Introduction
Multiple myeloma (MM) is characterized by the
abnormal accumulation of clonal malignant plasma cells; it accounts
for ~1% of neoplastic diseases and 13% of hematological
malignancies. MM is defined by the excessive production of heavy
and light chain monoclonal immunoglobulins, and bone marrow
plasmacytosis. It is associated with bone lesions, anemia,
cytopenia, hypercalcemia, peripheral neuropathy and renal
dysfunction (1). The treatment
for this malignancy has advanced significantly in recent decades;
however, the prognosis for patients with MM is poor despite the
available chemotherapeutic agents and emerging targeted treatments
(2). The relapse of MM occurs
frequently, and thus, novel therapeutic modalities are
required.
Pterostilbene
(trans-3,5-dimethoxy-4′-hydroxystilbene; PTE) (3), predominately occurring in
blueberries and certain varieties of grapes, is a naturally
occurring dimethyl ether-analogue of resveratrol. However, PTE is
more metabolically stable and has exhibited more favorable
pharmacological activities than resveratrol as its dimethyl ether
structure enhances lipophilicity and membrane permeability
(4). PTE has gained significant
attention for its potent antioxidant, anti-inflammatory and
anticarcinogenic properties (5,6).
However, the mechanism of the inhibitory effect of PTE on tumor
cells has not yet been completely characterized. Numerous previous
research efforts have focused on the ability of PTE to target
metabolic pathways that may be altered during insulin resistance or
metabolic syndrome, as well as in the abnormal metabolism
associated with cancer cells (7-9).
By contrast to normal human tissue, cancer cells,
including MM cells, depend on a high anabolic metabolism rate.
Cancer cell survival requires a high availability of de novo
lipogenesis, which is achieved by overexpressing key lipogenic
enzymes, including acetyl-CoA carboxylase (ACC) and fatty acid
synthase (FASN) (10,11). The expression of these enzymes is
positively correlated with increasing cancer stage, aggressiveness
and drug resistance (12,13). Pharmacological inhibitors that
block FASN or ACC1 activity can inhibit the survival of a range of
types of cancer cells (14-16). MM cells undergo extensive protein
synthesis, specifically that of immunoglobulin. Therefore, MM cells
are particularly reliant on protein metabolism homeostasis
(17,18). Two opposing pathways control
protein synthesis. The absence of amino acids induces the
phosphorylation of α-subunit of eukaryotic translation initiation
factor-2 (eIF2α), which interferes with eIF2 and consequently
hinders the initiation of translation. However, in the presence of
amino acids, mechanistic target of rapamycin (mTOR) is activated
and phosphorylates eIF4E-binding protein 1 (4E-BP1), which
facilitates eIF4 assembly, and thus protein synthesis (19-21).
One critical monitor that may regulate cellular and
organismal metabolic homeostasis is AMP-activated protein kinase
(AMPK), which coordinates cell survival and apoptosis in response
to nutrient and energy levels (22). AMPK is activated in response to
cellular stress or pharmacological inducers that inhibit anabolic
pathways. AMPK acts to decrease ATP consumption and promote
catabolic pathways that generate more ATP (23). Therefore, AMPK acts as a central
metabolic switch that governs metabolism. As a number of previous
studies have reported the important role of AMPK in the
pro-apoptotic pathway of cancer cells, AMPK is an attractive target
molecule for cancer treatment (24,25).
PTE is reported to mediate AMPK activation in
numerous cell types, including hepatocytes and vascular endothelial
cells (26,27). However, the effect of PTE on the
AMPK pathway in tumor cells has only been reported in prostate
cancer cells (9). The effect of
PTE on cancer metabolic regulation is also unclear. In the present
study, the potential of PTE as a non-toxic anti-neoplastic strategy
for patients with MM was investigated from a metabolic perspective
in MM cells. It was demonstrated that PTE effectively induced MM
cell apoptosis by blocking energy metabolism through the activation
of AMPK. The present study suggested that targeting AMPK activation
with PTE represents a relevant strategy for MM prevention and
therapy.
