Introduction
Host immune cells utilize Toll-like receptors (TLRs)
to recognize pathogen-associated molecular patterns to transmit
intracellular signals via kinase cascades, which ultimately
activate transcription factors, leading to the expression of
proinflammatory cytokines. However, excessive production of these
cytokines disrupts immune homeostasis, and even causes tissue
damage (1). To antagonize
aberrant inflammatory signaling and maintain homeostasis, negative
regulators are also required for inflammatory signaling. Previous
studies in both humans and mutant mice with inflammatory diseases
have verified some critical inhibitory pathways, including the
transforming growth factor-β (TGF-β) and interleukin-10 (IL-10)
pathways (2). Furthermore, the
deubiquitinases A20 and CYLD, the p110d kinase, and the SHP-1/2 and
MKP-1/5 phosphatases have been identified as negative regulators of
inflammation in the TLR4 signaling pathway (3–7).
cMaf is a transcription factor with basic leucine
zipper domains that mediate DNA binding to the cMaf recognition
element (8,9). cMaf is expressed in monocyte and
macrophage lineages, as well as in T cell subsets (10,11). Studies of transgenic cMaf T helper
(Th) cells revealed that cMaf is important for the development of
Th1 and Th17 cells (12–14). A functional analysis showed that
TGF-β induces cMaf expression, which suppresses IL-22 expression in
Th17 cells, enhances proinflammatory innate defense mechanisms in
epithelial cells, and provides crucial protection to tissues from
damage caused by inflammation and infection (15). Furthermore, cMaf regulates IL-10
expression in macrophages by directly binding to the IL-10 promoter
(16).
Mammalian target of rapamycin (mTOR) can form two
distinct complexes, mTOR complex 1 (mTORC1) and mTORC2, although
the kinase-specific inhibitor rapamycin mainly targets mTORC1,
which controls translation mainly by phosphorylating the
translation initiation factor eIF4E-binding protein 1 (4EBP-1) and
the S6 ribosomal kinase 1 (S6K1) (17,18). Moreover, mTOR also takes part in
controlling glycolysis by promoting the expression of the
transcription factors HIF-1α and c-Myc (19,20), and upregulating the expression of
nutrient transporters. mTOR also participates in the control of
lipid synthesis by activating the transcription factor sterol
regulatory element-binding protein (SREBP), as well as in the
control of autophagy (21,22).
However, the precise mechanisms by which mTOR responds to TLR4
signaling in macrophages remain to be identified.
In this study, we demonstrated that cMaf expression
is induced following lipopolysaccharide (LPS) challenge, which
suggests that cMaf functions in the TLR4 signaling pathway.
Knockdown of cMaf expression in macrophages impaired IL-10
production, but increased the expression of IL-1, IL-6, IL-12 and
tumor necrosis factor-α (TNF-α). A pathway analysis using
inhibitors indicated that TLR4 signaling activated extracellular
signal-related kinase (ERK) and phosphoinositide 3-kinase (PI3K),
which phosphorylate mTOR and control the translation, but not
transcription, of cMaf via the downstream kinases S6K1 and 4EBP-1,
thereby regulating inflammatory cytokine production in macrophages.
In vivo, altered TLR4 signaling resulted in pathological
symptoms of alveolar damage and led to more endotoxin-induced death
of cMaf-knockdown macrophages in mice.
Materials and methods
Animals
Wild-type mice (C57/BL6) were supplied by the
Shanghai Laboratory Animal Center (Shanghai, China). Animals were
treated humanely, and all animal experiments conformed to the
recommendations of the Guide for the Care and Use of Laboratory
Animals published by the U.S. National Institutes of Health (NIH
publication no. 85–23, revised 1996), and the study was approved by
the Ethics Committee of the Shanghai Institutes for Biological
Sciences, Chinese Academy of Sciences. Animal survival rates were
calculated at 1, 2, 3, 4, 5, 6, 7 and 8 days after LPS
injection.
For the animal survival study, the animal death was
used as the clinical endpoint. All mice were monitored every 6 h
for 7 days. Mice were anesthetized with ketamine followed by
cervical dislocation when they were found in a moribund state which
was characterized by labored breathing and/or no-responsiveness to
cape tapping. At the end of the study, all surviving mice were
anesthetized by ketamine overdose, followed by cervical
dislocation.
Reagents and cells
Roswell Park Memorial Institute (RPMI)-1640 medium,
fetal bovine serum (FBS), penicillin, and streptomycin were
purchased from Invitrogen (Shanghai, China). All signaling pathway
inhibitors were obtained from Calbiochem (San Diego, CA, USA).
RAW264.7 macrophages were purchased from the American Type Culture
Collection (Manassas, VA, USA) and cultured in RPMI-1640 medium
supplemented with 10% FBS, 100 U/ml penicillin, and 100 U/ml
streptomycin, and maintained in a humidified incubator with 5%
CO2 at 37°C. For preparation of peritoneal macrophages,
2 ml of 4% thioglycollate medium (R&D Systems, Minneapolis, MN,
USA) was injected into the peritoneal cavity of mice. After 3 days,
mice were euthanized, and the peritoneal cavity was flushed twice
with 6–8 ml of phosphate-buffered saline (PBS). Collected
peritoneal cells were seeded in cell culture dishes containing
complete RPMI-1640 medium (10% FBS). Non-adherent cells were washed
off, and adherent cells were stimulated the following day. To
generate bone marrow-derived macrophages (BMDMs), bone marrow cells
were cultured in RPMI-1640 medium supplemented with 30%
L929-conditioned medium (containing macrophage colony-stimulating
factor [M-CSF]) and 10% FBS. On day 4, non-adherent cells were
removed, and fresh RPMI-1640 medium supplemented with
L929-conditoned medium was added. BMDMs were used on 7–10 days.
