Introduction
Over the last two decades, neural stem cells (NSCs)
have become a major topic of interest from basic research to
translational experiments for the development of therapies for a
range of neurological disorders. NSCs have two defining
characteristics: Self-renewal and multipotentiality (1). Their capacity to propagate in
culture over several passages and differentiate into neuronal and
glial cell types renders them attractive as a model of neurogenesis
and neural cells, and as a therapeutic tool for treating
neurological disease. Previous studies have used a wide range of
NSCs from adult and fetal origins, but predominantly from rodent
models (2). However, in the case
of human NSCs (hNSCs), several constraints, including the limited
donor availability to derive fetal and adult NSCs, the low rate of
proliferation and the difficulty of long-term in vitro
expansion, mean it is not possible to produce the required cell
numbers while maintaining a stable phenotype across passages.
Therefore, it is important to develop in vitro expandable
cell sources for providing suitable hNSCs in sufficiently large
numbers.
The life span of hNSCs in vitro can be
improved by optimizing culture conditions (3) or via immortalization using the myc
transcription factor (4) and
maintaining a stable phenotype. Stable hNSC lines, including
ReNcell CX cells immortalized using c-myc and VM cells immortalized
with v-myc, are widely used in investigations in a variety of
neurological fields (5). ReNcell
lines have been shown to propagate perpetually in culture and
exhibit properties of hNSCs, including expression of NESTIN in an
undifferentiated state and differentiation into specific cell
types, including neuronal and glial cells, following deprivation of
growth factors in culture medium (6). It was previously reported that
ReNcell lines were used in disease modeling for Alzheimer's disease
(AD) (7,8); a three-dimensional culture model of
ReNcell VM cells with mutations in amyloid precursor protein and
presenilin 1 was able to recapitulate AD pathologies. However,
there are practical limitations to using immortalized hNSC lines
for clinical applications, including a higher risk of aberrant
growth, which may be circumvented by subjecting these cells to
extensive characteristic analyses.
Human embryonic stem cells (hESCs), used as
pluripotent cells, provide an unlimited and renewable source of
hNSCs. Several protocols have been developed to differentiate hESCs
into expandable hNSC populations, and to derive potentially
functional neurons and glial cells in a controlled manner (6,9,10).
Due to the high differentiation potential, in vitro
expandable NSCs derived from hESCs are one of the most accessible
models for human developmental neurobiology, although certain
ethical issues remain unresolved (11). hESC-derived NSCs can serve as an
in vitro model for the examination of human neural
development as newly derived NSCs are similar to embryonic
neuroepithelial cells. In addition, in long-term culture, these
cells are more likely to develop features similar to those of fetal
and adult NSCs (12). The hESCs
used in the production of hNSCs have the advantage of being capable
of propagation over multiple passages, offering a virtually
unlimited supply of hNSCs (13).
The present study aimed to compare and characterize
two representative hNSC sources to provide a well-defined in
vitro model comparable to human neuronal physiology for various
research applications. This involved examining whole-genome
expression using microarrays in ReNcell and hESC-derived NSCs, and
assessing their neuronal differentiation potential. To the best of
our knowledge, this is the first report to provide a comprehensive
analysis of the gene expression of ReNcell and hESC-derived NSCs.
The results extend the gene expression network for neural
differentiation and reveal common principles of transcriptional
regulation underlying the differentiation of hESCs into NSCs.
Materials and methods
hESC culture
H9 hESCs (cat. no. WA09; WiCell Research Institute,
Madison, WI, USA) were maintained on Matrigel (BD Biosciences, San
Diego, CA, USA) in mTeSR1 (StemCell Technologies, Vancouver, BC,
Canada) as previously described (14,15).
Differentiation of hESCs into hNSCs
The hNSCs were differentiated through the formation
of human neuroectodermal spheres (hNESs) as previously reported
with minor modifications (2,16).
The H9 hESCs (cat. no. WA09; WiCell Research Institute) were
maintained on Matrigel (BD Biosciences) in mTeSR1 (StemCell
Technologies) as previously described (14). Human embryoid bodies (hEBs) were
generated by culturing hESCs in hEB medium consisting of knockout
DMEM supplemented with 10% knockout serum replacement, 1%
non-essential amino acids, 1 mM L-glutamine (all from Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) and 0.1 mM
β-mercaptoethanol (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
on non-coated Petri dishes. The resulting hEBs were then cultured
in NES/NSC medium consisting of DMEM/F12, 1X N2/B27 (both from
Invitrogen; Thermo Fisher Scientific, Inc.), 20 ng/ml basic
fibroblast growth factor (bFGF; R&D Systems, Inc., Minneapolis,
MN, USA), 20 ng/ml epidermal growth factor (EGF) and 10 ng/ml
leukemia inhibitory factor (both from PeproTech, Inc., Rocky Hill,
NJ, USA). The hNESs were sub-cultured every week using a Mcllwain
tissue chopper (Mickle Engineering, Surrey, UK), and the medium was
replaced every 2 days. The hNESs were passaged at least five times
without disturbing the formation of neural rosettes. For terminal
differentiation, each hNES was allowed to attach to a
Matrigel-coated coverslip and was maintained without growth factors
for 2 weeks, as previously described (17,18). To count the total number of cells
within each hNES, the hNESs were dissociated into single-cell
suspensions with 0.1% trypsin-EDTA (Invitrogen; Thermo Fisher
Scientific, Inc.) for 3 min. Live cell numbers were counted using
trypan blue (Invitrogen; Thermo Fisher Scientific, Inc.) exclusion
under an Olympus fluorescence microscope (IX51; Olympus Corp.,
Tokyo, Japan).
ReNcell CX cell culture
ReNcell CX cells derived from the cortical region of
human fetal brain tissue (cat. no. SCC007; EMD Millipore, Temecula,
CA, USA) were cultured according to the manufacturer's protocol.
The ReNcell CX cells were maintained in ReNcell NSC maintenance
medium supplemented with 20 ng/ml EGF and 20 ng/ml bFGF (all from
EMD Millipore) on laminin-coated tissue culture dishes (BD
Biosciences). The culture medium was replaced every 2 days. For
terminal differentiation, the ReNcell CX cells were cultured for 5
days without growth factors, as previously described (19).