Materials and methods
Reagents
PTE, 3-methyladenine (3-MA), monodansylcadaverine
(MDC) and dimethyl sulfoxide (DMSO) were purchased from
Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). Compound C was
purchased from Selleck Chemicals (Houston, TX, USA).
PTE stock solution preparation
PTE was dissolved in DMSO to yield a 78 mM stock
solution that was stored at −20°C. The different doses of PTE (10,
20, 30, 40, 50, 60 and 70 µM) for each treatment group were
diluted with RPMI-1640 medium (Sigma-Aldrich; Merck KGaA). All
experiments used a corresponding volume of DMSO as a control
treatment.
Cell culture
RPMI-8226, NCI-H929, U266 and ARH-77 human MM cell
lines were purchased from American Type Culture Collection
(Manassas, VA, USA). The RPMI-8226, NCI-H929, U266 and ARH-77 cells
were cultured in RPMI-1640 medium supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and 100 U/ml penicillin/streptomycin (Beyotime Institute of
Biotechnology, Haimen, China). The cells were incubated in a
humidified incubator with 5% CO2 at 37°C.
Proliferation and viability assays
Cell viability was determined using a Cell Counting
kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan) assay. In
brief, 1×104 cells/well were seeded into 96-well plates
in a final volume of 100 µl of complete culture medium with
the specified concentrations of PTE. The CCK-8 reagent was added
and incubated for an additional 0.5-4 h at 37°C and the optical
density (OD) was measured at 450 nm using a microplate reader.
Western blotting
Following PTE treatment with or without 1 µM
compound C for 48 h, the MM cells were harvested and lysed in
radioimmunoprecipitation assay lysis solution (Beyotime Institute
of Biotechnology) containing phosphatase inhibitors and PMSF. A
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology) was used to determine the protein concentrations.
Protein lysates (30-50 µg per sample) were separated with
6-12% SDS-polyacrylamide gels (Beyotime Institute of Biotechnology)
and transferred to polyvinyl difluoride membranes (EMD Millipore,
Billerica, MA, USA). The membranes were blocked in 5% skimmed milk
in Tris-buffered saline for 1 h at room temperature and incubated
at 4°C overnight with primary antibodies against cleaved caspase 9
(Asp330; cat. no. 9501T), caspase 9 (cat. no. 9502T),
poly(ADP-ribose) polymerase (PARP; cat. no. 9532), phosphorylated
(p)-AMPK (Thr172; cat. no. 2535T), AMPK (cat. no. 5832T), FASN
(cat. no. 3180T), ACC (cat. no. 3676T), p-ACC (Ser79; cat. no.
11818T), p-mTOR (Ser2448; cat. no. 5536), mTOR (cat. no. 2983),
4E-BP1 (cat. no. 9644T), p4E-BP1 (Thr37/46) (cat. no. 2855T),
p-eIF2α (Ser51; cat. no. 3398T), eIF2α (cat. no. 5324T),
autophagy-related (ATG)5 (cat. no. 12994T), beclin1 (cat. no.
3495T) and light chain (LC)3B (cat. no. 3868T; all dilution,
1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA), and
primary antibody against cleaved caspase 3 (cat. no. ab32042;
dilution, 1:500; Abcam, Cambridge, MA, USA). The primary antibody
against β-actin was from Antgene (cat. no. ANT010S; dilution,
1:2,000; Wuhan, China). The membranes were incubated with
horseradish peroxidase-conjugated anti-rabbit (cat. no. ANT020;
dilution, 1:5,000; Antgene) or anti-mouse (cat. no. ANT019;
dilution, 1:5,000; Antgene) secondary antibodies for 60 min at room
temperature. The blots were developed using an enhanced
chemiluminescence reagent (Antgene). The protein expression levels
were normalized to the level of β-actin in the corresponding lanes
and Image Lab 3.0 software (Bio-Rad Laboratories, Inc., Hercules,
CA, USA) was used for densitometric analysis.