Quantitative real-time polymerase chain
reaction (qPCR)
TRIzol reagent (Invitrogen, Carlsbad, CA, USA) was
used to isolate total RNA from the target cells, and cDNA was
generated using the ReverTra Ace® qPCR RT kit (Toyobo,
Osaka, Japan). The SYBR® Premix Ex Taq™ kit (Takara,
Shiga, Japan) and an ABI 7500 LightCycler (Applied Biosystems,
Waltham, MA, USA) were used for the qPCR analysis. The following
primer sequences were used: IL-1β forward,
5′-CAACCAACAAGTGATATTCTCCATG-3′ and reverse,
5′-GATCCACACTCTCCAGCTGCA-3′; IL-6 forward,
5′-AGATAAGCTGGAGTCACAGAAGGAG-3′ and reverse,
5′-CGCACTAGGTTTGCCGAGTA-3′; IL-10 forward,
5′-ATTTGAATTCCCTGGGTGAGAAG-3′ and reverse,
5′-CACAGGGGAGAAATCGATGACA-3′; IL-12 forward,
5′-CACCCTTGCCCTCCTAAAC-3′ and reverse, 5′-CACCTGGCAGGTCCAGAG-3′;
TNF-α forward, 5′-GTCCCCAAAGGGATGAGAAGTT-3′ and reverse,
5′-GTTTGCTACGACGTGGGCTACA-3′; glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) forward, 5′-TGGAGAAACCTGCCAAGTATGA-3′ and
reverse, 5′-CTGTTGAAGTCGCAGGAGACAA-3′; cMaf forward,
5′-CTGAGCCAAGATTCATGTATGGG-3′ and reverse,
5′-CGTTGCACCATCCAAACAGT-3′.
RNA interference and transfection
RAW264.7 cells were seeded at a density of
10×106 cells/well in 6-well plates and transfected with
a control small interfering RNA (siRNA) or a cMaf siRNA using the
Silencer® siRNA Transfection II kit (Ambion, Austin, TX,
USA) according to the manufacturer's protocol. The siRNA sequences
for mouse cMaf were designed and synthesized by GenePharma Co.,
Ltd. (Shanghai, China). The siRNA-1 and 2 sequences were
5′-ACAGCGAGCCCAACCUUAUUG-3′ and 5′-CUAAUUCUUAGAGCUUCAUAU-3′,
respectively, and the control siRNA sequence was
5′-AAUGCCUACGUUAAGCUAUAC-3′.
Enzyme-linked immunosorbent assay
(ELISA)
IL-1, IL-6, IL-10, IL-12 and TNF-α levels in the
cell culture supernatants were quantified using ELISA kits (R&D
Systems) according to the manufacturer's instructions.
Western blotting
To prepare whole cell lysates, cells were washed
three times with PBS and incubated in lysis buffer for 30 min on
ice. After boiling in sodium dodecyl sulfate (SDS) loading buffer,
equal amounts of protein were subjected to SDS-polyacrylamide gel
electrophoresis, and transferred to a nitrocellulose membrane.
Then, the membrane was blocked in 5% nonfat dried milk containing
0.1% Tween-20 for 1 h at room temperature, and incubated at 4°C
overnight with primary antibodies against p-S6 (#4858, 1:1,000
dilution), p-p70S6K (#9234, 1:1,000 dilution), p-mTOR (#5536,
1:1,000 dilution), p-4EBP1 (#9451, 1:1,000 dilution) (Cell
Signaling Technology, Danvers, MA, USA). Anti-GAPDH antibody
(#AP0063, 1:2,000 dilution) was obtained from Bioworld
(Minneapolis, MN, USA). Anti-cMaf antibody (sc-7866, 1:500
dilution) was from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA). The membrane was washed three times with PBS and further
incubated with a horseradish peroxidase-conjugated secondary
antibody for 2 h at room temperature. After a final wash,
immunoreactive bands were visualized using the Scientific
SuperSignal West Pico Chemiluminescent Substrate reagent (Pierce,
Rockford, IL, USA).
Lentivirus and infection
To stably express the cMaf siRNAs in BMDMs,
pLKO.1-GFP or pLKO.1-puro plasmids expressing small hairpin RNAs
(shRNAs) targeting mouse cMaf were transiently transfected into
human embryonic kidney 293T cells, together with packaging plasmids
(psPAX2 and pMD2G), and virus-containing media were harvested at 48
h. Supernatants containing shRNA-expressing lentiviruses were added
to BMDMs at day 3. At 24 h, the medium was removed and fresh
RPMI-1640 medium containing 2 µg/ml puromycin was added.
After selection for 4–6 days, positive BMDMs were enriched and
replated for LPS stimulation. The two shRNA targeting sequences for
cMaf (cMaf-1, 5′-AGGAACTGAAGTACGTGATTC-3′; cMaf-2,
5′-GTGGTTCAGAGGATCCTTAAA-3′) were from Sigma-Aldrich (St. Louis,
MO, USA) TRC libraries (TRCN0000346430 and TRCN0000376317). BMDMs
infected with green fluorescent protein (GFP)-expressing
lentiviruses were detected by laser confocal microscope, and the
knockdown efficiency was determined by immunoblotting using the
anti-cMaf antibody.
In vivo knockdown and depletion of
macrophages
Procedures for delivering viruses encoding shRNAs
into mice and depleting cMaf in macrophages were performed as
described previously (23).
Histological examinations
Tissue samples were collected 72 h after endotoxin
shock and immediately fixed in 4% paraformaldehyde for 24 h. Then,
tissue sections were embedded in paraffin, cut into 4–5 µm
sections, and stained with hematoxylin and eosin. Two experienced
pathologists who were blinded to the protocol performed the
histological examinations. To grade the degree of lung injury, a
scoring system was used based on the following histological
features: edema, hyperemia and congestion, neutrophil margination
and tissue infiltration, intra-alveolar hemorrhage and debris, and
cellular hyperplasia. Each feature was graded as absent, mild,
moderate, or severe, with a score of 0 to 3, respectively, and
total scores were calculated for each animal.
Statistical analysis
Statistically significant differences between groups
were determined by two-tailed Student's t-test and two-way analysis
of variance (ANOVA). Survival was analyzed with the log-rank test.