Immunocytochemistry
Immunocytochemistry was performed as previously
described (20). In brief, the
cells were fixed in 4% formaldehyde and then permeabilized with PBS
containing 0.1% Triton X-100. Following blocking with 3% bovine
serum albumin (Sigma-Aldrich; Merck KGaA), the cells were incubated
at 4°C overnight with anti-neuron-specific class III β-tubulin
(TUJ1; 1:500; cat. no. PRB-435P; Covance, Inc., Princeton, NJ,
USA), anti-NESTIN (1:100; cat. no. MAB5326),
anti-microtubule-associated protein 2 (MAP2; 1:500; cat. no.
MAB3418), anti-glial fibrillary acidic protein (GFAP; 1:200; cat.
no. MAB3402) and anti-Ki67 (1:500; cat. no. AB9260; Chemicon) (all
from EMD Millipore), followed by incubation with Alexa Fluor
488-conjugated anti-mouse IgG (1:1,000; cat. no. A21202), Alexa
Fluor 594-conjugated anti-mouse IgG (1:1,000; cat. no. A21203),
Alexa Fluor 488-conjugated anti-rabbit IgG (1:1,000; cat. no.
A21441) or Alexa Fluor 594-conjugated anti-rabbit IgG (1:1,000;
cat. no. A21442) (all from Molecular Probes, Eugene, OR, USA) as
secondary antibodies for 1 h at room temperature. DAPI (1 mg/ml;
Invitrogen; Thermo Fisher Scientific, Inc.) was added to visualize
the nuclei. The slides were examined using an Axiovert 200M
microscope (Carl Zeiss AG, Gottingen, Germany).
Semi-quantitative reverse
transcription-polymerase chain reaction (RT-PCR) analysis
Total RNA was extracted from cells with an RNeasy
kit (Qiagen, Inc. Hilden, Germany) and reverse transcribed using a
Superscript IV First-Strand Synthesis System kit (Invitrogen;
Thermo Fisher Scientific, Inc.) as previously described (21). The resulting cDNA was diluted 1:10
with deionized water, and 1 µl of the diluted cDNA was added
to Accupower™ PCR PreMix (Bioneer Corp., Daejeon, Korea), 10 pmol/l
of specific primers and deionized water to a final volume of 20
µl. The RT-PCR analysis was performed under the following
conditions: 5 min at 95°C; 30–40 cycles of 30 sec at 95°C, 30 sec
at 60°C, 30 sec at 72°C, and 5 min extension at 72°C. GAPDH was
used as an internal control. The relative expression of target
genes was determined using the 2−ΔΔCq method (22). The primers used in this study are
listed in Table I.
 | Table IList of primers used in the present
study. |
Table I
List of primers used in the present
study.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| OCT4 |
GAGAAGGATGTGGTCCGAGTGTG C |
AGAGGAAAGGACACTGGTCCC |
| SOX2 |
AGAACCCCAAGATGCACAAC |
ATGTAGGTCTGCGAGCTGGT |
| SOX1 | GGGAAAACGGGCAAAATAAT
CC |
ATCTGGGCTTCAAGTGTT |
| SOX3 |
GACGCCTTGTTTAGCTTTGC |
TTCTCCCATTCACTCCTTGG |
| MSI1 |
ACCCCCACATTCTCTCACTG |
AAACCCAAAACACGAACAGC |
| TUJ1 |
ACCTCAACCACCTGGTATCG |
GGGTACCACTCCACGAAGTA |
| NESTIN |
CAGGAGAAACAGGGCCTACA |
TGGGAGCAAAGATCCAAGAC |
| GAPDH |
GAAGGTGAAGGTCGGAGTC |
GAAGATGGTGATGGGATTTC |
Microarray analysis
The microarray experiments were performed using the
Low RNA input linear amplification kit, cRNA cleanup module and
one-color (Cy3) Whole Human Genome Microarray 4X44K, according to
the manufacturer's protocol (Agilent Technologies, Inc., Santa
Clara, CA, USA) as previously described (23). The raw data was normalized using
global scale normalization and processed using GeneSpring software
version 11.0 (Agilent Technologies, Inc.,). Heatmap and
hierarchical clustering of genes was generated using MeV v. 4.9.0
software (http://www.tm4.org). Gene functions were
annotated using the GeneCard database (http://www.genecards.org/). The principal component
analysis (PCA) was performed with GeneSpring software. Biological
processes and protein classes were described using Protein Analysis
Through Evolutionary Relationships (PANTHER; http://www.pantherdb.org/). Kyoto Encyclopaedia of
Genes and Genomes (KEGG) analysis and functional annotation
clustering were performed using David Bioinformatics Resources 6.8
with the Database for Annotation, Visualization and Integrated
Discovery (DAVID; http://david.abcc.ncifcrf.gov).
