Introduction
Rhabdomyosarcoma (RD) is a type of soft tissue
sarcoma that commonly occurs in children and adolescents (1,2).
RD is originated from mesenchymal progenitors of myogenic lineages
(3) and is characterized by
undifferentiated myoblast-like cells (4). Although various therapeutic
strategies have been applied, including intense chemotherapy,
irradiation and surgery, patients with RD exhibit poor prognosis
(5). Thus, effective drugs with
minimal toxicity are urgently needed for RD treatment.
Energy consumption from metabolic activities of
cancer cells depends on glycolysis, even under oxygen-rich
conditions. Phosphofructokinase-1 (PFK-1) serves an important role
in glycolysis, which is activated by fructose 2,6-bisphosphate
(F2,6BP) (6). F2,6BP has been
reported to increase the glucose uptake and allosterically activate
PFK-1 (7,8). In cancer cells, F2,6BP has been
observed to be significantly increased compared with that in normal
tissue cells, indicating that upregulated F2,6BP may contribute to
cell proliferation in cancer tissues (9,10).
PFKFB, also known as
6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase, has been
regarded as a key glycolysis regulator in modulating F2,6BP,
lactate production and glucose uptake (11). PFKFB3, a member of the PFKFB
family, has a significantly high kinase activity, enhances the
F2,6BP level and glycolysis effect (12). The protein levels of PFKFB3 are
commonly elevated in various types of human cancer, including
breast (13), gastric (14) and bladder cancer (15). In addition, previous studies have
demonstrated that the inhibition of PFKFB3 was able to suppress the
glucose metabolism and cell proliferation in cancer cells (16,17). Therefore, strategies of
downregulating PFKFB3 activity may be a promising target for
anticancer therapy.
Targeting PFKFB3 suppression inhibited head and neck
squamous cell carcinoma growth and metastasis (18). In addition, the selective
antagonist of PFKFB3 suppressed the proliferation of the
transplanted tumor in several cells (19,20). PFK15, also known as
1-(4-pyridinyl)-3-(2-quinolinyl)-2-propen-1-one, is a small
molecular PFKFB3 inhibitor that has displayed a powerful activity
against PFKFB3 and anti-tumor effects (21). However, the functions of PFK15 and
the adenosine monophosphate-activated protein kinase (AMPK)
signaling pathway in RD cells remain largely unknown.
In the present study, the effects of PFK15 on the
viability, apoptosis, autophagy and proliferation of RD cells were
investigated. Furthermore, the study explored the possible roles of
the AMPK signaling pathway during these processes.
Materials and methods
Reagents and chemicals
PFK15 (cat. no. SML1009),
3-(3-pyridinyl)-1-(4-pyridinyl)-2-propen-1-one (3PO; cat. no.
407325), 3-bromopyruvate (3BP), 3-methyladenine (3MA), chloroquine
diphosphate salt (CQ; cat. no. C6628) and a polyclonal antibody
against light chain 3 (LC3; cat. no. L7543) were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). An antibody against
t-ERK1/2 (cat.no. 9102), antibody against poly (ADP-ribose)
polymerase 1 (PARP-1; cat. no. 9542), antibody against pAMPK (cat.
no. 2535; Thr 172), antibody against mTOR (cat. no. 2971; Ser2448),
antibody against P-AKT (cat.no. 9721; Ser 473), antibody against
t-Akt (cat. no. T308) and an antibody against phosphorylated
acetyl-CoA carboxylase [pACC (cat. no. 3661; Ser79)] were obtained
from Cell Signaling Technology, Inc. (Danvers, MA, USA). An
antibody against p62 (cat. no. sc-28359) was obtained from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA), while an antibody
against β-actin (cat. no. TA-09) was purchased from OriGene
Technologies, Inc. (Beijing, China). MTS [also known as
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium;
cat. no. G1111] was acquired from Promega Corporation (Madison, WI,
USA). The Alexa Fluor 594 goat anti-rabbit IgG (H+L) antibody (cat.
no. R37117), fetal bovine serum (FBS; Gibco) and enhanced
chemiluminescence reagent (Pierce) were obtained from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA).
5-Aminoimidazole-4-carboxamide-1-β-D-ribofuranoside (AICAR;
Sigma-Aldrich; Merck KGaA; cat. no. A9978), an activator of AMPK.
DAPI (VECTASHIELD® with DAPI; cat. no. H-1200) was
purchased from Fushen Biotechnology Co., Ltd. (Shanghai, China).
3PO was provided by Dr. Y.Q. Huo (Georgia Regents University,
Augusta, GA, USA). The siRNA specific for Autophagy Related 5
(Atg5; cat. no. sc-41445), Autophagy Related 7 (Atg7; cat. no.
sc-41447) was purchased from Santa Cruz Biotechnology, Inc.
together with the control (mock) siRNA (cat. no. sc-37007).
Cell culture and treatments
Human RD (ATCC® CCL-136™) cells were
obtained from the American Type Culture Collection (Manassas, VA,
USA). Cell cultures were maintained in Dulbecco's modified Eagle's
medium (DMEM) (4.5 g/l glucose,) containing 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.) and 1% Penicillin-Streptomycin (100 IU/ml)
and incubated in an atmosphere with 95% humidified air and 5%
CO2 at 37°C. Prior to treatment, the cells were grown to
70–80% confluence. Subsequently, PFK15 was diluted in dimethyl
sulfoxide (DMSO) at different concentrations (2, 4 and 6 mM) and
then added to the cells in growth medium. Cells in the control
group were only treated with DMSO. The concentration of DMSO in
DMEM was 1:2,000. 3PO with concentration of 10 μM was also
used for comparison.
Immunoblotting
Cells were homogenized with lysis buffer containing
Triton X-100/glycerol buffer (1% Triton X-100, 10% glycerol), 50 mM
Tris-HCl (pH 7.4), 1 mM dithiothreitol, 2 mM EGTA and 4 mM EDTA,
supplemented with 1% SDS, and 1% protease inhibitor cocktail (cat.
no. 04693159001; Roche Diagnostics, Basel, Switzerland). Next, the
cells were subjected to 13, 10 or 8% SDS-PAGE and transferred onto
a polyvinylidene difluoride membrane. The membrane was blocked in
5% milk for 1 h at the room temperature. The appropriate primary
antibodies (1:1,000) was incubated for 12 h at 4°C and horseradish
peroxidase-conjugated suitable secondary antibodies (1:5,000) were
used for immunoblotting. The secondary antibodies were incubated
for 1 h at room temperature. Subsequently, the blots were detected
by enhanced chemiluminescence. Several X-ray films were analyzed to
verify the linear range of the chemiluminescence signals, and the
densitometry was performed using ImageJ software version 1.46
(National Institutes of Health, Bethesda, MD, USA).
Cell viability
For viability analysis, cells in the PFK15-treated
and control groups were seeded into a 96-well plate at a density of
5,000–10,000 cells per well and allowed to adhere by culturing at
37°C overnight. For dose-dependent experiments, PFK15 was added in
increasing concentrations (2, 4 and 6 μM) and incubated for
24 h. For time-dependent experiments, PFK15 (6 μM) or 3BP
(25 μM) was added and incubated for 12, 24 or 48 h. For
synergistic effect analysis, PFK15 (4 μM) combined with CQ
(10 μM) or 3MA (1 mM) were added to the cells and incubated
for 48 h. Subsequent to these different treatments, 0.8 mg/ml MTS
was added to the samples and incubated for 3 h. The absorbance of
samples were then read at 492nm using a microtiter plate
spectrophotometer (BioTek Instruments, Inc., Winooski, VT,
USA).
