Introduction
Skin is characterized as the first defense against
harmful biological, physical and environmental pollutants,
including ultraviolet (UV) irradiation (1). Excessive UVB exposure has been
reported to disrupt biological macromolecules, including proteins,
lipids and nucleic acids, leading to various disorders of skin and
ultimately to skin cancer (2,3).
Additionally, solar UVB irradiation can result in sunburn,
proliferation, oxidative stress, apoptosis, oxidative stress and
erythema, which are associated with various signaling pathways,
including the toll-like receptor (TLR)4/nuclear factor-κB (NF-κB)
and mitogen-activate protein kinases (MAPKs) (4). Pro-inflammatory cytokines, including
interleukin (IL)-1β, tumor necrosis factor (TNF)-α, IL-18, IL-6 and
cyclooxygenase (COX)2, have been suggested to be crucial in the
progression of inflammatory responses (5,6).
UVB, as previously reported, can lead to the high release of COX2.
COX2 is involved in the inflammatory response, cell survival and
proliferation (7). The secretion
of pro-inflammatory cytokines is linked to activation of the
TLR4/NF-κB signaling pathway. TLRs are important in initiating the
inflammatory response. Several stimuli can activate TLR4. Once
activated, myeloid differentiation factor 88 (MyD88), a key adapter
protein for TLR4, leads to the direct activation of NF-κB and the
subsequent induction of pro-inflammatory cytokines, including
TNF-α, IL-1β and IL-6, which contribute to the inflammatory
response (8,9). Drugs inhibiting the activity of the
TLR inflammatory system afford potential beneficial effects for
various tissues or organs, including the skin, as do transgenic
methods of inhibiting TLR4/NF-κB-related genes (10). Reactive oxygen species (ROS),
reflecting levels of oxidative stress, are closely associated with
a variety of biomolecules, leading to damage of biological
structures, and contributing to cellular injury and ultimately
tissue destruction. The skin has evolved effective mechanisms for
the protection from ROS damage, which involves ROS detoxification
and DNA repair (11). Therefore,
the excessive production of ROS disrupts molecular balances,
promoting the pathogenesis of inflammatory skin diseases, which
identifies a target for drug investigations.
MAPKs, including p38, extracellular signal-regulated
kinase (ERK)1/2 and c-Jun N-terminal kinase (JNK), are important in
regulating apoptosis (12). P38
and JNK are often activated in human tumors, and are associated
with cell proliferation, migration and metastasis. p38-MAPK has
also been described as a tumor suppressor, particularly under
situations of stress (13). The
p38-MAPK pathway can be regulated by transcriptional and
post-transcriptional mechanisms to affect cell death signaling, and
proand anti-apoptotic B-cell lymphoma 2 proteins (14). Subsequently, the caspase signaling
pathway is altered, contributing to apoptosis (15). Therefore, targeting the
MAPK-regulated signaling pathway may be of potential value in
UVB-induced apoptosis.
Liquiritin, a major constituent of Glycyrrhiza
Radix, has various pharmacological activities (16). A previous study reported that
liquiritin may exert neuroprotective effects in cerebral
ischemia/reperfusion-induced brain damage through antioxidant and
anti-apoptotic mechanisms (17).
As reported previously, liquiritin ameliorates tissue injury
through suppressing MAPK and TLR4/MyD88 signaling pathways, which
reduces the inflammatory response (18). In addition, liquiritin suppresses
the progression of breast cancer, with a reliance on the regulation
of ROS (19). Therefore, the
present study hypothesized that liquiritin may be effective in
preventing skin injury induced by UVB, which is associated with
inflammation, ROS and apoptosis. However, until now, the role of
liquiritin in this regard remains to be elucidated. The present
study indicated for the first time, to the best of our knowledge,
that liquiritin can be used as an effective compound to protect the
skin from damage by UVB in viv and in vitro.
Liquiritin pre-treatment suppressed UVB-induced inflammation,
oxidative stress and apoptosis via inactivating the
TLR4/MyD88/NF-κB and MAPK/caspase signaling pathways.
Materials and methods
Animals and treatments
A total of 110 female, 6-8-week-old SKH-1 hairless
mice (18–20 g) were purchased from Shanghai Laboratory Animal
Research Center (Shanghai, China). The mice were acclimatized for 1
week prior to experiments in the specific pathogen-free conditions
in static microisolator cages with tap water ad libitum, and
were maintained under standard conditions of a 12 h dark/12 h light
cycle (8:00 a.m. to 8:00 p.m.) at a temperature of 23±2°C and
relative humidity of 50±5%. All animal experiments were performed
following the Guide for the Care and Use of Laboratory Animals,
issued by the National Institutes of Health in 1996 and approved by
Huai'an First People's Hospital, Nanjing Medical University
(Nanjing, China). The care and handling of mice were in accordance
with the ethical guidelines of Huai'an First People's Hospital. The
SKH-1 hairless mice were randomly divided into four groups with 15
mice in each group cage. The mice were exposed to UVB lamps
(GL20SE; Sankyo Denki Co., Ltd., Hiratsuka, Japan) equipped with a
controller to modulate UV dosage, at a distance of 20 cm between
the target skin and the light source. The treatment groups,
containing 15 animals in each group, were as follows: Group 1,
untreated animals (Con); Group 2, animals irradiated with UVB only
(UVB); Group 3, UVB irradiation with application of liquiritin (10
mg/kg) by gavage (UVB+10); Group 4, UVB irradiation with
application of liquiritin (20 mg/kg) by gavage (UVB+20) following
UVB (50 mJ/cm2) for 30 min. The murine skin exposure was
then performed at 50 mJ/cm2 of UVB, three times per week
for a consecutive 10-week period. Following another 7 weeks of
exposure with or without liquiritin treatment, serum samples of
animals were collected through eye bleeding 24 h following the
final UVB irradiation, and the dorsal skin tissues from mice were
excised and collected for following analysis (Fig. 1A). In addition, 30 mice were
treated with liquiritin (25, 50 and 100 mg/kg) in the absence of
UVB irradiation to investigate the hepatotoxicity of liquiritin in
mice. The mice treated with liquiritin were pre-treated at the
described concentrations for 2 weeks. The liquiritin (purity ≥98%)
used in the present study was purchased from the Chinese Institute
for the Control of Pharmaceutical and Biological Products (Beijing,
China).
Cell culture and treatments
Human epidermal cells (HACAT) and the L02 human
liver normal cell line were purchased from the American Type
Culture Collection (Manassas, VA, USA) and Nanjing KeyGEN Biotech
Co., Ltd. (Nanjing, China), respectively. The cells were cultured
in DMEM supplemented with 10% FBS (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), penicillin (100 U/ml) and
streptomycin (100 μg/ml) at 37°C in an atmosphere of 5%
CO2. The HACAT cells were cultured until 80% confluence
and then pretreated with various concentrations of liquiritin (40
and 80 μM). Following incubation for 2 h, the culture medium
was replaced with 1.5 ml of phosphate-buffered saline (PBS).
Subsequently, the HACAT cells were exposed to UVB (15
mJ/cm2) light at a 312 nm light source for 1 h.
Following UVB-exposure, the cells were treated with various
concentrations of liquiritin (40 and 80 μM) in serum-free
medium for another 22 h. The cells were then harvested for further
examination.
Cell viability analysis
The HACAT human epidermal cell line and L02 human
liver normal cell line were initially seeded in 96-well plates at a
density of 2×104 cells/well, respectively, prior to
incubation for 24 h at 37°C. The cell culture media were then
replaced with complete media containing the indicated
concentrations (0, 1.25, 2.5, 5, 10, 20, 40, 80 and 160 μM)
of liquiritin, prior to incubation for the indicated duration (0,
6, 12, 24, 36, 48, 72 and 96 h). Following incubation as indicated,
10 μl of MTT (Nanjing KeyGEN Biotech, Co., Ltd.) was
administrated to cells, followed by incubation for 4 h at 37°C
according to the manufacturer's protocol. Finally, the absorbance
was read at 570 nm on a microplate reader. The cell viability (%)
was evaluated as the ratio of surviving cells.
Crystal violet staining
Following the various treatments, the HACAT cells
were harvested and a Crystal Violet Staining Solution kit (Beyotime
Institute of Biotechnology, Haimen, China) was used to analyze the
HACAT cell proliferation, according to the manufacturer's
protocol.
Analysis of chemical indicators
The activities of enzymatic antioxidants in serum
and skin tissues, including superoxide dismutase (SOD) and catalase
(CAT), were analyzed using an SOD assay kit (Nanjing Jiancheng
Bioengineering Institute, Nanjing, China) and a CAT assay kit
(Nanjing Jiancheng Bioengineering Institute), respectively. The
level of malondialdehyde (MDA) in skin tissues was measured using
an MDA assay (Nanjing Jiancheng Bioengineering Institute) kit
according to the manufacturer's protocol. The levels of aspartate
aminotransferase (AST) and alanine aminotransferase (ALT) in serum
were calculated to determine the hepatotoxicity using specific kits
purchased from Nanjing Jiancheng Bioengineering Institute according
to the manufacturer's protocol. The levels of
H2O2 in the skin tissue sections were
calculated using a hydrogen peroxide assay kit (Nanjing Jiancheng
Bioengineering Institute). O2− in the skin
tissue samples was evaluated using the lucigenin chemiluminescence
method. Briefly, the mice skin tissue samples were weighed and
homogenized in homogenization buffer with HEPES and EDTA. Following
centrifugation at 1,000 x g for 10 min at 4°C, an aliquot (200
μl) of the supernatant was further incubated with 5
μM lucigenin in Krebs-HEPES buffer. The light emission was
measured with a Tecan Infinite 200 reader. The specificity for
O2− was evaluated by adding SOD (350 U/ml)
into the incubation medium. Protein content was then measured with
a BCA Protein Quantitative Analysis kit (Thermo Fisher Scientific,
Inc.).
ELISA methods
The skin samples were frozen in liquid nitrogen and
crushed into a powder with a multibead shocker. The powder was then
dissolved in cell lysis buffer for western blot analysis and
immunoprecipiation (Beyotime Institute of Biotechnology) with
protease inhibitor cocktail. The skin extract was prepared by
centrifugation at 12,000 x g for 10 min at 4°C, and the supernatant
was retained for further analysis. The sample protein
concentrations were calculated using a BCA protein assay kit
(Thermo Fisher Scientific, Inc.). The protein levels of IL-1β,
TNF-α, IL-18, IL-6 and COX2 were assessed using respective mouse
ELISA kits (R&D Systems, Inc., Minneapolis, MN, USA) according
to the manufacturer's protocol. Finally, the absorbance was read at
450 nm on a microplate reader.
Analyses of transepidermal water loss
(TEWL), hydration and elasticity
TEWL (g/m2/h) is a marker of epidermal
skin barrier function, and was measured with a Tewameter™ 300
(Courage and Khazaka Electronics GmbH, Köln, Germany). In brief,
under isoflurane anesthesia, a medical adhesive tape (Honsmed,
Shanghai, China) was attached to the mouse skin under a gentle
pressure, following which it was removed. The mice were
tape-stripped five times. TEWL was then recorded with the
Tewameter™ 300 device. The moisture levels of the stratum corneum
were evaluated through a CM825 corneometer (Courage and Khazaka
Electronics GmbH) and the skin elasticity, indicated by the
parameter F3, was assessed with a cutometer of DUAL MPA580 (Courage
and Khazaka Electronics GmbH) according to the manufacturer's
protocols.
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) assays
The TUNEL assay (BioVision, Inc., Milpitas, CA, USA)
was used to measure the apoptotic cells according to the
manufacturer's protocol. The skin sections were treated with
proteinase K (20 μg/ml) at 37°C for 15 min, followed by
incubation with TUNEL reaction mixture at 37°C for 1 h.
Subsequently, each section was exposed to an antibody solution
(anti-BrdU monoclonal antibody; 1:250; cat. no. ab6326; Abcam,
Cambridge, MA, USA) for 30 min at room temperature. The
TUNEL-stained cells were observed using a light microscope (Nikon,
Tokyo, Japan) equipped with an ocular micrometer in five randomly
selected fields per section. The level of apoptosis (%) was
calculated as the average number of stained cells in each field
divided by the total number of cells.