Results and discussion
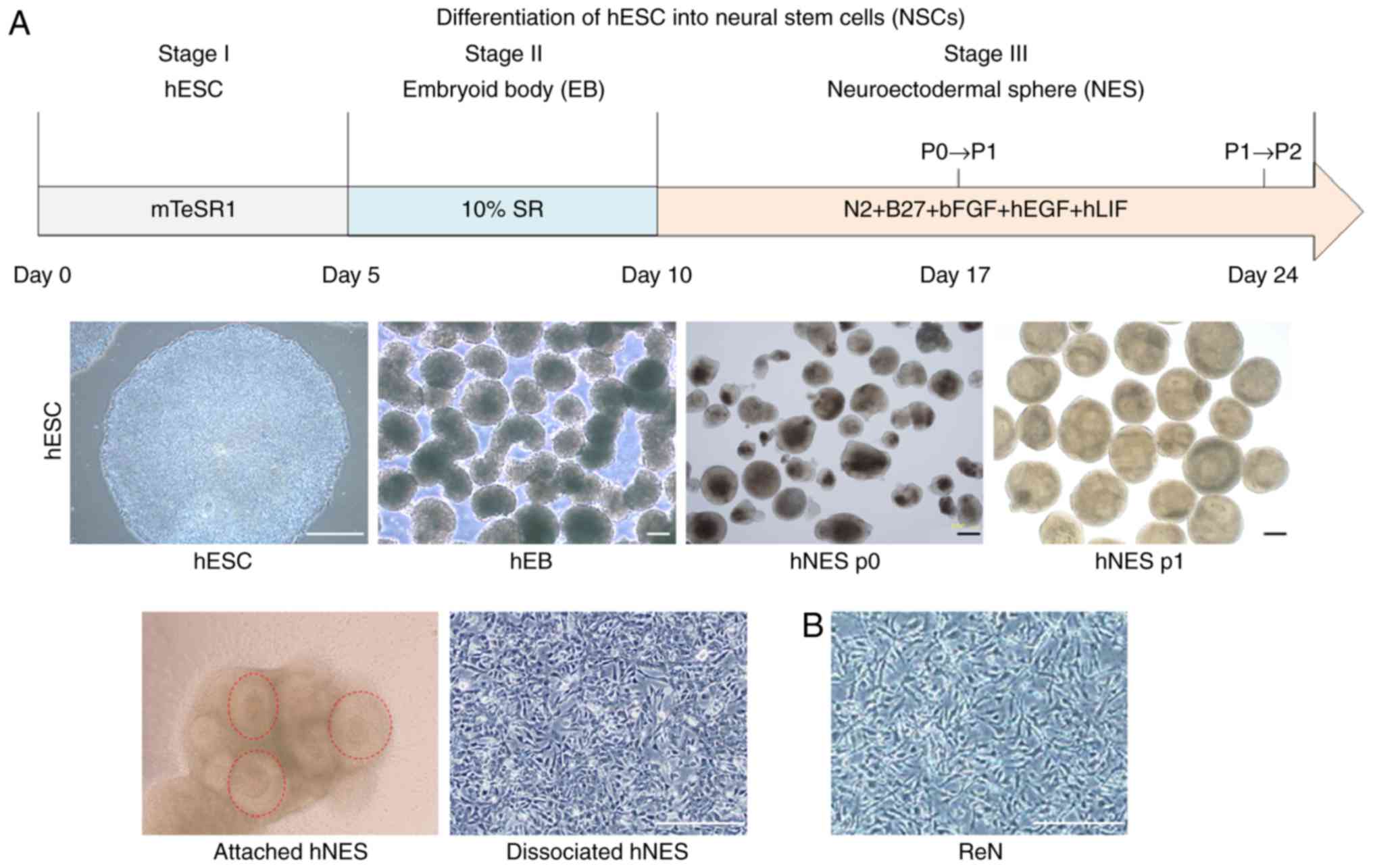
Generation of NSCs derived from hESCs via
hNES formation
In the present study, H9 hESCs were differentiated
into hNSCs based on the previously described hNES formation method
(17). The aggregates of hESCs
were cultured in hEB medium for 5 days, followed by transfer into
NES/NSC medium to drive neuronal fate commitment and promote
neuronal differentiation (Fig.
1A). During differentiation, following the first subculture,
hNESs containing neural rosette structures, a key structure
representing NSCs, appeared and retained the potential to form
neural rosette structures (Fig.
1A; red dotted circle). The hNESs were dissociated into single
NSCs by trypsin digestion and were cultured as adherent monolayers.
As reported in our previous study (1,17,24), the hNESs generated using this
method are characterized as NSCs as they have the potential to
differentiate into neuronal and glial cell types, and can be
serially passaged to form new hNESs.
Immortalized hNSC lines are in increasing demand due
to the inherent limitations of primary hNSCs, including limited
availability, poor expandability and associated ethical issues
(5). The ReNcell CX cell line,
which is a commercially available immortalized fetal cortical NSC
line, was used as a reference hNSC type in the present study. Under
normal growth conditions, ReNcell CX cells exhibited immature
neural morphology, similar to that of monolayer-cultured hNSCs
(Fig. 1). The ReNcell CX cells
grew rapidly as a monolayer on laminin, with a doubling time of ~24
h due to the c-MYC-based immortalization. Therefore, there are
safety concerns, including the risk that oncogenic c-MYC may render
this hNSC line tumorigenic following transplantation (19,25).