Colony growth assay
For colony growth assay, cells were seeded in a 6 cm
dish at a concentration of 300 cells/ml, PFK15-treated and cultured
for 2 weeks. To examine the association of AMPK activity with
autophagy and cell death caused by PFK15 in RD cells, the AMPK
agonist AICAR (100 μM) was used in subsequent experiments.
Next, cells were fixed in paraformaldehyde (4%) and stained by
trypan blue. The numbers of colonies in each sample were counted by
ImageJ software version 1.46 (National Institutes of Health,
Bethesda, MD, USA).
Fluorescence microscopy
Cells were fixed with paraformaldehyde (4%) for 12
min at room temperature and then examined by fluorescence
microscopy. Briefly, cells were blocked by PBS containing 3% bovine
serum albumin and 0.2% Triton X-100 for 1 h. Following washing
three times with PBS, the cells were incubated with the LC3
antibody (1:200) for 2 h at room temperature, washed with PBS for
three times, and then incubated with Alexa Fluor 594 goat
anti-rabbit IgG (H+L) antibody for 2 h at room temperature. The
nuclei were subsequently counterstained with DAPI/Antifade and
observed using a fluorescence microscope system. The number of the
punctate LC3 structures in each cell was counted, and at least 30
cells were included for each group in the immunofluorescent
assay.
Transmission electron microscopy
Electron microscopy was conducted as described
previously (22). Briefly, cells
were initially treated with DMSO (control group) or PFK15 6
μM (treatment group). Samples were then washed, trypsinized
and collected by centrifugation at 1,500 × g, at 4°C for 5 min.
Next, cells were fixed with 4% paraformaldehyde overnight and
post-fixed with 1% OsO4 in cacodylate buffer at room
temperature for 1 h, followed by stepwise dehydration with ethanol.
The dehydrated pellets were subsequently washed with propylene
oxide for 30 min at room temperature and embedded in Spurr resin
for sectioning. A transmission electron microscope (JEM1230; JEOL,
Ltd., Tokyo, Japan) was used for observation of the thin sections.
Morphometric analysis of the area fractions between autophagosomes
and whole cells were calculated using ImageJ software, and the data
were presented in a histogram graph.
Lactate assay
Cells were seeded in 6-well plates, after
PFK15-treated, the suspension was collected and subjected to
lactate assay. Levels of the secreted lactate were detected using
the lactate assay kit (K-DATE, Megazyme, Bray, Co. Wicklow,
Ireland). Data represent three independent experiments.
Statistical analysis
GraphPad Prism version 5 software (GraphPad
Software, Inc., La Jolla, CA, USA) was used for statistical
analyses. All data are expressed as the mean ± standard deviation.
Student's t-test was used to evaluate the statistical significant
differences between two groups. Multigroup comparisons of the means
were conducted by one-way analysis of variance with a post hoc
Student-Newman-Keuls test. A statistically significant difference
was regarded when the P-value was <0.05. Experiments were
repeated for a minimum of three times for each condition.
Results
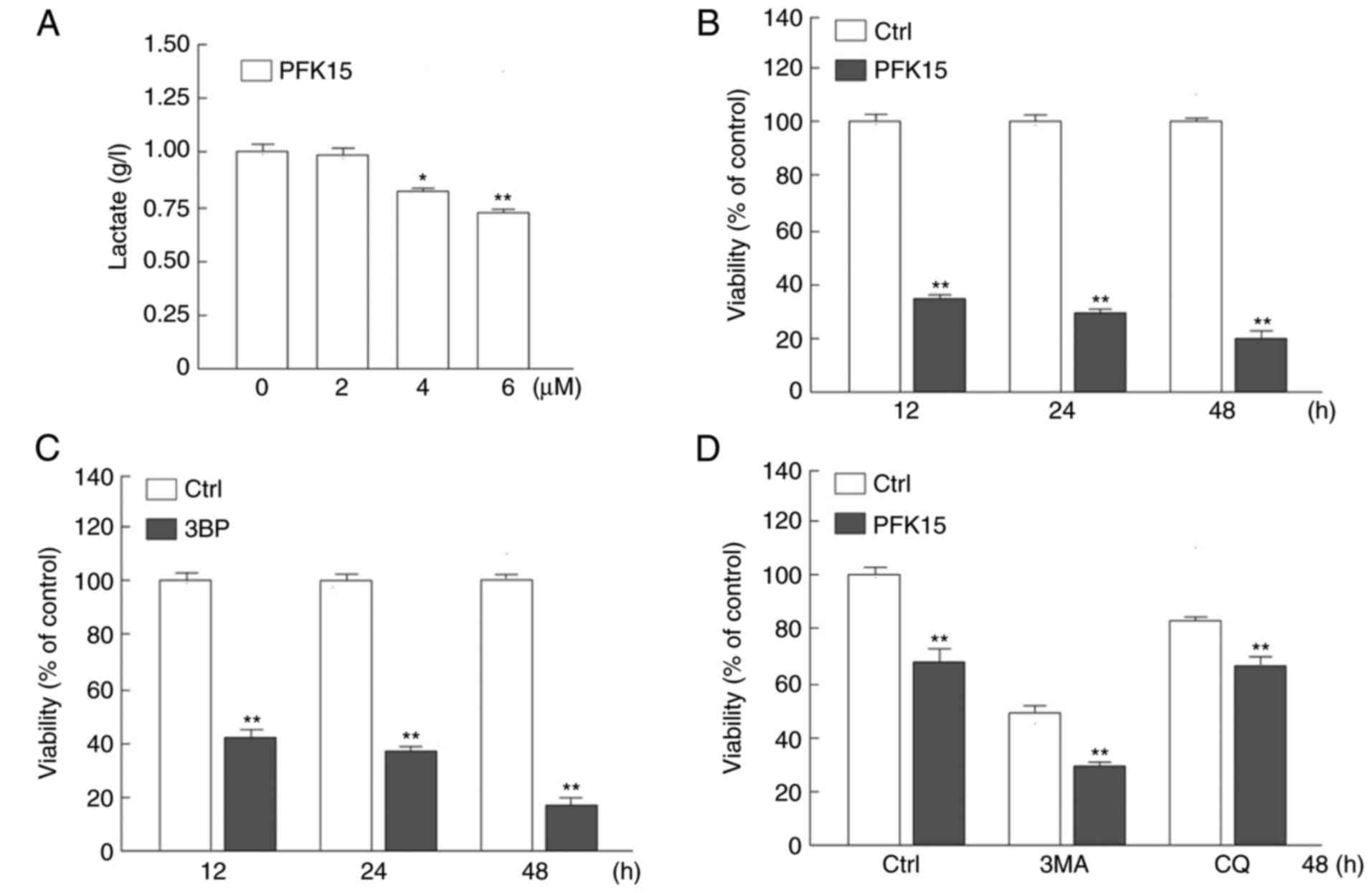
PFK15 suppresses the viability of RD
cells
As a potent small molecule antagonist of PFKFB3,
PFK15 has been verified to inhibit the proliferation and viability
of various cancer cells at low concentrations (14,21). In the present study, it was
observed that PFK15 significantly suppressed the glycolysis in RD
cells by monitoring the final product lactate, and the inhibitory
effect of PFK15 was dose-dependent. Treatment with 4 and 6
μM PFK15 significantly attenuated the lactate production by
20 and 27%, respectively, as compared with the control group (4
μM vs. control: Mean difference (MD)=−0.2, q=6.928,
P<0.05; 6 μM vs. control: MD=−0.3, q=10.39, P<0.01;
Fig. 1A). Thus, the concentration
of PFK15 was selected at 6 μM for the following
experiments.