Western blot analysis
The HACAT cells or mice skin tissue samples were
lysed using RIPA buffer containing a 1:100 dilution of protease
inhibitor and phosphatase inhibitor (Baomanbio, Shanghai, China).
The lysates were then centrifuged at 15,000 x g for 15 min at 4°C
to collect the supernatant. The protein concentrations were
evaluated using a BCA protein assay (Thermo Fisher Scientific,
Inc.), and equal protein quantities (40 μg) were separated
using 10% SDS-PAGE. The proteins were then electrophoretically
transferred onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA), and then incubated with
Tris-buffered saline containing 0.1% Tween-20 with 5% skim milk (BD
Biosciences, Franklin Lakes, NJ, USA) at room temperature for 2 h.
Primary antibodies dissolved in blocking buffer were used to detect
the target protein blots at 4°C overnight. The following antibodies
were used: Rabbit anti-phosphorylated (p-)P38 (1:1,000; cat. no.
4511; Cell Signaling Technology, Inc., Danvers, MA, USA), rabbit
anti-P38 (1:1,000; cat. no. 8690; Cell Signaling Technology, Inc.),
rabbit anti-poly(ADP-ribose) polymerase (PARP) (1:1,000; cat. no.
9532; Cell Signaling Technology, Inc.), rabbit anti-TLR4 (1:1,000;
cat. no. 14358; Cell Signaling Technology, Inc.), rabbit anti-MyD88
(1:1,000; cat. no. 4283; Cell Signaling Technology, Inc.), and
rabbit anti-p-inhibitor of NF-κB kinase (IKK)α (1:1,000; cat. no.
2682; Cell Signaling Technology, Inc.), rabbit anti-inhibitor of
NF-κB (IκB)α (1:500; cat. no. 4814; Cell Signaling Technology,
Inc.), rabbit anti-p-JNK (1:1,000; cat. no. ab4821; Abcam), rabbit
anti-JNK (1:1,000; cat. no. ab131499; Abcam), rabbit anti-caspase-3
(1:1,000; cat. no. ab13847; Abcam), rabbit anti-xanthine oxidase
(XO) (1:1,000; cat. no. ab133268; Abcam), rabbit anti-inducible
nitric oxide synthase (iNOS) (1:1,000; cat. no. ab15323; Abcam),
rabbit anti-NOX2 (1:1,000; cat. no. ab80508; Abcam), rabbit
anti-NOX4 (1:1,000; cat. no. ab216654; Abcam), rabbit anti-NF-κB
(1:1,000; cat. no. ab207297; Abcam), rabbit anti-p-NF-κB (1:1,000;
cat. no. ab86299; Abcam), mouse anti-caspase-9 (1:1,000; cat. no.
ab32539; Abcam) and anti-GAPDH (1:1,000; cat. no. ab8245; Abcam).
Then, the membranes were washed with tris-buffered saline with
Tween-20 (1%) three times, followed by incubation with a goat-anti
rabbit horseradish peroxidase-conjugated secondary antibody
(1:2,500; cat. no. ab6721; Abcam) at room temperature for 2 h. The
bands on the membrane were analyzed using chemiluminescence with
Pierce ECL Western Blotting Substrate reagents (Thermo Fisher
Scientific, Inc.). Protein expression levels were assigned a grey
value using ImageJ 1.38 software (National Institutes of Health,
Bethesda, MD, USA) and standardized to the housekeeping gene GAPDH
and expressed as a fold of the control. All experiments were
performed in triplicate and performed three times
independently.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA from the cells and skin tissue samples was
extracted using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Subsequently, RNA were quantified and subjected to reverse
transcription to prepare cDNA using a RevertAid First Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc.). For analysis,
single-stranded cDNA was prepared from RNA by reverse transcription
using oligo(dT) primers (SunShine Biotechnology International Co.,
Ltd., Somerset, NJ, USA). The RT reaction contained RNA samples
including 2 μg purified total RNA, 2 μl 10 μM
oligo(dT) primers, 5 μl 5xRT buffer (Promega Corporation,
Madison, WI, USA), 1 μl 40 U/μl MultiScribe reverse
transcriptase (Promega Corporation), 1.25 μl 10 mM dNTPs,
0.625 μl 40 U/μl RNase inhibitor (SunShine
Biotechnology International Co., Ltd.), and 25 μl
H2O-diethyl pyrocarbonate. qPCR was performed on a CFX96
Real-Time system (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
The sequences of primers were commercially synthesized and the
sequences of the primers are listed in Table I. The reaction conditions were as
follows: denaturation at 95°C for 10 min, followed by 40 cycles of
amplification and quantification at 95°C for 30 sec, 60°C for 30
sec and 72°C for 1 min. The melt curve conditions were as follows:
95°C for 15 sec, 60°C for 15 sec and 95°C for 15 sec. The mRNA
expression normalized to the expression of the housekeeper GAPDH
was measured using the 2−∆∆Cq method (20).
 | Table ISequences of primers used in reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I
Sequences of primers used in reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Primer sequence
(5′-3′) |
|---|
| Mouse-TNF-α | F:
AGATATTACAGTGCCATGGTT |
| R:
ACGAGTAGAGATCTCGAG |
| Mouse-COX2 | F:
CCAGACTTGGCTACCTCGTG |
| R:
CTGACTTAGTCGATAATTCT |
| Mouse-SOD1 | F:
GCCATTCCTGTCTCGTGGAG |
| R:
TAATGTATTGATCCTTGATTAT |
| Mouse-IL-18 | F:
AGTAAGTGGCATTACCGAC |
| R:
ACCAACGCAACAGTCTGCAG |
| Mouse-IL-6 | F:
GTCAAGACCACAGCTAGC |
| R:
CTGGCGACTACTAGTAGATA |
| Mouse-IL-1β | F:
ACTCGAGAGGAATCCTTACGA |
| R:
CACTCTCGTGACTCGCTA |
| Mouse-CAT | F:
CCTCGTTCACACTTCGTGTA |
| R:
GGAAGCACTCCAGTGCAGC |
| Mouse-Nrf2 | F:
CTACAGACCAATGCCTGAC |
| R:
ACGTAACTGTGCCATGGGGA |
| Mouse-SOD2 | F:
AGGCCGTGAGAGCTTGTGTA |
| R:
CGGCAGTAAGTGCCCTCTAC |
| Mouse-GAPDH | F:
GCGAGCTGAGACACTCTAG |
| R:
TAGGCATGCACCTCTGTTCA |
| Human-TNF-α | F:
ATATGTGCTGGTCATACTCAT |
| R:
AGAGATCGGCGTCAGATGA |
| Human-COX2 | F:
CCTTGCAGAGCTACCGAGT |
| R:
GTGACTTAGTATATTCATCC |
| Human-SOD1 | F:
GTCCTACGGTCTCGTGAGATAT |
| R:
GTTAATATTGGTAGTTCTCTG |
| Human-IL-18 | F:
AGCATTACCGACCTATTCCT |
| R:
AGCACCCAACGCAACAAGTG |
| Human-IL-6 | F:
CAGTAGAAGACCAAGTACAT |
| R:
CTGGCGACTACTTAGCTATAA |
| Human-IL-1β | F:
AATTGAGAGCTCGGATCCGT |
| R:
CTGACGCTATCACGTAGAGCA |
| Human-CAT | F:
CTCACTACGTCGTGTATTCCCT |
| R:
GTGAAGCACTCCAGTGCCAG |
| Human-Nrf2 | F:
CTCCAATGCCTGATCGCGCT |
| R:
AGGCCATGGCAAGTGTGAT |
| Human-SOD2 | F:
CGTGAGGCGAGCTACAGAA |
| R:
GGTAAGTTACGCTCCAGCCTC |
| Human-GAPDH | F:
GGACCGAGTACTGCATCAGCTA |
| R:
TCCTCGTGAAGGTTGACTGCGA |
Immunohistochemical analysis
The skin tissues obtained from the mice were fixed,
embedded in paraffin blocks and cut into 3-μm thick
sections. The skin sections were deparaffinized and stained with
hematoxylin and eosin (H&E) staining. The thickness of the skin
epidermis was assessed using Magnuspro software (Magnus pro 3.0
software; Olympus Corporation, Tokyo, Japan). The epidermal
thickness of the H&E-stained sections was assessed using
Image-J software (1.47v version; National Institutes of Health,
Bethesda, MD, USA). For immunohistochemical images, the skin tissue
sections were then exposed to HCl (3.5 M) for 20 min at room
temperature and washed using PBS three times. Subsequently, the
skin tissue sections were treated with peroxidase (0.3%) to block
endogenous peroxidase activity. The tissue sections were then
incubated with normal goat serum (5%; cat. no. 5425; Cell Signaling
Technology, Inc.) for 30 min, followed by incubation with primary
antibodies TLR4 and NOX-2; Abcam, Cambridge, MA, USA) at 1:100
dilution for 2 h at room temperature. The sections were then
incubated with HRP-conjugated compact polymer systems.
Diaminobenzidine (Chem Service, West Chester, PA, USA) was used as
the chromogen according to the manufacturer's protocol.
Statistical analysis
Data are expressed as the mean ± standard error of
the mean. The statistical analysis was performed using one-way
analysis of variance with Tukey's post-hoc test (on GraphPad Prism
6.01; GraphPad Software, Inc., La Jolla, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Liquiritin improves UVB-induced skin
injury in mice
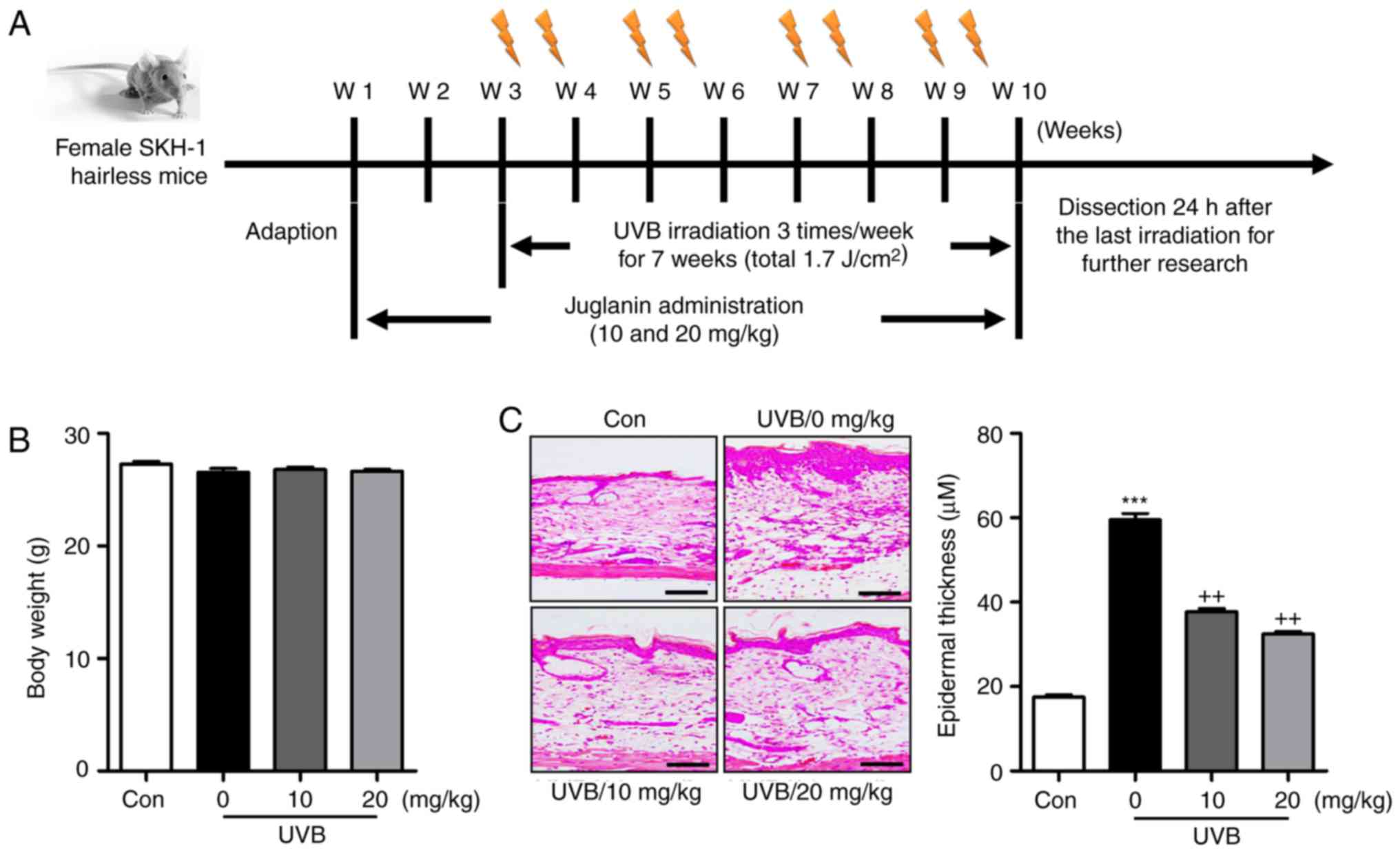
As shown in Fig.
