Thalidomide was initially synthesized as a
non-addictive, non-barbiturate sedative by the German
pharmaceutical company Chemie-Grünenthal in the late 1950's. It was
discovered to be an effective antiemetic and was then used to
control symptoms of nausea and vomiting in pregnant women. It was
not until 1961 that thalidomide was validated as the cause of the
largest man-made medical disaster in history with an estimated
10,000 children in 46 countries born with birth defects.
Subsequently, thalidomide was withdrawn from most countries. In the
following decades, the immunomodulatory and antiangiogenic effects
of thalidomide were discovered and recognised as a novel indication
for treatment with thalidomide. In 1965, Sheskin (1) serendipitously discovered that the
immunomodulatory effects of thalidomide could be used to
successfully cure erythema nodosum leprosum. In 1994, D'Amato et
al (2) discovered that orally
administered thalidomide inhibits angiogenesis induced by basic
fibroblast growth factor (FGF) in a rabbit cornea micropocket
assay. Due to these mechanisms, thalidomide was used to treat
multiple myeloma and shown to be highly effective (3). To date, studies have been performed
to examine the efficacy of thalidomide in the treatment of solid
tumors, such as prostate cancer, glioblastoma and squamous cell
carcinoma of the neck (4–8).
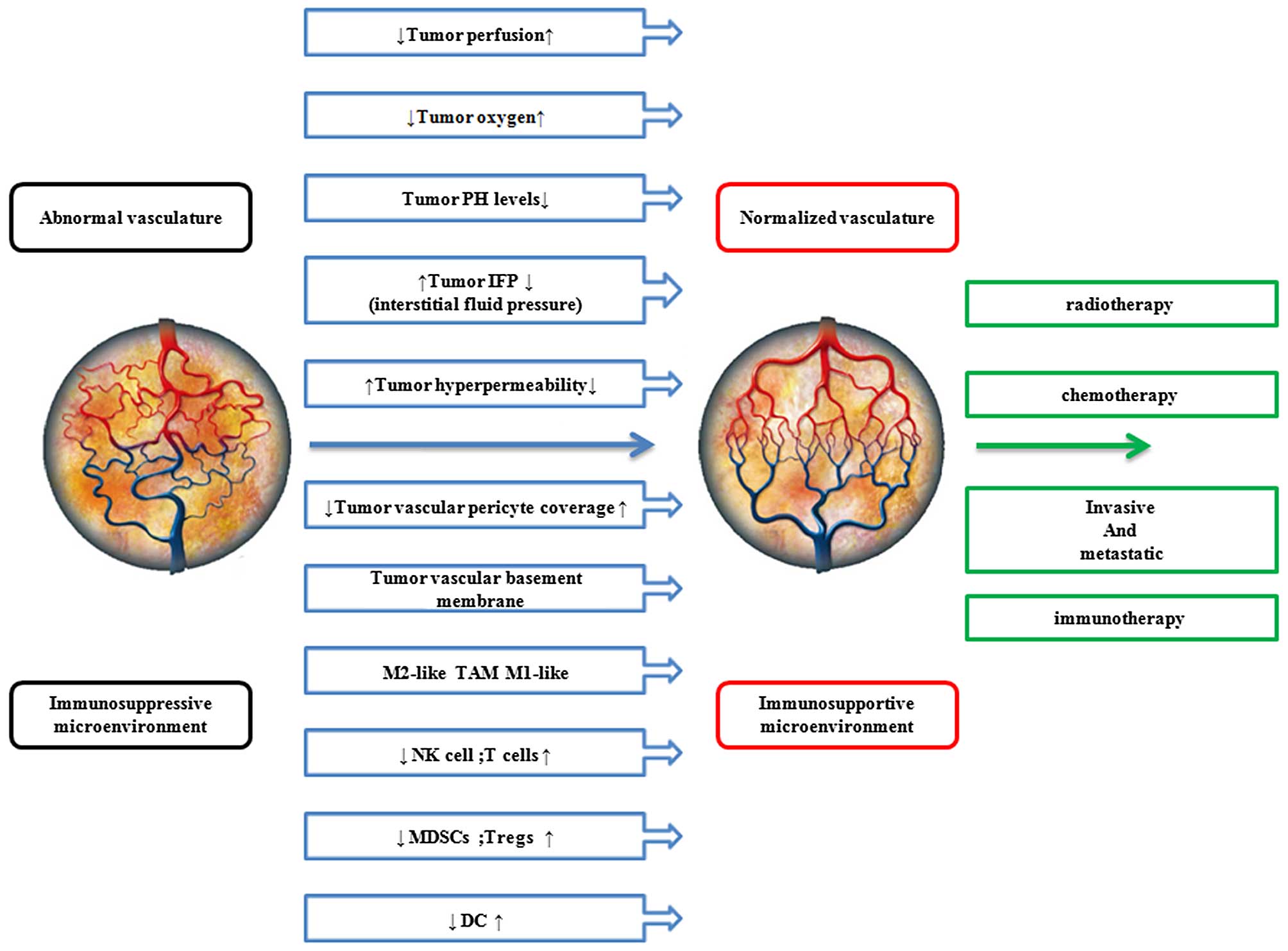
In 2005, Jain suggested that certain antiangiogenic
agents can also 'normalize' the abnormal structure and function of
tumor vasculature to make it more efficient for oxygen and drug
delivery (9). Subsequently,
several studies of thalidomide suggested that it reduces tumor
vessel density and tumor interstitial fluid pressure, while
improving perfusion and oxygenation in preclinical fibrosarcoma and
liver carcinoma models. Although the time of improved oxygenation
only lasted for 2–4 days of thalidomide treatment (10), it demonstrated that thalidomide
probably temporarily reversed tumor vessels towards a more normal
phenotype. In vitro, thalidomide analogs also inhibited
proliferative effects on human umbilical vein endothelial cells
(11). These studies explored the
vascular remodeling effect of thalidomide without associating
vascular normalization with effects on immune cells. The
immunomodulatory properties of thalidomide have been the focus of
mechanistic studies of the therapeutic effects of thalidomide in
patients with different types of cancer (12–14). The potent immunomodulatory
activity of thalidomide mainly alters the secretion and activity of
various cytokines (12–18). To some extent, these cytokines
control the balance between pro-angiogenic and anti-angiogenic
effects. Thalidomide has also been demonstrated to regulate immune
cells, such as regulatory T cells (Treg) and natural killer (NK)
cells, which resulted in enhanced tumor surveillance defence
(13). The potential 'vascular
normalization' of thalidomide may also create a microenvironment of
immune-activating cells in tumors, thereby leading to immune cell
polarization and cytokine changes. Thus, we suggest that the
interaction between immune cells and normal tumor vasculature is
the principal mechanism responsible for the effects of thalidomide
in the treatment of cancer.