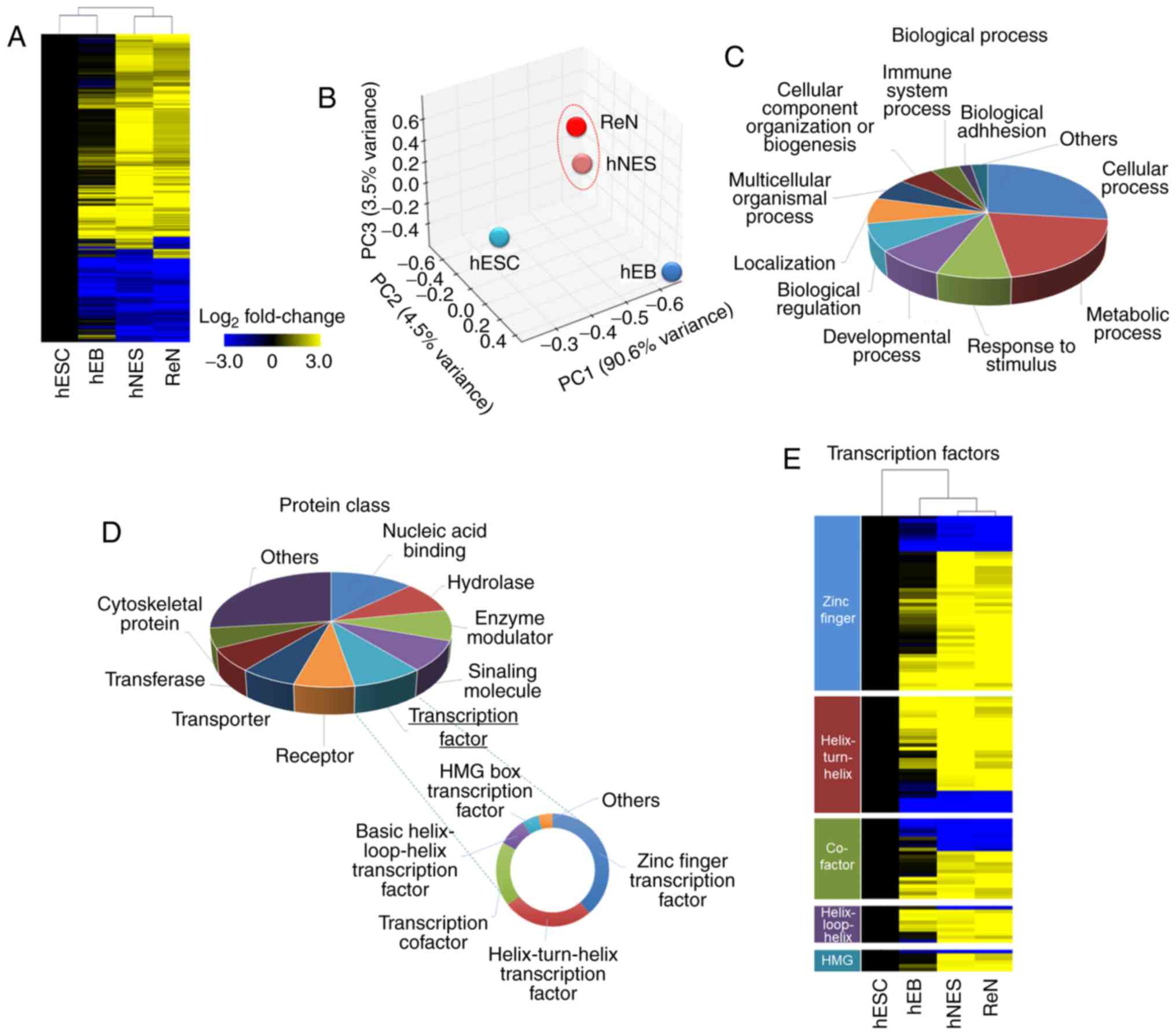
Comparative gene expression analysis of
hNESs derived from hESCs and ReNcell CX cells
To compare hNESs derived from hESCs and ReNcell CX
cells for use as an hNSC model, and examine the mechanisms
underlying lineage commitment in NSCs, microarray analyses were
performed in undifferentiated hESCs, hEBs (intermediate cells in
hNSC differentiation), hESC-derived hNESs and ReNcell CX cells. A
heatmap showing the hierarchical clustering results from the
whole-genome expression profiles indicated that differentiated
hNESs preferentially clustered with ReNcell CX cells (Fig. 2A). The principal component
analysis (PCA) also confirmed that hNESs and ReNcell CX cells were
distinctly separated from undifferentiated hESCs and hEBs (Fig. 2B). Accordingly, only ~12.4% of all
the genes were differentially expressed, with a fold-change
threshold of 2.0 between the hNESs and ReNcell CX cells. These data
indicated that the global transcription of differentiated hNESs is
similar to that of ReNcell CX cells.
To obtain the overall profile regarding common
aspects of hNSC identity, a total of 1,711 commonly upregulated and
856 commonly downregulated genes between hNESs and ReNcell CX cells
with a fold-change of >5 were identified as the hNSC-specific
transcriptome and analyzed using the PANTHER classification system.
The top biological process term was cellular process (26.9%)
(Fig. 2C). Other major processes
corresponding to these hNSC-related genes included metabolic
process (20.4%), response to stimulus (8.7%), developmental process
(8.3%), biological regulation (7.6%), localization (7.3%),
multicellular organismal process (6.3%) and cellular component
organization or biogenesis (5.6%), as shown in Fig. 2C. In addition, the predominant
protein class was nucleic acid binding (12.9%), followed by
hydrolase (8.9%), enzyme modulator (8.6%), signaling molecule
(8.5%), transcription factor (8.3%), receptor (7.0%), transporter