1B, the present study first examined the body weights of mice
treated under different conditions to assess the role of liquiritin
in mice. The results indicated there was no significant difference
in the body weight of mice treated with various concentrations of
liquiritin with UVB exposure, indicating that the doses used in the
present study, at least in part, were safe for the animals.
Subsequently, the H&E staining suggested that the epidermal
thickness of the skin of the mice was higher in the UVB-treated
mice in absence of liquiritin, compared with that of mice in the
Con group, and was reduced in the mice exposed to liquiritin
treatments, compared with that in the UVB-only group of mice
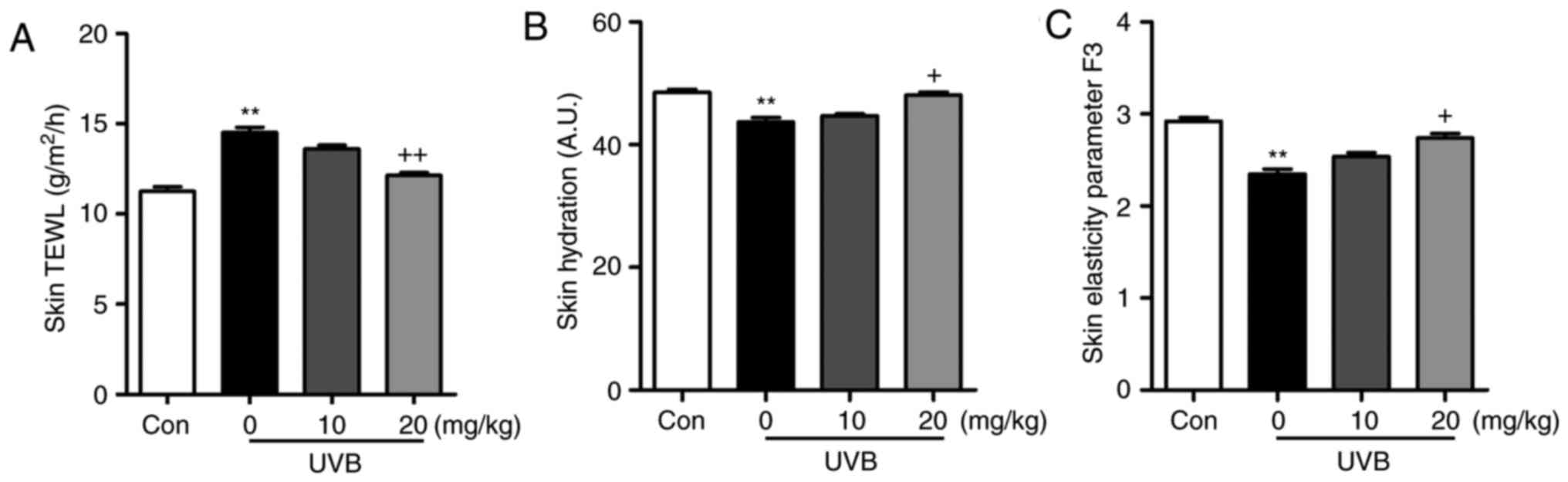
(Fig. 1C). Furthermore, the skin
TEWL, skin hydration and skin elasticity parameter F3 were highly
induced in the UVB-irradiated mice, which were comparable to those
in the Con group. Following liquiritin treatments, these indicators
were reduced (Fig. 2A–C). These
data indicated that liquiritin had a potential role in ameliorating
skin injury in mice exposed to UVB.
Liquiritin reduces the inflammatory
response in UVB-induced skin in mice
The inflammatory response has been characterized as
an essential factor in accelerating the progression of various
diseases, including liver injury, heart fibrosis and several types
of cancer (21). In the present
study, the levels of pro-inflammatory cytokines in the skin tissue
samples were measured and showed that IL-1β (Fig. 3A), TNF-α (Fig. 3B), IL-18 (Fig. 3C), IL-6 (Fig. 3D) and COX2 (Fig. 3E) were released at high levels in
the skin of mice exposed to UVB, which were significantly reduced
by liquiritin administration in a dose-dependent manner. The
results of the RT-qPCR analysis also indicated that the mRNA levels
of IL-1β, TNF-α, IL-18, IL-6 and COX2 were expressed at high levels
in the UVB-treated mice. Similar results were observed in the
protein levels, determined using western blot analysis. Of note,
liquiritin exerted a suppressive effect on the altered release of
these signals, with levels comparable to those in the Con group of
mice (Fig. 3F and G). These data
indicated that liquiritin improved UVB-induced skin injury through
reducing the release of pro-inflammatory cytokines.
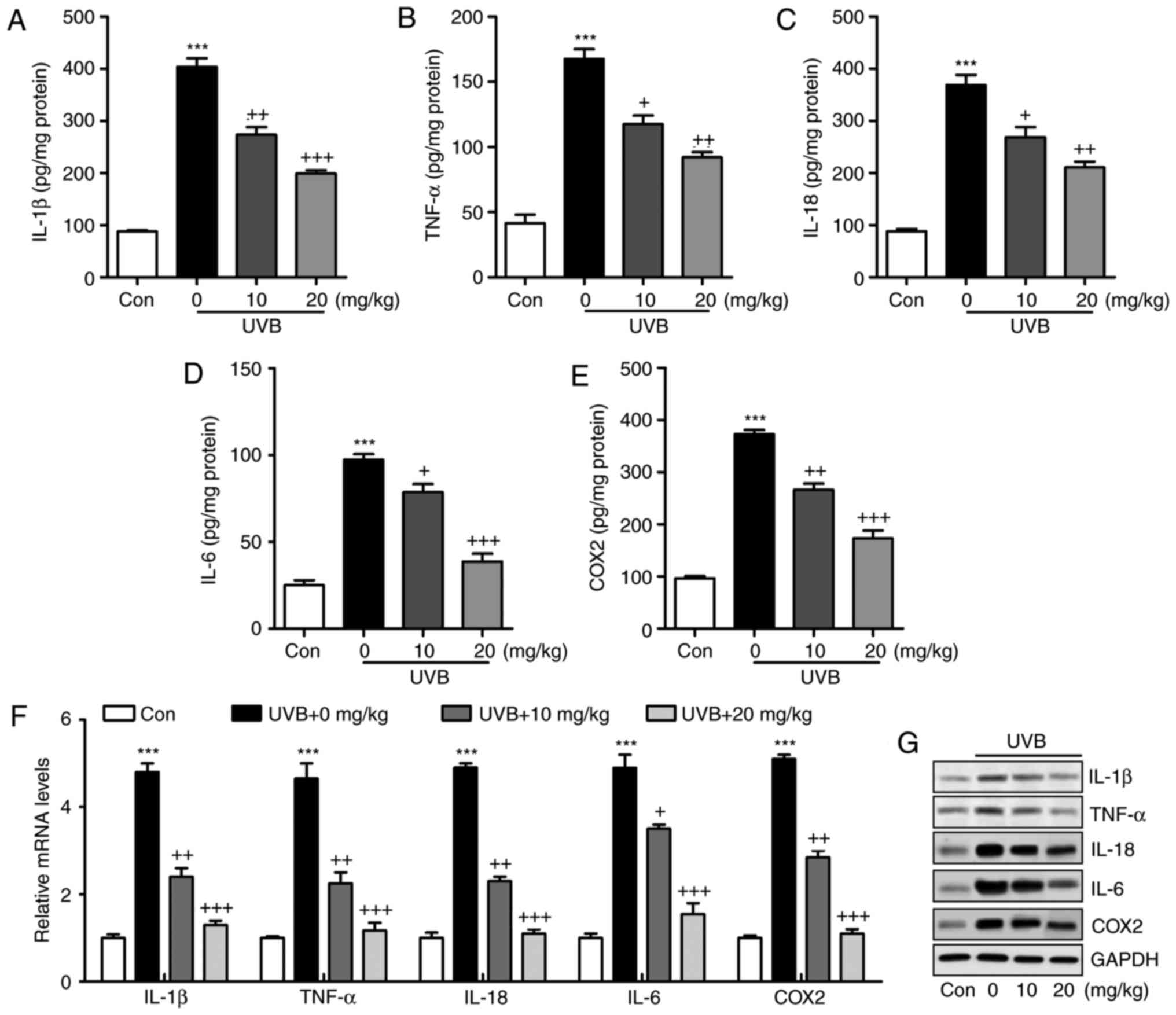 | Figure 3Liquiritin reduces the release of
pro-inflammatory cytokines in UVB-induced skin in mice. Expression
of pro-inflammatory cytokines (A) IL-1β, (B) TNF-α, (C) IL-18, (D)
IL-6 and (E) COX2 in the skin tissue samples were calculated using
ELISA methods. (F) Reverse transcription-quantitative polymerase
chain reaction and (G) western blot assays were used to evaluate
the gene and protein levels of IL-1β, TNF-α, IL-18, IL-6 and COX2
in the skin tissue of mice obtained from mice induced by UVB. Data
are presented as the mean ± standard error of the mean (n=10).
***P<0.001, compared with the Con group.
+P<0.05, ++P<0.01 and
+++P<0.001, compared with the UVB-only group. UVB,
ultraviolet B; Con, control; IL, interleukin; TNF-α, tumor necrosis
factor-α; COX2, cyclooxygenase 2. |
Activation of the TLR4/MyD88 signaling pathway can
lead to the direct activation of NF-κB, contributing to
pro-inflammatory cytokine secretion (22). The TLR4/MyD88 and NF-κB signaling
pathways were investigated in the present study. As shown in
Fig. 4A, the immunohistochemical
analysis indicated that the level of TLR4 was induced in the skin
tissue sections of mice exposed to UVB, which was reduced by
liquiritin administration. Similar results were observed for TLR4
and MyD88 protein levels using western blot analysis (Fig. 4B). Additionally, the protein
levels of p-IκBα, p-IKKα, and p-NF-κB were high in the UVB-only
group. By contrast, IκBα was reduced under UVB exposure. Liquiritin
treatment significantly decreased the phosphorylation of IKKα,
IκBα, and NF-κB, and increased the levels of IκBα in a
dose-dependent manner (Fig. 4C).
Together, these data indicated that the liquiritin-ameliorated
inflammation induced by UVB was dependent on TLR4/MyD88/NF-κB
inactivation.