The cell viability was then detected by an MTS assay
in RD cells following PFK15 treatment for different time periods.
At the 12-h time point, PFK15 evidently decreased the viability of
RD cells compared with the control group (P<0.01; Fig. 1B). With longer treatment time, the
survival rate of RD cells decreased from ~31% at 24 h to 19% at 48
h compared with that in the control group (P<0.01; Fig. 1B).
3BP is an inhibitor of key enzyme hexokinase 2 in
glycolysis and is also an inhibitor of phosphatidylinositol
3-kinase, which is widely used as an inhibitor of autophagy
(23). 3BP inhibits the activity
of PI3K, which is a known negative regulator of autophagy.
Theoretically 3BP could promotes autophagy, however, our un-shown
data indicated that 3BP inhibited autophagy. In addition, CQ is a
known inhibitor of autolysosome degradation that is widely used in
monitoring the autophagic flux. In the current study, after 12 h of
treatment with 3BP (25 μM), the cell viability was
significantly reduced as compared with the control group (Fig. 1C). Furthermore, when treated with
PFK15 and 3MA (1 mM) but not CQ, the cell viability was further
decreased as a result of a synergistic effect (Fig. 1D). One single drug treatment can
induce various types of cell death, thus PFK15 may cause apoptotic
and autophagic cell death, as well as other types of cell death.
However, as the autophagy initiation inhibitor 3MA further
increased the cell viability loss, it is suggested that autophagy
may serve a protective role upon PFK15 treatment. As inhibition of
autophagy exacerbated the cell viability loss upon PFK15 treatment,
it was hypothesized that autophagy may in part ameliorate the
PFK15-induced cell viability loss. Furthermore, although widely
used as an inhibitor of autophagic flux, CQ functions during the
autolysosome fusion and degradation processes, which may be reason
for the distinct phenotype between CQ and 3MA on PFK15-induced cell
viability loss in Fig. 1D. These
results indicated that PFK15 was able to decrease RD cell
autophagy.
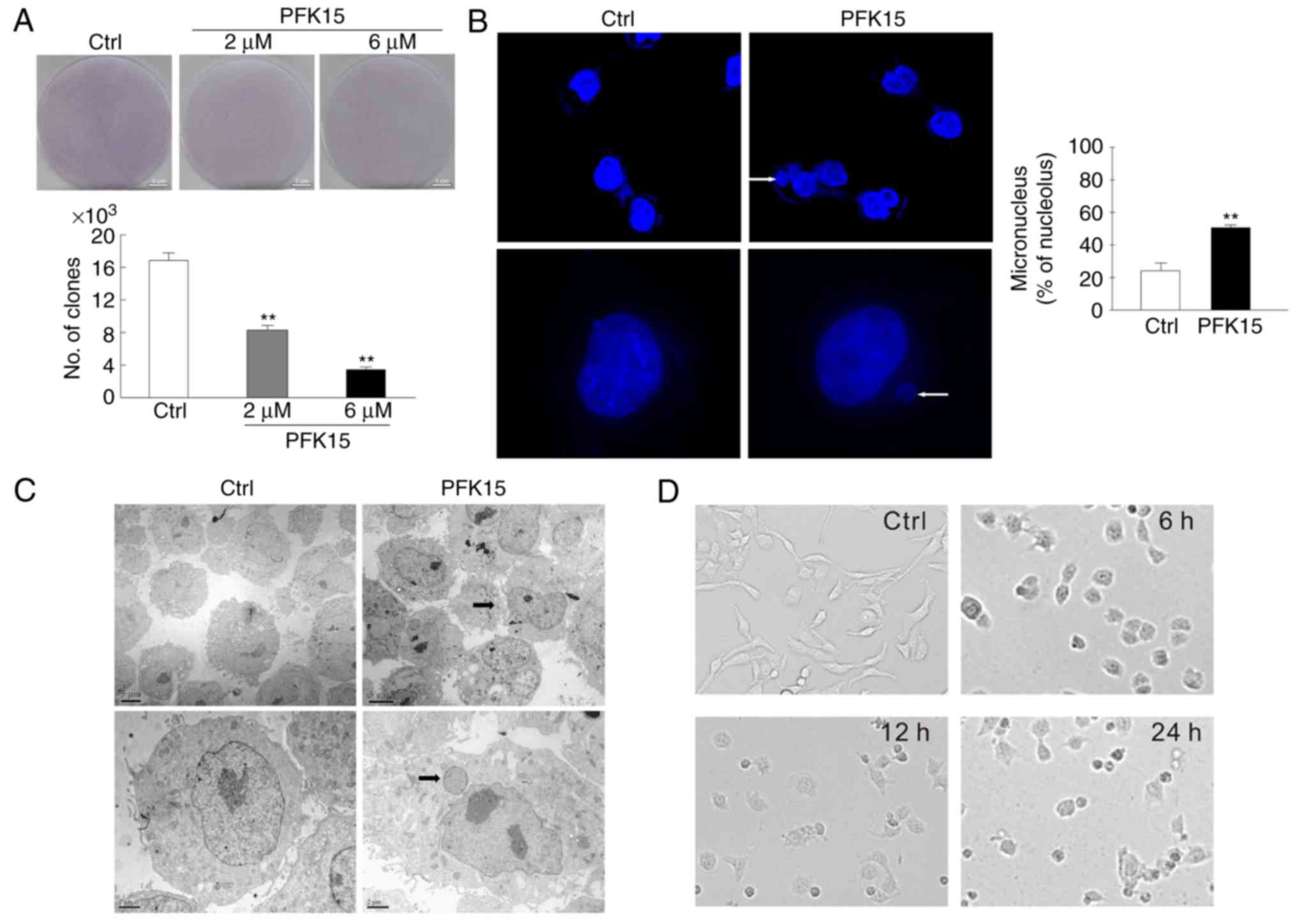
PFK15 inhibits clone formation and causes
abnormal nuclear morphology
A colony formation assay was performed to further
analyze the effect of PFK15 on the cell proliferation. As a result,
the number of clones were significantly reduced in the 2 and 6
μM PFK15 treatment groups as compared with the controls (6
μM vs. control: MD=−13, q=22.52, P<0.01; 2 μM vs.
control: MD=−9, q=15.59, P<0.01; Fig. 2A). Furthermore, the nuclear
component was distinct from the primary nuclei in RD cells treated
with 6 μM PFK15 for 2 h, as observed by immunofluorescence
microscopy (Fig. 2B) and
transmission electron microscopy (Fig. 2C). Vehicle control cells exhibited
a wavy plasma membrane edge, which is a characteristic of normal
morphology. By contrast, the integrity of the nuclear membrane of
PFK15-treated cells disappeared, while protuberance was detected in
the nuclear membrane. The nucleus condensed and the chromosomes
gathered to assemble chromatin. Gradually, the protuberance was
separated from nucleus and formed micronuclei, which subsisted in
the cytoplasm. Compared with the control group, an evident
accumulation of micronuclei was observed in PFK15-treated RD cells
(P<0.01; Fig. 2B). Similar
results were observed by transmission electron microscopy (Fig. 2C). The cells were spindle-shaped
and well-attached to the culture plate in the control group, while
the PFK15-treated cells were loosely attached, rounded and randomly
oriented. Following PFK15 treatment for 24 h, the cells exhibited
reduction in size, karyorrhexis and even death (Fig. 2D).
