Infectious viral hepatitis is a major public health
problem worldwide. The number of global viral hepatitis deaths has
recently increased. More than half of these viral hepatitis-related
deaths are associated with chronic hepatitis B virus (HBV)
infection (1). China has made
tremendous progress in controlling HBV infection, largely owing to
the enactment of a universal hepatitis B vaccination program and
implementation of effective health management programs (2). However, the prevalence of the HBV
surface antigen (HBsAg) (7.18%) is still high in Chinese adults
(3). HBV infection is a major
cause of inflammatory liver disease. Of note, HBV is considered to
be a 'mysterious' virus that is not readily sensed by the host
immune system (4). A large number
of studies have demonstrated that effective antiviral therapy and
successful HBV clearance in patients is associated with the breadth
and depth of the proliferation and responses of HBV-specific T
cells (5). However, compelling
evidence has suggested that the host immune system often exhibits
weak or absent virus-specific T-cell reactivity following chronic
HBV infection. This is referred to as T cell 'exhaustion', which is
characterized by poor effector cytotoxic responses, decreased
cytokine production and upregulated expression of multiple
inhibitory molecules, including programmed cell death-1 (PD-1),
cytotoxic T lymphocyte-associated antigen-4 (CTLA-4) and lymphocyte
activation gene-3 (6).
In a previous study, programmed death-1 (PD-1)
pathway-mediated inhibitory signals were demonstrated to serve a
key role in CD8+ T cell exhaustion during persistent
viral infection (18). However,
the exhaustion could not be completely reversed by PD-1 blockade
alone, and full restoration required a combined PD-1/CTLA-4
blockade (20). The critical
immunoregulatory role of CTLA-4 in induced peripheral immune
tolerance is illustrated by the massive and fatal
lymphoproliferation that occurs in CTLA-4-deficient mice (21). During the symptomatic phase of
acute Hepatitis A (AHA), CTLA-4 is highly expressed on
virus-specific CD8+ T cells, and functions as an
inhibitory molecule that suppresses cytotoxic T-cells and prevents
the destruction of virus-infected hepatocytes to avoid the
occurrence of severe acute hepatitis (22). However, during hepatitis C virus
(HCV) infection, high expression of CTLA-4 on CD8+ T
cells lead to increased susceptibility of the cells to spontaneous
apoptosis (23). By contrast,
functional skewing of the global CD8+ T cell population
led to impairment in their ability to produce cytokines
[interleukin (IL)-2, interferon (IFN)-γ and tumor necrosis factor
(TNF) α] and to proliferate in cells with chronic hepatitis B virus
(CHB) infection (24). Similar
findings have been reported by Wongjitrat et al (25); CD8+ expressing CTLA-4
molecules in CHB-infected patients were significantly higher
compared with healthy controls, and CD8+ T cells
presenting CTLA-4 might contribute to the impaired immune response
and the failure of immunological control of the persisting
pathogens. However, it is astounding that children and young adults
with CHB infection in the period of immune tolerance (IT) are not
associated with an immune profile of T cell tolerance, but have an
HBV-specific immune profile (26). In addition, the expression of
CTLA-4 and other inhibitory receptors (such as lymphocyte
activating 3, hepatitis A virus cellular receptor 2, and leukocyte
associated immunoglobulin like receptor 1) was not increased on
HBV-specific CD8+ T cells from peripheral blood
mononuclear cells (PBMCs). This may seem controversial to the
viewpoint that immunity is not activated in younger CHB patients.
Velazquez et al (27) may
propose a possible explanation, as this review considered that the
CD8+ T cells expression of the C-C motif chemokine
ligand 3 (CCL3), which is involved in migration, was impaired in
the immune tolerant cohort, compared with healthy controls and
immune active CHB patients. The absence of CCL3 may prompt a
potential migratory defect that could hamper functional
HBV-specific CD8+ T cells from gaining access to
virus-infected hepatocytes. Under these conditions, immune
tolerance would be largely a matter of sequestration.
However, continuous exposure to high concentrations
of HBV antigens (HBeAg, HBsAg, HBx) exhausts a large proportion or
the majority of CD8+ T cells, which is associated with
gradual upregulation of CTLA-4 expression (6). There is insufficient information to
explain the relationship between the upregulated expression of
CTLA-4 on CD8+ T cells and chronic HBV infection. Recent
findings by Peng et al confirming that HBeAg may increase
the expression of CTLA-4 on CD8+ T cells and that this
was associated with a high HBV DNA load, may lead to some
breakthroughs, but the pathway underlying this effect remains
unclear (28). It is possible
that research focusing on other diseases may provide clues into
these mechanisms. Recently, it was demonstrated in tumor models
that the mannose receptor (MR) antigens were internalized and
processed specifically for cross-presentation, which may have
resulted in upregulated expression of CTLA-4 on CD8+ T
cells. The regulatory effect of the MR was mediated by a direct
interaction with CD45 on the CD8+ T cells, inhibiting
its phosphatase activity, which prevented the expression of B-cell
lymphoma 6 (Bcl-6), a transcriptional inhibitor that directly binds
the CTLA-4 promoter and regulates its activity (29). This pathway may be one reason for
the immune escape of tumor cells that induces the development and
metastasis of tumors. However, the upregulation of CTLA-4 is
accompanied by a high level of BCL2 interacting mediator of cell
death (Bim) on CD8+ T cells following HBV infection. Bim
is a proapoptotic member of the BCL2 family, a protein important
for regulating apoptosis (30).