Flow cytometry analysis
Following PTE treatment, or PTE co-treatment with 1
µM compound C or 3 mM 3MA, cell apoptosis was assayed with
Annexin V/propidium iodide staining (PI; BD Biosciences, San Jose,
CA, USA), followed by fluorescence-activated cell sorting analysis.
RPMI-8226 cells were treated with 0, 20, 40 or 60 µM PTE and
ARH-77 cells were treated with 0, 10, 20 or 40 µM PTE for 24
h. Next, 1×106 cells were resuspended in 100 µl
Annexin V-binding buffer containing 5 µl Annexin V and PI.
Following incubation at room temperature for 15 min, 400 µl
binding buffer was added to the cells, which were analyzed using a
FACSAriaII flow cytometer (BD Biosciences). The rate of apoptotic
events was defined as the sum of the number of cells in the early
(Annexin V+/PI-) and late (Annexin
V+/PI+) stages of apoptosis. FlowJo 7.6
software (FlowJo LLC, Ashland, OR, USA) was used to analyze the
flow cytometry data.
Visualization of autophagic vacuoles with
MDC
MDC was used as an autophagolysosome-specific marker
to analyze the autophagic process. Cells were seeded into 6-well
plates and treated with 40 µM PTE for 24 h. Autophagic
vacuoles were labeled with MDC by incubating cells with 50
µM MDC at 37°C for 30 min. The cells were washed with PBS
three times in 5 min intervals. The cells were then immediately
analyzed using a fluorescence microscope (magnification, ×200;
Nikon Corporation, Tokyo, Japan).
Transmission electron microscopy
(TEM)
The autophagosome ultrastructure of MM cells was
observed by TEM. RPMI-8226 cells were exposed to 40 µM PTE
for 0, 7 and 24 h. The cells were fixed with 2.5% glutaraldehyde at
4°C for 2 h, followed by incubation with 2%
OsO4 for 2-3 h at 4°C. Following dehydration
in a graded ethanol series, cells were infiltrated and embedded in
EMbed-812 (Electron Microscopy Sciences, Hatfield, PA, USA) at 60°C
for 48 h. Thin (60-100 nm) sections were stained at room
temperature with uranyl acetate for 15 min and lead citrate for 10
min, prior to being examined using a Tecnai G2 20 TWIN electron
microscope (magnification, ×1,700 and ×5,000; FEI, Eindhoven, the
Netherlands).
Animal model and treatment
A total of 10 female NOD/SCID mice (weight, 15.7±1.0
g; age, 3-4 weeks; Nanjing Bioscience Company, Nanjing, China) were
individually maintained in a temperature-controlled (23±2°C with
50-60% relative humidity) room with a 12-h light/dark cycle and
ad libitum access to water and food. All experimental
procedures and protocols were approved by the Committee on Animal
Handling of Huazhong University of Science and Technology (Wuhan,
China). The mice were subcutaneously injected with 2×107
RPMI-8226 cells in 200 µl serum-free RPMI-1640 medium. The
treatment began when the tumor volume reached ~100 mm3
(~3 weeks). Tumor volumes were calculated using the following
formula: V = A×B2/2, where A is the largest diameter and
B the smallest. 10 mice were randomly divided into the control (5%
DMSO) or PTE (50 mg/kg in 5% DMSO) groups (n=5). Intraperitoneal
injections were performed 5 days a week for 21 days. Tumor sizes
were evaluated daily. All mice were sacrificed on day 21 and their
tumors were excised. Tumor tissues were fixed in 10% formalin at
room temperature for 24 h for further analysis. Blood was also
collected at sacrifice and serum samples were stored at −80°C until
further analysis.
Terminal deoxynucleotidyl transferase
mediated dUTP nick-end labeling (TUNEL) staining
A TUNEL assay kit (cat. no. 11684817910; Roche
Diagnostics, Ltd., Basel, Switzerland) was used to detect apoptosis
in the tumor tissues, according to the manufacturer's protocol.