In this review, we have discussed the regulation of
the interplay between tumor vessels and immune cells by
thalidomide. Targeted therapy with thalidomide may improve the
efficacy of anticancer therapy.
The mechanism responsible for the antiangiogenic
effects of thalidomide has been discovered and defined. D'Amato
et al have determined the critical role of angiogenesis in
the progression and metastasis of cancer, based on antiangiogenic
effects in rabbits, and thalidomide was first examined as a novel
agent for the treatment of patients with resistant myeloma
(2,3). Subsequently, the potential
antiangiogenic effects of thalidomide were also demonstrated in
humans and several other species (19–22). Many clinical and preclinical
investigations showed that the antiangiogenic activity of
thalidomide is associated with a series of events as well as
certain molecules. Verheul et al (23) demonstrated that thalidomide is
capable of inhibiting the effects of FGF2 or vascular endothelial
growth factor (VEGF) on tumor growth in an animal model. The
decreased plasma levels of angiogenic growth factors in patients
with multiple myeloma treated with thalidomide were documented by
Bertolini et al (24).
Such effects were also observed in other types of cancer, such as
lung adenocarcinoma as well as head and neck squamous cell
carcinoma (25,26). Furthermore, the effects of
insulin-like growth factor (IGF)-1 and FGF2 can be reversed by
thalidomide. Thalidomide eliminates their stimulation of the
transcription of αv and β3 integrin subunit genes through
intercalating into the GC box in the promoters of IGF-1 and FGF2
genes, which inhibits the angiogenic effects of the αvβ3 integrin
dimer (27,28). It has also been noted that
thalidomide is involved in the inhibition of cyclooxygenase-2
(COX-2), a key enzyme responsible for the formation of
prostaglandins (PGs) (29). COX-2
was shown to be required for angiogenesis in a rat corneal model
and COX-2-derived PGE2, which is a potent inducer of the angiogenic
switch during the progression of mammary cancer (30,31). Many important molecules involved
in embryonic vascular development and patterning have been reported
to exhibit changed expression patterns following thalidomide
exposure, for example, actin, tubulin, integrins, VEGF,
platelet-derived growth factor (PDGF)β, nitric oxide, ceramide,
angiopoietins (Ang), Notch, hypoxia-inducible factor (HIF),
Slit2/Robo signaling and reactive oxygen species (27,28,32–35). These molecules are associated with
angiogenesis in both physiological and pathological states, and
thalidomide can inhibit angiogenesis through affecting changes in
the expression of these molecules. However, it remains unclear
whether the mechanism of thalidomide regulation can exert different
effects on tumor angiogenesis.
It is now well established that angiogenesis plays a
major role in the aggressive behavior and metastatic potential of
various solid tumors. However, the antitumor effects of thalidomide
did not always correlate with reductions in tumor microvessels in
patients. Despite observing a strong association between
thalidomide and decreased microvessel density in xenografted
primary human myeloma cells in a mouse model which responded to
treatment, it was not possible to conclude whether reduced
microvessel density is a primary result of the anti-angiogenic
activity of thalidomide or secondary to reduced tumor burden
(2). Patients or the animal model
treated with thalidomide appeared to exhibit survival benefits due
to the alleviation of hypoxia and the increased
perfusion/oxygenation as well as the reduced microvessel density of
the tumor (3). Thus, thalidomide
was discovered to induce tumor reoxygenation and extensive vascular
remodeling when combined with radiotherapy and chemotherapy
(10,36). These findings suggest that
thalidomide is capable of remodeling tumor vessels and promoting
the normalization of tumor vasculature. Therefore, the mechanisms
addressing the anti-angiogenic activity of thalidomide are complex
and yet to be completely elucidated. In addition, it was discovered
that thalidomide and its analogs can decrease levels of
pro-angiogenic factors and increase anti-angiogenic factors in
vivo and in vitro. Lebrin et al analyzed whether
the target vessel effect of thalidomide in individuals with
hereditary hemorrhagic telangiectasia (HHT) correlated with PDGF-B
(37). Thalidomide treatment
stimulated mural cell coverage and rescued vessel wall defects. It
also increased PDGF-B expression in endothelial cells and
stimulated mural cell activation. Moreover, simultaneous treatment
of postnatal wild-type mice with a high dose of thalidomide and
imatinib mesylate, a potent inhibitor of PDGF receptor tyrosine
kinase activity, was sufficient to restore postnatal retinal
vascular development. Thus, blocking PDGF signaling counteracts the
effects of thalidomide on angiogenesis and on the apposition of
pericytes to the endothelial cells of the blood vessels. Therefore,
the data has established PDGF as a key mediator of the effects of
thalidomide. This study established the association between
thalidomide and vascular normalization in HHT. At the same time,
the investigator discovered that vascular normalization is
dependent on the dose of thalidomide. High doses of thalidomide
(100 µg ml−1) caused vessel pruning and inhibited
vessel formation. By contrast, lower doses (10–50 µg
ml−1) enhanced endothelial sprouting, and low
concentrations (25 µg ml−1) stimulated the
recruitment of mural cells to the vessel branches (37). Clinical manifestations of HHT
include arteriovenous malformations, which resemble the morphology
of heterogeneous tumor vessels. The data demonstrated the ability
of thalidomide to induce vessel maturation, which could be a
potential indication for the study of thalidomide in cancer
treatment. The vascular normalization hypothesis proposes that
direct or indirect antiangiogenic therapy usually restores the
imbalance between pro- and anti-angiogenic factors back towards
equilibrium. As a result, vessel structure and function become
normal (38).
Cytokines are inflammatory mediators that play
important roles in the pathogenesis of cancer. Over the last few
decades, efforts have focused on ways to use cytokines to
manipulate the host immune response towards recognising and
eradicating cancer cells (39).
Thalidomide regulates the secretion and activity of various
cytokines including interleukin (IL), tumor necrosis factor (TNF),
interferon (IFN) and growth factor. These modulatory effects of
thalidomide may contribute to the suppression of proliferation,
inflammation and angiogenesis as well as of the immune system.