The activation of Bim can induce BCL2 associated X (Bax)
expression, thus increasing cell apoptosis via the mitochondrial
pathways (31). In addition, it
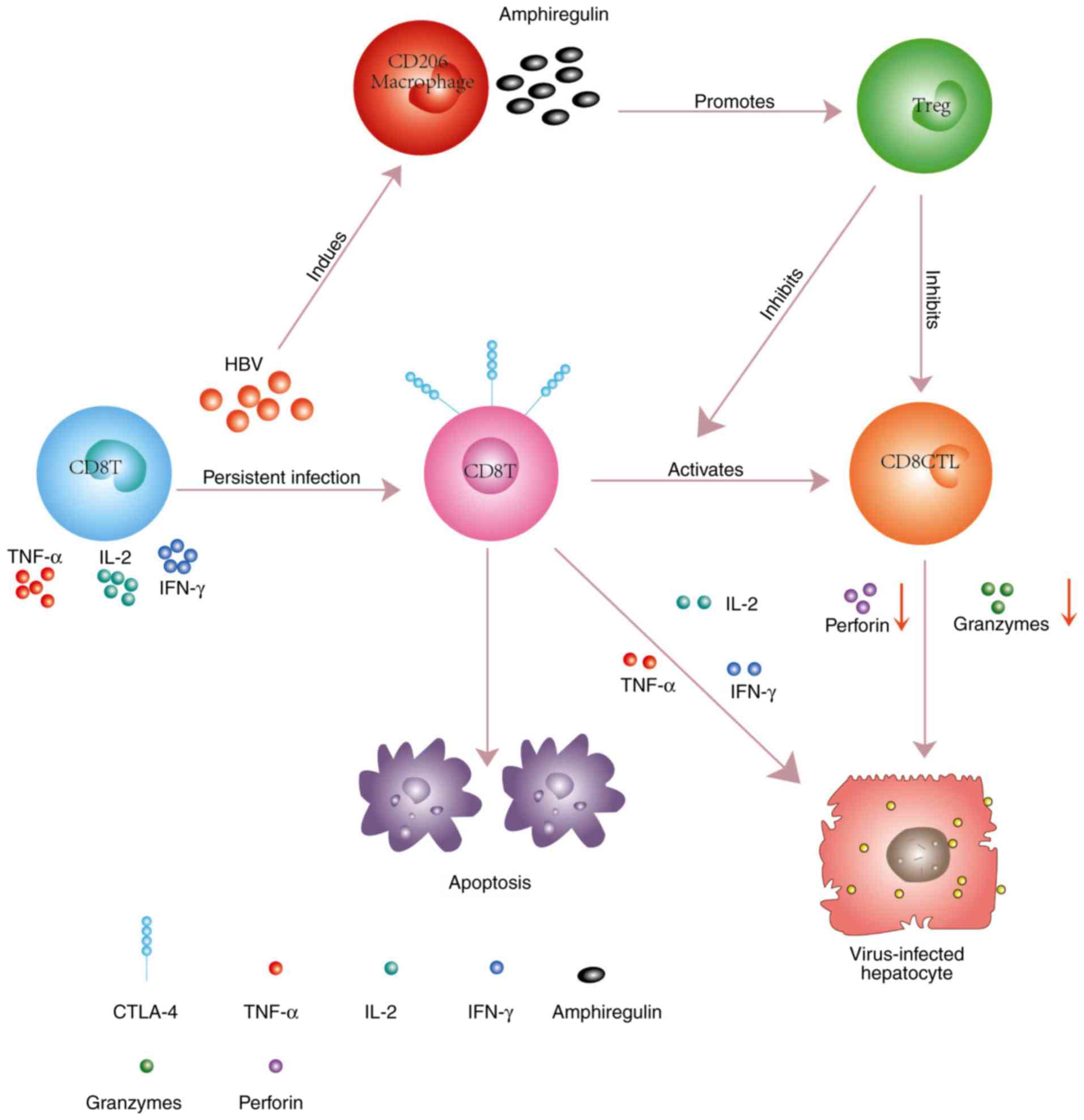
was demonstrated that CD206+ macrophage-derived
amphiregulin promoted the immunosuppressive activity of
intrahepatic regulatory T cells (Treg) by inducing mammalian target
of rapamycin (mTOR) activation, upregulating CTLA-4 expression on
Treg cells, and subsequently restraining the antiviral activity of
CD8+ T cells during chronic HBV infection (32). As a result, the proliferation of
CD8+ T cells and their activation into CD8+
cytotoxic T cells (CTL) was blocked (Fig. 1). Encouragingly, the
CD8+ T cells could be rescued by reducing the HBV DNA
load, and the immune response to the HBV protein was enhanced
(33). Schurich et al
(30) similarly demonstrated that
the function of IFN-γ-secreting HBV-specific CD8+ T
cells could improve after viral load reduction, but without any
effect on the CTLA-4 or Bim levels. It should be noted that the
latter study was based on a cohort of CHB patients commencing
antiviral therapy, all of whom had been treated for 18 months or
less. Long-term antiviral therapy may be necessary to change the
CTLA-4 and Bim levels and their impact on CD8+ T
cells.
The classical immunological synapse is described as
a central supramolecular activation cluster, which involves
molecules, such as the TCR, CD28, protein kinase Cθ (PCKθ), and
lymphocyte-specific protein tyrosine kinase (Lck) (46). In the presence of CTLA-4,
CD4+ T cells fail to form this cluster of TCR, Lck and
PCKθ. Lck binds the cytoplasmic domain of CD4 and CD8 molecules in
T cells, which are continuously phosphorylated in resting T cells.
After being recruited to the TCR-pMHC-complex with CD4+
or CD8+ T cells, Lck phos-phorylates the tyrosine in the
immunoreceptor tyrosine-based activation (ITAM) motif of CD3's
cytoplasmic domain, which is responsible for TCR-induced
intracellular signal initiation from tyrosine phosphorylation
(47). Therefore, the absence of
Lck blocks intracellular signal transduction. A lack of TCR
clustering in the immune synapse affects antigen presentation via
the MHCII receptor and the TCR receptor (signal 1). The nature of
activation (signal 1), defined by the strength of the TCR
stimulation, can affect the polarization of T helper cells towards
Th1 or Th2, in which a high affinity interaction favors Th1
development and a low affinity interaction drives Th2 development
(48). Smeets et al
(49) recently confirmed that
phorbol 12-myristate 13-acetate (PMA)/CD3 stimulation enhances the
Th1-like response in a Lck- and PKC-dependent manner, whereas
PMA/CD28 stimulation results in a Th2-like phenotype, independent
of the proximal TCR-tyrosine kinase Lck. It is intriguing to
speculate that a lack of Lck and PKCθ in the immune synapses of
CD4+ Th cells may polarize them towards a Th2
phenotype.