(6.6%), transferase (6.5%) and cytoskeletal protein (5.6%), as
shown in Fig. 2D. A significant
number of genes were identified as transcription factors, which are
known to have prominent roles in lineage specification and
developmental processes (26).
Therefore, the contribution of these transcription factors to the
hNSC identity was dissected. The important enriched transcription
factor categories were zinc finger transcription factor (38.4%),
helix-turn-helix transcription factor (25.6%), transcription
cofactor (18.4%), basic helix-loop-helix transcription factor
(8.0%), HMG box transcription factor (4.8%), and nuclear hormone
receptor binding (4.0%), as shown in Fig. 2D. The expression of several
transcription factors from the microarray data were analyzed
further, and the transcription factor expression levels were
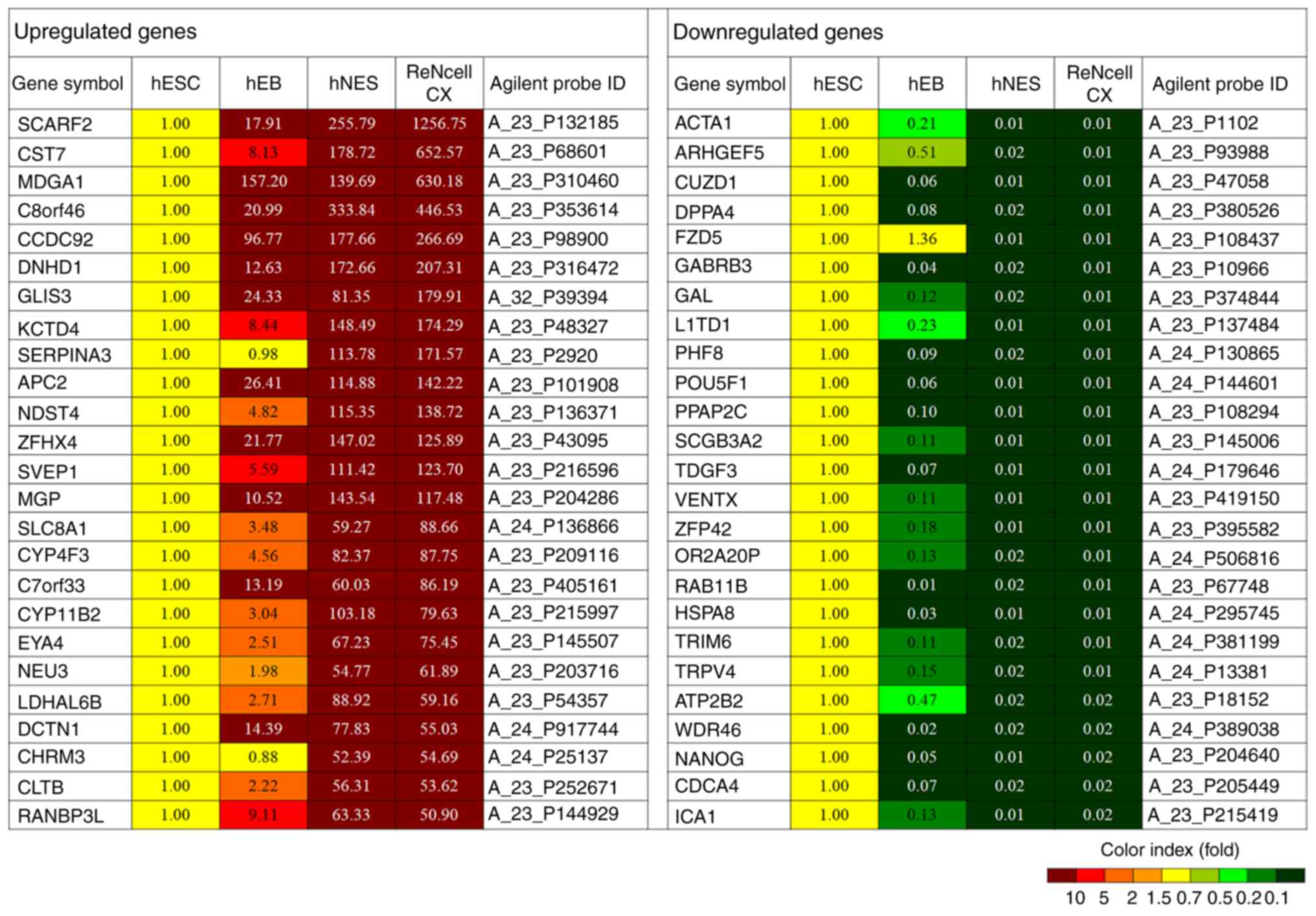
similar between the hNESs and ReNcell CX cells (Fig. 2E). Representative genes are shown
in Fig. 3.
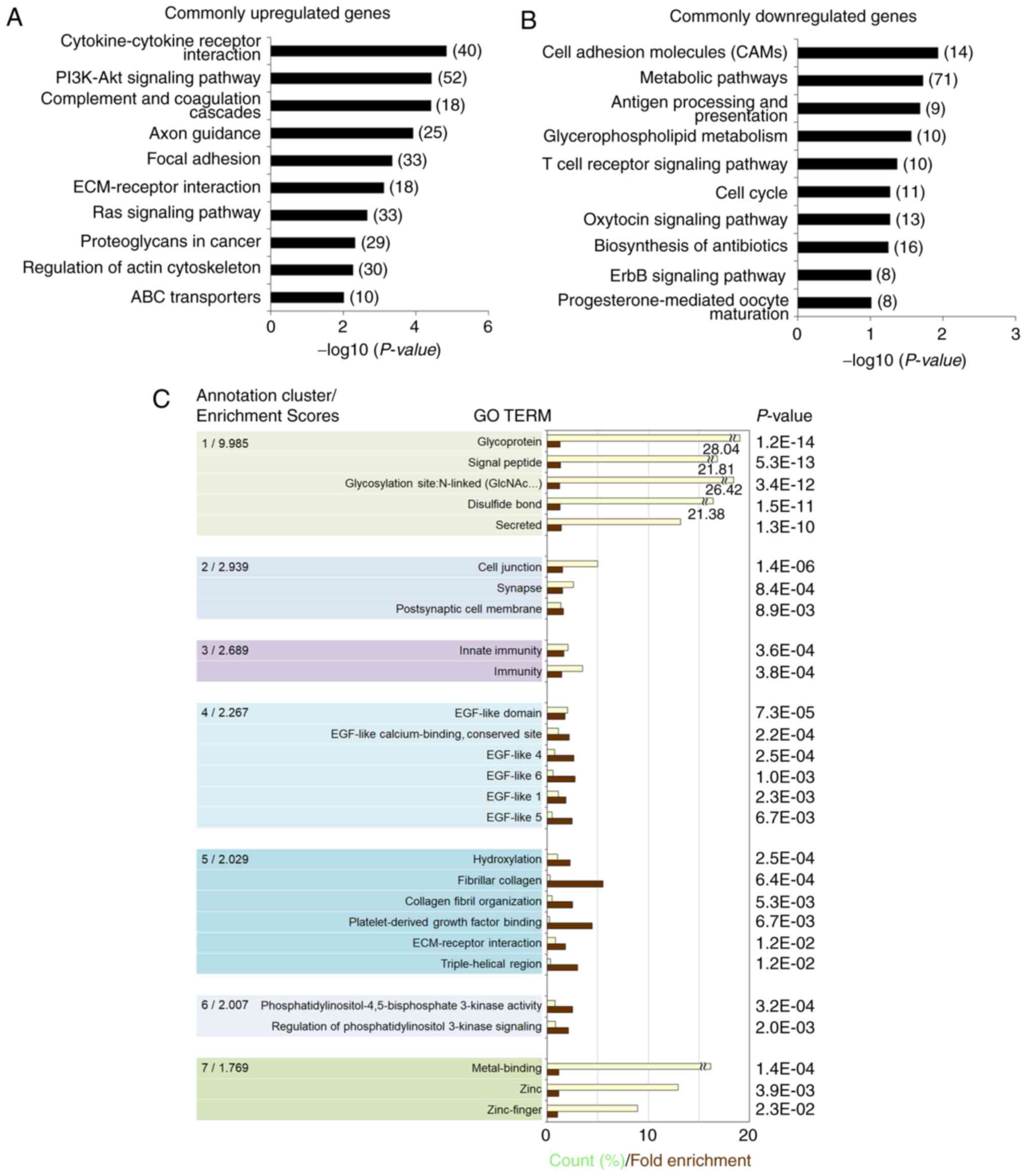
Gene ontology (GO) and pathway enrichment
analysis in hNESs derived from hESCs and ReNcell CX cells
To elucidate the signaling pathways and molecular
mechanisms associated with the hNSC identity, the present study
analyzed the hNSC-specific transcriptome. Pathway analysis based on
the KEGG database showed that the commonly upregulated genes were
significantly associated with the following pathways:
Cytokine-cytokine receptor interaction, PI3K-Akt signaling,
complement and coagulation cascades, axon guidance, focal adhesion,
ECM-receptor interaction, Ras signaling, proteoglycans in cancer,
regulation of actin cytoskeleton, and ABC transporters (Fig. 4A). Cell adhesion molecules,
metabolic pathways, antigen processing and presentation,
glycerophospholipid metabolism, T cell receptor signaling, cell
cycle, oxytocin signaling, biosynthesis of antibiotics, ErbB
signaling, and progesterone-mediated oocyte maturation were
enriched for the commonly downregulated genes in hNESs and ReNcell
CX cells (Fig. 4B).