Thalidomide can inhibit the effects of FGF2 or VEGF resulting in
anti-angiogenic effects on tumor growth in animal model (28). As a pro-inflammatory cytokine,
TNF-α plays a critical role in cancer. Thalidomide inhibited TNF-α
synthesis by increasing the degradation of TNF-α mRNA which
resulted in the decreased expression of multiple cell surface
adhesion molecules. These broad effects have been attributed to the
blocking of the activation of nuclear factor-κB (NF-κB) which is a
key transcriptional regulator of inflammatory genes. Thalidomide
achieved the modulations by suppressing IκB kinase activity and
contributing to the suppression of the proliferation of cancer
cells. On the one hand, thalidomide acts to costimulate
CD8+ T cells resulting in the enhanced production of
IL-2, IL-12 and IFN-α (17). On
the other hand, IL-2 in turn enhances T cell proliferation whereas
IL-12 and IFN-α activate NK cells to eliminate cancer cells
(12,13). Thus, thalidomide significantly
enhances antiangiogenic and antitumor activities by regulating
changes in cytokine expression in the tumor milieu. Moreover,
thalidomide also increases the susceptibility of cancer cells to
apoptosis by downregulating the antiapoptotic protein Bcl-2 and
enhancing the sensitivity to Fas-induced apoptosis, and
downregulating NF-κB activity (40,41). In addition to immunomodulatory
properties, thalidomide is associated with DNA damage and oxidative
stress through a signaling pathway mediating different cytokines.
In 2010, thalidomide was found to bind to the protein cereblon
(21) which led to the activation
of immune cells and the secretion of cytokines. Thus, another novel
signaling pathway between cytokines and immune cells via the
downstream targets of the complex with thalidomide was established.
While low amounts of cereblon in multiple myeloma cells correlate
with clinical drug resistance and poor survival outcomes (42), this novel mechanism of thalidomide
may lead to the identification of a new class of therapeutics for
treatment of another type of cancer.
Since the vascular changes alone are unlikely to
explain the inhibition of tumor growth by thalidomide, the immune
response of tumors through vascular normalization is probably
involved. The vascular system ensures the delivery of immune cells
to all organs and tissues (43).
Therefore abnormal tumor vasculature creates a hypoxic
microenvironment that polarizes immune cells toward immune
suppression. Simultaneously, hypoxia alters various metabolic
pathways in cancer cells leading to the accumulation of
immunosuppressive metabolites (44,45). Hence, treatments resulting in
vascular normalization may be an effective modality to potentiate
immunomodulatory effects (46).
Thalidomide, an agent possessing the potential function of vascular
normalization, stimulates vessel maturation and alleviates hypoxia,
so the normalization of the tumor microenvironment by repairing the
function of tumor vessels may be a promising strategy to enhance
immunomodulatory effects and slow tumor progression. RGS5
deficiency and the histidine-rich glycoprotein (HRG) have been
demonstrated to show a mechanistic link between vessel
normalization and enhancing immune cell infiltration and function
(47,48). Many studies have demonstrated the
benefits of antiangiogenic treatment on the efficacy of cancer
immunotherapy in terms of pharmacological and clinical implications
(Table I). These results suggest
that there is a strong correlation between immune cells and
vascular normalization, as shown in Fig. 1. This may represent a potential
mechanism of thalidomide.
Thalidomide was demonstrated to transiently
'normalize' the abnormal structure and function of tumor
vasculature in order to enhance the efficiency of oxygen and drug
delivery (20). It has also been
shown to possess potent anti-inflammatory and immunomodulatory
activities. Tumor-related angiogenesis supports tumor growth and is
also a main obstacle for successful immune therapy as it prevents
the migration of immune effector cells into established tumor
parenchyma. Thalidomide potentially remodeled tumor vessels and
then attracted immune effector cells into the established tumor
parenchyma. Thalidomide probably polarizes the immune cell
phenotype and forms an immunosupportive tumor microenvironment by
vascular normalization resulting in the inhibition of tumor growth.
The molecular mechanisms responsible for the effects of thalidomide
on immune cells and vascular normalization will be the focus of
future studies as therapeutic effects in patients with different
types of cancer were associated with both properties.
In clinical investigations, the type and density of
intra-tumoral immune cells were validated as reliable parameters
for the evaluation of patient clinical outcomes in certain types of
cancer (49–51). Therapies targeting immune cells
has been an efficient strategy for treating tumors. Although the
immunomodulatory activity of thalidomide mainly lies in its
capability to alter the secretion and activity of various
cytokines, thalidomide has been extensively investigated for T cell
costimulation and modulation of the activities of NK cells.
Thalidomide suppressed tumor growth by activating T cells in
HIV-infected individuals and NK cells in mice (13,17). Lenalidomide and pomalidomide,
which are thalidomide analogues, strongly inhibit the proliferation
of Tregs as well as suppressing the function of Tregs. This outcome
suggests that thalidomide may potentially inhibit Tregs, which may
help to overcome an important barrier to tumor-specific immunity in
cancer patients (52).