GraphPad Prism version 5.0 software was used for all analyses.
P<0.05 was considered statistically significant and P<0.01 as
highly significant.
Results
cMaf regulates inflammatory cytokine
production in macrophages upon TLR4 activation
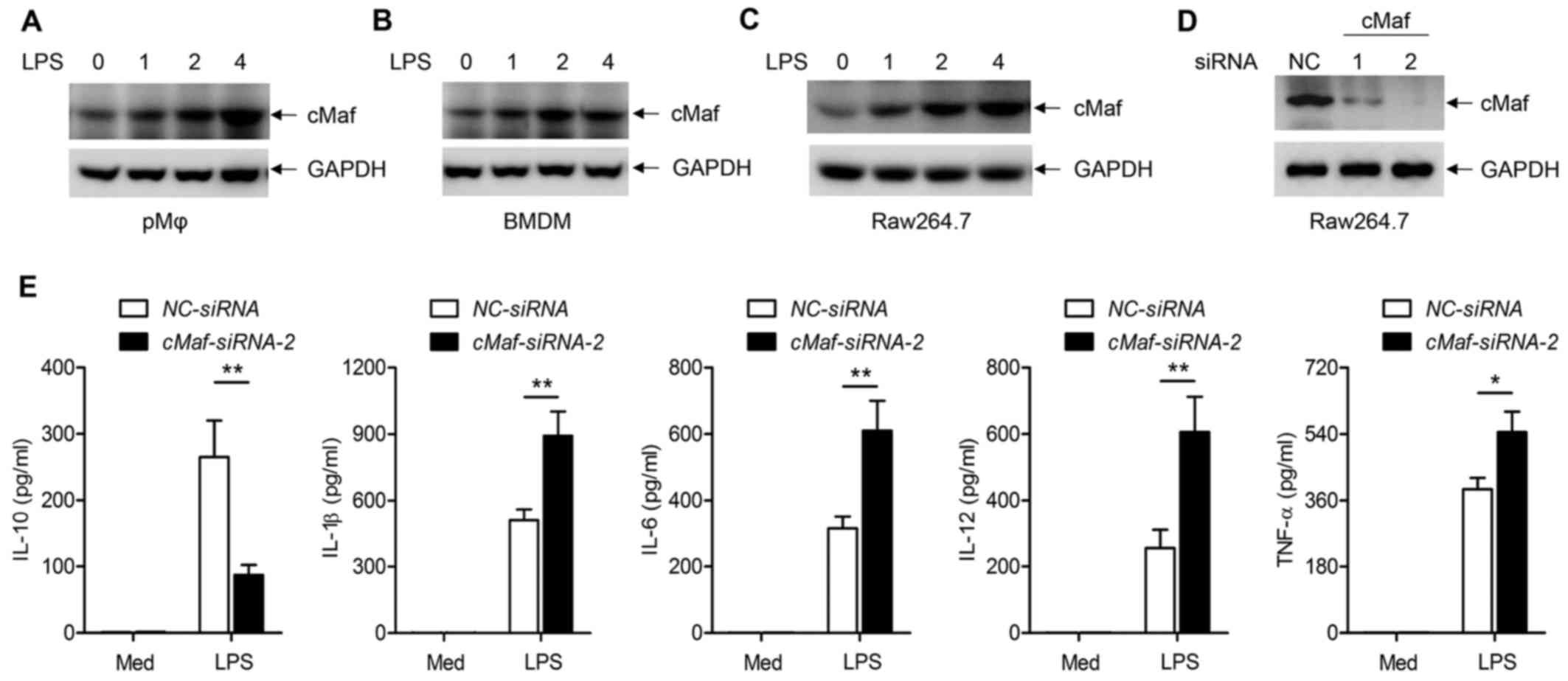
To understand the functions of cMaf in inflammatory
responses in macrophages, we first tested whether LPS directly
affects cMaf expression in three typical macrophage types
(peritoneal macrophages, BMDMs, and RAW64.7 cells). Upon treatment
with LPS for 0, 1, 2 and 4 h, there was a time-dependent increase
of cMaf expression in the three different macrophage types, as
determined by western blotting (Fig.
2A–C). Furthermore, we explored whether this increased cMaf
expression accounted for the LPS-induced inflammatory response.
First, we silenced endogenously expressed cMaf in RAW264.7 cells
with two specific siRNAs, and western blot results verified that
the expression of endogenous cMaf was obviously reduced (Fig. 2D). The supernatants of RAW264.7
cells with different treatment were collected and inflammatory
cytokines were detected by ELISAs. The results showed that
inflammatory cytokines, such as IL-1, IL-6, IL-10, IL-12 and TNF-α,
were significantly induced by LPS. However, knockdown of cMaf in
peritoneal macrophages impaired the production of IL-10, but
increased the LPS-mediated production of IL-1, IL-6, IL-12 and
TNF-α (Fig. 2E). These data show
that LPS-induced cMaf regulates inflammatory cytokine production in
macrophages.