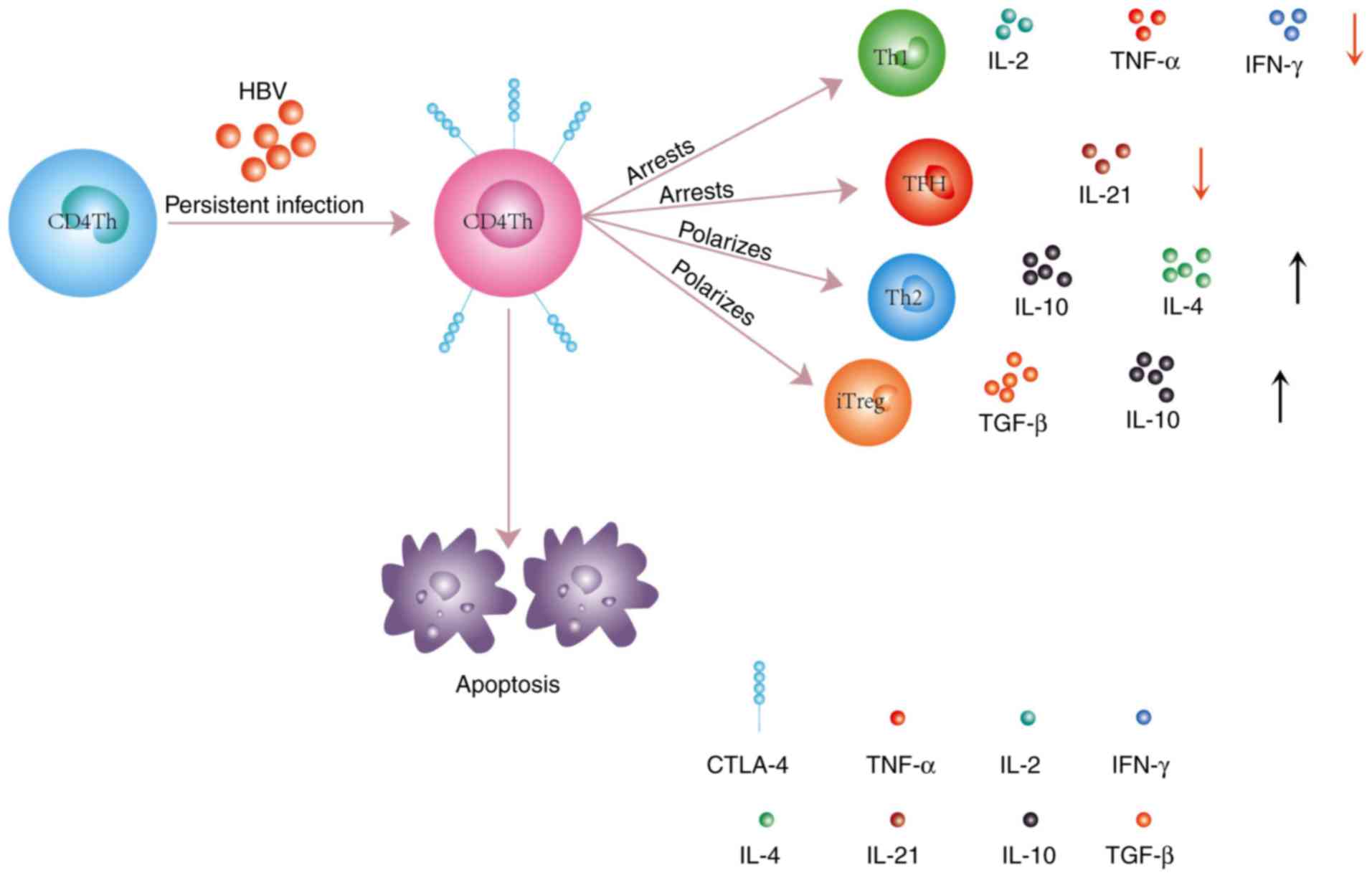
Regardless of the mechanism, it has been clearly
demonstrated that CTLA-4 induces the differentiation of
CD4+ Th cells toward the Th2 cell phenotype, and
increases the frequencies of these cells in the liver and
peripheral blood. However, higher expression of CTLA-4 might be a
consequence of a low responsiveness by Th2 cells to CTLA-4
function, and a lack of the TCR cluster in the immune synapse
hinders the proliferation of these cell and their synthesis of
cytokines (IL-3, IL-4 and IL-5) (47). It is possible that CTLA-4 may play
a coordinating role with Th2 cells through humoral immunity. Yin
et al (50) immunized mice
with DNA plasmids (HBcAg fused to the extracellular domain of
CTLA-4, termed pCTLA-4-HBc) prior to challenging by hydrodynamic
injection (HI) of pAAV/HBV1.2, and demonstrated that the clearance
of HBsAg was completed more quickly (within 16 days) in the
immunized mice compared with the control mice. Of note, >50% of
the control mice were still positive for HBsAg on day 22.
Meanwhile, the Th1/Th2 bias (the Th2 type cells were increased) and
anti-HBs antibody response developed rapidly in the mice immunized
with pCTLA-4-HBc. Similarly, Zhou et al demonstrated that a
CTLA4-fused DNA vaccine, which was constructed by linking the
extracellular domain of CTLA-4 with HBsAg, led to breakdown of IT
to viral infection in HBV transgenic mice (51). The underlying mechanism may be an
increase of Th, Th2 and HBsAg-specific CD8+ T cell
responses, as well as CTL, following vaccination with this vaccine.
These findings underline the complex impact of CTLA-4 on the immune
response.
Treg cells are broadly subdivided into two major
subtypes: Thymus-derived natural Treg (nTreg) cells and
peripherally-inducible Treg (iTreg) cells (57). iTreg cells have a large repertoire
of self-specific and non-self-specific T-cell receptors and have
been detected in several human infectious diseases (58). Highlighting the importance of
CTLA-4 in the suppression of the immune response, CTLA-4 expression
and signaling are essential for Treg cells to execute their
suppressive function (59).
CTLA-4 can induce a costimulatory blockade either by sequestering
or removing costimulatory ligands from the surface of APCs (perhaps
via transendocytosis), and might also stimulate these cells to
secrete indoleamine 2,3-dioxygenase (IDO), which limits the
availability of tryptophan and metabolically suppresses the
activation of naïve T cells (60). Direct mutation of CTLA-4 or CTLA-4
deficiency leads to defective Treg cell function, which is
associated with an impaired ability to control the levels of the
CTLA-4 ligands, CD80 and CD86 (61). In addition, CTLA-4 may strengthen
the secretion of cytokines, such as TGF-β and IL-10, by the Treg
cells, to suppress a wide range of cells and cytokines (62). Yang et al (63), Stoop et al (64) and Xu et al (65) have demonstrated that patients with
CHB infection have a higher frequency of Treg cells in peripheral
blood and liver compared with healthy controls. A recent study
concluded that there were increased levels of Treg cells
accompanied by CTLA-4 during CHB infection, and that these cells
were associated with the antiviral immune response and disease
progression (66). Previously,
Feng et al (67) have
demonstrated that HBcAg peptide-specific Treg cells from liver and
peripheral blood of CHB patients express CTLA-4 and have a pivotal
role in modulating IT. Enhanced HBcAg peptide-specific CTL
frequencies by blocking of CTLA-4 may account for spontaneous Acute
exacerbations (AEs) in the process of CHB infection. One possible
mechanism for the change in the natural history of CHB infection by
blocking CTLA-4 may be the inhibition of TGF-β secretion, which
functions as a mediator of Treg cell suppressor effectors (68). Recently, Zhang et al
(69) reported similar findings;
they demonstrated that CTLA-4 expression levels were significantly
higher on Treg cells from patients with HBsAg-positive
hepatocellular carcinoma compared with healthy controls, whereas no
difference was observed in HBsAg-negative HCC patients.