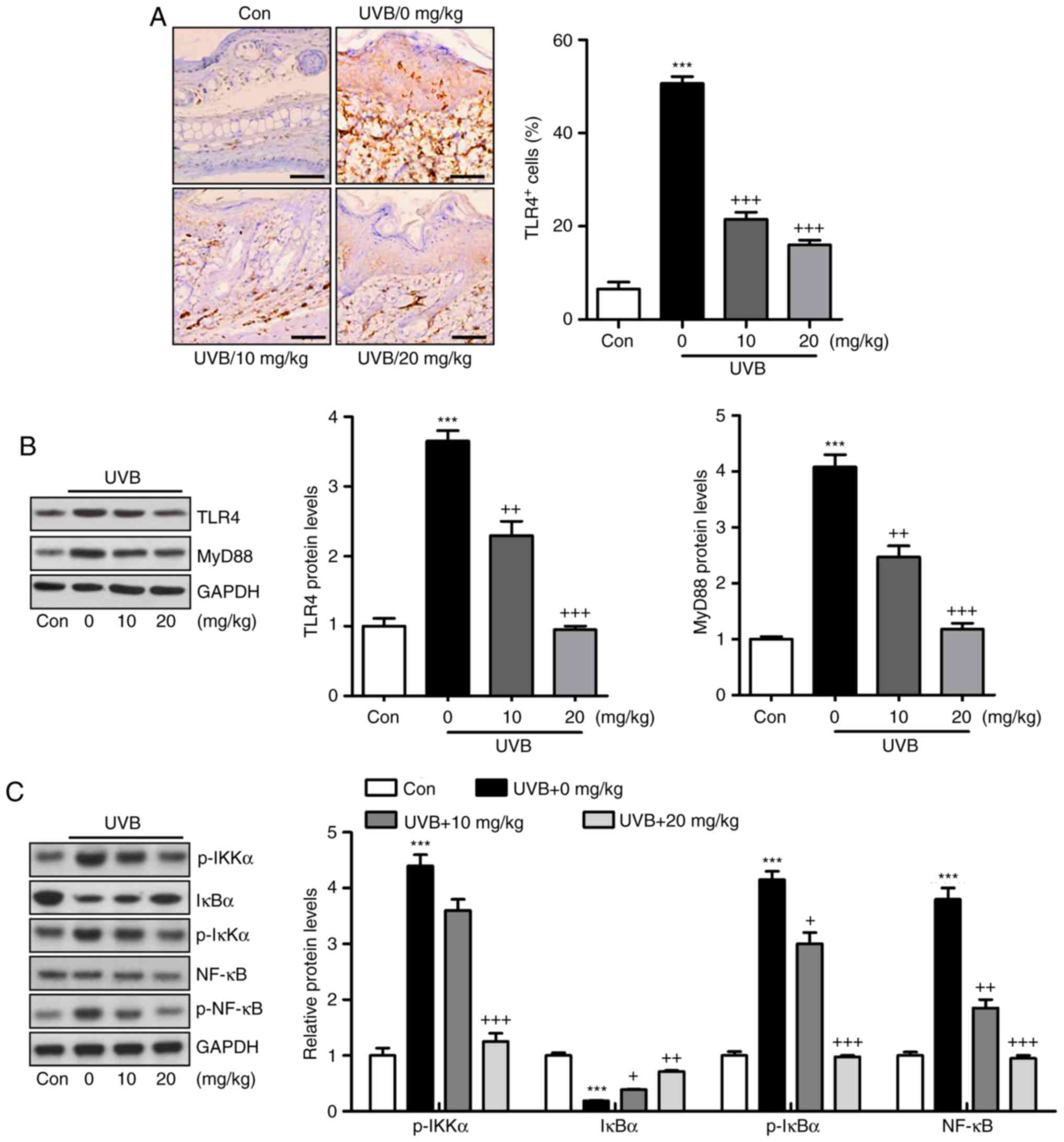 | Figure 4Liquiritin-reduced pro-inflammatory
cytokines release is dependent on the TLR4/NF-κB signaling pathway.
(A) Immunohistochemical analysis was performed to assess
TLR4-positive cells in various groups of mice exposed to UVB
irradiation. Scale bar, 100 μm. (B) Western blot analysis
was performed to determine the expression levels of TLR4 and MyD88,
shown in the representative images and quantification in the
histogram. (C) Protein levels of p-IKKα, IκBα, p-IκBα, and p-NF-κB
were assessed using western blot analysis, and quantification of
these proteins is shown in the histogram. Data are presented as the
mean ± standard error of the mean (n=10). ***P<0.001,
compared with the Con group. +P<0.05,
++P<0.01 and +++P<0.001, compared with
the UVB-only group. UVB, ultraviolet B; Con, control; TLR4,
toll-like receptor 4; MyD88, myeloid differentiation factor 88;
NF-κB, nuclear factor-κB; IκBα, inhibitor of NF-κB α; IKKα, IκB
kinase α; p-, phosphorylated. |
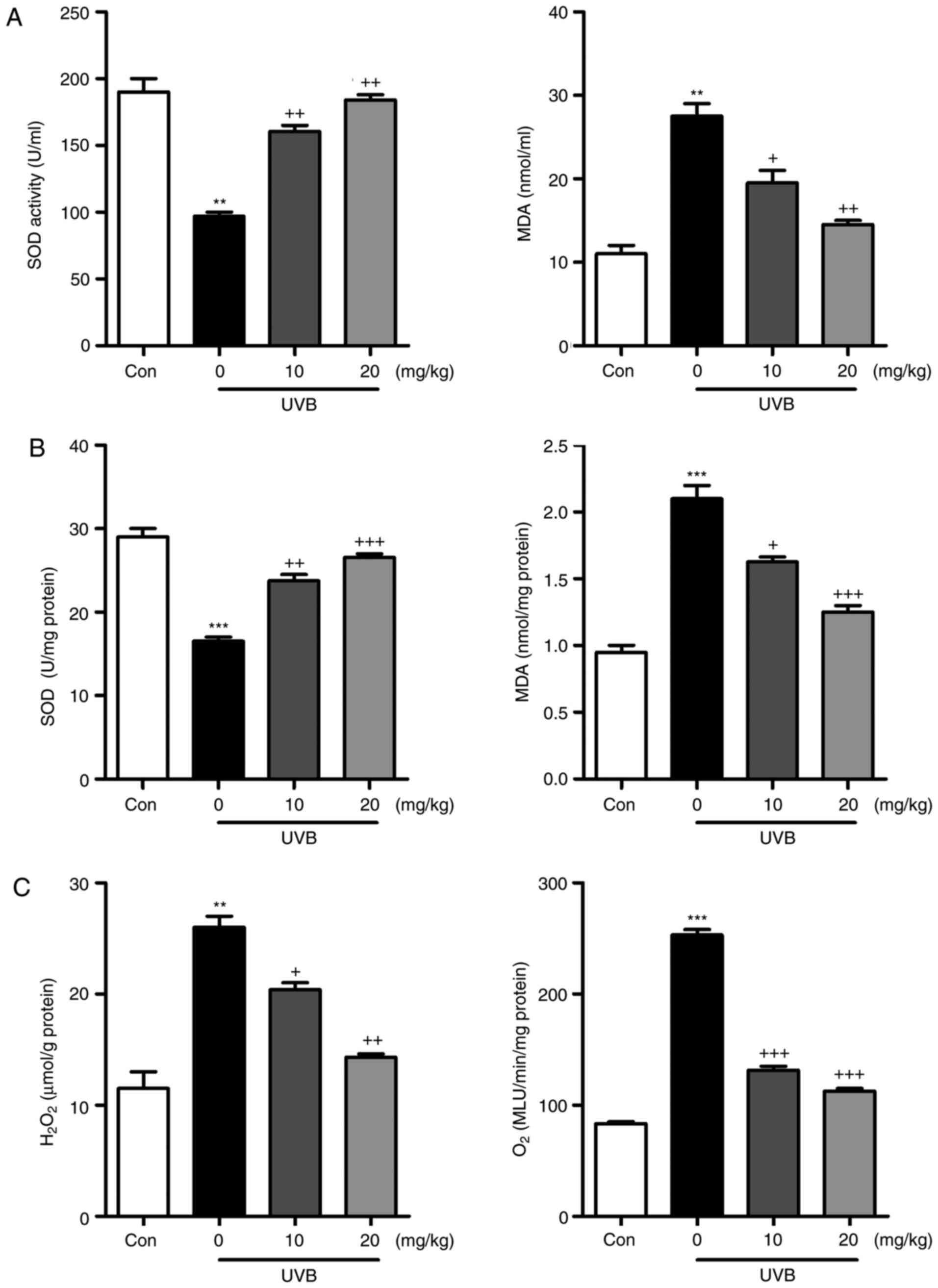
Liquiritin inhibits UVB-induced oxidative
stress in mice
Oxidative stress is reported to be key in the
induction of skin injury induced by UVB (11,12). Therefore, antioxidants and
oxidants were measured in the present study. As shown in Fig. 5A and B, it was found that the
activity of SOD in serum and skin tissue samples of mice exposed to
UVB was lower, compared with that in the Con group of mice. Of
note, liquiritin treatment upregulated the activity of SOD in the
serum and in the skin tissue samples. By contrast, the levels of
MDA were found to be higher following UVB exposure, which were
downregulated due to liquiritin administration. The levels of
antioxidants H2O2 and O .2 in the
skin were found to be accelerated in the UVB-treated mice, and were
reduced by liquiritin treatment (Fig.
5C). Oxidative stress was present in the skin of mice according
to these results, which was reversed following liquiritin
administration. Subsequently, the present study attempted to
examine the molecular mechanism by which liquiritin exerted its
effect in reducing oxidative stress. As shown in Fig. 6A, RT-qPCR analysis indicated that
the gene expression levels of SOD1, SOD2, CAT and nuclear factor
erythroid 2-related factor 2 (Nrf2), which are important
antioxidants for inhibiting oxidants and suppressing ROS, were
decreased following UVB irradiation. In the liquiritin-treated
groups, the expression of these signals was increased, further
indicating that liquiritin is potentially involved in ameliorating
oxidative stress via enhancing the expression of antioxidants. The
molecules XO, iNOS, NOX-4 and NOX-2, which are crucial in promoting
ROS generation, were induced under UVB exposure. Liquiritin
treatment at various concentrations significantly reduced the
expression of these molecules, demonstrating its inhibitory effects
on ROS (Fig. 6B). NOX-2 is known
to be important in contributing to ROS generation. Therefore,
immunohistochemical analysis was performed to further investigate
how liquiritin affects the expression of NOX-2 in UVB-induced skin
injury (Fig. 6C). The
representative images indicated that liquiritin reduced the
UVB-induced levels of NOX-2 positive cells, which was in accordance
with the results of western blot analysis. Taken together, the
results indicated that liquiritin reduced oxidative stress to
ameliorate UVB-induced skin injury in vivo.
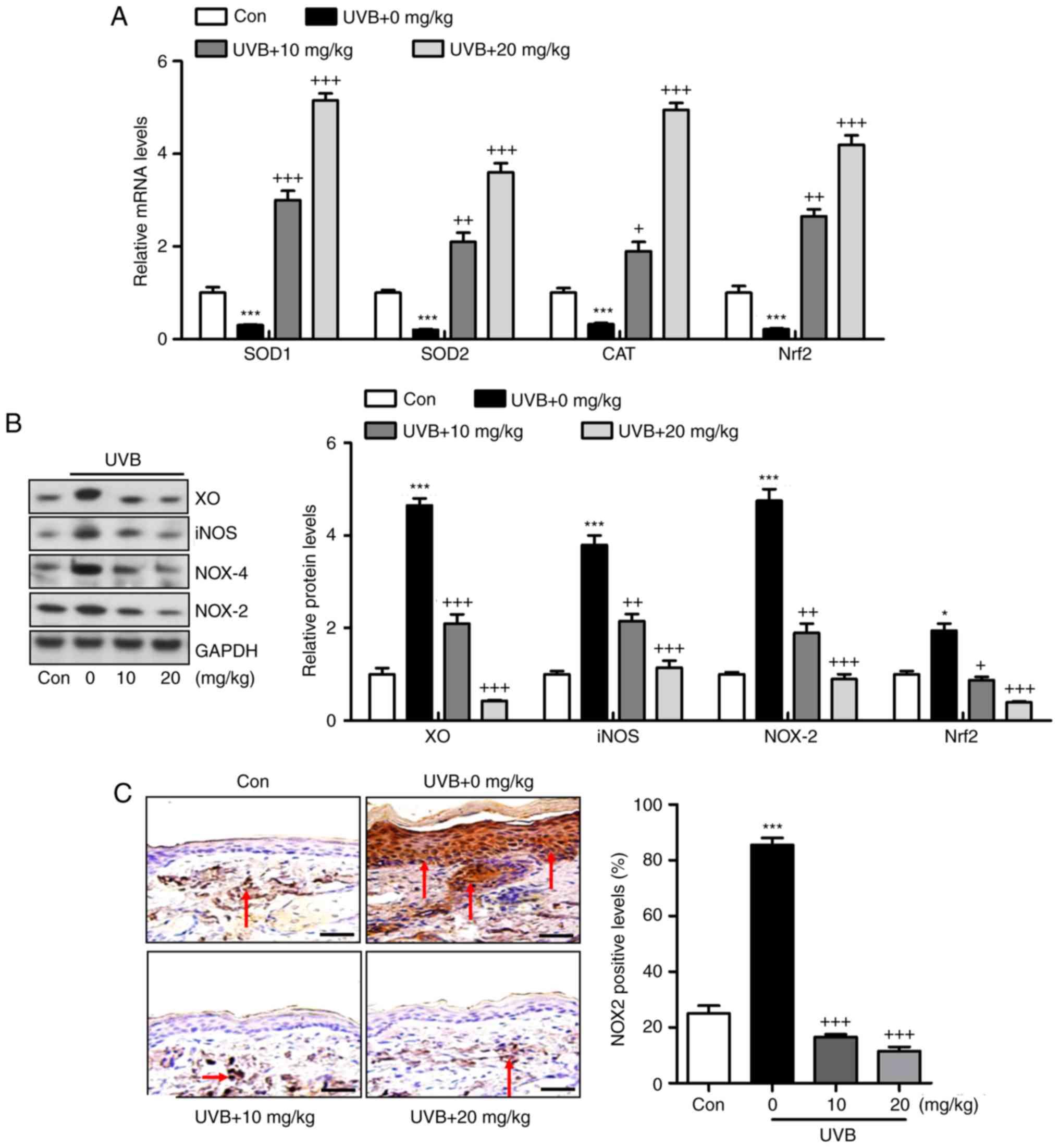 | Figure 6Liquiritin reduces oxidative stress
via promoting the expression of antioxidants and suppressing the
expression of oxidants in the skin tissue samples. (A) mRNA levels
of SOD1, SOD2, CAT and Nrf2 in the skin tissue specimens were
calculated using reverse transcription-quantitative polymerase
chain reaction analysis. (B) Western blot assays were performed to
examine protein levels of XO, iNOS, NOX-2 and NOX-4 in the skin
tissue samples of mice treated under various conditions. (C)
Expression levels of NOX-2 were measured using immunohistochemical
analysis in the skin tissue sections from mice. The red arrows
refer to NOX2-positive cells. Scale bar, 100 μm. Data are
presented as the mean ± standard error of the mean (n=10).