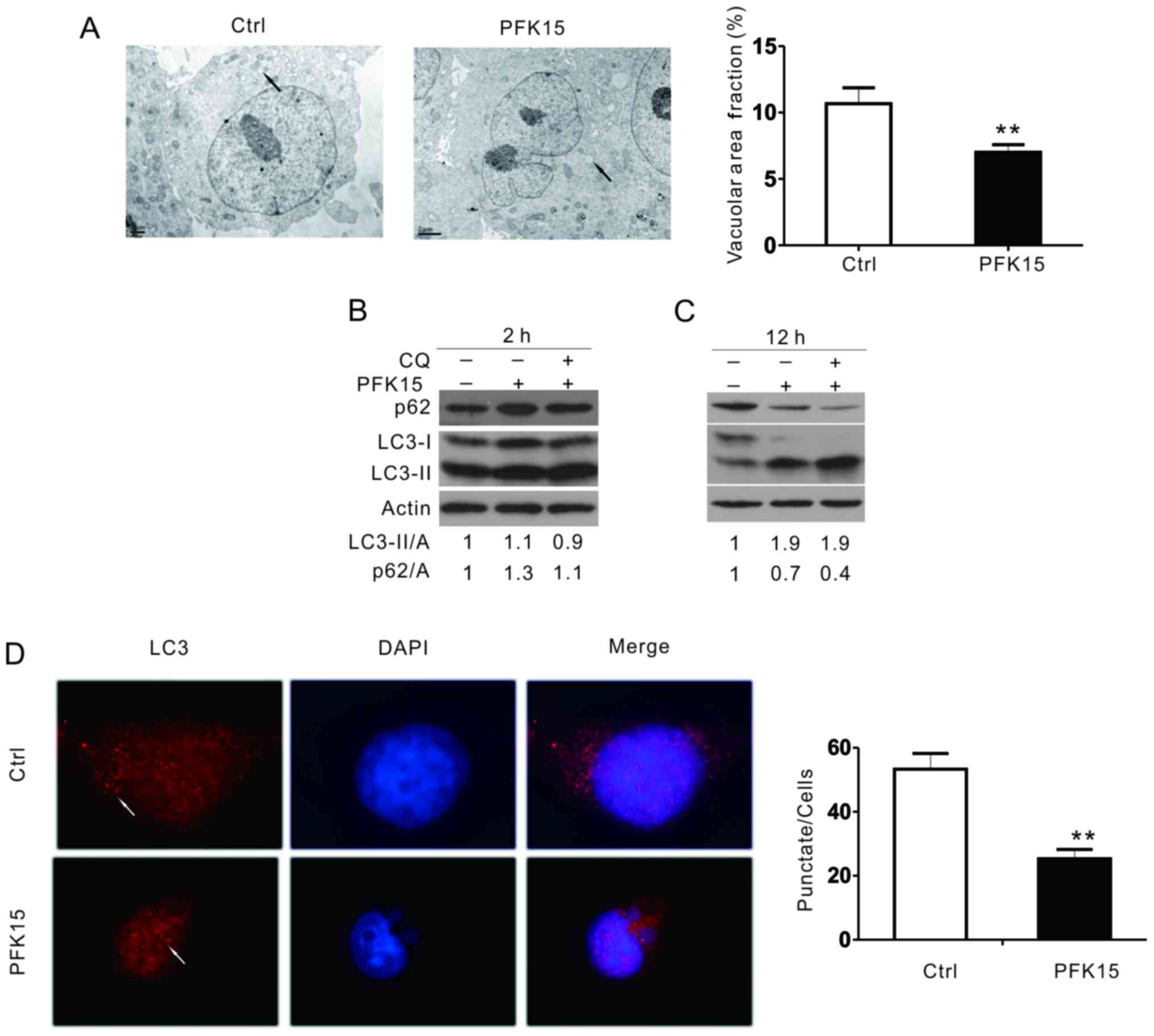
PFK15 inhibits autophagy in RD cells
Transmission electron microscopy, considered as one
of the most convincing approaches to detect autophagosome formation
(24,25), was used in the current study to
determine the effect of PFK15 on autophagy. Compared with the DMSO
control, PFK15 markedly attenuated the accumulation of membrane
vacuoles (Fig. 3A). When
autophagy occurs, LC3-I can be converted into its
phosphatidylethanolamine-conjugated form, LC3-II, which is
associated with autophagosomes. As observed in the immunoblotting
assay, PFK15 increased the level of LC3-II and p62 after 2 h of
treatment in RD cells (Fig. 3B).
After 12 h of treatment, PFK15 further increased the LC3-II level,
whereas it decreased the p62 level (Fig. 3C). However, combined incubation
with CQ, a known inhibitor of autolysosome degradation that is
widely used in monitoring autophagic flux, failed to further
increase the level of LC3-II and block p62 degradation in the
PFK15-treated cells, suggesting that PFK15 inhibited the autophagic
flux in RD cells. An immunofluorescence assay was also performed to
monitor the aggregation of LC3 in RD cells. Consistently, the
punctate of LC3 was significantly decreased when cells were treated
with PFK15 (Fig. 3D).
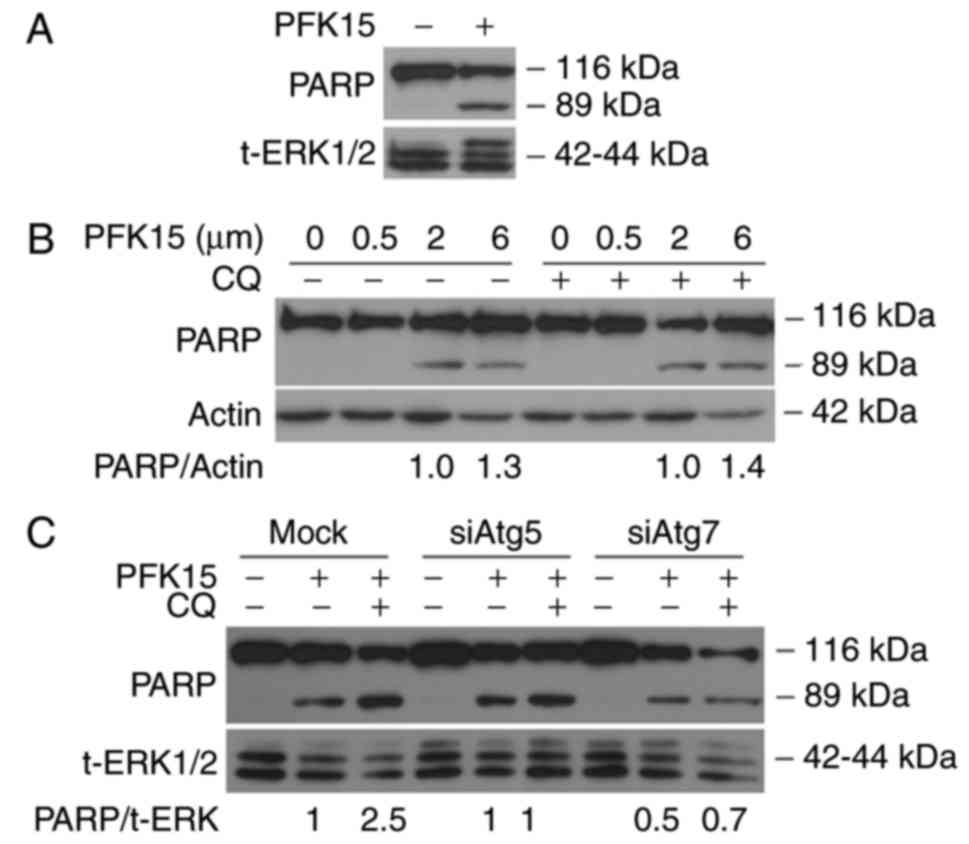
PFK15 induces the cleavage of PARP-1
PARP-1 has been reported to be involved in several
cellular processes, including cell autophagy and apoptosis, and the
cleavage of PARP-1 has been used as a marker of caspase-dependent
apoptosis (26). In the current
study, immunoblotting analysis was performed to evaluate the effect
of PFK15 on PARP-1, and the results revealed that PFK15 induced the
cleavage of PARP-1 in a dose-dependent manner (Fig. 4A and B), indicating that PFK15
activated the apoptotic pathway. However, addition of CQ did not
significantly induce cleavage of PARP-1 in RD cells (Fig. 4B). The autophagy-associated
proteins Atg5 and Atg7 serve important roles in the maturation of
autophagosomes by promoting the elongation of phagophores (27). In order to further analyze the
association between cell autophagy and apoptosis caused by PFK15,
an autophagy inhibition model was constructed by silencing the Atg5
and Atg7 genes. The results demonstrated that the cleavage of
PARP-1 was decreased when Atg5 and Atg7 were knocked down,
indicating that the cell apoptosis caused by PFK15 was suppressed
by the inhibition of earlier autophagy (Fig. 4C).