Furthermore, when the human hepatoma cell line HepG2.2.15
(transfected with HBV) and its parental cell line HepG2 were
co-cultured with healthy donor PBMCs, the results demonstrated that
HepG2.2.15 cells strongly increased the frequencies of Treg cells
and CTLA-4 expression compared with the HepG2 cells. Notably, the
Treg cells from CHB patients can help HCC tumor antigens escape
tumor immunosurveillance. It appears that high expression of CTLA-4
on Treg cells is a risk factor in HBsAg-positive HCC. Inhibition of
Treg cells and CTLA-4 may therefore represent a novel therapeutic
approach for CHB patients. However, these studies have failed to
explain the mechanism by which CHB infection upregulated CTLA-4 on
Treg cells. Liu et al (70) may have provided evidence in that
direction, by demonstrating that IL-10-producing regulatory
B-cells, which are a new subset of B-cells that exert
immunosuppressive functions in autoimmunity and infections, can
enhance Treg cells in chronic HBV infection and upregulate the
expression of CTLA-4. Therefore, it can be speculated that an
imbalance of cytokines in the microenvironment of the liver may be
one of the reasons. A recent study indicated that total Treg cell
frequencies were affected by IL-2, and the strongest Treg cell
suppressive phenotype was observed under low-dose IL-2. It was
hypothesized that low-dose IL-2 promotes STAT5 phosphorylation and
subsequently upregulates CTLA-4 (71). Nevertheless, the relationship
between the proteins of HBV and CTLA-4 expression following CHB
infection remain unclear.
There have recently been conflicting findings
regarding the function of Treg cells (Table I), suggesting that their
activation may not be entirely dependent on CTLA-4. Treg activation
can be mediated by forkhead box P3 (Foxp3), as it serves a central
role in directing the regulatory program (72). Foxp3 expression is essentially
confined to CD4+CD25+ cells and is
responsible for the regulatory activity of this subset of cells.
Accordingly, the adoptive transfer of
CD4+CD25+ T cells from wild type mice can
rescue scurfy mice, which are deficient in Treg cells, from
lymphoproliferative syndrome. Retroviral expression and transgenic
expression of Foxp3 in CD25− T cells have been
demonstrated to endow them with regulatory functions, and even
induce regulatory activity in CD8+ T cells (73). Consistent with the large body of
evidence obtained in mouse models, mutations in the Foxp3 gene in
humans are associated with defective immune regulation, manifesting
as a syndrome that has been termed 'immune dysregulation
polyendocrinopathy enteropathy Xlinked' (IPEX) (74). Together, these findings suggest
that Foxp3 and CTLA-4 exhibit some redundant functions and can
function independently of one another. More recently, Paterson
et al (75) have put
forward new evidence that Treg cells remain functionally
suppressive and produce an overabundance of IL-10 following the
deletion of CTLA-4. The authors demonstrated that the deletion of
CTLA-4 in mice during adulthood did not precipitate systemic
autoimmunity, but surprisingly conferred protection against
experimental autoimmune encephalomyelitis (EAE). Furthermore, they
demonstrated that the deletion of CTLA-4 was accompanied by the
activation and expansion of both conventional CD4+ T
cells and Treg cell subsets. Notwithstanding, Fontenot et al
reported that all mice receiving a mix of scurfy and
CTLA-4−/− bone marrow died, and while transgenic
overexpression of Foxp3 could delay the lethality in
CTLA-4−/− mice, it could not completely rescue the mice
(73). Hence, effective immune
regulation by Treg cells requires the co-expression of Foxp3 and
CTLA-4. In conclusion, these findings suggest that upregulation of
CTLA-4 in Treg cells in HBV infection patients is involved in the
immunopathogenesis of acute Hepatitis B (AHB) to CHB.
Cytokines mediate the non-cytolytic clearance of
HBV, and have been demonstrated to control HBV replication and to
contribute to curing HBV in several disease models (76). IFN-γ restricts HBV entry by
inducing soluble factors that bind to heparan sulfate proteoglycan
(HSPG) and block HBV attachment (77). IL-1β regulates sodium taurocholate
cotransporting polypeptide (NTCP) expression to inhibit HBV
infection (78). IFN-γ and TNF-α
interfere with covalently closed circular DNA (cccDNA) integrity
and stability by inducing cccDNA deamination and subsequent
degradation in HBV-infected primary human hepatocytes and HepaRG
cells (79). IL-6 may reduce
cccDNA and HBsAg secretion (80).
CTLA-4 has been demonstrated to be a major and specific regulator
of the production of both pro- and anti-inflammatory cytokines. In
the dominant negative transforming growth factor β receptor II
(dnTGFβRII) mouse model, CTLA-4 Ig quickly reduced the level of
intrahepatic proinflammatory cytokines, thereby reducing the bile
duct damage (81). Blockade of
CTLA-4 resulted in increased secretion of granulocyte-macrophage
colony stimulating factor (GM-CSF), IL-1, IL-2 and IFN-γ by
CD4+ Th cells (82).
Similar to CD4+ Th cells, CD8+ T cells
exhibited an enhanced ability to control HBV infection following
inhibition of CTLA-4. Virus-specific CD8+ T cells can
function in two different ways during an interaction with
HBV-producing hepatocytes. When they are cytolytic, they can induce
the clearance of intrahepatic cccDNA by killing a fraction of the
infected cells. The noncytolytic activity is mediated by an array
of cytokines, including IFN-γ, TNF-α and IL-2 (83). When CTLA-4 is silenced by RNA
interference in in vitro cultures of PBMCs derived from
CHB-infected patients, an enhanced secretion of IFN-γ and IL-2 were
observed. Furthermore, when the CD8+ T cells from CHB
patients were treated with anti-CTLA-4 antibodies, the levels of
IFN-γ, TNF-α and IL-2 were significantly increased following
stimulation with an HBV peptide (28,84). Finally, Pedicord et al
(85) confirmed that anti-CTLA-4
antibodies could enhance the memory formation, function, and
maintenance of CD8+ T cells. The above studies confirmed
that CTLA-4 decreases the synthesis of many proinflammatory
cytokines. However, further studies are required to determine the
specific mechanisms involved.