***P<0.001, compared with the Con group.
+P<0.05, ++P<0.01 and
+++P<0.001, compared with the UVB-only group. UVB,
ultraviolet B; Con, control; SOD, superoxide dismutase; CAT,
catalase; Nrf2, nuclear factor erythroid 2-related factor 2; XO,
xanthine oxidase; iNOS, inducible nitric oxide synthase; NOX,
nitric oxide. |
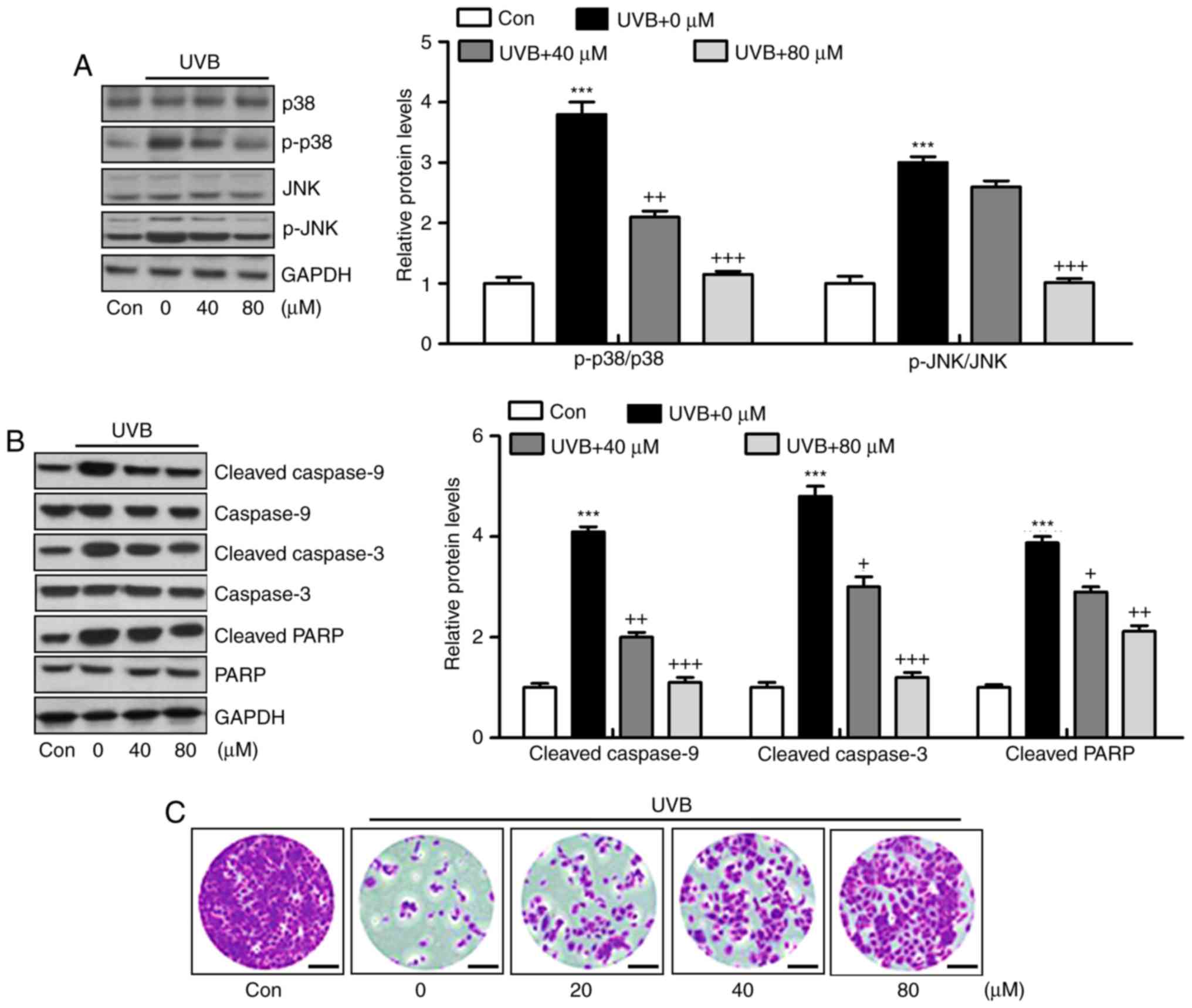
Liquiritin ameliorates UVB-induced skin
injury dependent on MAPK and caspase signaling pathways
Liquiritin has been reported to inhibit cancer
progression via the modulation of apoptosis (23). Therefore, it was hypothesized that
liquiritin may improve UVB-induced skin injury associated with the
apoptotic response. The MAPK signaling pathway is known to affect
apoptosis under various conditions (24). In the present study, it was found
that MAPKs, p38 and JNK were phosphorylated by UVB in the skin
tissue samples from mice, and were reduced by liquiritin treatment
(Fig. 7A). The caspase signaling
pathway is known to regulate the apoptotic response (25). The results of western blot
analysis indicated that cleaved caspase-9, caspase-3 and PARP were
expressed at high levels in the UVB-treated mice, suggesting that
apoptosis was induced under UVB exposure, leading to eventual cell
death. However, liquiritin inhibited the activation of these
signals, suppressing apoptosis and cell death (Fig. 7B). Finally, TUNEL analysis
directly indicated that liquiritin reduced UVB-induced apoptosis
(Fig. 7C). Together, these data
indicated that UVB-induced skin injury was closely associated with
apoptosis, which was inhibited by liquiritin treatment.
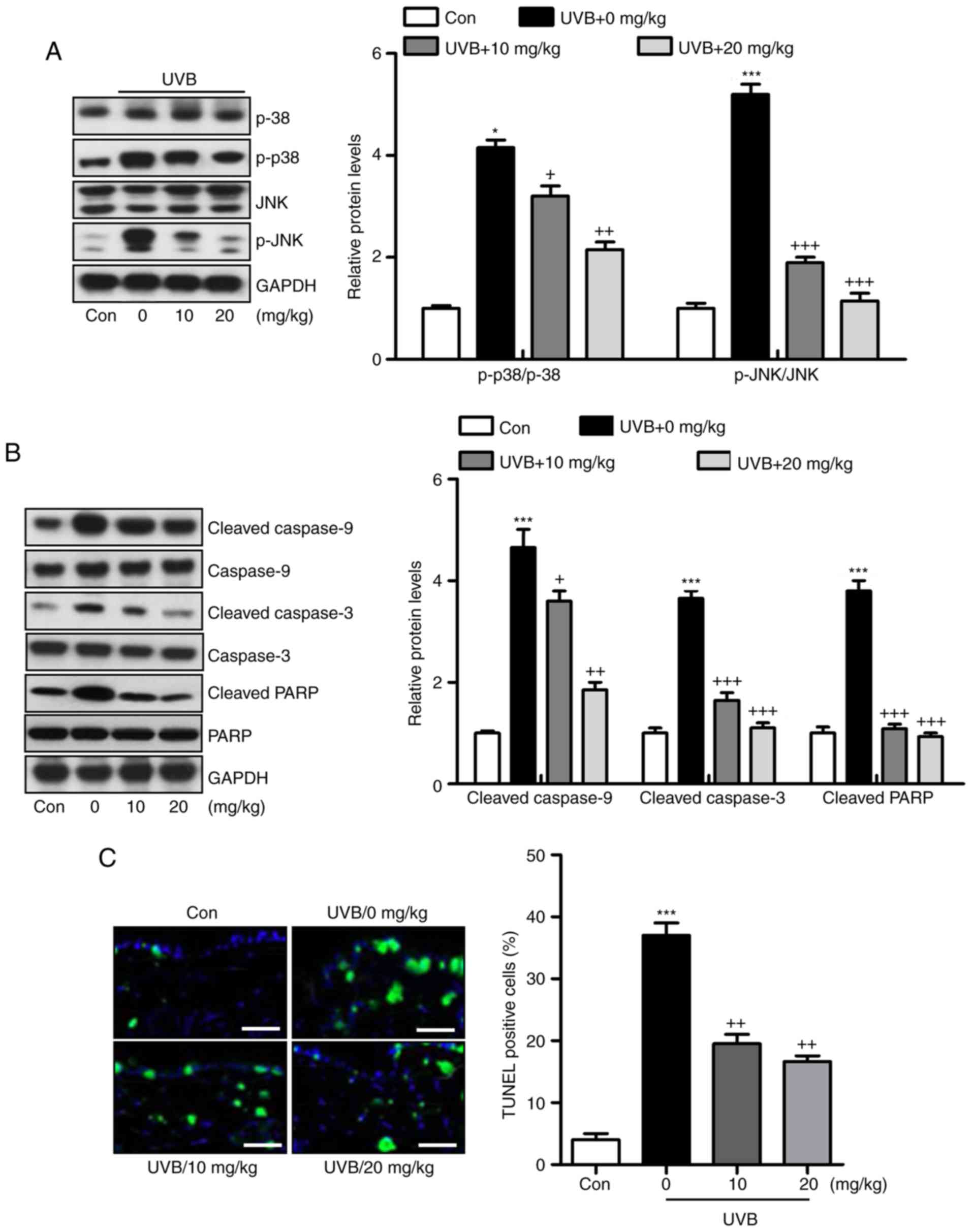 | Figure 7Liquiritin ameliorates UVB-induced
skin injury dependent on mitogen-activated protein kinase and
caspase signaling pathways. (A) Expression levels of p-p38 and
p-JNK were assessed using western blot analysis. (B) Immumoblotting
analysis was performed to evaluate the levels of cleaved caspase-9,
caspase-3 and PARP in the skin tissue samples isolated from mice
exposed to UVB irradiation treated with or without liquiritin. (C)