PFK15 downregulates the activity of AMPK,
pAKT and mammalian target of rapamycin complex 1 (mTORC1)
mTORC1 is a protein complexes, in which mTOR is the
core component. The p-mTOR (S2448) could reflect the activity of
mTORC1. mTORC1 is usually known as a negative regulator of
autophagy. AMPK, a sensor for intracellular adenosine nucleotides,
responds to alterations in energy status, and coordinates the cell
growth, autophagy and metabolism (28). Since it is generally
phosphorylation of PFKFB3 on Ser461, the present study attempted to
clarify whether AMPK serves a regulatory role in the PFK15-mediated
effects in RD cells (29). The
results demonstrated that, after 2-h treatment, PFK15 suppressed
the level of phosphorylated AMPK (pAMPK) and its substrate pACC by
44 and 20%, respectively. Similar trends were also identified at
the 4 and 12 h time points (Fig.
5A). At the 12 h time point, PFK15 had a significant inhibitory
effect on pAMPK and pACC, reducing their levels by 88 and 61%,
respectively. Similar results were also obtained upon treatment
with 3PO, another inhibitor of PFKFB3. Inhibition of PFKFB3 using
3PO was successful in reducing cancer growth and increasing
apoptosis, but only at certain time points within the circadian
cycle (30). Considering that the
decreased glycolysis caused by PFK15 failed to activated the AMPK
activity in a feedback loop, whereas it decreased the levels of
pAMPK and pACC, it is suggested that PFKFB3 may function upstream
of AMPK under certain circumstance. The activity of AKT, which is
another negative regulator of autophagy, was also investigated in
the present study. It was observed that PFK15 evidently upregulated
the pAKT (Ser473) level, which inhibits autophagy by activating
mTORC1 (Fig. 5B). As glycolysis
is closely associated with the energy sensor AMPK, the present
study only focused on the AMPK related data; other data will be
reported in future studies.
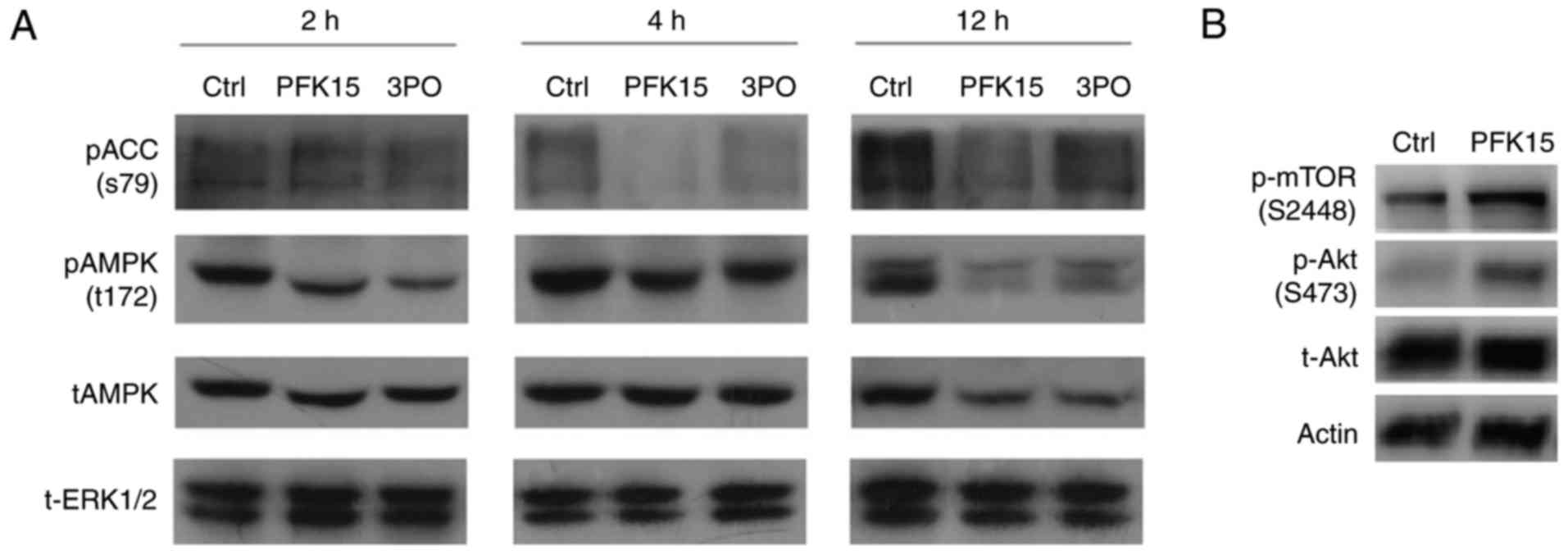 | Figure 5PFK15 regulates AMPK proteins
expression. (A) RD cells were treated for 2, 4 and 12 h with 6
μM PFK15, and western blot analysis was conducted to examine
the levels of t-AMPK, p-AMPK and p-ACC proteins. Total ERK was used
as a loading control. (B) Immunoblotting assay was performed to
determine the p-mTOR, p-AKT and t-AKT levels in cells treated with
PFK15 (6 μM). RD, rhabdomyosarcoma; AMPK, adenosine
monophosphate-activated protein kinase; AKT, protein kinase B; ACC,
acetyl-CoA carboxylase; ERK, extracellular signal-regulated kinase;
mTOR, mammalian target of rapamycin; p-, phosphorylated; t-, total;
Ctrl, control. |
AICAR rescues the downregulation of PFK15
on autophagy and activity of AMPK
It was observed that, compared with PFK15 treatment
alone, the combination of AICAR and PFK15 decreased LC3-II
expression with CQ treatment and decreased p62 degradation at the 2
and 12 h time points. Furthermore, CQ addition further increased
the LC3-II level, as well as blocked the p62 degradation,
indicating a complete autophagic flux (Fig. 6A). Thus, it is likely that AICAR
rescued the PFK15-dependent inhibition of autophagic flux in RD
cells. Subsequently, the AMPK activity was detected following the
indicated treatment in RD cells. Addition of AICAR inhibited the
downregulation of AMPK phosphorylation at the 2 and 12 h time
points (Fig. 6B). This
consistency of AMPK activity and autophagy supported that PFK15
inhibited autophagy by decreasing AMPK signaling. The current study
also examined the effect of AICAR on PFK15-dependent cell death,
and the results demonstrated that pre-treatment with AICAR provided
protection against PFK15-induced cell death at the 24 and 48 h time
points (Fig. 6C). These findings
indicated that PFK15 inhibited the cell growth, while AICAR partly
recovered the cell growth (PFK15 vs. DMSO: Mean diff.= −13,
q=14.16, P<0.01; PFK15+AICAR vs. DMSO: Mean diff.= −6.667,
q=7.263, P<0.01; Fig. 6D).