This review summarized the current literature
highlighting the negative effects of CTLA-4 in the context of
chronic HBV infection (Table
II). Although the specific mechanisms leading to the
upregulation of CTLA-4 on T cells remain unclear, CTLA-4 has been
demonstrated to inhibit HBV-specific T cell immune responses.
Anti-CTLA-4 antibodies have exhibited immunomodulatory effects in
patients with hepatocellular carcinoma and chronic hepatitis C
(86). The mechanism underlying
the antitumor and antiviral activity of these antibodies may be
associated with a recovery of the function of the host T cells, as
well as decreased deletion of T cells by blockade of CTLA-4.
However, severe immune-mediated adverse events may limit the
clinical potential of such treatments. Therefore, future studies
are needed to explore the specific effector T-cell response that
may be regulated by CTLA-4, as well as potential T cell-independent
mechanisms.
This study was funded by the National Natural
Science Foundation of China (grant no. 81373860).
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
HC wrote the manuscript. WZ edited and proofread
this manuscript. RWZ was a major contributor in commenting and
revising the manuscript. All authors read and approved the final
manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
The authors thank Ying Yu Wang for assistance with
figure production.
|
1
|
Stanaway JD, Flaxman AD, Naghavi M,
Fitzmaurice C, Vos T, Abubakar I, Abu-Raddad LJ, Assadi R, Bhala N,
Cowie B, et al: The global burden of viral hepatitis from 1990 to
2013: Findings from the global burden of disease study 2013.
Lancet. 388:1081–1088. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lu FM, Li T, Liu S and Zhuang H:
Epidemiology and prevention of hepatitis B virus infection in
China. J Viral Hepat. 17(Suppl 1): S4–S9. 2010. View Article : Google Scholar
|
|
3
|
Liu J, Zhang S, Wang Q, Shen H, Zhang M,
Zhang Y, Yan D and Liu M: Seroepidemiology of hepatitis B virus
infection in 2 million men aged 21-49 years in rural China: A
population-based, cross-sectional study. Lancet Infect Dis.
16:80–86. 2016. View Article : Google Scholar
|
|
4
|
Blendis L, Lurie Y and Oren R: Occult HBV
infection-both hidden and mysterious. Gastroenterology.
125:1903–1905. 2003. View Article : Google Scholar
|
|
5
|
Park JJ, Wong DK, Wahed AS, Lee WM, Feld
JJ, Terrault N, Khalili M, Sterling RK, Kowdley KV, Bzowej N, et
al: Hepatitis B virus - specific and global T-cell dysfunction in
chronic hepatitis B. Gastroenterology. 150:684–695.e5. 2016.
View Article : Google Scholar
|
|
6
|
Ye B, Liu X, Li X, Kong H, Tian L and Chen
Y: T-cell exhaustion in chronic hepatitis B infection: Current
knowledge and clinical significance. Cell Death Dis. 6:e16942015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ling V, Wu PW, Finnerty HF, Sharpe AH,
Gray GS and Collins M: Complete sequence determination of the mouse
and human CTLA4 gene loci: Cross-species DNA sequence similarity
beyond exon borders. Genomics. 60:341–355. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lindsten T, Lee KP, Harris ES, Petryniak
B, Craighead N, Reynolds PJ, Lombard DB, Freeman GJ, Nadler LM,
Gray GS, et al: Characterization of CTLA-4 structure and expression
on human T cells. J Immunol. 151:3489–3499. 1993.PubMed/NCBI
|
|
9
|
Adams AB, Ford ML and Larsen CP:
Costimulation blockade in autoimmunity and transplantation: The CD
28 pathway. J Immunol. 197:2045–2050. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brunet JF, Denizot F, Luciani MF,
Roux-Dosseto M, Suzan M, Mattei MG and Golstein P: A new member of
the immunoglobulin superfamily - CTLA-4. Nature. 328:267–270. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tang ST, Tang HQ, Zhang Q, Wang CJ, Wang
YM and Peng WJ: Association of cytotoxic T-lymphocyte associated
antigen 4 gene polymorphism with type 1 diabetes mellitus: A
meta-analysis. Gene. 508:165–187. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li Q, Wang B, Pan F, Zhang R, Xiao L, Guo
H, Ma S and Zhou C: Association between cytotoxic T-lymphocyte
antigen 4 gene polymorphisms and primary biliary cirrhosis in
Chinese population: Data from a multicenter study. J Gastroenterol
Hepatol. 28:1397–1402. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Du L, Yang J and Huang J, Ma Y, Wang H,
Xiong T, Xiang Z, Zhang Y and Huang J: The associations between the
polymorphisms in the CTLA-4 gene and the risk of Graves' disease in
the Chinese population. BMC Med Genet. 14:462013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang R, Hao Y, Fan Y, Yang C, Wu K, Cao S
and Wu C: Association between cytotoxic T-lymphocyte-associated
antigen 4+49A/G polymorphism and persistent hepatitis B virus
infection in the Asian population: Evidence from the current
studies. Genet Test Mol Biomarkers. 17:601–606. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Isogawa M, Furuichi Y and Chisari FV:
Oscillating CD8+ T cell effector functions after antigen
recognition in the liver. Immunity. 23:53–63. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Benechet AP and Iannacone M: Determinants
of hepatic effector CD8+ T cell dynamics. J Hepatol.
66:228–233. 2017. View Article : Google Scholar
|
|
17
|
Bengsch B, Martin B and Thimme R:
Restoration of HBV-specific CD8+ T cell function by PD-1
blockade in inactive carrier patients is linked to T cell
differentiation. J Hepatol. 61:1212–1219. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Barber DL, Wherry EJ, Masopust D, Zhu B,
Allison JP, Sharpe AH, Freeman GJ and Ahmed R: Restoring function
in exhausted CD8 T cells during chronic viral infection. Nature.