ERK1/2 and PI3K phosphorylate mTOR to
regulate the translation of cMaf
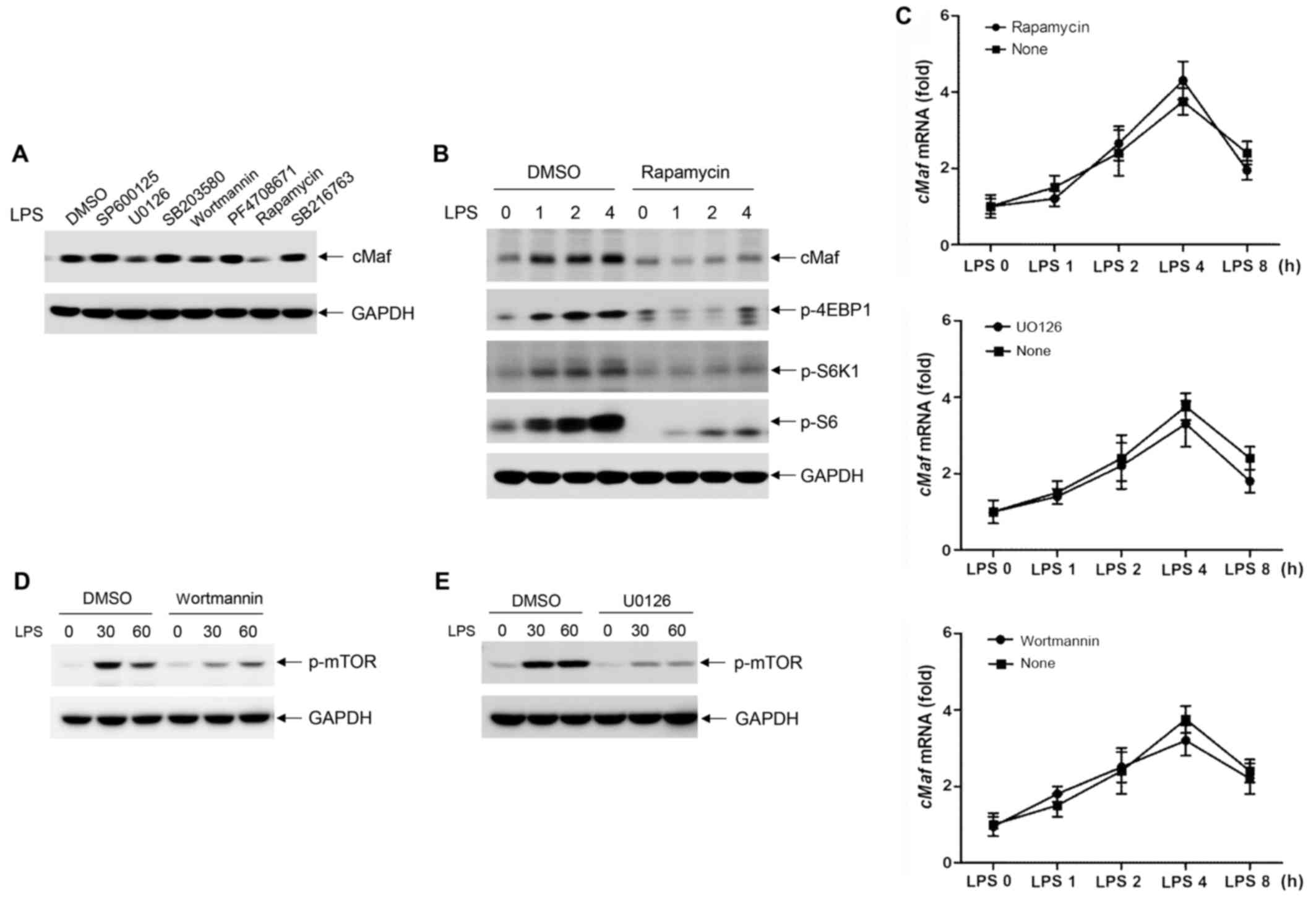
To analyze the regulation of cMaf in TLR4-stimulated
macrophages, the inhibitors SP600125, U0126, SB203580, wortmannin,
PF4708671, rapamycin and SB216763 were used to determine the
involvement of Jun-kinase (JNK), ERK1/2, p38 mitogen-activated
protein kinase, PI3K, S6K1, mTOR and glycogen synthase kinase,
respectively, in LPS-stimulated cMaf expression. Interestingly, we
found that pretreatment of BMDMs with U0126, wortmannin, or
rapamycin significantly reduced LPS-induced cMaf expression;
however, cMaf expression remained unaltered in infected macrophages
that were pretreated with the other inhibitors (Fig. 3A).
It is known that rapamycin inhibits mTOR. To further
validate that the mTOR pathway is involved in LPS-induced cMaf
expression, we treated BMDMs with rapamycin for 0, 1, 2 and 4 h. As
shown in Fig. 3B, rapamycin
strongly inhibited the phosphorylation of 4EBP1 and S6K1, which are
downstream of mTOR, and western blot analyses showed that blocking
the mTOR pathway significantly inhibited LPS-induced cMaf
expression (Fig. 3B). However,
qPCR analyses showed that cMaf mRNA remained unaltered in infected
macrophages that were pretreated with rapamycin as well as U0126 or
wortmannin (Fig. 3C).
Furthermore, the results showed that U0126 and
wortmannin also reduced LPS-mediated cMaf expression (Fig. 3A). To investigate the roles of the
PI3K and ERK pathways in LPS-induced mTOR activation, wortmannin
and U0126 were used to inhibit the activation of PI3K and ERK,
respectively. As shown in Fig. 2D and
E, LPS-mediated mTOR phosphorylation was significantly
inhibited by both U026 and wortmannin. These data suggest that
ERK1/2 and PI3K phosphorylate mTOR to regulate cMaf expression at
the translational level.