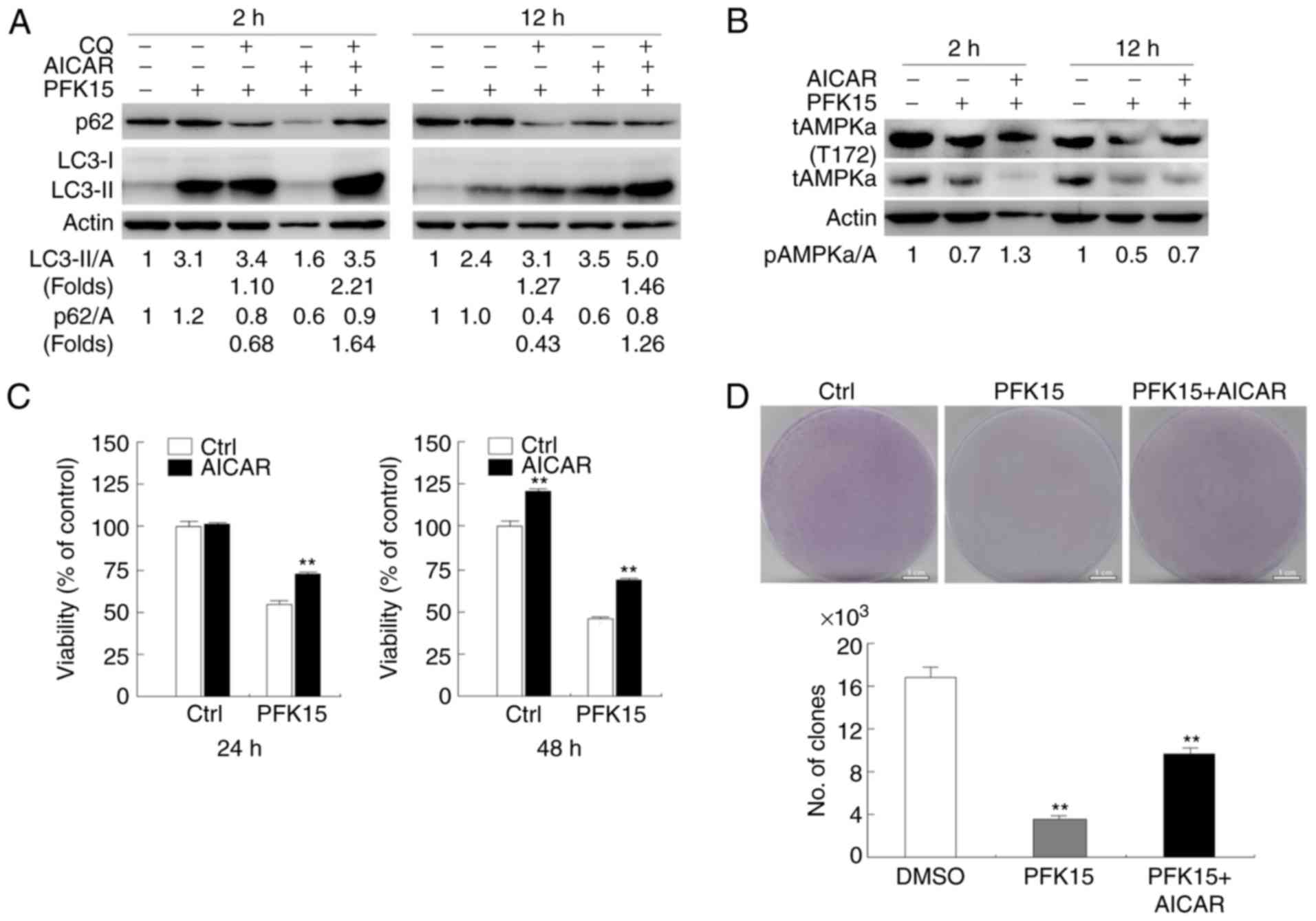 | Figure 6AICAR attenuates PFK15-induced
suppression of autophagy and cell viability loss, partly blocking
the decrease of p-AMPK. (A) Western blots analyzing the expression
levels of p62 and LC3 in RD cells treated with AICAR and PFK15 in
the presence or absence of CQ (10 μM). (B) Effect of AICAR
(0.5 mM) treatment for 2 or 12 h on AMPK activity in the absence
and presence of PFK15 (6 μM), as reflected by the
phosphorylation of AMPK. (C) RD cells were treated with PFK15 and
AICAR for 24 and 48 h, and cell viability was analyzed by an MTS
assay. (D) Colony growth assays were performed in RD cells treated
with PFK15 (6 μM) in the absence and presence of AICAR. RD,
rhabdomyosarcoma; AMPK, adenosine monophosphate-activated protein
kinase; *P<0.05 vs. control and
**P<0.01 vs. control. AICAR,
5-aminoimidazole-4-carboxamide-1-β-D-ribofuranoside; CQ,
chloroquine diphosphate salt; LC3, light chain 3; p-,
phosphorylated; t-, total; Ctrl, control. |
Discussion
Currently, the deregulation of cellular energetics
by anti-glycolysis is a novel strategy in the treatment of cancer
cells, which has attracted much attention (31). As an antagonist of PFKFB3, PFK15
has been suggested to suppress the tumor cell viability by
inhibiting the glycolysis (21).
However, the underlying pharmacological mechanism is not yet
clearly understood. In the present study, the appearance of cleaved
PARP-1 and micronucleus protuberance was different in PFK15-treated
cells compared with control group, indicating that PFK15 inhibited
RD cell viability by inducing apoptosis.
Autophagy, the 'self-eating' system (32), is known as one of the most
important systems in solid tumor. In cancer, autophagy-mediated
recycling can be used to maintain energy homeostasis and
mitochondrial function, for providing elevated metabolic demand of
growth and proliferation (33).
At the beginning of autophagy, the isolation membrane is developed
to package the organelles and other materials in the cytoplasm to
form autophagosomes. During this process, LC3 is cleaved into a
soluble form known as LC3-I, which is then modified into a
membrane-bound form, termed LC3-II (34). At the second step, the packaged
cell organelles are digested by lysosomal enzymes, while LC3 and
the other autophagosomes are digested. In the present study,
treatment with PFK15 increased the LC3-II level in the RD cells.
However, the autophagy inhibitor CQ did not further increase the
PFK15-induced LC3-II level and failed to block the degradation of
p62, suggesting that PFK15 inhibited the second step of autophagy,
in which the accumulated LC3-II was not digested by lysosomal
enzymes.