439:682–687. 2006. View Article : Google Scholar
|
|
19
|
Legat A, Speiser DE, Pircher H, Zehn D and
Fuertes Marraco SA: Inhibitory receptor expression depends more
dominantly on differentiation and activation than 'Exhaustion' of
human CD8 T cells. Front Immunol. 4:4552013. View Article : Google Scholar
|
|
20
|
Nakamoto N, Cho H, Shaked A, Olthoff K,
Valiga ME, Kaminski M, Gostick E, Price DA, Freeman GJ, Wherry EJ
and Chang KM: Synergistic reversal of intrahepatic HCV-specific CD8
T cell exhaustion by combined PD-1/CTLA-4 blockade. PLoS Pathog. 5.
pp. e10003132009, View Article : Google Scholar
|
|
21
|
Waterhouse P, Penninger JM, Timms E,
Wakeham A, Shahinian A, Lee KP, Thompson CB, Griesser H and Mak TW:
Lymphoproliferative disorders with early lethality in mice
deficient in Ctla-4. Science. 270:985–988. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cho H, Kang H, Kim CW, Kim HY, Jang JW,
Yoon SK and Lee CD: Phenotypic characteristics of PD-1 and CTLA-4
expression in symptomatic acute hepatitis A. Gut Liver. 10:288–294.
2016. View Article : Google Scholar :
|
|
23
|
Radziewicz H, Ibegbu CC, Hon H, Osborn MK,
Obideen K, Wehbi M, Freeman GJ, Lennox JL, Workowski KA, Hanson HL
and Grakoui A: Impaired hepatitis C virus (HCV)-specific effector
CD8+ T cells undergo massive apoptosis in the peripheral
blood during acute HCV infection and in the liver during the
chronic phase of infection. J Virol. 82:9808–9822. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Das A, Hoare M, Davies N, Lopes AR, Dunn
C, Kennedy PT, Alexander G, Finney H, Lawson A, Plunkett FJ, et al:
Functional skewing of the global CD8 T cell population in chronic
hepatitis B virus infection. J Exp Med. 205:2111–2124. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wongjitrat C, Sukwit S, Chuenchitra T,
Seangjaruk P, Rojanasang P, Romputtan P and Srisurapanon S: CTLA-4
and its ligands on the surface of T- and B-lymphocyte subsets in
chronic hepatitis B virus infection. J Med Assoc Thai. 96(Suppl 1):
S54–S59. 2013.PubMed/NCBI
|
|
26
|
Kennedy PTF, Sandalova E, Jo J, Gill U,
Ushiro-Lumb I, Tan AT, Naik S, Foster GR and Bertoletti A:
Preserved T-cell function in children and young adults with
immune-tolerant chronic hepatitis B. Gastroenterology. 143:637–645.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Velazquez VM and Grakoui A: Immune
quiescence and hepatitis B virus: Tolerance has its limits.
Gastroenterology. 143:529–532. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Peng G, Luo B, Li J, Zhao D, Wu W, Chen F
and Chen Z: Hepatitis B e-antigen persistency is associated with
the properties of HBV-specific CD8 T cells in CHB patients. J Clin
Immunol. 31:195–204. 2011. View Article : Google Scholar
|
|
29
|
Schuette V, Embgenbroich M, Ulas T, Welz
M, Schulte- Schrepping J, Draffehn AM, Quast T, Koch K, Nehring M,
König J, et al: Mannose receptor induces T-cell tolerance via
inhibition of CD45 and up-regulation of CTLA-4. Proc Natl Acad Sci
USA. 113:10649–10654. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schurich A, Khanna P, Lopes AR, Han KJ,
Peppa D, Micco L, Nebbia G, Kennedy PT, Geretti AM, Dusheiko G and
Maini MK: Role of the coinhibitory receptor cytotoxic T lymphocyte
antigen-4 on apoptosis-Prone CD8 T cells in persistent hepatitis B
virus infection. Hepatology. 53:1494–1503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang YM, Zhang GY, Wang Y, Hu M, Zhou JJ,
Sawyer A, Cao Q, Wang Y, Zheng G, Lee VW, et al: Exacerbation of
spontaneous autoimmune nephritis following regulatory T cell
depletion in B cell lymphoma 2-interacting mediator knock-out mice.
Clin Exp Immunol. 188:195–207. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dai K, Huang L, Sun X, Yang L and Gong Z:
Hepatic CD206-positive macrophages express amphiregulin to promote
the immunosuppressive activity of regulatory T cells in HBV
infection. J Leukoc Biol. 98:1071–1080. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nitschke K, Luxenburger H, Kiraithe MM,
Thimme R and Neumann-Haefelin C: CD8+ T-cell responses
in hepatitis B and C: The (HLA-) A, B, and C of hepatitis B and C.
Dig Dis. 34:396–409. 2016. View Article : Google Scholar
|
|
34
|
Farhan RK, Vickers MA, Ghaemmaghami AM,
Hall AM, Barker RN and Walsh GM: Effective antigen presentation to
helper T cells by human eosinophils. Immunology. 149:413–422. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li M, Sun XH, Zhu XJ, Jin SG, Zeng ZJ,
Zhou ZH, Yu Z and Gao YQ: HBcAg induces PD-1 upregulation on
CD4+ T cells through activation of JNK, ERK and PI3K/AKT
pathways in chronic hepatitis-B-infected patients. Lab Invest.
92:295–304. 2012. View Article : Google Scholar
|
|
36
|
Wang H, Wu D, Wang X, Chen G, Zhang Y, Yan
W, Luo X, Han M and Ning Q: Hepatitis B virus surface
protein-induced hPIAS1 transcription requires TAL1, E47, MYOG, NFI,
and MAPK signal pathways. Biol Chem. 397:1173–1185. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Raziorrouh B, Heeg M, Kurktschiev P,
Schraut W, Zachoval R, Wendtner C, Wächtler M, Spannagl M, Denk G,
Ulsenheimer A, et al: Inhibitory phenotype of HBV-specific
CD4+ T-cells is characterized by high PD-1 expression
but absent coregulation of multiple inhibitory molecules. PLoS One.
9:e1057032014. View Article : Google Scholar
|
|
38
|
Walker LS and Sansom DM: The emerging role
of CTLA4 as a cell-extrinsic regulator of T cell responses. Nat Rev
Immunol. 11:852–863. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Halpert MM, Konduri V, Liang D, Chen Y,
Wing JB, Paust S, Levitt JM and Decker WK: Dendritic cell-secreted
cytotoxic T-lymphocyte-associated protein-4 regulates the T-cell
response by downmodulating bystander surface B7. Stem Cells Dev.