mTOR regulates inflammatory gene
expression in LPS-stimulated macrophages
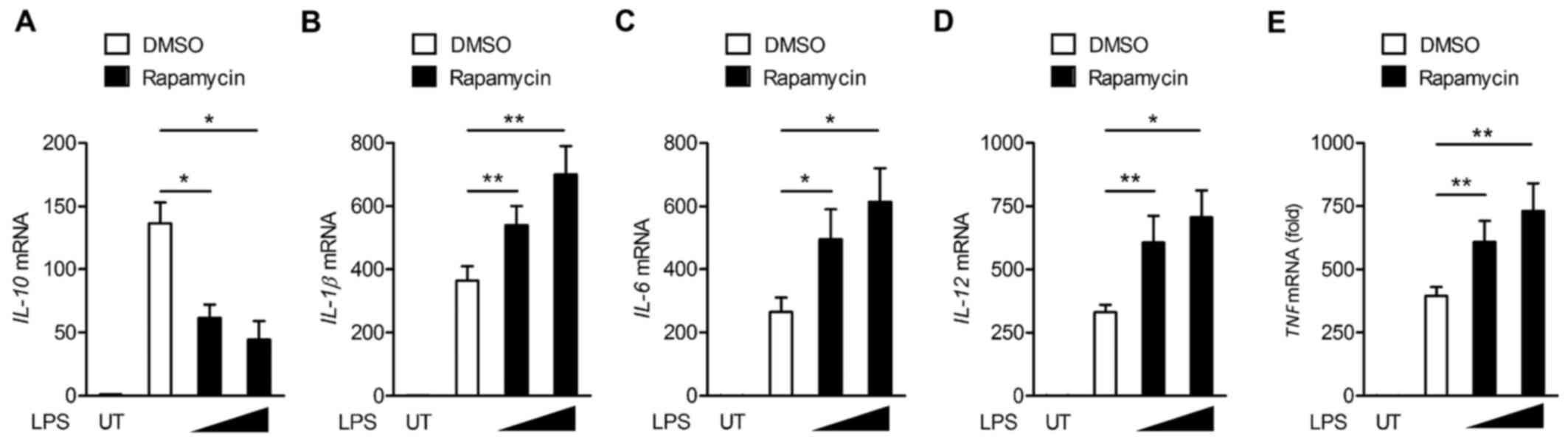
Our prior results indicated that the activation of
mTOR is involved in LPS-induced cMaf expression. Next, we assessed
whether mTOR regulates inflammatory gene expression in LPS-induced
macrophages. A qPCR analysis showed that the inhibition of mTOR by
rapamycin impaired the production of IL-10 in LPS-treated BMDMs,
but increased the production of IL-1, IL-6, IL-12 and TNF-α
(Fig. 4).
mTOR regulates inflammatory cytokine
production via cMaf in LPS-stimulated macrophages
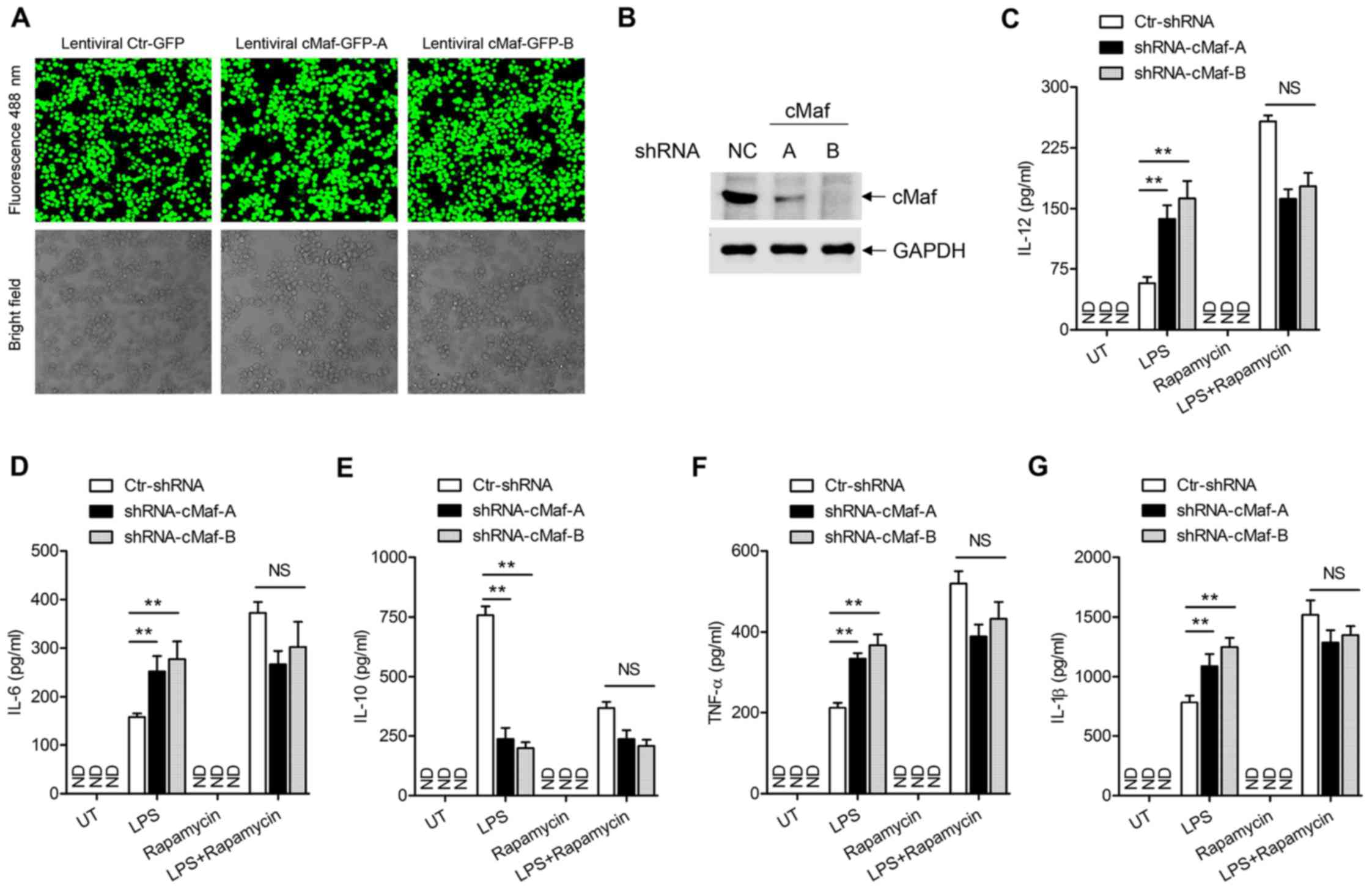
To further verify the importance of cMaf in the
mTOR-mediated regulation of inflammatory cytokines in
LPS-stimulated macrophages, BMDMs were stably transfected with
recombinant lentiviruses expressing a control GFP shRNA or two
distinct cMaf shRNA. The lentiviral transfection and protein
knockdown efficiency in BMDMs stably expressing the control or cMaf
shRNAs was verified by laser confocal microscope (Fig. 5A) and western blotting in Fig. 5B. Next, we determined whether
depleting cMaf could neutralize the effect of rapamycin on the TLR4
signaling pathway. ELISA analyses showed that the production of
these cytokines was mostly unaffected when cMaf expression was
knocked down in BMDMs (Fig.