PARP-1 is a nuclear enzyme that catalyzes the
transfer of ADP-ribose polymers onto itself and other proteins in
response to DNA strand breaks (35). It is one of the main cleavage
targets of caspase-3 in vivo (36), and cleavage of PARP-1 serves as a
marker of cells undergoing apoptosis (37). During apoptosis, PARP-1 breaks
into two fragments (89 and 24 kDa), which is a useful hallmark in
cell death (38). In the present
study, it was observed that PFK15 induced the cleavage of PARP-1 in
RD cells, indicating that PFK15 activated the apoptotic pathway.
There is a close connection between autophagy and apoptosis, since
autophagy is able to promote, suppress and accompany apoptosis. The
current study observed that inhibition of autophagy by silencing
Atg5 and Atg7 attenuated the PFK15-induced caspase-dependent
apoptosis, particularly by silencing Atg7. Along with the promoting
effect of 3MA on PFK15-induced cell viability loss, these results
indicated that PFK15 induced multi-type cell death other than
caspase-dependent apoptosis. In addition, the distinct effect of CQ
on PFK15-induced PARP-1 cleavage between siRNA knockdown mock group
and PFK15 treated group (Fig. 4B)
may be due to siRNA transfection altering the cell state via an
uncertain mechanism. As demonstrated in the present study, there
was crosstalk between autophagy and apoptosis. Xi et al
(39) reported that, in RD cells,
inhibition of autophagosomes at the stage of autophagosome and
lysosome fusion promoted apoptosis. Notably, another previous study
confirmed that induction of autophagy was a useful therapeutic
approach for overcoming drug resistance to certain therapeutic
agents, particularly those that typically induce an apoptotic
response (40). Consistent with
the results of the present study, PARP serves an important function
in the crosstalk between autophagy and apoptosis. Furthermore,
PFK15-induced apoptosis was suppressed by autophagy inhibition.
In addition, the present results indicated that
PFK15-induced cell death was mediated by AMPK; however, AICAR was
able to attenuate this effect. As an evolutionary conserved
fuel-sensing enzyme, AMPK is activated in shortage of energy and
suppressed by its surfeit (41).
Previous studies have demonstrated that AMPK serves a dual role in
cancer (42). Under some
conditions, activation of AMPK signaling inhibited cancer cell
growth and tumorigenesis (43,44). Convincing evidence has accumulated
indicating that AMPK signaling is a conditional tumor suppressor
pathway (45,46). Hadad et al (47) have also reported that reduced
pAMPK and pACC signals were inversely correlated with the
histological grade, as well as axillary node metastasis in breast
cancer. However, in certain cancer cells, AMPK downregulation is
beneficial to therapy, while with the administration of
pharmacological activators of AMPK the antineoplastic effect
disappears or is decreased (42,48). In the present study, it was
observed that PFK15 suppressed the levels of pAMPK and pACC at
different treated times, and upon treatment with AICAR, an agonist
of AMPK, the RD cell activity and autophagic flux were partially
recovered. Taken together, these findings suggested that PFK15 was
able to inhibit autophagy and cell viability through the AMPK
signaling pathway.
There are also certain limitations to the present
study. Due to the limitation of experimental conditions,
experiments involving an in vivo xenograft model were not
performed In the future, PFK15-induced inhibition of autophagy and
proliferation in an in vivo xenograft model will be
researched.
In conclusion, the present study provided novel
insights into the antitumor activity of PFK15 in RD cells. PFK15
inhibited autophagy and cell viability through AMPK signaling, and
AMPK functioned downstream of PFKFB3. These findings may provide a
theoretical basis for the use of PFKFB3 as a target for the
clinical treatment of RD.
Acknowledgments
Not applicable.
References
|
1
|
Huh WW and Skapek SX: Childhood
rhabdomyosarcoma: New insight on biology and treatment. Curr Oncol
Rep. 12:402–410. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Leaphart C and Rodeberg D: Pediatric
surgical oncology: Management of rhabdomyosarcoma. Surg Oncol.
16:173–185. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xia SJ, Pressey JG and Barr FG: Molecular
pathogenesis of rhabdomyosarcoma. Cancer Biol Ther. 1:97–104. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Keller C and Guttridge DC: Mechanisms of
impaired differentiation in rhabdomyosarcoma. FEBS J.
280:4323–4334. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Malempati S and Hawkins DS:
Rhabdomyosarcoma: Review of the Children's Oncology Group (COG)
soft-tissue sarcoma committee experience and rationale for current
COG studies. Pediatr Blood Cancer. 59:5–10. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Van Schaftingen E, Jett MF, Hue L and Hers
HG: Control of liver 6-phosphofructokinase by fructose
2,6-bisphosphate and other effectors. Proc Natl Acad Sci USA.
78:3483–3486. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Atsumi T, Chesney J, Metz C, Leng L,
Donnelly S, Makita Z, Mitchell R and Bucala R: High expression of
inducible 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase
(iPFK-2; PFKFB3) in human cancers. Cancer Res. 62:5881–5887.
2002.PubMed/NCBI
|
|
8
|
Van Schaftingen E, Hue L and Hers HG:
Fructose 2,6-bisphosphate, the probably structure of the glucose-
and glucagon-sensitive stimulator of phosphofructokinase. Biochem
J. 192:897–901. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Miralpeix M, Azcon-Bieto J, Bartrons R and
Argiles JM: The impairment of respiration by glycolysis in the
Lewis lung carcinoma. Cancer Lett. 50:173–178. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bando H, Atsumi T, Nishio T, Niwa H,
Mishima S, Shimizu C, Yoshioka N, Bucala R and Koike T:
Phosphorylation of the 6-phosphofructo-2-kinase/fructose
2,6-bisphosphatase/PFKFB3 family of glycolytic regulators in human
cancer. Clin Cancer Res. 11:5784–5792. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ros S and Schulze A: Balancing glycolytic
flux: The role of 6-phosphofructo-2-kinase/fructose
2,6-bisphosphatases in cancer metabolism. Cancer Metab. 1:82013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yalcin A, Telang S, Clem B and Chesney J:
Regulation of glucose metabolism by
6-phosphofructo-2-kinase/fructose-2, 6-bisphosphatases in cancer.
Exp Mol Pathol. 86:174–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ge X, Lyu P, Gu Y, Li L, Li J, Wang Y,
Zhang L, Fu C and Cao Z: Sonic hedgehog stimulates glycolysis and
proliferation of breast cancer cells: Modulation of PFKFB3
activation. Biochem Biophys Res Commun. 464:862–868. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu W, Ye L, Zhang J, Yu P, Wang H, Ye Z
and Tian J: PFK15, a small molecule inhibitor of PFKFB3 induces
cell cycle arrest, apoptosis and inhibits invasion in gastric
cancer. PLoS One. 11:e01637682016. View Article : Google Scholar
|
|
15
|
Hu KY, Wang de G, Liu PF, Cao YW, Wang YH,
Yang XC, Hu CX, Sun LJ and Niu HT: Targeting of MCT1 and PFKFB3
influences cell proliferation and apoptosis in bladder cancer by
altering the tumor microenvironment. Oncol Rep. 36:945–951. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
O'Neal J, Clem A, Reynolds L, Dougherty S,
Imbert-Fernandez Y, Telang S, Chesney J and Clem BF: Inhibition of
6-phosphofructo-2-kinase (PFKFB3) suppresses glucose metabolism and
the growth of HER2+ breast cancer. Breast Cancer Res Treat.
160:29–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Feng Y and Wu L: mTOR up-regulation of
PFKFB3 is essential for acute myeloid leukemia cell survival.