25:774–787. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tai X, Van Laethem F, Pobezinsky L,
Guinter T, Sharrow SO, Adams A, Granger L, Kruhlak M, Lindsten T,
Thompson CB, et al: Basis of CTLA-4 function in regulatory and
conventional CD4+ T cells. Blood. 119:5155–5163. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ville S, Poirier N, Blancho G and Vanhove
B: Co-stimulatory blockade of the CD28/CD80-86/CTLA-4 balance in
transplantation: Impact on memory T cells? Front Immunol.
6:4112015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cloutier JF and Veillette A: Cooperative
inhibition of T-cell antigen receptor signaling by a complex
between a kinase and a phosphatase. J Exp Med. 189:111–121. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhu J, Yamane H and Paul WE:
Differentiation of effector CD4 T cell populations (*). Annu Rev
Immunol. 28:445–489. 2010. View Article : Google Scholar
|
|
44
|
Tang ZS, Hao YH, Zhang EJ, Xu CL, Zhou Y,
Zheng X and Yang DL: CD28 family of receptors on T cells in chronic
HBV infection: Expression characteristics, clinical significance
and correlations with PD-1 blockade. Mol Med Rep. 14:1107–1116.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dilek N, Poirier N, Hulin P, Coulon F,
Mary C, Ville S, Vie H, Clémenceau B, Blancho G and Vanhove B:
Targeting CD28, CTLA-4 and PD-L1 costimulation differentially
controls immune synapses and function of human regulatory and
conventional T-cells. PLoS One. 8:e831392013. View Article : Google Scholar :
|
|
46
|
Grakoui A, Bromley SK, Sumen C, Davis MM,
Shaw AS, Allen PM and Dustin ML: The immunological synapse: A
molecular machine controlling T cell activation. Science.
285:221–227. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Alegre ML, Shiels H, Thompson CB and
Gajewski TF: Expression and function of CTLA-4 in Th1 and Th2
cells. J Immunol. 161:3347–3356. 1998.PubMed/NCBI
|
|
48
|
Turner MS, Isse K, Fischer DK, Turnquist
HR and Morel PA: Low TCR signal strength induces combined expansion
of Th2 and regulatory T cell populations that protect mice from the
development of type 1 diabetes. Diabetologia. 57:1428–1436. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Smeets RL, Fleuren WW, He X, Vink PM,
Wijnands F, Gorecka M, Klop H, Bauerschmidt S, Garritsen A, Koenen
HJ, et al: Molecular pathway profiling of T lymphocyte signal
transduction pathways; Th1 and Th2 genomic fingerprints are defined
by TCR and CD28-mediated signaling. BMC Immunol. 13:122012.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yin Y, Wu C, Song J, Wang J, Zhang E, Liu
H, Yang D, Chen X, Lu M and Xu Y: DNA immunization with fusion of
CTLA-4 to hepatitis B virus (HBV) core protein enhanced Th2 type
responses and cleared HBV with an accelerated kinetic. PLoS One.
6:e225242011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhou C, Peng G, Jin X, Tang J and Chen Z:
Vaccination with a fusion DNA vaccine encoding hepatitis B surface
antigen fused to the extracellular domain of CTLA4 enhances
HBV-specific immune responses in mice: Implication of its potential
use as a therapeutic vaccine. Clin Immunol. 137:190–198. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fazilleau N, Mark L, McHeyzer-Williams LJ
and McHeyzer- Williams MG: Follicular helper T cells: Lineage and
location. Immunity. 30:324–335. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tivol EA, Borriello F, Schweitzer AN,
Lynch WP, Bluestone JA and Sharpe AH: Loss of CTLA-4 leads to
massive lymphoproliferation and fatal multiorgan tissue
destruction, revealing a critical negative regulatory role of
CTLA-4. Immunity. 3:541–547. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang CJ, Heuts F, Ovcinnikovs V,
Wardzinski L, Bowers C, Schmidt EM, Kogimtzis A, Kenefeck R, Sansom
DM and Walker LS: CTLA-4 controls follicular helper T-cell
differentiation by regulating the strength of CD28 engagement. Proc
Natl Acad Sci USA. 112:524–529. 2015. View Article : Google Scholar
|
|
55
|
Qureshi OS, Zheng Y, Nakamura K, Attridge
K, Manzotti C, Schmidt EM, Baker J, Jeffery LE, Kaur S, Briggs Z,
et al: Trans-endocytosis of CD80 and CD86: A molecular basis for
the cell-extrinsic function of CTLA-4. Science. 332:600–603. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Li Y, Ma S, Tang L, Li Y, Wang W, Huang X,
Lai Q, Zhang M, Sun J, Li CK, et al: Circulating chemokine (C-X-C
Motif) receptor 5(+) CD4(+) T cells benefit hepatitis B e antigen
seroconversion through IL-21 in patients with chronic hepatitis B
virus infection. Hepatology. 58:1277–1286. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Abbas AK, Benoist C, Bluestone JA,
Campbell DJ, Ghosh S, Hori S, Jiang S, Kuchroo VK, Mathis D,
Roncarolo MG, et al: Regulatory T cells: Recommendations to
simplify the nomenclature. Nat Immunol. 14:307–308. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Shevach EM and Thornton AM: tTregs,
pTregs, and iTregs: Similarities and differences. Immunol Rev.
259:88–102. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kolar P, Knieke K, Hegel JK, Quandt D,
Burmester GR, Hoff H and Brunner-Weinzierl MC: CTLA-4 (CD152)
controls homeostasis and suppressive capacity of regulatory T cells
in mice. Arthritis Rheum. 60:123–132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Guntermann C and Alexander DR: CTLA-4
suppresses proximal TCR signaling in resting human CD4+
T cells by inhibiting ZAP-70 Tyr319 phosphorylation: A
potential role for tyrosine phosphatases. J Immunol. 168:4420–4429.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hou TZ, Verma N, Wanders J, Kennedy A,
Soskic B, Janman D, Halliday N, Rowshanravan B, Worth A, Qasim W,
et al: Identifying functional defects in patients with immune
dysregulation due to LRBA and CTLA-4 mutations. Blood.