Tumor tissues from mice were fixed in 10% formalin at room
temperature for 24 h. Paraffin-embedded sections (4-µm) from
tumor tissues were deparaffinized and hydrated by sequential
immersion in xylene and a graded alcohol series (100, 100, 95, 90,
80 and 70%). Next, the sections were incubated with the TUNEL
reaction mixture (TdT+dUTP, v/v=1:9) at 37°C for 1 h, and
hematoxylin was used to stain the nuclei at room temperature for 5
min. The sections were sealed with neutral gum and visualized using
a fluorescence microscope (magnification, ×100; Nikon Corporation).
Five fields of view for each section were observed.
Histology and immunohistochemistry
(IHC)
Tumors from mice were fixed in 10% formalin at room
temperature for 24 h. Paraffin-embedded sections (4-µm) of
tumor tissues were deparaffinized and hydrated by sequential
immersion in xylene and a graded alcohol series (100, 100, 95, 90,
80 and 70%). Following antigen retrieval with 10 mM citrate buffer
(pH=6.0) at 95°C for 10 min, endogenous tissue peroxidase activity
was blocked with 3% hydrogen peroxide at room temperature for 10
min. Next, non-specific sites were blocked with 5% bovine serum
albumin (Roche Diagnostics) at room temperature for 20 min. The
sections were incubated with antibodies against p-AMPK (Thr172;
cat. no. 2535S; dilution, 1:100; Cell Signaling Technology, Inc.),
p-mTOR (cat. no. bs-3494R; dilution, 1:200; Biosynthesis
Biotechnology Co., Ltd., Beijing, China), FASN (cat. no. 3180S;
dilution, 1:50; Cell Signaling Technology, Inc.) and cleaved
caspase-3 (cat. no. RLC006; dilution, 1:50; Ruiying Biological,
Suzhou, China) at 4°C overnight. Next, the sections were incubated
with the following secondary antibodies: horseradish
peroxidase-conjugated anti-rabbit or anti-mouse (cat. no. K5007;
dilution, 1:200; Dako; Agilent Technologies, Santa Clara, CA, USA)
for 50 min at room temperature. The labelled cells were observed
using a fluorescence microscope (Nikon Corporation) for cleaved
caspase-3 (magnification, ×100) and for p-AMPK, FASN and p-mTOR
(magnification, ×200). Five fields of view for each section were
observed.
In vivo toxicity assay
Inner canthus blood samples (100 µl) were
collected from the mice prior to sacrifice. Blood cells were
isolated by density gradient centrifugation at 3,000 x g at room
temperature for 10 min, and creatine kinase-MB (CK-MB), cardiac
troponins T (cTnT), alanine aminotrans-ferase (ALT), aspartate
amino transferase (AST), alkaline phosphatase (ALP), total
bilirubin (TBIL), urea nitrogen (BUN) and serum creatinine (Scr)
were detected in the serum. Concentrations of CK-MB and cTnT were
determined quantitatively using the commercially available CK-MB
ELISA kit (cat. no. FLSW-M1025; Kehua Bio-Engineering co., Ltd.,
Shanghai, China) and cTnT ELISA kit (cat. no. FLSW-M763; Kehua
Bio-Engineering co., Ltd.) according to the manufacturer's
protocol. The absorbance at 450 nm was measured using an automated
microplate reader. The other serum indexes, including ALT, AST,
ALP, TBIL, BUN and Scr, were measured by routine methods using a
fully automatic biochemical analyzer (Automatic Analyzer AU2700;
Beckman Coulter, Inc., Brea, CA, USA), according to the
manufacturer's protocol.
Statistical analyses
SPSS 15.0 software (SPSS Inc, Chicago, IL, USA) was
used to analyze all recorded data. Data are presented as the mean ±
standard deviation of three independent experiments. Statistically
significant differences between multiple group comparisons were
determined using one-way analysis of variance followed by Dunnett's
test. Comparisons between two groups were made using unpaired
Student's t-tests. P<0.05 was considered to indicate a
statistically significant difference.