Biochem Biophys Res Commun. 483:897–903. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li HM, Yang JG, Liu ZJ, Wang WM, Yu ZL,
Ren JG, Chen G, Zhang W and Jia J: Blockage of glycolysis by
targeting PFKFB3 suppresses tumor growth and metastasis in head and
neck squamous cell carcinoma. J Exp Clin Cancer Res. 36:72017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Calvo MN, Bartrons R, Castaño E, Perales
J, Navarro-Sabaté A and Manzano A: PFKFB3 gene silencing decreases
glycolysis, induces cell-cycle delay and inhibits
anchorage-independent growth in HeLa cells. FEBS Lett.
580:3308–3314. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lea MA, Guzman Y and Desbordes C:
Inhibition of growth by combined treatment with inhibitors of
lactate dehydrogenase and either phenformin or inhibitors of
6-phosphofructo-2-kinase/fructose-2,6-bis phosphatase 3. Anticancer
Res. 36:1479–1488. 2016.PubMed/NCBI
|
|
21
|
Clem BF, O'Neal J, Tapolsky G, Clem AL,
Imbert-Fernandez Y, Kerr DA II, Klarer AC, Redman R, Miller DM,
Trent JO, et al: Targeting 6-phosphofructo-2-kinase (PFKFB3) as a
therapeutic strategy against cancer. Mol Cancer Ther. 12:1461–1470.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Scagliusi SM, Jorge V and Hugo K:
Cytopathology of callus cells infected with grapevine
leafroll-associated virus 3. Trop Plant Pathol. 27:384–388.
2002.
|
|
23
|
Liu D, Yang Y, Liu Q and Wang J:
Inhibition of autophagy by 3-MA potentiates cisplatin-induced
apoptosis in esophageal squamous cell carcinoma cells. Med Oncol.
28:105–111. 2011. View Article : Google Scholar
|
|
24
|
Lucocq JM and Hacker C: Cutting a fine
figure: On the use of thin sections in electron microscopy to
quantify autophagy. Autophagy. 9:1443–1448. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD,
Adeli K, et al: Guidelines for the use and interpretation of assays
for monitoring autophagy (3rd edition). Autophagy. 12:1–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Muñozgámez JA, Rodríguezvargas JM,
Quilespérez R, Aguilar-Quesada R, Martín-Oliva D, de Murcia G,
Menissier de Murcia J, Almendros A, Ruiz de Almodóvar M and Oliver
FJ: PARP-1 is involved in autophagy induced by DNA damage.
Autophagy. 5:61–74. 2009. View Article : Google Scholar
|
|
27
|
Pham DL, Kim SH, Losol P, Yang EM, Shin
YS, Ye YM and Park HS: Association of autophagy related gene
polymorphisms with neutrophilic airway inflammation in adult
asthma. Korean J Intern Med. 31:375–385. 2016. View Article : Google Scholar :
|
|
28
|
Carling D: The AMP-activated protein
kinase cascade-a unifying system for energy control. Trends Biochem
Sci. 29:18–24. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Novellasdemunt L, Obach M, Millán-Ariño L,
Manzano A, Ventura F, Rosa JL, Jordan A, Navarro-Sabate A and
Bartrons R: Progestins activate
6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3 (PFKFB3) in
breast cancer cells. Biochem J. 442:345–356. 2012. View Article : Google Scholar
|
|
30
|
Telang S, Clem BF, Klarer AC, Clem AL,
Trent JO, Bucala R and Chesney J: Small molecule inhibition of
6-phosphofructo-2-kinase suppresses t cell activation. J Transl
Med. 10:952012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mizushima N and Komatsu M: Autophagy:
Renovation of cells and tissues. Cell. 147:728–741. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
White E, Mehnert JM and Chan CS:
Autophagy, metabolism, and cancer. Clin Cancer Res. 21:5037–5046.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim DK, Yang JS, Maiti K, Hwang JI, Kim K,
Seen D, Ahn Y, Lee C, Kang BC, Kwon HB, et al: A
gonadotropin-releasing hormone-II antagonist induces autophagy of
prostate cancer cells. Cancer Res. 69:923–931. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
D'Amours D, Desnoyers S, D'Silva I and
Poirier GG: Poly(ADP-ribosyl)ation reactions in the regulation of
nuclear functions. Biochem J. 342:249–268. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Andrabi SA, Kim NS, Yu SW, Wang H, Koh DW,
Sasaki M, Klaus JA, Otsuka T, Zhang Z, Koehler RC, et al:
Poly(ADP-ribose) (PAR) polymer is a death signal. Proc Natl Acad
Sci USA. 103:18308–18313. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Amé JC, Spenlehauer C and de Murcia G: The
PARP super-family. Bioessays. 26:882–893. 2004. View Article : Google Scholar
|
|
38
|
Kaufmann SH, Desnoyers S, Ottaviano Y,
Davidson NE and Poirier GG: Specific proteolytic cleavage of
poly(ADP-ribose) polymerase: An early marker of
chemotherapy-induced apoptosis. Cancer Res. 53:3976–3985.
1993.PubMed/NCBI
|
|
39
|
Xi X, Zhang X, Wang B, Wang T, Wang J,
Huang H, Wang J, Jin Q and Zhao Z: The interplays between autophagy
and apoptosis induced by enterovirus 71. PLoS One. 8:e569662013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yokoyama T, Kondo Y, Bögler O and Kondo S:
The role of autophagy and apoptosis in the drug resistance of
cancer. Springer US, Drug Resistance in Cancer Cells. 53–71. 2009.
View Article : Google Scholar
|
|
41
|
Luo Z, Zang M and Guo W: AMPK as a
metabolic tumor suppressor: Control of metabolism and cell growth.
Future Oncol. 6:457–470. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Okoshi R, Ozaki T, Yamamoto H, Ando K,
Koida N, Ono S, Koda T, Kamijo T, Nakagawara A and Kizaki H:
Activation of AMP-activated protein kinase induces p53-dependent
apoptotic cell death in response to energetic stress. J Biol Chem.
283:3979–3987. 2008. View Article : Google Scholar
|
|
43
|
Jose C, Hébert-Chatelain E, Bellance N,
Larendra A, Su M, Nouette-Gaulain K and Rossignol R: AICAR inhibits
cancer cell growth and triggers cell-type distinct effects on
OXPHOS biogenesis, oxidative stress and Akt activation. Biochim
Biophys Acta. 1807:707–718. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kasznicki J, Sliwinska A and Drzewoski J:
Metformin in cancer prevention and therapy. Ann Transl Med.
2:572014.PubMed/NCBI
|
|
45
|
Faubert B, Vincent EE, Poffenberger MC and
Jones RG: The AMP-activated protein kinase (AMPK) and cancer: Many
faces of a metabolic regulator. Cancer Lett. 356:165–170. 2015.
View Article : Google Scholar
|
|
46
|
Zadra G, Batista JL and Loda M: Dissecting
the dual role of AMPK in cancer: From experimental to human
studies. Mol Cancer Res. 13:1059–1072. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hadad SM, Baker L, Quinlan PR, Robertson
KE, Bray SE, Thomson G, Kellock D, Jordan LB, Purdie CA, Hardie DG,
et al: Histological evaluation of AMPK signalling in primary breast
cancer. BMC Cancer. 9:3072009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Buzzai M, Jones RG, Amaravadi RK, Lum JJ,
DeBerardinis RJ, Zhao F, Viollet B and Thompson CB: Systemic
treatment with the antidiabetic drug metformin selectively impairs
p53-deficient tumor cell growth. Cancer Res. 67:6745–6752. 2007.
View Article : Google Scholar : PubMed/NCBI
|