5C–G). Together, these results demonstrate that mTOR inhibits
the production of inflammatory cytokines and increases IL-10
production via cMaf.
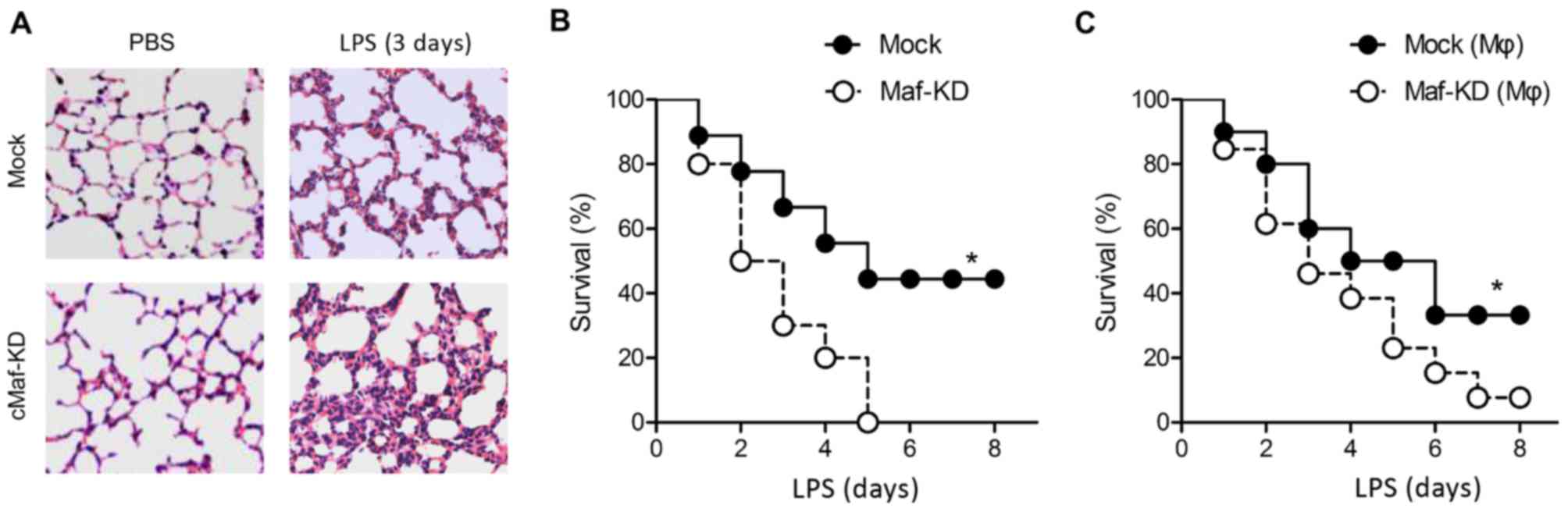
cMaf-knockdown mice are more susceptible
to endotoxin shock
To verify our mechanistic understanding of cMaf
in vivo, we knocked down cMaf expression in mice by
injecting a cMaf-specific shRNA, and we subjected the mice to
septic shock. Mice in which cMaf expression was knocked down
(cMaf-KD mice) displayed more severe lung injuries than did mice
that underwent a mock knockdown of cMaf (mock mice) 3 days after
acute challenge with LPS (Fig.
6A). Accordingly, the survival rate of LPS-challenged cMaf-KD
mice was significantly lower than that of their LPS-treated mock
counterparts. Administration of LPS caused a 50% mortality rate
within 8 days in the mock mice, whereas cMaf-KD mice were more
susceptible to endotoxin shock, as evidenced by the almost 100%
mortality rate within 8 days (Fig.
6B). These results are consistent with the cytokine production
results in cMaf-KD macrophages (Fig.
2E). To further investigate the importance of cMaf in
macrophages, we performed an adoptive cell transfer in an
experimental model in which cMaf expression was knocked down in the
donor cells through the use of a cMaf shRNA. Upon LPS challenge,
the survival rate of mice that received macrophages that were
treated with a control shRNA with a scrambled sequence was 36%,
while that of mice that received macrophages that were treated with
cMaf-specific shRNA was only 5% (Fig.
6C). These results indicate that the effect of cMaf on the
inflammatory response is largely mediated via macrophages.
Discussion
Aberrant inflammatory responses can cause unexpected
damage to a host. mTOR functions as a key signal transducer for
TLR-mediated inflammation. Subtle regulation of mTOR activity is
crucial for maintaining immunological homeostasis. Here, our
results show that cMaf, a leucine-zipper transcription factor, is
targeted by mTOR in TLR4 signaling, and that cMaf primes
TLR4-related inflammatory responses in macrophages.
In the present study, we observed the inducible
expression of cMaf in different macrophages (peritoneal
macrophages, BMDMs and RAW294.7 cells) following short-term LPS
stimulation. As reported previously, cMaf expression also could be
induced by cytokines such as IL-10, M-CSF, or TGF-β, which are
expressed in T cells and macrophages (24).