Results
PTE reduces MM cell viability and induces
apoptosis
PTE treatment has demonstrated antitumor activity in
several types of human cancer cells (28,29). Therefore, the growth inhibition
effect of PTE on MM cells was assessed. Different concentrations of
PTE between 0 and 70 µM were applied to the MM RPMI-8226,
ARH-77, U266 and NCI-H929 cell lines for 48 h. Consistent with
other previous studies (30,31), a significant reduction in cell
viability was observed following the PTE treatment of MM cells. PTE
induced the concentration-dependent decrease in viability of
RPMI-8226, ARH77, U266 and NCI-H929 cells, with IC50
values at 48 h of 33.8, 16.6, 63.5 and 44.5 µM, respectively
(Fig. 1A). The relatively
sensitive RPMI-8226 and ARH-77 cells were used for subsequent
experiments.
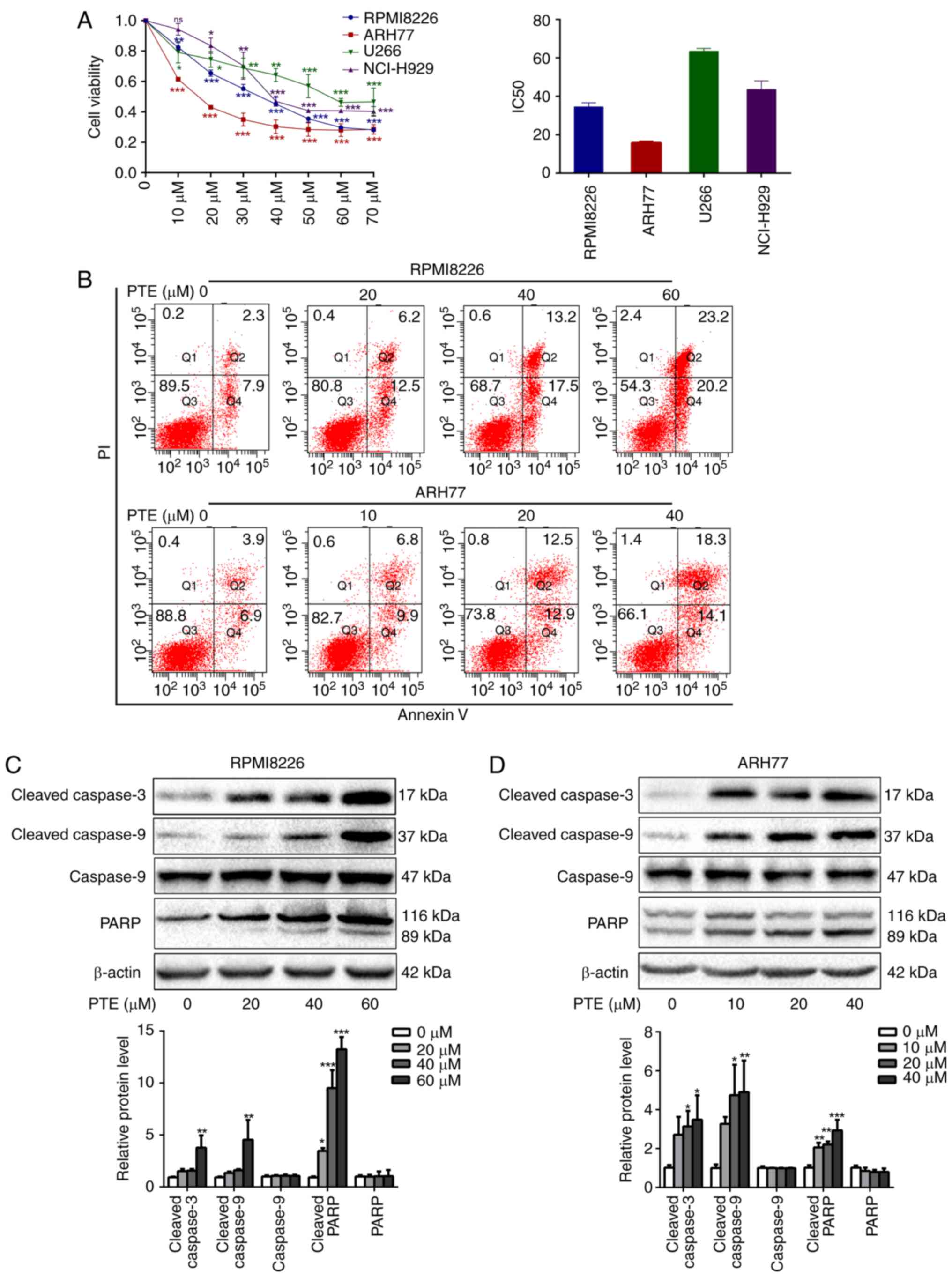 | Figure 1PTE reduces MM cell viability and
induces cell apoptosis. (A) RPMI-8226, ARH-77, U266 and NCI-H929
cells were treated with PTE and their viability was measured using
a Cell Counting kit-8 assay. The IC50 value is shown.
(B) Flow cytometric analysis of the effect of PTE on apoptosis
using an Annexin V-FITC/PI kit. DMSO served as the control. (C)
RPMI-8226 and (D) ARH-77 cells were treated with DMSO (control) or
different concentrations of PTE for 48 h, the expression levels of
cleaved caspase 3, cleaved caspase 9 and cleaved PARP were
determined using western blotting. β-actin served as a loading
control. *P<0.05, **P<0.01,
***P<0.001 vs. DMSO group. PTE, pterostilbene; MM,
multiple myeloma; FITC, fluorescein isothiocyanate; PI, propidium
iodide; DMSO, dimethyl sulfoxide; PARP, poly(ADP-ribose)
polymerase. |