129:1458–1468. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Mellor AL and Munn DH: IDO expression by
dendritic cells: Tolerance and tryptophan catabolism. Nat Rev
Immunol. 4:762–774. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Yang G, Liu A, Xie Q, Guo TB, Wan B, Zhou
B and Zhang JZ: Association of
CD4+CD25+Foxp3+ regulatory T cells
with chronic activity and viral clearance in patients with
hepatitis B. Int Immunol. 19:133–140. 2007. View Article : Google Scholar
|
|
64
|
Stoop JN, van der Molen RG, Baan CC, van
der Laan LJ, Kuipers EJ, Kusters JG and Janssen HL: Regulatory T
cells contribute to the impaired immune response in patients with
chronic hepatitis B virus infection. Hepatology. 41:771–778. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Xu D, Fu J, Jin L, Zhang H, Zhou C, Zou Z,
Zhao JM, Zhang B, Shi M, Ding X, et al: Circulating and liver
resident CD4+CD25+ regulatory T cells
actively influence the antiviral immune response and disease
progression in patients with hepatitis B. J Immunol. 177:739–747.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Tavakolpour S, Alavian SM and Sali S:
Manipulation of regulatory cells' responses to treatments for
chronic hepatitis B virus infection. Hepat Mon. 16:e379272016.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Feng IC, Koay LB, Sheu MJ, Kuo HT, Sun CS,
Lee C, Chuang WL, Liao SK, Wang SL, Tang LY, et al: HBcAg-specific
CD4+CD25+ regulatory T cells modulate immune
tolerance and acute exacerbation on the natural history of chronic
hepatitis B virus infection. J Biomed Sci. 14:43–57. 2007.
View Article : Google Scholar
|
|
68
|
Chen W, Jin W and Wahl SM: Engagement of
cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) induces
transforming growth factor beta (TGF-beta) production by murine
CD4+ T cells. J Exp Med. 188:1849–1857. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kalathil S, Lugade AA, Miller A, Iyer R
and Thanavala Y: Higher frequencies of
GARP+CTLA-4+Foxp3+ T regulatory
cells and myeloid-derived suppressor cells in hepatocellular
carcinoma patients are associated with impaired T-cell
functionality. Cancer Res. 73:2435–2444. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Liu Y, Cheng LS, Wu SD, Wang SQ, Li L, She
WM, Li J, Wang JY and Jiang W: IL-10-producing regulatory B-cells
suppressed effector T-cells but enhanced regulatory T-cells in
chronic HBV infection. Clin Sci. 130:907–919. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Jeffery HC, Jeffery LE, Lutz P, Corrigan
M, Webb GJ, Hirschfield GM, Adams DH and Oo YH: Low-dose
interleukin-2 promotes STAT-5 phosphorylation, Treg survival and
CTLA-4-dependent function in autoimmune liver diseases. Clin Exp
Immunol. 188:394–411. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Walker LS: Treg and CTLA-4: Two
intertwining pathways to immune tolerance. J Autoimmun. 45:49–57.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Fontenot JD, Gavin MA and Rudensky AY:
Foxp3 programs the development and function of CD4+CD25+ regulatory
T cells. Nat Immunol. 4:330–336. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
74
|
Bennett CL, Christie J, Ramsdell F,
Brunkow ME, Ferguson PJ, Whitesell L, Kelly TE, Saulsbury FT,
Chance PF and Ochs HD: The immune dysregulation,
polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused
by mutations of FOXP3. Nat Genet. 27:20–21. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Paterson AM, Lovitch SB, Sage PT, Juneja
VR, Lee Y, Trombley JD, Arancibia-Cárcamo CV, Sobel RA, Rudensky
AY, Kuchroo VK, et al: Deletion of CTLA-4 on regulatory T cells
during adulthood leads to resistance to autoimmunity. J Exp Med.
212:1603–1621. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Xia Y and Protzer U: Control of hepatitis
B virus by cytokines. Viruses. 9:pii: E182017. View Article : Google Scholar
|
|
77
|
Xia Y and Cheng X: Secreted
interferon-inducible factors restrict hepatitis B and C virus entry
in vitro. J Immunol Res. 2017:48289362017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Le Vee M, Gripon P, Stieger B and Fardel
O: Down-regulation of organic anion transporter expression in human
hepatocytes exposed to the proinflammatory cytokine interleukin
1beta. Drug Metab Dispos. 36:217–222. 2008. View Article : Google Scholar
|
|
79
|
Xia Y, Stadler D, Lucifora J, Reisinger F,
Webb D, Hösel M, Michler T, Wisskirchen K, Cheng X, Zhang K, et al:
Interferon-γ and tumor necrosis Factor-α produced by T cells reduce
the HBV persistence form, cccDNA, without cytolysis.
Gastroenterology. 150:194–205. 2016. View Article : Google Scholar
|
|
80
|
Bouezzedine F, Fardel O and Gripon P:
Interleukin 6 inhibits HBV entry through NTCP down regulation.
Virology. 481:34–42. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Dhirapong A, Yang GX, Nadler S, Zhang W,
Tsuneyama K, Leung P, Knechtle S, Ansari AA, Coppel RL, Liu FT, et
al: Therapeutic effect of cytotoxic T lymphocyte antigen
4/immunoglobulin on a murine model of primary biliary cirrhosis.
Hepatology. 57:708–715. 2013. View Article : Google Scholar :
|
|
82
|
Kolar P, Hoff H, Maschmeyer P, Burmester
GR and Brunner-Weinzierl MC: CTLA-4 (CD152) blockade does not cause
a pro-inflammatory cytokine profile in regulatory T cells. Clin Exp
Rheumatol. 29:254–260. 2011.PubMed/NCBI
|
|
83
|
Phillips S, Chokshi S, Riva A, Evans A,
Williams R and Naoumov NV: CD8+ T cell control of
hepatitis B virus replication: Direct comparison between cytolytic
and noncytolytic functions. J Immunol. 184:287–295. 2010.
View Article : Google Scholar
|
|
84
|
Yu Y, Wu H, Tang Z and Zang G: CTLA4
silencing with siRNA promotes deviation of Th1/Th2 in chronic
hepatitis B patients. Cell Mol Immunol. 6:123–127. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Pedicord VA, Montalvo W, Leiner IM and
Allison JP: Single dose of anti-CTLA-4 enhances CD8+
T-cell memory formation, function, and maintenance. Proc Natl Acad
Sci USA. 108:266–271. 2011. View Article : Google Scholar
|
|
86
|
Sangro B, Gomez-Martin C, de la Mata M,
Iñarrairaegui M, Garralda E, Barrera P, Riezu-Boj JI, Larrea E,
Alfaro C, Sarobe P, et al: A clinical trial of CTLA-4 blockade with
tremelimumab in patients with hepatocellular carcinoma and chronic
hepatitis C. J Hepatol. 59:81–88. 2013. View Article : Google Scholar : PubMed/NCBI
|