An efficient immune response depends on the
appropriate distribution and positioning of immune cells within
dynamic tissue microenvironments. The different immune cells can
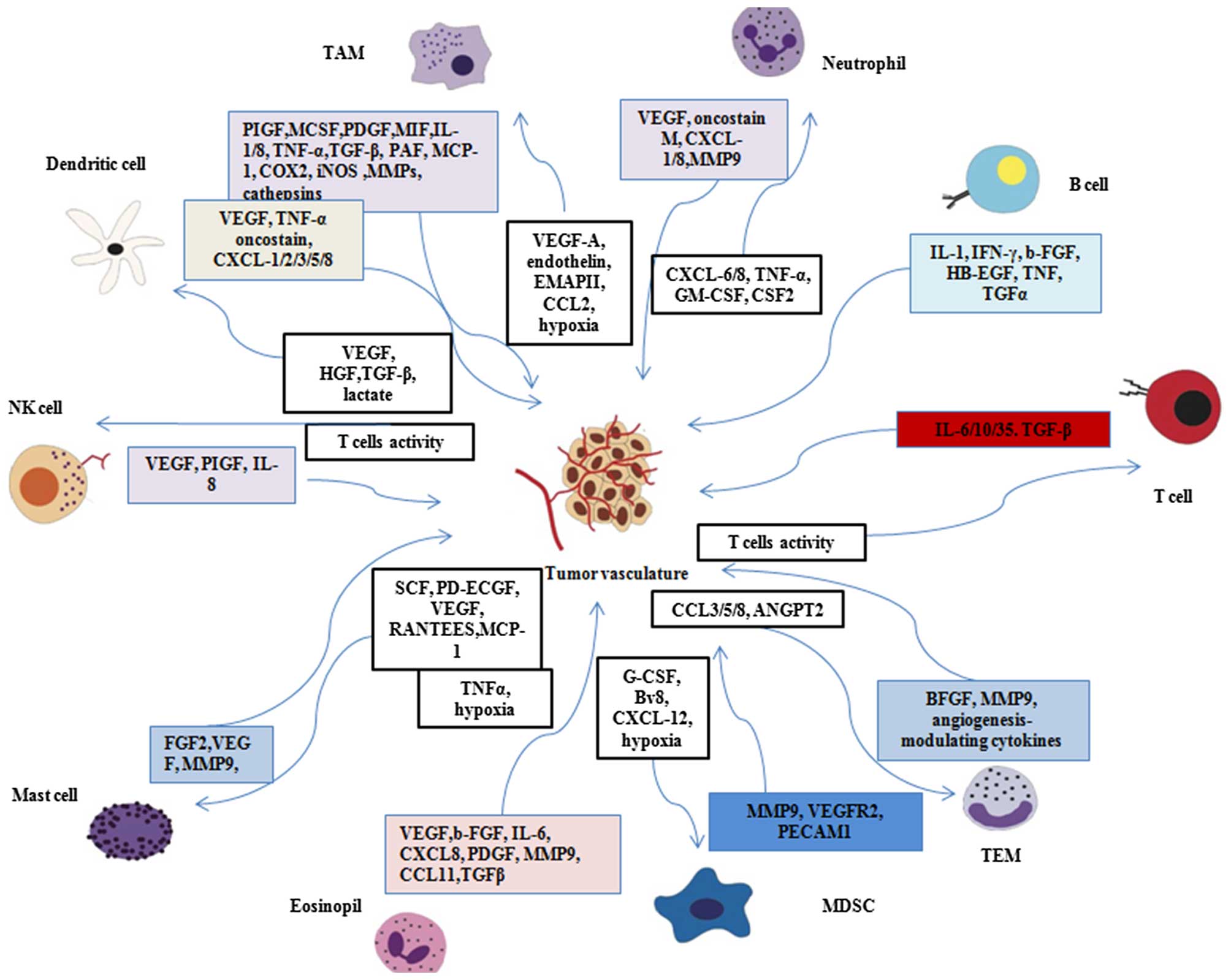
also affect blood vessel neoformation and remodeling (53–55). As summarized in Fig. 2, every type of immune cell has
been shown to affect the process of tumor angiogenesis either
directly or indirectly by releasing regulatory cytokines.
Thalidomide may probably affect angiogenesis and vascular remolding
by affecting immune cells.
General immune cells exert functions through the
release of cytokines. In solid tumors, macrophages are often
attracted to the hypoxic areas of tumor sites due to the secretion
of hypoxia-induced chemoattractants by tumor cells. Such
chemoattractants include different cytokines such as VEGF,
endothelin, endothelial monocyte-activating polypeptide II (EMAP
II) (56) and CCL2 (57). Subsequently, tumor-associated
macrophages (TAMs) are recruited to the hypoxic areas. This
microenvironment promotes the metabolic adaptation of TAMs to
hypoxia by upregulating HIF-1, HIF-2 and VEGF (58–60). Macrophages are attracted to the
hypoxic areas of tumor site, which produce a large amount of
pro-angiogenic factors, in addition to angiogenesis-modulating
enzymes under the regulation of specific signaling pathways (i.e.,
NF-κB and mTOR) and transcription factors (i.e., HIFs and Stat3),
which contribute to tumor angiogenesis. These molecules can switch
angiogenesis or remold tumor vessels, resulting in the alleviation
of hypoxia and increased tumor perfusion/oxygenation. A study
determined the effects of thalidomide in the pathogenesis of
asthmatic airways using a mouse model of allergic asthma as well as
the underlying mechanism responsible for these effects. When the
ovalbumin-sensitized mice were treated with thalidomide, the mRNA
expression of RELM-α, Arg-1, CD206 and YM-1 in the lungs was
significantly reduced. Thalidomide inhibited the expression of M2
macrophages (protumoral and pro-angiogenic TAMs) in vivo and
in vitro (61). Rolny
et al (48) treated
tumor-bearing (T241 fibrosarcoma and Panc02 tumor) mice with
clodronate liposomes to chemically deplete TAMs, which suggested
that M2-like TAMs induced vessel normalization. This study further
identified an inhibitory effect of thalidomide on alternative
activation of macrophages as a potential antitumor mechanism.
Furthermore, thalidomide probably mediates tumor vessel
normalization through the alternative activation of
macrophages.
NK cells are cells of the innate immune system that
arise from a common lymphoid progenitor cell. These cells are
characterized by a high cytolytic capacity against cancer cells. In
addition to their important role in immunosurveillance, NK cells
contribute to neovascularization. The expression of IFN inducible
protein (IP)-10, IFN-α, IL-12 and IL-18 was markedly upregulated
following the administration of thalidomide in order to modulate NK
cell activity in a murine model of melanoma (13). The upregulated cytokines are
associated with angiogenesis. The CD56(+) CD16(−) NK subset
[decidual NK (dNK) cells] in non-small cell lung cancer patients
was associated with VEGF, placental growth factor (PIGF), and
IL-8/CXCL8 production. This suggests that thalidomide probably
affects vessel remolding by regulating the secretion of
pro-angiogenic cytokines by NK cells (62). However, the precise role of
thalidomide and NK cells in tumor vascular normalization remains to
be defined. More studies have suggested that thalidomide may also
act as an inducer of T cell stimulatory signals, inducing T cell
proliferation in association with IFN-α and IL-2 production
(15,63). T cells can directly regulate the
level of MHC expression through IFN-γ secretion (64,65), but also improve the regulatory
functions of endothelial cells (ECs), namely, the regulation of
blood vessel formation and remodeling, blood flow, permselectivity,
blood fluidity and hemostasis (66). T cells not only affect the
cytoskeletal rearrangement in ECs and thrombi but are also
implicated in the regulation of the inhibitory molecule programmed
cell death-1 ligand (PD-L1) by T cell-derived cytokines. Thus,
thalidomide modulated immune cells to improve vascular
normalization through the production of cytokines.
