in male
mouse reproductive system: the role of advanced glycation end
product-receptor for advanced glycation end product axis. Exp Biol
Med (Maywood). 239:937–947. 2014. View Article : Google Scholar
|
|
21
|
Mallidis C, Agbaje IM, Rogers DA, Glenn
JV, Pringle R, Atkinson AB, Steger K, Stitt AW and McClure N:
Advanced glycation end products accumulate in the reproductive
tract of men with diabetes. Int J Androl. 32:295–305. 2009.
View Article : Google Scholar
|
|
22
|
O'Neill J, Czerwiec A, Agbaje I, Glenn J,
Stitt A, McClure N and Mallidis C: Differences in mouse models of
diabetes mellitus in studies of male reproduction. Int J Androl.
33:709–716. 2010. View Article : Google Scholar
|
|
23
|
Mallidis C, Czerwiec A, Filippi S, O'Neill
J, Maggi M and McClure N: Spermatogenic and sperm quality
differences in an experimental model of metabolic syndrome and
hypogonadal hypogonadism. Reproduction. 142:63–71. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Karimi J, Goodarzi MT, Tavilani H,
Khodadadi I and Amiri I: Relationship between advanced glycation
end products and increased lipid peroxidation in semen of diabetic
men. Diabetes Res Clin Pract. 91:61–66. 2011. View Article : Google Scholar
|
|
25
|
Karimi J, Goodarzi MT, Tavilani H,
Khodadadi I and Amiri I: Increased receptor for advanced glycation
end products in spermatozoa of diabetic men and its association
with sperm nuclear DNA fragmentation. Andrologia. 44(Suppl 1):
280–286. 2012. View Article : Google Scholar
|
|
26
|
Cheung KK, Luk AO, So WY, Ma RC, Kong AP,
Chow FC and Chan JC: Testosterone level in men with type 2 diabetes
mellitus and related metabolic effects: a review of current
evidence. J Diabetes Investig. 6:112–123. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
El Baba K and Azar ST: Low testosterone
and diabetes. Curr Diabetes Rev. 9:418–421. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Iwashima Y, Eto M, Horiuchi S and Sano H:
Advanced glycation end product-induced peroxisome
proliferator-activated receptor gamma gene expression in the
cultured mesangial cells. Biochem Biophys Res Commun. 264:441–448.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chemes H, Cigorraga S, Bergadá C,
Schteingart H, Rey R and Pellizzari E: Isolation of human Leydig
cell mesenchymal precursors from patients with the androgen
insensitivity syndrome: testosterone production and response to
human chorionic gonadotropin stimulation in culture. Biol Reprod.
46:793–801. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Klinefelter GR, Hall PF and Ewing LL:
Effect of luteinizing hormone deprivation in situ on
steroidogenesis of rat Leydig cells purified by a multistep
procedure. Biol Reprod. 36:769–783. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bose HS, Lingappa VR and Miller WL: The
steroidogenic acute regulatory protein, StAR, works only at the
outer mitochondrial membrane. Endocr Res. 28:295–308. 2002.
View Article : Google Scholar
|
|
32
|
Payne AH and Hales DB: Overview of
steroidogenic enzymes in the pathway from cholesterol to active
steroid hormones. Endocr Rev. 25:947–970. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ott C, Jacobs K, Haucke E, Navarrete
Santos A, Grune T and Simm A: Role of advanced glycation end
products in cellular signaling. Redox Biol. 2:411–429. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jahan H and Choudhary MI: Glycation,
carbonyl stress and AGEs inhibitors: a patent review. Expert Opin
Ther Pat. 25:1267–1284. 2015.PubMed/NCBI
|
|
35
|
Jin X, Yao T, Zhou Z, Zhu J, Zhang S, Hu W
and Shen C: Advanced glycation end products enhance macrophages
polarization into M1 phenotype through activating RAGE/NF-κB
pathway. BioMed Res Int. 2015:7324502015. View Article : Google Scholar
|
|
36
|
Amaral S, Oliveira PJ and Ramalho-Santos
J: Diabetes and the impairment of reproductive function: possible
role of mitochondria and reactive oxygen species. Curr Diabetes
Rev. 4:46–54. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wautier MP, Chappey O, Corda S, Stern DM,
Schmidt AM and Wautier JL: Activation of NADPH oxidase by AGE links
oxidant stress to altered gene expression via RAGE. Am J Physiol
Endocrinol Metab. 280:E685–E694. 2001.PubMed/NCBI
|
|
38
|
Rong G, Tang X, Guo T, Duan N, Wang Y,
Yang L, Zhang J and Liang X: Advanced oxidation protein products
induce apoptosis in podocytes through induction of endoplasmic
reticulum stress. J Physiol Biochem. 71:455–470. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu J, Xiong M, Huang B and Chen H:
Advanced glycation end products upregulate the endoplasmic
reticulum stress in human periodontal ligament cells. J
Periodontol. 86:440–447. 2015. View Article : Google Scholar
|
|
40
|
Wu L, Wang D, Xiao Y, Zhou X, Wang L, Chen
B, Li Q, Guo X and Huang Q: Endoplasmic reticulum stress plays a
role in the advanced glycation end product-induced inflammatory
response in endothelial cells. Life Sci. 110:44–51. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu J, Huang K, Cai GY, Chen XM, Yang JR,
Lin LR, Yang J, Huo BG, Zhan J and He YN: Receptor for advanced
glycation end-products promotes premature senescence of proximal
tubular epithelial cells via activation of endoplasmic reticulum
stress-dependent p21 signaling. Cell Signal. 26:110–121. 2014.
View Article : Google Scholar
|
|
42
|
Piperi C, Adamopoulos C, Dalagiorgou G,
Diamanti-Kandarakis E and Papavassiliou AG: Crosstalk between
advanced glycation and endoplasmic reticulum stress: emerging
therapeutic targeting for metabolic diseases. J Clin Endocrinol
Metab. 97:2231–2242. 2012. View Article : Google Scholar : PubMed/NCBI
|