To obtain a more comprehensive understanding of the
functions of the hNSC-specific transcriptome, GO term enrichment
analysis was performed through DAVID functional annotation
clustering, which shows functionally linked groups by reducing the
redundancy in the annotation (11,27). The following seven significant
annotation clusters were identified in the hNSC-specific
transcriptome, which were related to glycoprotein, cell junction
(synapse), immunity, EGF-like domain, ECM-receptor interaction
(fibrillar collagen), regulation of PI3K signaling and
metal-binding based on statistical criteria (P<0.05 and an
enrichment score of at least 1.7) (Fig. 4C).
Although the majority of the genes analyzed showed
similar expression patterns, differentially expressed genes found
only in a small portion of genes (12.4% of all genes given the
2.0-fold cutoff criterion) between hNESs and ReNcell CX cells were
enriched in the following pathways: TGF-β signaling, acute myeloid
leukemia, malaria, transcriptional misregulation in cancer,
pathways in cancer, axon guidance, signaling pathways regulating
pluripotency of stem cells, and melanoma (Fig. 5). The majority of the
overrepresented pathways were identified as cancer-related
pathways; this may be due to the oncogenic c-MYC having been
functionally linked to cancer-related pathways (28,29).
Neuronal differentiation of hNESs derived
from hESCs and ReNcell CX cells in vitro
To further characterize hNESs derived from hESCs
molecularly, semi-quantitative RT-PCR analysis was performed for
the expression of NSC markers. Transcripts of ESC markers,
including OCT4 and NANOG, were decreased during differentiation
(Fig. 6A). The levels of
molecular markers for NSCs, including superoxide dismutase (SOX)1,
SOX2, SOX3, musashi-1 (MSI1), TUJ1 and NESTIN, were increased in
the hESC-derived hNESs (Fig. 6A).
Dissociated hNES cells and ReNcell CX cells were also immunostained
for NSC markers, including NESTIN and TUJ1 (Fig. 6B). To functionally characterize
neuronal differentiation capacity, hNESs and ReNcell CX cells were
differentiated following the withdrawal of growth factors.
Following 15 days of differentiation, MAP2-positive neuronal cells
and GFAP-positive glial cells were observed in the differentiated
hNESs and ReNcell CX cells (Fig.
6B). Of note, the hESC-derived hNESs continued to expand over
five passages without losing any of their features. It was possible
to scale-up hNES production by ~662-fold in terms of cell number
(mean values) following five passages, with the majority of cells
undergoing active proliferation, as indicated by Ki-67 labeling at
passage 5 (Fig. 6C).
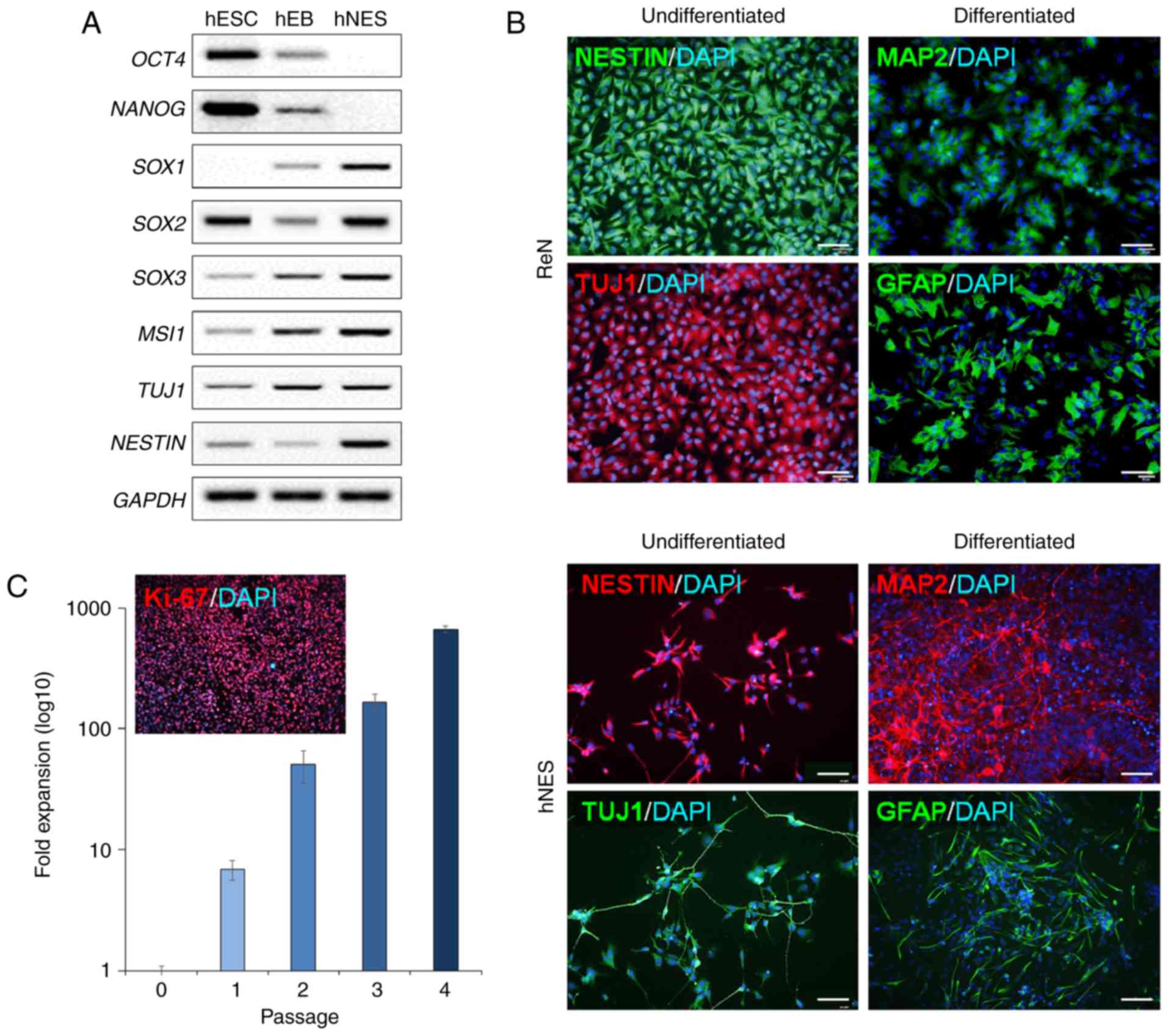 | Figure 6Neuronal differentiation potential of
hESC-derived NSCs and ReNcell CX cells. (A) Semi-quantitative
reverse transcription-polymerase chain reaction analysis of NSC
markers during hNES differentiation. (B) Neuronal differentiation
capacity of hESC-derived hNESs and ReNcell CX cells under
differentiation conditions. Immunostaining of undifferentiated
hNSCs, including hNESs and ReNcell CX cells, with the NSC marker
nestin and the neuronal marker TUJ1, and differentiated hNSCs with
the mature neuronal marker, MAP2, and the mature glial cell marker,
GFAP. Scale bar=200 µm. (C) Expansion rate and
representative images of Ki-67 staining of hESC-derived hNESs.
Fold-expansion was compared with hNESs at passage 0. hNESs contain
Ki-67-positive proliferating NSCs. hESCs, human embryonic stem
cells; hNESs, human neuroectodermal spheres; ReN, ReNcell CX cells;
hEB, human embryonic body; NSC, neural stem cell; MAP2,
microtubule-associated protein 2; GFAP, glial fibrillary acidic
protein; TUJ1, neuron-specific class III β-tubulin; OCT4,