Angiogenesis was demonstrated to be important in
supporting tumor growth, whereas infiltrating cancer cells
contributed to the tumor microenvironment through the secretion of
growth factors, cytokines and important molecules in the cancer
cells (67). Similarly, cytokines
were crucial in the development of tumors, the activation of the
immune response and angiogenesis in cancer. Thalidomide directly
altered the expression of cytokines but also regulated the
secretion of cytokines from immune cells indirectly. Through the
two pathways, thalidomide achieved antiangiogenesis and vascular
normalization in cancer. As modulators of tumor angiogenesis,
cytokines have been therapeutic targets in angiogenesis (68).
TAMs are capable of releasing a number of cytokines
(such as IL-1, IL-8 or TNF-α), which was the novel pivotal factor
of TAMs in facilitating tumor angiogenesis (69,70). Various mouse models have shown
that neutrophils are crucially involved in the process of tumor
angiogenesis. The depletion of neutrophils impaired angiogenesis in
mice inoculated with CXCL8 as well as in a transgenic RIPK1-Tag2
mouse model of pancreatic carcinoma (72). Mast cells driving neo-angiogenesis
have been documented to express Ang-1 (71). Tumor-associated dendritic cells
(DCs) can directly drive tumor angiogenesis through the release of
pro-angiogenic cytokines such as TNF-α, CXCL8 and osteopontin
(71). The above-mentioned
studies suggest that a complex relationship exists between immune
cells, tumor vessels and cytokines. Cytokines probably link
vascular normalization with immune cells. As thalidomide is capable
of altering the secretion and activity of various cytokines and
inducing an immune response, it may affect the interactions between
immune cells and the tumor vasculature through these cytokines.
Over the last few decades, thalidomide has undergone
a remarkable metamorphosis from a notorious drug inducing birth
defects into a highly effective therapy for treating leprosy and
multiple myeloma. Today, thalidomide is used to treat a wide range
of medical conditions successfully, which include cancer.
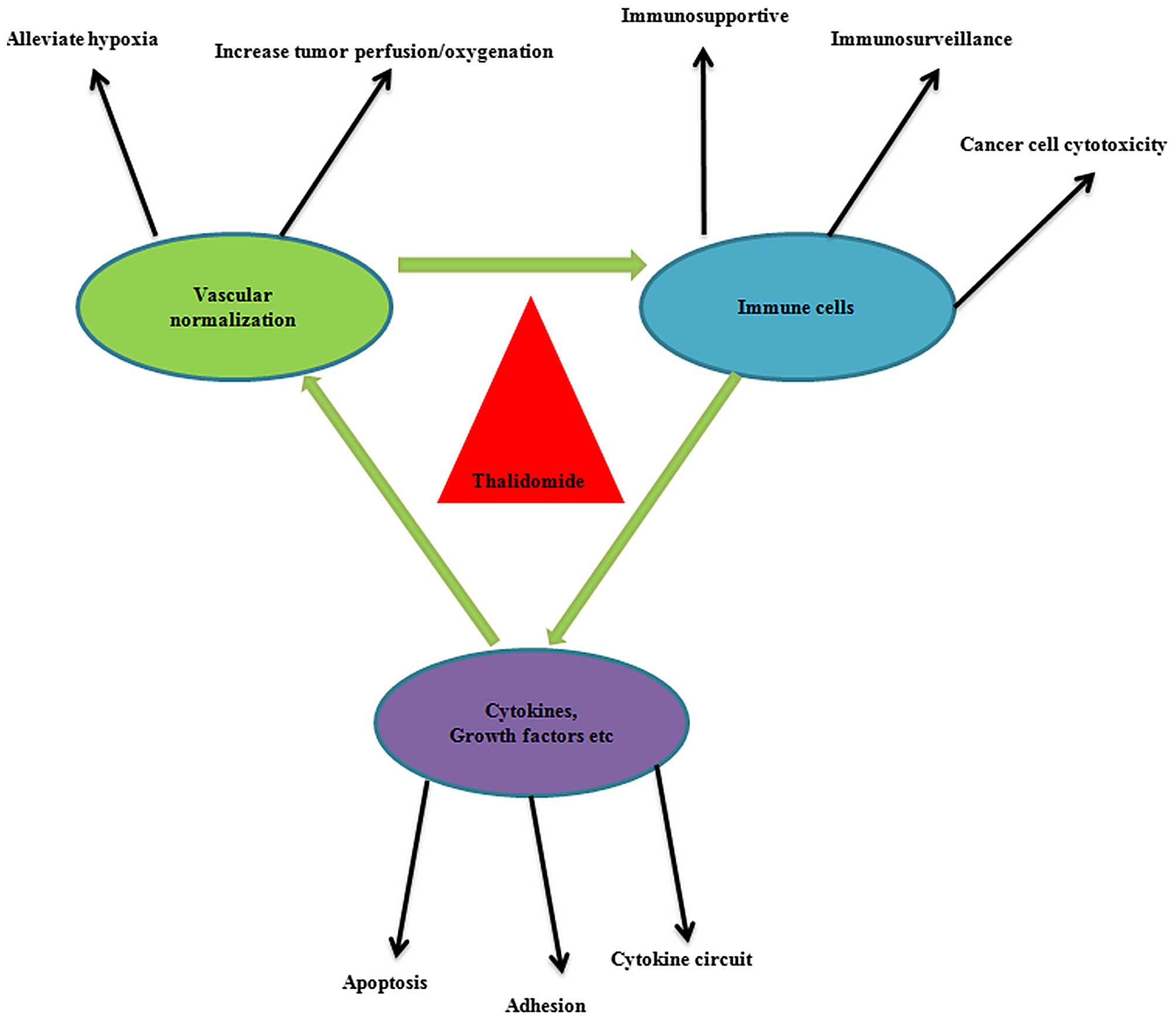
The mechanisms associated with antiangiogenesis and
the potent immunomodulatory effects of thalidomide obtained the
most support. Subsequently, the studies of the antiangiogenic
activity of thalidomide were guided into a novel direction by a
hypothesis regarding the vascular normalization of tumors. As
summarized in Fig. 3, the
potential normalization of tumor vessels by thalidomide may be an
effective strategy for reconditioning the tumor immune
microenvironment in order to produce anticancer effects. On the
other hand, it is recognized that the immune cell compartment
within the tumor is a major driver of angiogenesis and vascular
remodeling (72). Angiogenic
activity within a tumor is affected by many different types of
immune cell which release cytokines and angiogenic factors. Tumors
are highly heterogeneous. Despite the multiple mechanisms of action
responsible for the therapeutic effects of thalidomide discussed
previously, the precise mode of action of thalidomide remains to be
completely elucidated. Further studies are warranted in order to
clarify whether thalidomide needs to target different
pro-angiogenic factors to induce vascular normalization in
different tumors, at different times, as well as on
interconnections between the immune cells and tumor vasculature and
thereby elucidate the therapeutic effects of thalidomide in cancer.