To investigate the mechanisms of the growth
inhibitory effect of PTE on MM cells, the effects of PTE on the
rate of apoptosis were analyzed by flow cytometry. PTE treatment
for 24 h induced early and late apoptosis to a greater extent
compared with the DMSO-treated controls in RPMI-8226 and ARH-77
cells in a dose-dependent manner (Fig. 1B). Compared with the control group
(2.3+7.9%), apoptosis was increased at 24 h in the 20 µM
(6.2+12.5%), 40 µM (13.2+17.5%) and 60 µM
(23.2+20.2%) PTE-treated groups in RPMI-8226 cells. Similar
findings were observed in ARH77 cells, in which apoptosis was
increased at 24 h in the 10 µM (6.8+9.9%), 20 µM
(12.5+12.9%) and 40 µM (18.3+14.1%) PTE-treated groups,
compared with the control group (3.9+6.9%). Taken together, these
data indicated that PTE treatment increased the rate of apoptosis
of MM cells.
These results coincided with a marked increase in
the expression of the pro-apoptotic proteins, cleaved PARP and
cleaved caspase 3/9, following PTE exposure for 48 h in RPMI-8226
and ARH-77 cells (Fig. 1C and D).
Taken together, these results suggested that PTE treatment
attenuates MM cell viability and induces apoptosis.
PTE treatment activates the AMPK pathway
in MM cells
As the survival of MM cells depends on high rates of
anabolic metabolism, and AMPK is a critical energy sensor in
cellular metabolism (23,32), it was assessed whether PTE
treatment activated the AMPK pathway in MM cells. RPMI-8226 and
ARH-77 cells were exposed to different concentrations of PTE for 48
h, and the western blot data from these cells demonstrated that PTE
induced the phosphorylation of AMPK in a dose-dependent manner
(Fig. 2A).