octamer-binding protein 4; SOX, superoxide dismutase; MSI1,
musashi-1. |
The aim of the present study was to characterize
hNSCs and to select a more suitable hNSC model for developing
human-based platforms for applications in various neurological
fields. The hNSC-specific transcriptome data from hESC-derived
hNESs and ReNcell CX hNSCs were described. Global gene expression
profiling enables a systems-based analysis of the biological
processes through GO and pathway enrichment analyses, and of genes
driving differentiation into hNSCs. The comparative analysis of the
global gene expression showed that the hESC-derived hNESs were
similar to the ReNcell CX hNSCs, as shown by PCA and hierarchical
sample clustering. As described above, the hNESs derived from hESCs
in the simple protocol exhibited differentiation potential, based
on the specific terminal differentiation markers of neuronal and
glial cells that were detected. The hNESs showed stable
proliferation and were expanded for at least five passages without
loss of NSC characteristics. These results indicated that
hESC-derived hNESs may be used as a relevant hNSC model, similar to
ReNcell CX cells, in several neurological research fields.
hNSC differentiation protocols remain inefficient,
with poor yields of terminal differentiation and maturation into
specific neuronal cell types. However, increased understanding of
the mechanisms underlying hESC-based NSC differentiation through
fine-tuning protocols for the efficient derivation, long-term
maintenance and neuronal cell type-specific differentiation of
hNSCs may provide novel insights into human neurodevelopment and
the process of NSC fate specification.
Acknowledgments
This study was supported by the National Research
Foundation of Korea grant funded by the Ministry of Science, ICT
and Future Planning (grant nos. 2016R1A2B4013501 and
NRF-2016M3A9C4953144) and a grant from the KRIBB Research
Initiative Program. The funders had no involvement in study design,
data collection or analysis, decision to publish or preparation of
the manuscript.
Abbreviations:
|
bFGF
|
basic fibroblast growth factor
|
|
EGF
|
epidermal growth factor
|
|
hEB
|
human embryoid body
|
|
hESC
|
human embryonic stem cell
|
|
hNSC
|
human neural stem cell
|
|
NES
|
neuroectodermal sphere
|
|
NSC
|
neural stem cell
|
|
PCA
|
principal component analysis
|
References
|
1
|
Oh JH, Son MY, Choi MS, Kim S, Choi AY,
Lee HA, Kim KS, Kim J, Song CW and Yoon S: Integrative analysis of
genes and miRNA alterations in human embryonic stem cells-derived
neural cells after exposure to silver nanoparticles. Toxicol Appl
Pharmacol. 299:8–23. 2016. View Article : Google Scholar
|
|
2
|
Son MY, Sim H, Son YS, Jung KB, Lee MO, Oh
JH, Chung SK, Jung CR and Kim J: Distinctive genomic signature of
neural and intestinal organoids from familial Parkinson's disease
patient-derived induced pluripotent stem cells. Neuropathol Appl
Neurobiol. 43:584–603. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Otsu M, Nakayama T and Inoue N:
Pluripotent stem cell-derived neural stem cells: From basic
research to applications. World J Stem Cells. 6:651–657. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao X, Yang Z, Liang G, Wu Z, Peng Y,
Joseph DJ, Inan S and Wei H: Dual effects of isoflurane on
proliferation, differentiation, and survival in human
neuroprogenitor cells. Anesthesiology. 118:537–549. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Díaz-Coránguez M, Segovia J, López-Ornelas
A, Puerta-Guardo H, Ludert J, Chávez B, Meraz-Cruz N and
González-Mariscal L: Transmigration of neural stem cells across the
blood brain barrier induced by glioma cells. PLoS One. 8:pp.