Although PI3K and ERK have been implicated in IL-10
gene expression (25), the
underlying mechanisms are not fully understood. Here, we first
showed that the expression of the transcription factor cMaf was
controlled by mTOR at the translational level, and that cMaf
converged with upstream ERK and PI3K signaling pathways in response
to TLR4 stimulation. cMaf is reported to bind to the IL-10 gene
promoter, which induces IL-10 expression in macrophages (16). Our data showed that blocking ERK,
PI3K and mTOR with inhibitors dramatically downregulated IL-10
expression both at the mRNA and protein levels. This confirmed the
phenotypes of cMaf-KD macrophage upon LPS stimulation, which is
consistent with previously reported results. Interestingly, cMaf-KD
macrophages also exhibited upregulated IL-1, IL-6, IL-12 and TNF-α
expression, which is consistent with the inhibition of mTOR. The
most probable reason for this may be the inhibition of IL-10
signaling. IL-10 utilizes the IL-10 receptor, and the adaptor Janus
kinase phoshorylates signal transducer and activator of
transcription 3 (STAT3) at tyrosine 705. STAT3 functions as a
repressor that limits inflammatory gene expression via feedback
regulation, as was reported previously (25–28). Further studies are required to
measure the expression of inflammatory genes in IL-10 knockout
macrophages. Alternatively, cMaf functions as a dual regulator by
assembling into complexes with different partners.
Together, our study showed that cMaf expression was
induced in macrophages in response to LPS challenge, and the fact
that cMaf protected mice from septic shock indicated that cMaf may
improve host fitness, thereby enabling the survival of certain
infectious diseases. Nevertheless, an analysis of the expression of
inflammatory cytokines in cMaf-KD macrophages indicated that
cMaf-mediated inhibition of inflammatory damage mostly depended on
macrophages. A signaling pathway analysis indicated the existence
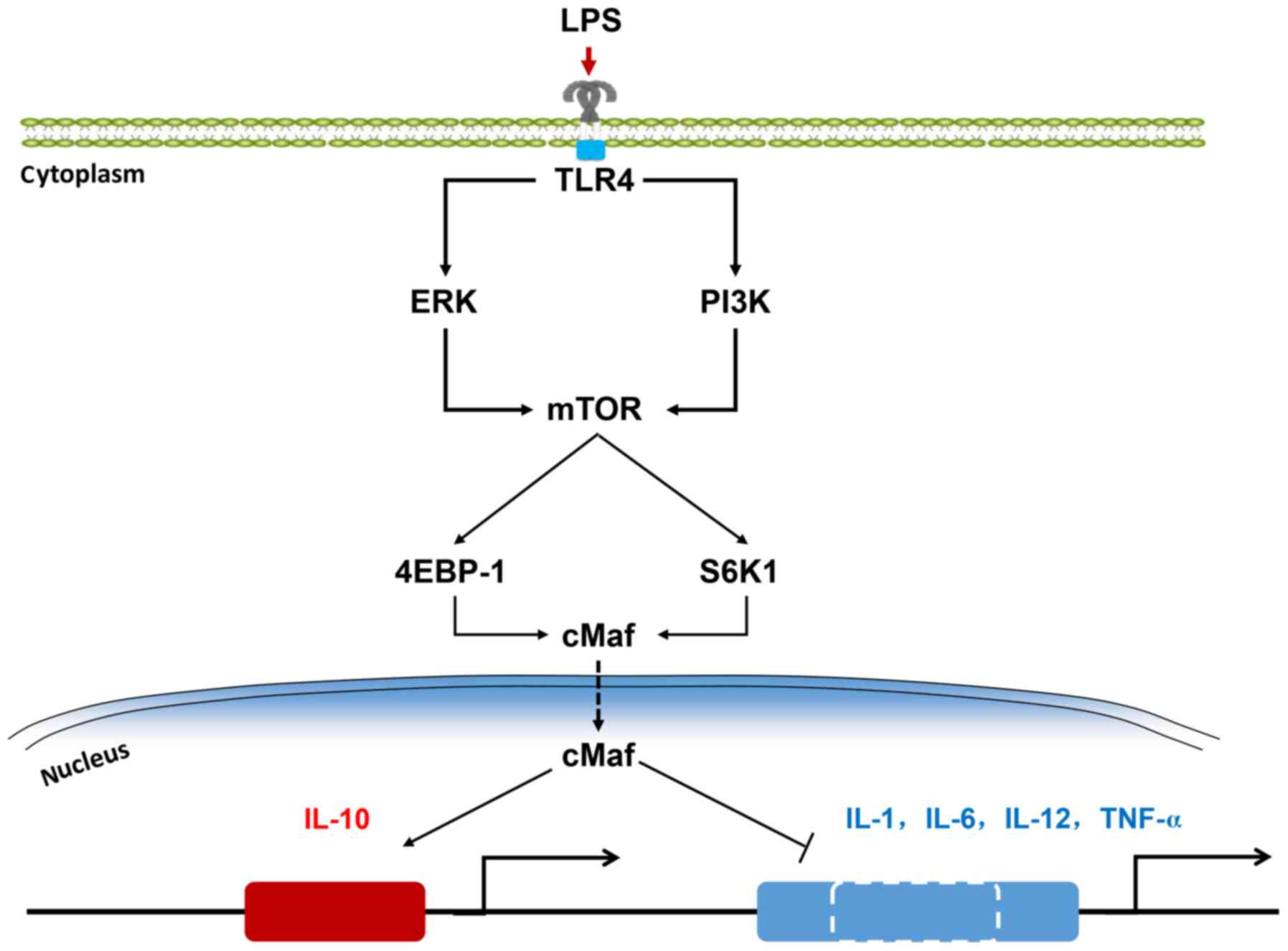
of an ERK- or PI3K-driven mTOR-cMaf regulatory axis (Fig. 1). Previous studies showed that the
Ras-ERK and PI3K pathways are related with cellular growth or
migration, and that macrophages have been linked to tumor genesis
as a result of altered immune system function (23). Therefore, we speculate that cMaf
may also function in regulating tumor-related inflammatory
responses via macrophages. Overall, the study of the combined
functions of cMaf and mTOR in macrophage-related inflammation
provide new insights into the search of new drug targets for acute
inflammatory response.
Acknowledgments
Not applicable.
Notes
[1]
Funding
No funding was received.
[2] Availability
of data and material
All the data and material in the manuscript are
fully available without restriction.
[3] Authors'
contributions
CS conceived and designed the experiment. YW and CL
performed the experiment. GZ analyzed the data. All authors read
and approved the manuscript.