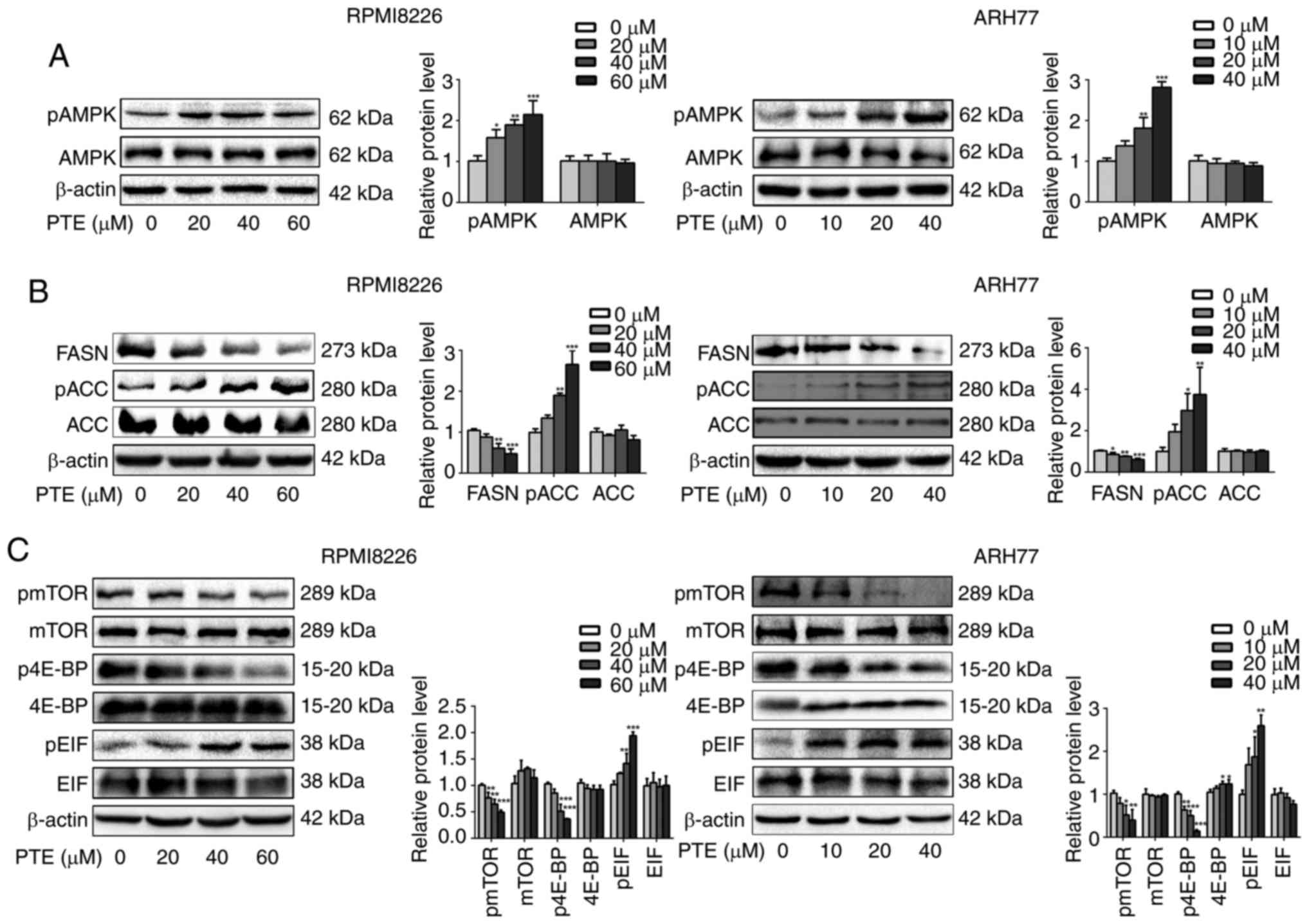 | Figure 2PTE suppresses the expression of
metabolic proteins in MM cells. RPMI-8226 (left) and ARH-77 (right)
cells were treated with DMSO (control) or different concentrations
of PTE for 48 h. Western blotting was used to analyze the protein
expression levels of (A) p-AMPK (Thr 172); (B) FASN, p-ACC (Ser 79)
and ACC; (C) p-mTOR (Ser 2448), mTOR, p-4E-BP1 (Thr 37/46), 4E-BP1,
eIF2α and p-eIF2α. β-actin served as a loading control. The
presented western blotting data are representative of those
obtained in at least three separate experiments.