e606552013, View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hovakimyan M, Müller J, Wree A, Ortinau S,
Rolfs A and Schmitt O: Survival of transplanted human neural stem
cell line (ReNcell VM) into the rat brain with and without
immunosuppression. Ann Anat. 194:429–435. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Choi SH, Kim YH, Quinti L, Tanzi RE and
Kim DY: 3D culture models of Alzheimer's disease: A road map to a
ʻcure-in-a-dishʼ. Mol Neurodegener. 11:752016. View Article : Google Scholar
|
|
8
|
Kim YH, Choi SH, D'Avanzo C, Hebisch M,
Sliwinski C, Bylykbashi E, Washicosky KJ, Klee JB, Brüstle O, Tanzi
RE and Kim DY: A 3D human neural cell culture system for modeling
Alzheimer's disease. Nat Protoc. 10:985–1006. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Woo SM, Kim J, Han HW, Chae JI, Son MY,
Cho S, Chung HM, Han YM and Kang YK: Notch signaling is required
for maintaining stem-cell features of neuroprogenitor cells derived
from human embryonic stem cells. BMC Neurosci. 10:972009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Breunig JJ, Haydar TF and Rakic P: Neural
stem cells: Historical perspective and future prospects. Neuron.
70:614–625. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Son MY, Kwak JE, Kim YD and Cho YS:
Proteomic and network analysis of proteins regulated by REX1 in
human embryonic stem cells. Proteomics. 15:2220–2229. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chambers SM, Fasano CA, Papapetrou EP,
Tomishima M, Sadelain M and Studer L: Highly efficient neural
conversion of human ES and iPS cells by dual inhibition of SMAD
signaling. Nat Biotechnol. 27:275–280. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dhara SK and Stice SL: Neural
differentiation of human embryonic stem cells. J Cell Biochem.
105:633–640. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Son MY, Lee MO, Jeon H, Seol B, Kim JH,
Chang JS and Cho YS: Generation and characterization of
integration-free induced pluripotent stem cells from patients with
autoimmune disease. Exp Mol Med. 48:pp. e2322016, View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jung KB, Son YS, Lee H, Jung CR, Kim J and
Son MY: Transcriptome dynamics of human pluripotent stem
cell-derived contracting cardiomyocytes using an embryoid body
model with fetal bovine serum. Mol Biosyst. 13:1565–1574. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim DS, Ryu JW, Son MY, Oh JH, Chung KS,
Lee S, Lee JJ, Ahn JH, Min JS, Ahn J, et al: A Liver-specific Gene
Expression panel predicts the differentiation status of in vitro
hepatocyte models. Hepatology. 66:1662–1674. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Son MY, Kwak JE, Seol B, Lee DY, Jeon H
and Cho YS: A novel human model of the neurodegenerative disease
GM1 gangliosidosis using induced pluripotent stem cells
demonstrates inflammasome activation. J Pathol. 237:98–110. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Son MY, Kim HJ, Kim MJ and Cho YS:
Physical passaging of embryoid bodies generated from human
pluripotent stem cells. PLoS One. 6:pp. e191342011, View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nakagawa M, Takizawa N, Narita M, Ichisaka
T and Yamanaka S: Promotion of direct reprogramming by
transformation-deficient Myc. Proc Natl Acad Sci USA.
107:14152–14157. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kwak JE, Son MY, Son YS, Son MJ and Cho
YS: Biochemical and molecular characterization of novel mutations
in GLB1 and NEU1 in patient cells with lysosomal storage disorders.
Biochem Biophys Res Commun. 457:554–560. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jung KB, Lee H, Son YS, Lee JH, Cho HS,
Lee MO, Oh JH, Lee J, Kim S, Jung CR, et al: In vitro and in vivo
imaging and tracking of intestinal organoids from human-induced
pluripotent stem cells. FASEB J. Aug 29–2017.Epub ahead of print.
View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
23
|
Son MY, Kim YD, Seol B, Lee MO, Na HJ, Yoo
B, Chang JS and Cho YS: Biomarker discovery by modeling behcet's
disease with patient-specific human induced pluripotent stem cells.
Stem Cells Dev. 26:133–145. 2017. View Article : Google Scholar
|
|
24
|
Donato R, Miljan EA, Hines SJ, Aouabdi S,
Pollock K, Patel S, Edwards FA and Sinden JD: Differential
development of neuronal physiological responsiveness in two human
neural stem cell lines. BMC Neurosci. 8:362007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Oganesyan Li Z, Mooney D, Rong R,
Christensen X, Shahmanyan MJ, Perrigue D, Benetatos PM, Tsaturyan
J, Aramburo LS, et al: L-MYC expression maintains self-renewal and
prolongs multipotency of primary human neural stem cells. Stem Cell
Reports. 7:483–495. 2016. View Article : Google Scholar
|
|
26
|
Li X, Xu J, Bai Y, Wang X, Dai X, Liu Y,
Zhang J, Zou J, Shen L and Li L: Isolation and characterization of
neural stem cells from human fetal striatum. Biochem Biophys Res
Commun. 326:425–434. 2005. View Article : Google Scholar
|
|
27
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mazan-Mamczarz K, Hagner PR, Dai B, Wood
WH, Zhang Y, Becker KG, Liu Z and Gartenhaus RB: Identification of
transformation-related pathways in a breast epithelial cell model
using a ribonomics approach. Cancer Res. 68:7730–7735. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Miller DM, Thomas SD, Islam A, Muench D
and Sedoris K: c-Myc and cancer metabolism. Clin Cancer Res.
18:5546–5553. 2012. View Article : Google Scholar : PubMed/NCBI
|