TUNEL-positive levels were evaluated in the skin tissue sections of
mice to examine the effect of liquiritin on apoptosis induced by
UVB. Scale bar, 100 μm. Data are presented as the mean ±
standard error of the mean (n=10). *P<0.05 and
***P<0.001, compared with the the Con group.
+P<0.05, ++P<0.01 and
+++P<0.001, compared with the UVB-only group. UVB,
ultraviolet B; Con, control, JNK, c-Jun N-terminal kinase; PARP,
poly(ADP-ribose) polymerase; p-, phosphorylated; TUNEL, Terminal
deoxynucleotidyl transferase dUTP nick end labeling. |
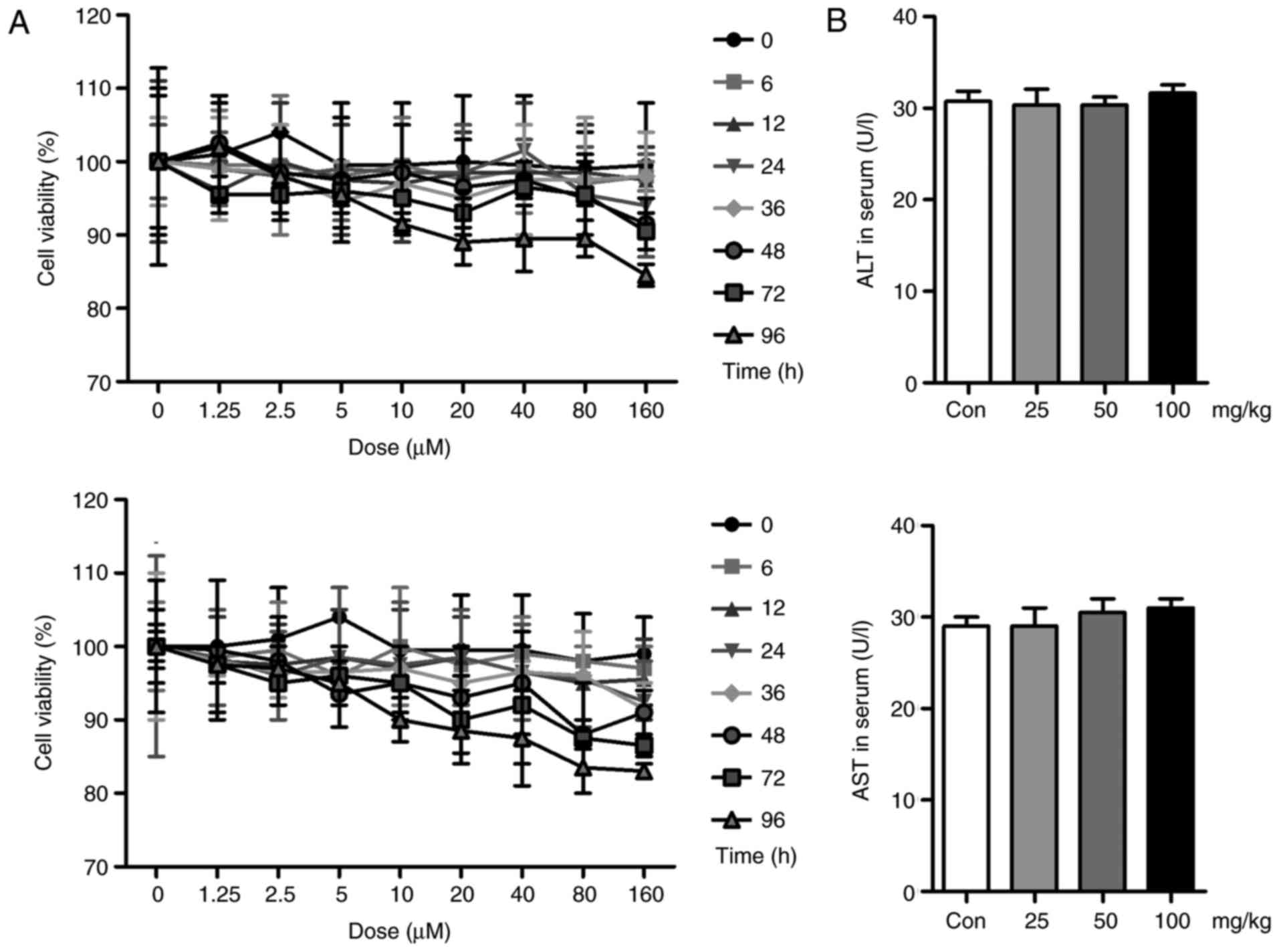
Liquiritin has no significant effect on
cell viability and hepatotoxicity
In vivo, liquiritin has been shown to
ameliorate UVB-induced skin injury. In the present study, in
vitr experiments were performed to further examine the effects
of liquiritin on UVB-induced skin damage. First, the cell viability
was assessed to investigate the safety or cytotoxicity of
liquiritin in cells. As shown in Fig.
8A, the HACAT and L02 cells were exposed to various
concentrations of liquiritin for different durations. However, no
significant difference was observed among the different groups,
with the exception of the highest dose of liquiritin in the L02
cells for the longest duration (96 h). These data, in part, showed
that liquiritin at the concentrations and durations examined in the
present study was not cytotoxic to cells. Additionally, no
significant difference was found in the levels of ALT and AST in
the serum of mice treated with different doses of liquiritin,
compared with those in the Con group, further indicating the safety
of liquiritin (Fig. 8B).
Liquiritin reduces the inflammatory
response in UVB-induced HACAT cells in vitro
The HACAT cells were irradiated with UVB and exposed
to liquiritin treatment at different concentrations, as indicated,
for analysis. The results of the RT-qPCR analysis suggested that
the mRNA levels of IL-1β, TNF-α, IL-18, IL-6 and COX2 were high in
the UVB-irradiated cells, and were downregulated by liquiritin
treatment, which was in accordance with the results of the in
viv experiments (Fig. 9A). In
addition, the UVB-induced higher protein levels of TLR4 and MyD88
were reduced by liquiritin treatment in vitro, further
confirming that the TLR4/MyD88 signaling pathway was involved in
the liquiritin-ameliorated skin injury induced by UVB (Fig. 9B). In addition, the phosphorylated
levels of IKKα, IκBα, and NF-κB proteins were upregulated in the
UVB-only group. Similar to the in viv experiment, IκBα was
reduced under UVB exposure. Liquiritin treatment at various
concentrations decreased the phosphorylation of IKKα, IκBα and
NF-κB, and increased the level of IκBα (Fig. 9C).
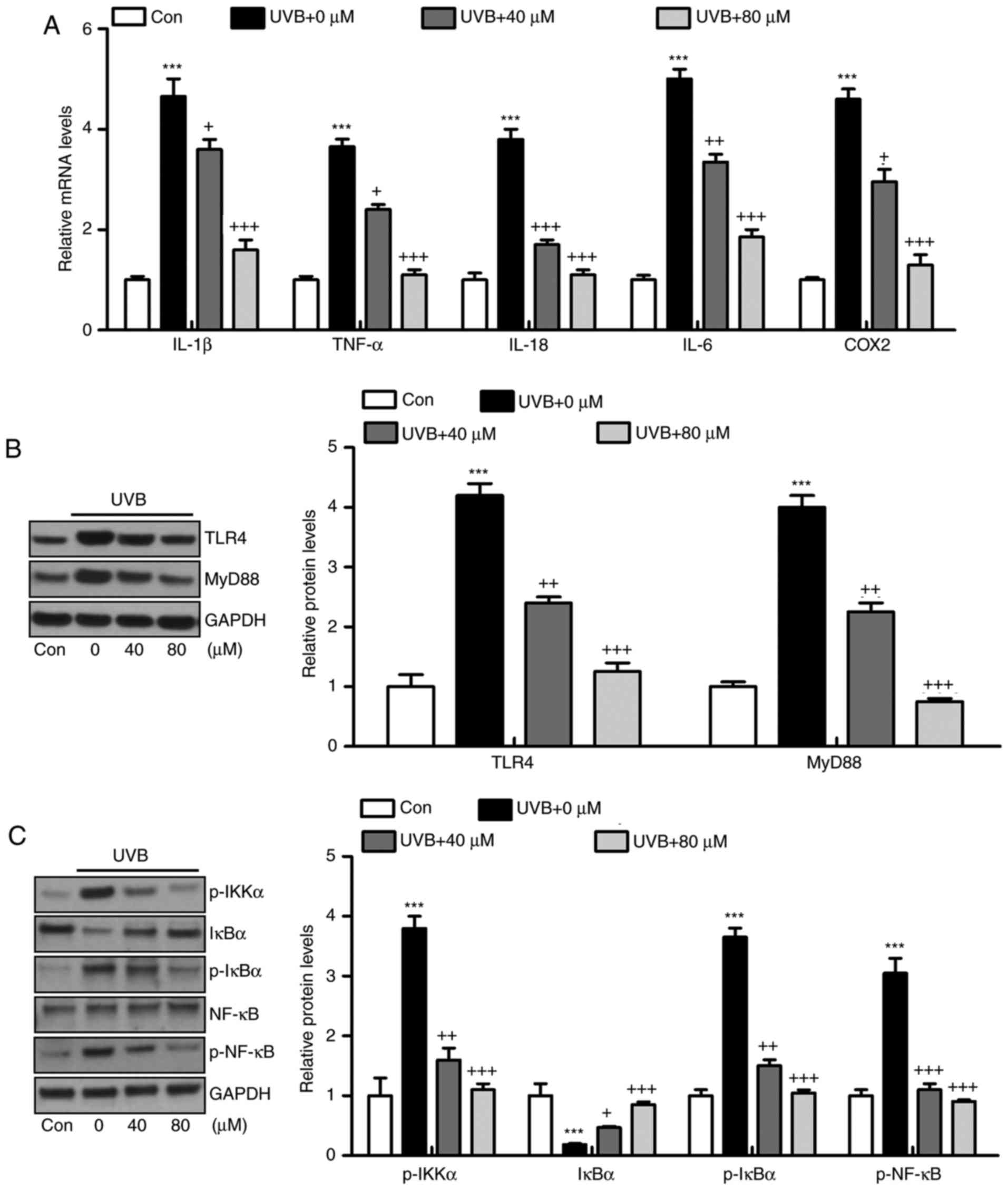 | Figure 9Liquiritin reduces inflammatory
response in UVB-induced HACAT cells in vitro. HACAT cells
were pre-treated with liquiritin at 40 and 80 μM
concentrations for 2 h, followed by UVB irradiation together for
another 22 h. All cells were then harvested and further analysis
was performed to evaluate the role of liquiritin in regulating
UVB-induced cells in vitro. (A) Pro-inflammatory cytokines
IL-1β, TNF-α, IL-18, IL-6 and COX2 were measured using reverse
transcription-quantitative polymerase chain reaction analysis. (B)
Western blot analysis was performed to evaluate protein levels of
TLR4 and MyD88 in vitro. (C) Levels of p-IKKα, IκBα, p-IκBα
and p-NF-κB were assessed using western blot analysis. Data are
presented as the mean ± standard error of the mean (n=8).
***P<0.001, compared with the Con group.
+P<0.05, ++P<0.01 and
+++P<0.001, compared with the UVB-only group. UVB,
ultraviolet B; Con, control; IL, interleukin; TNF-α tumor necrosis
factor-α; COX2, cyclooxygenase 2; TLR4, toll-like receptor 4;
MyD88, myeloid differentiation factor 88; NF-κB, nuclear factor-κB;
IκBα, inhibitor of NF-κB α; IKKα, IκB kinase α; p-,
phosphorylated. |
Liquiritin suppresses oxidative stress
and apoptosis in HACAT cells exposed to UVB in vitro
The oxidative stress confirmed above was also
investigated in vitro. In accordance with the results in
vivo, the mRNA levels of SOD1, SOD2, CAT and Nrf2 induced by
UVB were significantly reduced by liquiritin treatment (Fig. 10A). By contrast, the protein
levels of antioxidants XO, iNOS, NOX-4 and NOX-2 were induced by
UVB exposure, as demonstrated via western blot analysis, and
liquiritin administration significantly reduced the expression of
these molecules (Fig. 10B). In
addition, the protein levels of phosphorylated p38 and JNK induced
by UVB in HACAT cells were upregulated, and liquiritin reduced MAPK
activation (Fig. 11A). Cleaved
caspase-9, caspase-3 and PARP were also expressed at high levels
under UVB irradiation. Consistently, liquiritin exhibited a
suppressive effect in regulating caspase-9, caspase-3 and PARP
cleavage in vitro (Fig.
11B). Finally, it was found that the confluence of HACAT cells
exposed to UVB, determined using crystal violet staining, was
decreased, which was reversed by liquiritin in a dose-dependent
manner (Fig. 11C). These data
indicated that liquiritin reduced UVB-induced oxidative stress and
apoptosis in vitro, which was consistent with the findings
of the in viv experiments.