Future studies regarding the association between the effects of
immune cells and vascular normalization action modes are also of
paramount significance. These studies may enable thalidomide to be
used in more effective ways.
|
1
|
Sheskin J: Thalidomide in the treatment of
lepra reactions. Clin Pharmacol Ther. 6:303–306. 1965. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
D'Amato RJ, Loughnan MS, Flynn E and
Folkman J: Thalidomide is an inhibitor of angiogenesis. Proc Natl
Acad Sci USA. 91:4082–4085. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Singhal S, Mehta J, Desikan R, Ayers D,
Roberson P, Eddlemon P, Munshi N, Anaissie E, Wilson C, Dhodapkar
M, et al: Antitumor activity of thalidomide in refractory multiple
myeloma. N Engl J Med. 341:1565–1571. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Drake MJ, Robson W, Mehta P, Schofield I,
Neal DE and Leung HY: An open-label phase II study of low-dose
thalidomide in androgen-independent prostate cancer. Br J Cancer.
88:822–827. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marx GM, Pavlakis N, McCowatt S, Boyle FM,
Levi JA, Bell DR, Cook R, Biggs M, Little N and Wheeler HR: Phase
II study of thalidomide in the treatment of recurrent glioblastoma
multiforme. J Neurooncol. 54:31–38. 2001. View Article : Google Scholar
|
|
6
|
Figg WD, Dahut W, Duray P, Hamilton M,
Tompkins A, Steinberg SM, Jones E, Premkumar A, Linehan WM, Floeter
MK, et al: A randomized phase II trial of thalidomide, an
angiogenesis inhibitor, in patients with androgen-independent
prostate cancer. Clin Cancer Res. 7:1888–1893. 2001.PubMed/NCBI
|
|
7
|
Eleutherakis-Papaiakovou V, Bamias A and
Dimopoulos MA: Thalidomide in cancer medicine. Ann Oncol.
8:1151–1160. 2004. View Article : Google Scholar
|
|
8
|
Kesari S, Schiff D, Henson JW, Muzikansky
A, Gigas DC, Doherty L, Batchelor TT, Longtine JA, Ligon KL, Weaver
S, et al: Phase II study of temozolomide, thalidomide, and
celecoxib for newly diagnosed glioblastoma in adults. Neuro Oncol.
10:300–308. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jain RK: Normalization of tumor
vasculature: an emerging concept in antiangiogenic therapy.
Science. 307:58–62. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ansiaux R, Baudelet C, Jordan BF, Beghein
N, Sonveaux P, De Wever J, Martinive P, Grégoire V, Feron O and
Gallez B: Thalidomide radiosensitizes tumors through early changes
in the tumor microenvironment. Clin Cancer Res. 11:743–750.
2005.PubMed/NCBI
|
|
11
|
El-Aarag BY, Kasai T, Zahran MA, Zakhary
NI, Shigehiro T, Sekhar SC, Agwa HS, Mizutani A, Murakami H, Kakuta
H and Seno M: In vitro anti-proliferative and anti-angiogenic
activities of thalidomide dithiocarbamate analogs. Int
Immunopharmacol. 21:283–292. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Davies FE, Raje N, Hideshima T, Lentzsch
S, Young G, Tai YT, Lin B, Podar K, Gupta D, Chauhan D, et al:
Thalidomide and immunomodulatory derivatives augment natural killer
cell cytotoxicity in multiple myeloma. Blood. 98:210–216. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kawamata A, Ito D, Odani T, Isobe T, Iwase
M, Hatori M and Nagumo M: Thalidomide suppresses melanoma growth by
activating natural killer cells in mice. Oncol Rep. 16:1231–1236.
2006.PubMed/NCBI
|
|
14
|
von Moos R, Stolz R, Cerny T and Gillessen
S: Thalidomide: from tragedy to promise. Swiss Med Wkly. 133:77–87.
2003.PubMed/NCBI
|
|
15
|
Corral LG, Haslett PA, Muller GW, Chen R,
Wong LM, Ocampo CJ, Patterson RT, Stirling DI and Kaplan G:
Differential cytokine modulation and T cell activation by two
distinct classes of thalidomide analogues that are potent
inhibitors of TNF-alpha. J Immunol. 163:380–386. 1999.PubMed/NCBI
|
|
16
|
Corral LG and Kaplan G: Immunomodulation
by thalidomide and thalidomide analogues. Ann Rheum Dis. 58(Suppl
1): I107–I113. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Haslett PA, Klausner JD, Makonkawkeyoon S,
Moreira A, Metatratip P, Boyle B, Kunachiwa W, Maneekarn N,
Vongchan P, Corral LG, et al: Thalidomide stimulates T cell
responses and interleukin 12 production in HIV-infected patients.
AIDS Res Hum Retroviruses. 15:1169–1179. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Muller GW, Chen R, Huang SY, Corral LG,
Wong LM, Patterson RT, Chen Y, Kaplan G and Stirling DI:
Amino-substituted thalidomide analogs: potent inhibitors of
TNF-alpha production. Bioorg Med Chem Lett. 9:1625–1630. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Joussen AM, Germann T and Kirchhof B:
Effect of thalidomide and structurally related compounds on corneal
angiogenesis is comparable to their teratological potency. Graefes
Arch Clin Exp Ophthalmol. 237:952–961. 1999. View Article : Google Scholar
|
|
20
|
Therapontos C, Erskine L, Gardner ER, Figg
WD and Vargesson N: Thalidomide induces limb defects by preventing
angiogenic outgrowth during early limb formation. Proc Natl Acad
Sci USA. 106:8573–8578. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ito T, Ando H, Suzuki T, Ogura T, Hotta K,
Imamura Y, Yamaguchi Y and Handa H: Identification of a primary
target of thalidomide teratogenicity. Science. 327:1345–1350. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yabu T, Tomimoto H, Taguchi Y, Yamaoka S,
Igarashi Y and Okazaki T: Thalidomide-induced antiangiogenic action
is mediated by ceramide through depletion of VEGF receptors, and is
antagonized by sphingosine-1-phosphate. Blood. 106:125–134. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Verheul HM, Panigrahy D, Yuan J and
D'Amato RJ: Combination oral antiangiogenic therapy with
thalidomide and sulindac inhibits tumour growth in rabbits. Br J
Cancer. 79:114–118. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bertolini F, Mingrone W, Alietti A,
Ferrucci PF, Cocorocchio E, Peccatori F, Cinieri S, Mancuso P,
Corsini C, et al: Thalidomide in multiple myeloma, myelodysplastic
syndromes and histiocytosis. Analysis of clinical results and of
surrogate angiogenesis markers. Ann Oncol. 12:987–990. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li X, Liu X, Wang J, Wang Z, Jiang W, Reed
E, Zhang Y, Liu Y and Li QQ: Effects of thalidomide on the
expression of angiogenesis growth factors in human A549 lung
adenocarcinoma cells. Int J Mol Med. 11:785–790. 2003.PubMed/NCBI
|
|
26
|
Vasvari GP, Dyckhoff G, Kashfi F, Lemke B,
Lohr J, Helmke BM, Schirrmacher V, Plinkert PK, Beckhove P and
Herold-Mende CC: Combination of thalidomide and cisplatin in an
head and neck squamous cell carcinomas model results in an enhanced
antiangiogenic activity in vitro and in vivo. Int J Cancer.