[4] Ethics
approval and consent to participate
Approved by Ethics Committee of the Shanghai
Institutes for Biological Sciences, Chinese Academy of
Sciences.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
[7] Authors'
information
YW, Clinical laboratory, Shanghai Pudong New
District Zhoupu Hospital, Shanghai; CL, GZ and CS, Clinical
laboratory, Yantai Yuhuangding Hospital, Yantai, Shangdong.
References
|
1
|
Beutler B: Inferences, questions and
possibilities in Toll-like receptor signalling. Nature.
430:257–263. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Murray PJ and Smale ST: Restraint of
inflammatory signaling by interdependent strata of negative
regulatory pathways. Nat Immunol. 13:916–924. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liew FY, Xu D, Brint EK and O'Neill LA:
Negative regulation of Toll-like receptor-mediated immune
responses. Nat Rev Immunol. 5:446–458. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Aksoy E, Taboubi S, Torres D, Delbauve S,
Hachani A, Whitehead MA, Pearce WP, Berenjeno IM, Nock G, Filloux
A, et al: The p110δ isoform of the kinase PI(3)K controls the
subcellular compartmentalization of TLR4 signaling and protects
from endotoxic shock. Nat Immunol. 13:1045–1054. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cohen P: Immune diseases caused by
mutations in kinases and components of the ubiquitin system. Nat
Immunol. 15:521–529. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lu YC, Yeh WC and Ohashi PS: LPS/TLR4
signal transduction pathway. Cytokine. 42:145–151. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sun SC: Deubiquitylation and regulation of
the immune response. Nat Rev Immunol. 8:501–511. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Katsuoka F and Yamamoto M: Small Maf
proteins (MafF, MafG, MafK): History, structure and function. Gene.
586:197–205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tsuchiya M, Misaka R, Nitta K and Tsuchiya
K: Transcriptional factors, Mafs and their biological roles. World
J Diabetes. 6:175–183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kataoka K: Multiple mechanisms and
functions of maf transcription factors in the regulation of
tissue-specific genes. J Biochem. 141:775–781. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sumiya Y, Ishikawa M, Inoue T, Inui T,
Kuchiike D, Kubo K, Uto Y and Nishikata T: Macrophage activation
mechanisms in human monocytic cell line-derived macrophages.
Anticancer Res. 35:4447–4451. 2015.PubMed/NCBI
|
|
12
|
Sato K, Miyoshi F, Yokota K, Araki Y,
Asanuma Y, Akiyama Y, Yoh K, Takahashi S, Aburatani H and Mimura T:
Marked induction of c-Maf protein during Th17 cell differentiation
and its implication in memory Th cell development. J Biol Chem.
286:14963–14971. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xu J, Yang Y, Qiu G, Lal G, Wu Z, Levy DE,
Ochando JC, Bromberg JS and Ding Y: c-Maf regulates IL-10
expression during Th17 polarization. J Immunol. 182:6226–6236.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Zhang Y, Gu W and Sun B: TH1/TH2
cell differentiation and molecular signals. Adv Exp Med Biol.
841:15–44. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rutz S, Noubade R, Eidenschenk C, Ota N,
Zeng W, Zheng Y, Hackney J, Ding J, Singh H and Ouyang W:
Transcription factor c-Maf mediates the TGF-β-dependent suppression
of IL-22 production in T(H)17 cells. Nat Immunol. 12:1238–1245.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cao S, Liu J, Song L and Ma X: The
protooncogene c-Maf is an essential transcription factor for IL-10
gene expression in macrophages. J Immunol. 174:3484–3492. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma XM and Blenis J: Molecular mechanisms
of mTOR-mediated translational control. Nat Rev Mol Cell Biol.
10:307–318. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Weichhart T, Hengstschläger M and Linke M:
Regulation of innate immune cell function by mTOR. Nat Rev Immunol.
15:599–614. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheng SC, Quintin J, Cramer RA, Shepardson
KM, Saeed S, Kumar V, Giamarellos-Bourboulis EJ, Martens JH, Rao
NA, Aghajanirefah A, et al: mTOR- and HIF-1α-mediated aerobic
glycolysis as metabolic basis for trained immunity. Science.
345:1250684. 2014. View Article : Google Scholar
|
|
20
|
Laplante M and Sabatini DM: mTOR signaling
in growth control and disease. Cell. 149:274–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dazert E and Hall MN: mTOR signaling in
disease. Curr Opin Cell Biol. 23:744–755. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lewis CA, Griffiths B, Santos CR, Pende M
and Schulze A: Regulation of the SREBP transcription factors by
mTORC1. Biochem Soc Trans. 39:495–499. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiao S, Zhang Z, Li C, Huang M, Shi Z,
Wang Y, Song X, Liu H, Li C, Chen M, et al: The kinase MST4 limits
inflammatory responses through direct phosphorylation of the
adaptor TRAF6. Nat Immunol. 16:246–257. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Daassi D, Hamada M, Jeon H, Imamura Y, Nhu
Tran MT and Takahashi S: Differential expression patterns of MafB
and c-Maf in macrophages in vivo and in vitro. Biochem Biophys Res
Commun. 473:118–124. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lucas M, Zhang X, Prasanna V and Mosser
DM: ERK activation following macrophage FcgammaR ligation leads to
chromatin modifications at the IL-10 locus. J Immunol. 175:469–477.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Benkhart EM, Siedlar M, Wedel A, Werner T
and Ziegler-Heitbrock HW: Role of Stat3 in
lipopolysaccharide-induced IL-10 gene expression. J Immunol.
165:1612–1617. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Staples KJ, Smallie T, Williams LM, Foey
A, Burke B, Foxwell BM and Ziegler-Heitbrock L: IL-10 induces IL-10
in primary human monocyte-derived macrophages via the transcription
factor Stat3. J Immunol. 178:4779–4785. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu H, Pardoll D and Jove R: STATs in
cancer inflammation and immunity: A leading role for STAT3. Nat Rev
Cancer. 9:798–809. 2009. View
Article : Google Scholar : PubMed/NCBI
|