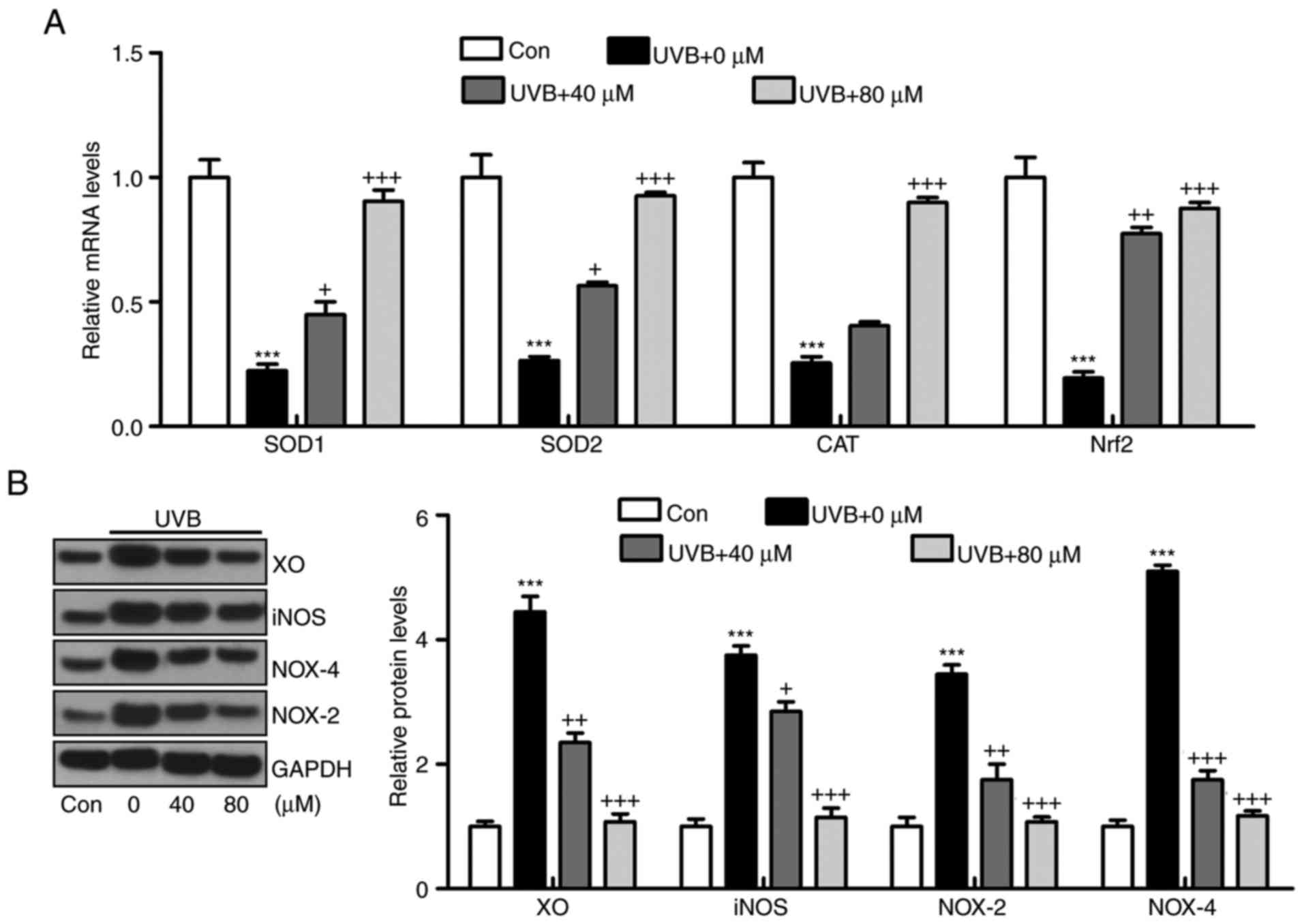 | Figure 10Liquiritin suppresses oxidative
stress via promotion of antioxidants and reduction of oxidants
in vitro. (A) mRNA levels of SOD1, SOD2, CAT and Nrf2 were
measured using reverse transcription-quantitative polymerase chain
reaction analysis. (B) Protein levels of XO, iNOS, NOX-4 and NOX-2
were evaluated using using western blot analysis. Data are
presented as the mean ± standard error of the mean (n=8).
***P<0.001, compared with the Con group.
+P<0.05, ++P<0.01 and
+++P<0.001, compared with the UVB-only group. UVB,
ultraviolet B; Con, control; SOD, superoxide dismutase; CAT,
catalase; Nrf2, nuclear factor erythroid 2-related factor 2; XO,
xanthine oxidase; iNOS, inducible nitric oxide synthase; NOX,
nitric oxide. |
Discussion
Skin cancer is known to be one of the most common
types of cancer in the world. Solar UVB irradiation is a ubiquitous
environmental carcinogen, leading to various cutaneous disorders,
including melanoma and non-melanomatous skin cancer (26). According to previous studies,
plant-derived products have potential antioxidant, anticancer,
antimutagenic and anti-inflammatory properties, which have been
examined for the prevention of skin injury induced by UVB (27). In the present study, liquiritin,
abundant in crude Glycyrrhiza Radix, may be a potential candidate
for use in suppressing UVB-induced hairless mice to examine its
effects on the release of pro-inflammatory cytokines, development
of oxidative stress and progression of apoptosis through various
signaling pathways. The findings of present study indicated that
the administration of liquiritin inhibited the UVB-exposed
inflammatory response, oxidative stress and apoptotic response,
which were dependent on inhibition of the TLR4/NF-κB, ROS-related
and MAPK/caspase signaling pathways.
Accumulating evidence indicates that on exposure to
UVB irradiation, the inflammatory response leads to skin damage by
generating the secretion of pro-inflammatory cytokines, including
IL-1β, TNF-α, IL-18, IL-6 and COX2. The increased expression of
pro-inflammatory cytokines can accelerate inflammatory cell
infiltration, contributing to tissue damage or organ injury,
including airway inflammation of the lung (28,29). In addition, in skin damage induced
under various situations, the hyper-proliferation of keratinocytes
is induced, which is closely associated with the secretion of
pro-inflammatory cytokines (30).
Similarly, in the present study, UVB irradiation significantly
promoted the release of pro-inflammatory cytokines, which were
effectively reduced by liquiritin treatment in the skin tissues of
UVB-induced mice in vivo. In vitro, the UVB-exposed
HACAT cells exhibited higher levels of pro-inflammatory cytokines.
Liquiritin treatment suppressed the expression of these cytokines.
The activation of NF-κB leads to increased synthesis and secretion
of pro-inflammatory cytokines (31). Upon stimulation of TLR-4, MyD88
binds to the cytoplasmatic domain of TLR-4, which activates IKK.
The activated IKK kinase leads to the phosphorylation and
degradation of IκB in the proteasome, and to the release of NF-κB
from the NF-IKB-κB complex, enabling the translocation of NF-κB to
the nucleus, where the expression of genes encoding
pro-inflammatory cytokines is induced (28,31). In the present study, the
TLR4/MyD88 signaling pathway showed a high level of activation
following UVB-irradiation in viv and in vitro
Subsequently, the NF-κB signaling pathway was phosphorylated and
released from the NF-IκB-κB complex, which enhanced
pro-inflammatory cytokine secretion and skin injury.
ROS, including H2O2 and
superoxide anions (O2•−) produced by cells
are involved in the regulation of different cellular functions,
including apoptosis, proliferation, transcription activation and
intracellular signaling (32).
Functioning as a barrier, the skin protects us from being injured
by environmental insults, including UV light and toxic chemicals,
which induces the generation of ROS (33). ROS-induced damage is reported to
be involved in the progression and formation of skin tumors, and in
the pathogenesis of inflammatory skin diseases, including atopic
dermatitis, contact dermatitis and psoriasis (34,35). Aerobic organisms have effective
antioxidant networks to defend against oxidative stress, involving
primary enzymes, including SOD and CAT, and inducible phase II
detoxifying enzymes, including HO-1 and the activation of Nrf-2
(36). For example, SOD has an
antioxidant role in organisms; SOD is necessary as superoxide
reacts with sensitive and important cellular targets, including NO
radicals. Additionally, H2O2 can be
detoxified to H2O by the scavenging enzyme of CAT. These
enzymes act together in the metabolic pathway of free radicals.
HO-1 is regulated by the activation of its transcription factor
Nrf2 to inhibit ROS generation (37). By contrast, MDA is produced by ROS
degrading polyunsaturated lipids and is used as a biomarker to
measure the level of oxidative stress in an organism (34). In the present study,
H2O2, O2•− and MDA were
found to be higher in the UVB-induced mice, and reduced by
liquiritin administration, indicating the effects of liquiritin to
diminish ROS. On the other, RT-qPCR analysis indicated that UVB
irradiation reduced the expression of these enzymes at the gene
level, and these levels were significantly enhanced by liquiritin
administration, reducing ROS generation to ameliorate oxidative
stress. XO is an important enzyme, which catalyzes the oxidation of
hypoxanthine to xanthine and further catalyzes the oxidation of
xanthine to uric acid, producing H2O2 and
superoxide (38). The enzyme is
vital in the development of various diseases (38,39). NO has been implicated as important
mediator in the inflammatory process through accelerating the
production of ROS. The excessive production of NO is produced by
iNOS. NOX-dependent signaling is critical in the development of
tissue injury, including that in the heart, liver and lung
(40). NOX-2 and NOX-4, as the
major NADPH oxidase isoform expressed in the tissue samples, are
principal sources of oxidative stress (41,42). In the present study, it was found
that UVB irradiation significantly upregulated XO, iNOS, NOX2 and
NOX4, enhancing the progression of ROS. Liquiritin exhibited an
inhibitory effect on the expression of these signals. Subsequently,
oxidative stress was improved.
The MAPK signaling pathway is also key in a number
of biological processes, followed by caspase cleavage, contributing
to PARP apoptotic signals (24,25,43). P38 and JNK MAPKs have been
reported to mediate cellular apoptosis, proliferation and
differentiation (44). Apoptosis,
or programmed cell death, is a regulated process, which allows a
cell to self-degrade to enable the body to eliminate unwanted or
dysfunctional cells (45). The
intrinsic pathway is initiated through the release of signaling
factors by mitochondria within the cell, and its two apoptotic
pathways are executed predominantly by a class of cysteine
proteases, termed caspases (46).
It is well known that activation of the p38 MAPK and JNK pathways
leads to the induction of apoptosis through the phosphorylation of
a variety of pro-apoptotic downstream effectors, whereas the ERK1/2
signal pathway is more often associated with cell survival
(47). In the present study, UVB
induction promoted the phosphorylation of p38 and JNK.
Mitochondrial dysfunction induces the activation of caspase-9 and,
subsequently, activates effector caspases, including caspase-3.
Following caspase-3 activation, the cleavage of PARP occurs,
contributing to apoptosis (48).
In accordance with the description above, the present study showed
that UVB exposure elevated the cleavage of caspase-9, caspase-3 and
PARP, leading to apoptosis, evidenced by TUNEL analysis. Liquiritin
was found to suppress the activation of p38 and JNK. Consequently,
caspase cleavage and PARP cleavage were reduced. Eventually,
UVB-induced cell death may be prevented.
In conclusion, the findings of the present study
indicated that liquiritin application to mice exposed to UVB leads
to a significant reduction in the release of pro-inflammatory
cytokines through the inactivation of TLR4/MyD88/NF-κB.
Additionally, oxidative stress was suppressed by liquiritin through
promoting the expression of antioxidants and inhibiting levels of
oxidants. Finally, cell survival was enhanced due to liquiritin
treatment via the suppression of apoptosis. Overall, the findings
suggested that liquiritin may be developed as an effective
photochemopreventive candidate to prevent UVB-induced skin
damage.
Acknowledgments
The authors thank Dr. Ruixia Yang for her technical
support.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
XQL, LMC and MJ conceived and designed the study.
XQL, LMC, JL, YLM, YHK and HL performed the experiments. XQL and
LMC wrote the study. XQL, LMC, YHK and MJ reviewed and edited the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All animal experiments were performed following the
Guide for the Care and Use of Laboratory Animals, issued by the
National Institutes of Health in 1996 and approved by Huai'an First
People's Hospital, Nanjing Medical University (Nanjing, China). The
care and handling of mice were in accordance with the ethical
guidelines of Huai'an First People's Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that there are no competing
interests.