121:1697–1704. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stephens TD, Bunde CJ and Fillmore BJ:
Mechanism of action in thalidomide teratogenesis. Biochem
Pharmacol. 59:1489–1499. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stephens TD and Fillmore BJ: Hypothesis:
thalidomide embryopathy-proposed mechanism of action. Teratology.
61:189–195. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fujita J, Mestre JR, Zeldis JB,
Subbaramaiah K and Dannenberg AJ: Thalidomide and its analogues
inhibit lipopolysaccharide-mediated linduction of cyclooxygenase-2.
Clin Cancer Res. 7:3349–3355. 2001.PubMed/NCBI
|
|
30
|
Chang SH, Liu CH, Conway R, Han DK,
Nithipatikom K, Trifan OC, Lane TF and Hla T: Role of prostaglandin
E2-dependent angiogenic switch in cyclooxygenase 2-induced breast
cancer progression. Proc Natl Acad Sci USA. 101:591–596. 2004.
View Article : Google Scholar :
|
|
31
|
Yamada M, Kawai M, Kawai Y and Mashima Y:
The effect of selective cyclooxygenase-2 inhibitor on corneal
angiogenesis in the rat. Curr Eye Res. 19:300–304. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vargesson N: Thalidomide-induced limb
defects: resolving a 50-year-old puzzle. BioEssays. 31:1327–1336.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vargesson N: Thalidomide embryopathy: An
enigmatic challenge. ISRN Dev Biol. 241016:2013 View Article : Google Scholar
|
|
34
|
Feng Q, Tan HH, Ge ZZ, Gao YJ, Chen HM and
Xiao SD: Thalidomide-induced angiopoietin 2, Notch1 and Dll4
downregulation under hypoxic condition in tissues with
gastrointestinal vascular malformation and human umbilical vein
endothelial cells. J Dig Dis. 15:85–95. 2014. View Article : Google Scholar
|
|
35
|
Li Y, Fu S, Chen H, Feng Q, Gao Y, Xue H,
Ge Z, Fang J and Xiao S: Inhibition of endothelial Slit2/Robo1
signaling by thalidomide restrains angiogenesis by blocking the
PI3K/Akt pathway. Dig Dis Sci. 59:2958–2966. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Segers J, Di Fazio V, Ansiaux R, Martinive
P, Feron O, Wallemacq P and Gallez B: Potentiation of
cyclophosphamide chemotherapy using the anti-angiogenic drug
thalidomide: importance of optimal scheduling to exploit the
'normalization' window of the tumor vasculature. Cancer Lett.
244:129–135. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lebrin F, Srun S, Raymond K, Martin S, van
den Brink S, Freitas C, Bréant C, Mathivet T, Larrivée B, Thomas
JL, et al: Thalidomide stimulates vessel maturation and reduces
epistaxis in individuals with hereditary hemorrhagic
telangiectasia. Nat Med. 16:420–428. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Goel S, Duda DG, Xu L, Munn LL, Boucher Y,
Fukumura D and Jain RK: Normalization of the vasculature for
treatment of cancer and other diseases. Physiol Rev. 91:1071–1121.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Floros T and Tarhini AA: Anticancer
cytokines: biology and clinical effects of Interferon-α2,
Interleukin (IL)-2, IL-15, IL-21, and IL-12. Semin Oncol.
42:539–548. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mitsiades N, Mitsiades CS, Poulaki V,
Chauhan D, Richardson PG, Hideshima T, Munshi NC, Treon SP and
Anderson KC: Apoptotic signaling induced by immunomodulatory
thalidomide analogs in human multiple myeloma cells: therapeutic
implications. Blood. 99:4525–4530. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Marriott JB, Clarke IA, Czajka A, Dredge
K, Childs K, Man HW, Schafer P, Govinda S, Muller GW, Stirling DI
and Dalgleish AG: A novel subclass of thalidomide analogue with
anti-solid tumor activity in which caspase-dependent apoptosis is
associated with altered expression of bcl-2 family proteins. Cancer
Res. 63:593–599. 2003.PubMed/NCBI
|
|
42
|
Schuster SR, Kortuem KM, Zhu YX, Braggio
E, Shi CX, Bruins LA, Schmidt JE, Ahmann G, Kumar S, Rajkumar SV,
et al: The clinical significance of cereblon expression in multiple
myeloma. Leuk Res. 38:23–28. 2014. View Article : Google Scholar :
|
|
43
|
Chung AS, Lee J and Ferrara N: Targeting
the tumour vasculature: insights from physiological angiogenesis.