References
|
1
|
Yager J: The skin as an immune organ
Advances in Veterinary Dermatology. Ihrke PJ, Mason IS and White
SD: Pergamon Press; Oxford: pp. 311993
|
|
2
|
Afaq F, Adhami VM and Mukhtar H:
Photochemoprevention of ultraviolet B signaling and
photocarcinogenesis. Mutat Res. 571:153–173. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Muthusamy V and Piva TJ: A comparative
study of UV-induced cell signalling pathways in human
keratinocyte-derived cell lines. Arch Dermatol Res. 305:817–833.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Muthusamy V and Piva TJ: The UV response
of the skin: A review of the MAPK, NFkappaB, and TNFalpha signal
transduction pathways. Arch Dermatol Res. 302:5–17. 2010.
View Article : Google Scholar
|
|
5
|
Masferrer JL, Leahy KM, Koki AT, Zweifel
BS, Settle SL, Woerner BM, Edwards DA, Flickinger AG, Moore RJ and
Seibert K: Antiangiogenic and antitumor activities of
cyclooxygenase-2 inhibitors. Cancer Res. 60:1306–1311.
2000.PubMed/NCBI
|
|
6
|
Kim SB, Kang OH, Joung DK, Mun SH, Seo YS,
Cha MR, Ryu SY, Shin DW and Kwon DY: Anti-inflammatory effects of
tectroside on UVB-induced HaCaT cells. Int J Mol Med. 31:1471–1476.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Buckman SY, Gresham A, Hale P, Hruza G,
Anast J, Masferrer J and Pentland AP: COX-2 expression is induced
by UVB exposure in human skin: Implications for the development of
skin cancer. Carcinogenesis. 19:723–729. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Karpurapu M, Wang X, Deng J, Park H, Xiao
L, Sadikot RT, Frey RS, Maus UA, Park GY, Scott EW and Christman
JW: Functional PU.1 in macrophages has a pivotal role in NF-κB
activation and neutrophilic lung inflammation during endotoxemia.
Blood. 118:5255–5266. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Medvedev AE, Lentschat A, Wahl LM,
Golenbock DT and Vogel SN: Dysregulation of LPS-induced Toll-like
receptor 4-MyD88 complex formation and IL-1 receptor-associated
kinase 1 activation in endotoxin-tolerant cells. J Immunol.
169:5209–5216. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee J, Giordano S and Zhang J: Autophagy,
mitochondria and oxidative stress: Cross-talk and redox signalling.
Biochem J. 441:523–540. 2012. View Article : Google Scholar :
|
|
11
|
Acker T, Fandrey J and Acker H: The good,
the bad and the ugly in oxygen-sensing: ROS, cytochromes and
prolyl-hydroxylases. Cardiovasc Res. 71:195–207. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Burotto M, Chiou VL, Lee JM and Kohn EC:
The MAPK pathway across different malignancies: A new perspective.
Cancer. 120:3446–3456. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hsieh SC, Huang MH, Cheng CW, Hung JH,
Yang SF and Hsieh YH: α-Mangostin induces mitochondrial dependent
apoptosis in human hepatoma SK-Hep-1 cells through inhibition of
p38 MAPK pathway. Apoptosis. 18:1548–1560. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lamy E, Herz C, Lutz-Bonengel S, Hertrampf
A, Márton MR and Mersch-Sundermann V: The MAPK pathway signals
telomerase modulation in response to isothiocyanate-induced DNA
damage of human liver cancer cells. PLoS One. 8:e532402013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hui K, Yang Y, Shi K, Luo H, Duan J, An J,
Wu P, Ci Y, Shi L and Xu C: The p38 MAPK-regulated PKD1/CREB/Bcl-2
pathway contributes to selenite-induced colorectal cancer cell
apoptosis in vitro and in vivo. Cancer Lett. 354:189–199. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang W, Hu X, Zhao Z, Liu P, Hu Y, Zhou J,
Zhou D, Wang Z, Guo D and Guo H: Antidepressant-like effects of
liquiritin and isoliquiritin from Glycyrrhiza uralensis in the
forced swimming test and tail suspension test in mice. Prog
Neuropsychopharmacol Biol Psychiatry. 32:1179–1184. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun YX, Tang Y, Wu AL, Liu T, Dai XL,
Zheng QS and Wang ZB: Neuroprotective effect of liquiritin against
focal cerebral ischemia/reperfusion in mice via its antioxidant and
antiapoptosis properties. J Asian Nat Prod Res. 12:1051–1060. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu TY, Khor TO, Saw CL, Loh SC, Chen AI,
Lim SS, Park JH, Cai L and Kong AN:
Anti-inflammatory/anti-oxidative stress activities and differential
regulation of Nrf2-mediated genes by non-polar fractions of tea
Chrysanthemum zawadski and licoric Glycyrrhiza uralensis. AAPS J.
13:1–13. 2011. View Article : Google Scholar
|
|
19
|
Dong SJ, Inoue A, Zhu Y, Tanji M and
Kiyama R: Activation of rapid signally pathways and the subsequent
transcriptional regulation for the proliferation of breast cancer
MCF-7 cells by the treatment with an extract of Glycyrrhiza glabra
root. Food Chem Toxicol. 45:2470–2478. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Weiler C, Nerlich AG, Bachmeier BE and
Boos N: Expression and distribution of tumor necrosis factor alpha
in human lumbar intervertebral discs: A study in surgical specimen
and autopsy controls. Spine (Phila Pa 1976). 30:44–54. 2005.
View Article : Google Scholar
|
|
22
|
Le Maitre CL, Hoyland JA and Freemont AJ:
Catabolic cytokine expression in degenerate and herniated human
intervertebral discs: IL-1beta and TNFalpha expression profile.
Arthritis Res Ther. 9:R772007. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fu Y, Hsieh ZC, Guo JQ, Kunicki J, Lee
YWT, Darzynkiewicz Z and Wu JM: Licochalcone-A, a novel flavonoid
isolated from licorice root (Glycyrrhiza glabra), causes G2 and
late-G1 arrests in androgen-independent PC-3 prostat cancer cells.
Biochem Biophys Res Commun. 322:263–270. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kuo WH, Chen JH, Lin HH, Chen BC, Hsu JD
and Wang CJ: Induction of apoptosis in the lung tissue from rats
exposed to cigarette smoke involves p38/JNK MAPK pathway. Chem Biol
Interact. 155:31–42. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
American Cancer Society: Cancer Facts and
Figures, 2010. American Cancer Society; Atlanta, GA: 2010
|
|
26
|
Suzuki Y, Nakabayashi Y, Nakata K, Reed JC
and Takahashi R: X-linked inhibitor of apoptosis protein (XIAP)
inhibits caspase-3 and -7 in distinct modes. J Biol Chem.
276:27058–27063. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Perrone D, Ardito F, Giannatempo G,
Dioguardi M, Troiano G, Lo Russo L, DE Lillo A, Laino L and Lo
Muzio L: Biological and therapeutic activities, and anticancer
properties of curcumin (Review). Exp Ther Med. 10:1615–1623. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nilsson MB, Langley RR and Fidler IJ:
Interleukin-6, secreted by human ovarian carcinoma cells, is a
potent proangiogenic cytokine. Cancer Res. 65:10794–10800. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cho JW, Lee KS and Kim CW: Curcumin
attenuates the expression of IL-1beta, IL-6, and TNF-alpha as well
as cyclin E in TNF-alpha-treated HaCaT cells; NF-kappaB and MAPKs
as potential upstream targets. Int J Mol Med. 19:469–474.
2007.PubMed/NCBI
|
|
30
|
Chen W, Tang Q, Gonzales MS and Bowden GT:
Role of p38 MAP kinases and ERK in mediating ultraviolet-B induced
cyclooxygenase-2 gene expression in human keratinocytes. Oncogene.
20:3921–3926. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wullaert A, Bonnet MC and Pasparakis M:
NF-κB in the regulation of epithelial homeostasis and inflammation.
Cell Res. 21:146–158. 2011. View Article : Google Scholar
|
|
32
|
Rajagopalan S, Kurz S, Münzel T, Tarpey M,
Freeman BA, Griendling KK and Harrison DG: Angiotensin II-mediated
hypertension in the rat increases vascular superoxide production
via membrane NADH/NADPH oxidase activation. Contribution to
alterations of vasomotor tone. J Clin Invest. 97:1916–1923. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Touyz RM and Briones AM: Reactive oxygen
species and vascular biology: Implications in human hypertension.
Hypertens Res. 34:5–14. 2011. View Article : Google Scholar
|
|
34
|
Jia SJ, Jiang DJ, Hu CP, Zhang XH, Deng HW
and Li YJ: Lysophosphatidylcholine-induced elevation of asymmetric
dimethylarginine level by the NADPH oxidase pathway in endothelial
cells. Vascul Pharmacol. 44:143–148. 2006. View Article : Google Scholar
|
|
35
|
Hanna IR, Taniyama Y, Szöcs K, Rocic P and
Griendling KK: NAD(P)H oxidase-derived reactive oxygen species as
mediators of angiotensin II signaling. Antioxid Redox Signal.
4:899–914. 2002. View Article : Google Scholar
|
|
36
|
Luo Z, Teerlink T, Griendling K, Aslam S,
Welch WJ and Wilcox CS: Angiotensin II and NADPH oxidase increase
ADMA in vascular smooth muscle cells. Hypertension. 56:498–504.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Was H, Dulak J and Jozkowicz A: Heme
oxygenase-1 in tumor biology and therapy. Curr Drug Targets.
11:1551–1570. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang G, Qian P, Jackson FR, Qian G and Wu
G: Sequential activation of JAKs, STATs and xanthine
dehydrogenase/oxidase by hypoxia in lung microvascular endothelial
cells. Int J Biochem Cell Biol. 40:461–470. 2008. View Article : Google Scholar
|
|
39
|
Ladilov Y, Schäfer C, Held A, Schäfer M,
Noll T and Piper HM: Mechanism of Ca(2+) overload in endothelial
cells exposed to simulated ischemia. Cardiovasc Res. 47:394–403.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Makino J, Kamiya T, Hara H and Adachi T:
TPA induces the expression of EC-SOD in human monocytic THP-1
cells: Involvement of PKC, MEK/ERK and NOX-derived ROS. Free Radic
Res. 46:637–644. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Carbone F, Teixeira PC, Braunersreuther V,
Mach F, Vuilleumier N and Montecucco F: Pathophysiology and
treatments of oxidative injury in ischemic stroke: Focus on the
phagocytic NADPH oxidase 2. Antioxid Redox Signal. 23:460–489.
2015. View Article : Google Scholar :
|
|
42
|
Ansari MA and Scheff SW: NADPH-oxidase
activation and cognition in Alzheimer disease progression. Free
Radic Biol Med. 51:171–178. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pearson G, Robinson F, Beers Gibson T, Xu
BE, Karandikar M, Berman K and Cobb MH: Mitogen-activated protein
(MAP) kinase pathways: Regulation and physiological functions.
Endocr Rev. 22:153–183. 2001.PubMed/NCBI
|
|
44
|
Yuan L, Wang J, Xiao H, Wu W, Wang Y and
Liu X: MAPK signaling pathways regulate mitochondrial-mediated
apoptosis induced by isoorientin in human hepatoblastoma cancer
cells. Food Chem Toxicol. 53:62–68. 2013. View Article : Google Scholar
|
|
45
|
Lin C, Holland RE Jr, Donofrio JC, McCoy
MH, Tudor LR and Chambers TM: Caspase activation in equine
influenza virus induced apoptotic cell death. Vet Microbiol.
84:357–365. 2002. View Article : Google Scholar
|
|
46
|
Vermes I, Haanen C and Reutelingsperger C:
Flow cytometry of apoptotic cell death. J Immunol Methods.
243:167–190. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Muslin AJ: MAPK signalling in
cardiovascular health and disease: Molecular mechanisms and
therapeutic targets. Clin Sci (Lond). 115:203–218. 2008. View Article : Google Scholar
|
|
48
|
Newhouse K, Hsuan SL, Chang SH, Cai B,
Wang Y and Xia Z: Rotenone-induced apoptosis is mediated by p38 and
JNK MAP kinases in human dopaminergic SH-SY5Y cells. Toxicol Sci.
79:137–146. 2004. View Article : Google Scholar : PubMed/NCBI
|