Nat Rev Cancer. 10:505–514. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sitkovsky MV, Kjaergaard J, Lukashev D and
Ohta A: Hypoxia-adenosinergic immunosuppression: tumor protection
by T regulatory cells and cancerous tissue hypoxia. Clin Cancer
Res. 14:5947–5952. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Palazon A, Aragones J, Morales-Kastresana
A, de Landazuri MO and Melero I: Molecular pathways: hypoxia
response in immune cells fighting or promoting cancer. Clin Cancer
Res. 18:1207–1213. 2012. View Article : Google Scholar
|
|
46
|
Huang Y, Goel S, Duda DG, Fukumura D and
Jain RK: Vascular normalization as an emerging strategy to enhance
cancer immunotherapy. Cancer Res. 73:2943–2948. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hamzah J, Jugold M, Kiessling F, Rigby P,
Manzur M, Marti HH, Rabie T, Kaden S, Gröne HJ, Hämmerling GJ, et
al: Vascular normalization in Rgs5-deficient tumours promotes
immune destruction. Nature. 453:410–414. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Rolny C, Mazzone M, Tugues S, Laoui D,
Johansson I, Coulon C, Squadrito ML, Segura I, Li X, Knevels E, et
al: HRG inhibits tumor growth and metastasis by inducing macrophage
polarization and vessel normalization through downregulation of
PlGF. Cancer Cell. 19:31–44. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Curiel TJ, Coukos G, Zou L, Alvarez X,
Cheng P, Mottram P, Evdemon-Hogan M, Conejo-Garcia JR, Zhang L,
Burow M, et al: Specific recruitment of regulatory T cells in
ovarian carcinoma fosters immune privilege and predicts reduced
survival. Nat Med. 10:942–949. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
50
|
Galon J, Costes A, Sanchez-Cabo F,
Kirilovsky A, Mlecnik B, Lagorce-Pagès C, Tosolini M, Camus M,
Berger A, Wind P, et al: Type, density, and location of immune
cells within human colorectal tumors predict clinical outcome.
Science. 313:1960–1964. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tan TT and Coussens LM: Humoral immunity,
inflammation and cancer. Curr Opin Immunol. 19:209–216. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Galustian C, Meyer B, Labarthe MC, Dredge
K, Klaschka D, Henry J, Todryk S, Chen R, Muller G, Stirling D, et
al: The anti-cancer agents lenalidomide and pomalidomide inhibit
the proliferation and function of T regulatory cells. Cancer
Immunol Immunother. 58:1033–1045. 2009. View Article : Google Scholar
|
|
53
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
de Visser KE and Coussens LM: The
inflammatory tumor microenvironment and its impact on cancer
development. Contrib Microbiol. 13:118–137. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lin EY, Li JF, Gnatovskiy L, Deng Y, Zhu
L, Grzesik DA, Qian H, Xue XN and Pollard JW: Macrophages regulate
the angiogenic switch in a mouse model of breast cancer. Cancer
Res. 66:11238–11246. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Murdoch C, Giannoudis A and Lewis CE:
Mechanisms regulating the recruitment of macrophages into hypoxic
areas of tumors and other ischemic tissues. Blood. 104:2224–2234.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Murdoch C, Muthana M, Coffelt SB and Lewis
CE: The role of myeloid cells in the promotion of tumour
angiogenesis. Nat Rev Cancer. 8:618–631. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Burke B, Tang N, Corke KP, Tazzyman D,
Ameri K, Wells M and Lewis CE: Expression of HIF-1alpha by human
macrophages: implications for the use of macrophages in
hypoxia-regulated cancer gene therapy. J Pathol. 196:204–212. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lewis CE and Pollard JW: Distinct role of
macrophages in different tumor microenvironments. Cancer Res.
66:605–612. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Murdoch C and Lewis CE: Macrophage
migration and gene expression in response to tumor hypoxia. Int J
Cancer. 117:701–708. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Lee HS, Kwon HS, Park DE, Woo YD, Kim HY,
Kim HR, Cho SH, Min KU, Kang HR and Chang YS: Thalidomide inhibits
alternative activation of macrophages in vivo and in vitro: a
potential mechanism of anti-asthmatic effect of thalidomide. PLoS
One. 10:e01230942015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Bruno A, Focaccetti C, Pagani A,
Imperatori AS, Spagnoletti M, Rotolo N, Cantelmo AR, Franzi F,
Capella C, Ferlazzo G, et al: The proangiogenic phenotype of
natural killer cells in patients with non-small cell lung cancer.
Neoplasia. 15:133–142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Haslett PA, Corral LG, Albert M and Kaplan
G: Thalidomide costimulates primary human T lymphocytes,
preferentially inducing proliferation, cytokine production, and
cytotoxic responses in the CD8+ subset. J Exp Med. 187:1885–1892.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Pober JS, Gimbrone MA Jr, Cotran RS, Reiss
CS, Burakoff SJ, Fiers W and Ault KA: Ia expression by vascular
endothelium is inducible by activated T cells and by human gamma
interferon. J Exp Med. 157:1339–1353. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Pober JS, Gimbrone MA Jr, Lapierre LA,
Mendrick DL, Fiers W, Rothlein R and Springer TA: Overlapping
patterns of activation of human endothelial cells by interleukin 1,
tumor necrosis factor, and immune interferon. J Immunol.
137:1893–1896. 1986.PubMed/NCBI
|
|
66
|
Choi J, Enis DR, Koh KP, Shiao SL and
Pober JS: T lymphocyte-endothelial cell interactions. Annu Rev
Immunol. 22:683–709. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Trédan O, Lacroix-Triki M, Guiu S,
Mouret-Reynier MA, Barrière J, Bidard FC, Braccini AL, Mir O,
Villanueva C and Barthélémy P: Angiogenesis and tumor
microenvironment: bevacizumab in the breast cancer model. Target
Oncol. 10:189–198. 2015. View Article : Google Scholar
|
|
68
|
Rivas-Fuentes S, Salgado-Aguayo A, Pertuz
Belloso S, Gorocica Rosete P, Alvarado-Vásquez N and Aquino-Jarquin
G: Role of chemokines in non-small cell lung cancer: angiogenesis
and inflammation. J Cancer. 6:938–952. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Dirkx AE, Oude Egbrink MG, Wagstaff J and
Griffioen AW: Monocyte/macrophage infiltration in tumors:
modulators of angiogenesis. J Leukoc Biol. 80:1183–1196. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Fischer C, Jonckx B, Mazzone M, Zacchigna
S, Loges S, Pattarini L, Chorianopoulos E, Liesenborghs L, Koch M,
De Mol M, et al: Anti-PlGF inhibits growth of
VEGF(R)-inhibitor-resistant tumors without affecting healthy
vessels. Cell. 131:463–475. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Nakayama T, Yao L and Tosato G: Mast
cell-derived angiopoietin-1 plays a critical role in the growth of
plasma cell tumors. J Clin Invest. 114:1317–1325. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Stockmann C, Schadendorf D, Klose R and
Helfrich I: The impact of the immune system on tumor: angiogenesis
and vascular remodeling. Front Oncol. 4:692014. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Jain RK: Antiangiogenesis strategies
revisited: from starving tumors to alleviating hypoxia. Cancer
Cell. 26:605–622. 2014. View Article : Google Scholar : PubMed/NCBI
|