*P<0.05, **P<0.01,
***P<0.001 vs. the DMSO group. PTE, pterostilbene;
MM, multiple myeloma; DMSO, dimethyl sulfoxide; p-, phosphorylated;
AMPK, AMP-activated protein kinase; FASN, fatty acid synthase; ACC,
acetyl-CoA carboxylase; mTOR, mechanistic target of rapamycin;
4E-BP1, 4E-binding protein 1; eIF2α, eukaryotic translation
initiation factor-2α. |
The activity of the de novo fatty acid
synthesis key enzymes FASN and ACC is negatively regulated by AMPK
(33). Therefore, it was then
investigated whether PTE decreased lipid synthesis by decreasing
FASN expression or inhibiting ACC activity. It was observed that
the FASN protein expression level was decreased, and ACC was
phosphorylated in a dose-dependent manner when RPMI-8226 and ARH-77
cells were treated with PTE (Fig.
2B). The inhibition of lipogenic key enzymes may induce MM
cells into a low de novo lipogenesis state.
As the survival of MM cells is dependent on
extensive protein synthesis and the activation of AMPK inhibits the
mTOR protein synthesis pathway (34,35), it was next examined whether the
activation of AMPK by PTE affected the mTOR signaling pathway. It
was demonstrated that PTE treatment for 48 h could inhibit mTOR
phosphorylation in a dose-dependent manner in RPMI-8226 and ARH-77
cells. The best understood roles of mTOR in mammalian cells are
regarding the control of mRNA translation by phosphorylating 4E-BP1
(20), it was observed that the
inhibition of mTOR phosphorylation was accompanied by the decreased
phosphorylation of 4E-BP1. Furthermore, eIF2α phosphorylation was
increased in a dose-dependent manner (Fig. 2C). These results indicated that
PTE activates AMPK phosphorylation to suppress the expression level
and activity of lipogenesis- and mRNA translation-associated
enzymes, potentially driving MM cells into a low nutrient
state.
PTE-induced apoptosis is dependent on the
activation of AMPK in MM cells
To more definitively establish the role of AMPK in
mediating the apoptosis-promoting effect of PTE, a well-established
pharmacological inhibitor of AMPK activity, compound C, was
utilized. RPMI-8226 cells were treated with 60 µM PTE and
ARH-77 cells with 40 µM PTE, in the absence or presence of
compound C (1 µM). As expected, treatment with PTE alone
activated AMPK phosphorylation. In addition, the expression levels
of the lipogenesis-associated key enzyme FASN were decreased, and
the activation of the mRNA translation-associated 4E-BP1 was
suppressed in RPMI-8226 and ARH-77 cells after 48 h. However, the
FASN expression level and 4E-BP1 activity were restored in the
presence of compound C (Fig.
3A-C). Furthermore, pre-treating RPMI-8226 and ARH-77 cells
with compound C for 1 h and then exposing the cells to PTE for 24 h
partially reduced the ability of PTE to induce apoptosis (Fig. 3D). These results coincided with
the expression of the pro-apoptotic protein, cleaved PARP.
Following treatment with PTE for 48 h, the protein expression
levels of cleaved PARP were increased, which were restored by
co-treatment with compound C (Fig.
3E). Taken together, these results demonstrated that PTE
decreased the expression level and activity of lipogenesis- and
mRNA translation-associated proteins through the activation of the
AMPK pathway, and the activation of AMPK at least in part, induced
MM cells apoptosis.
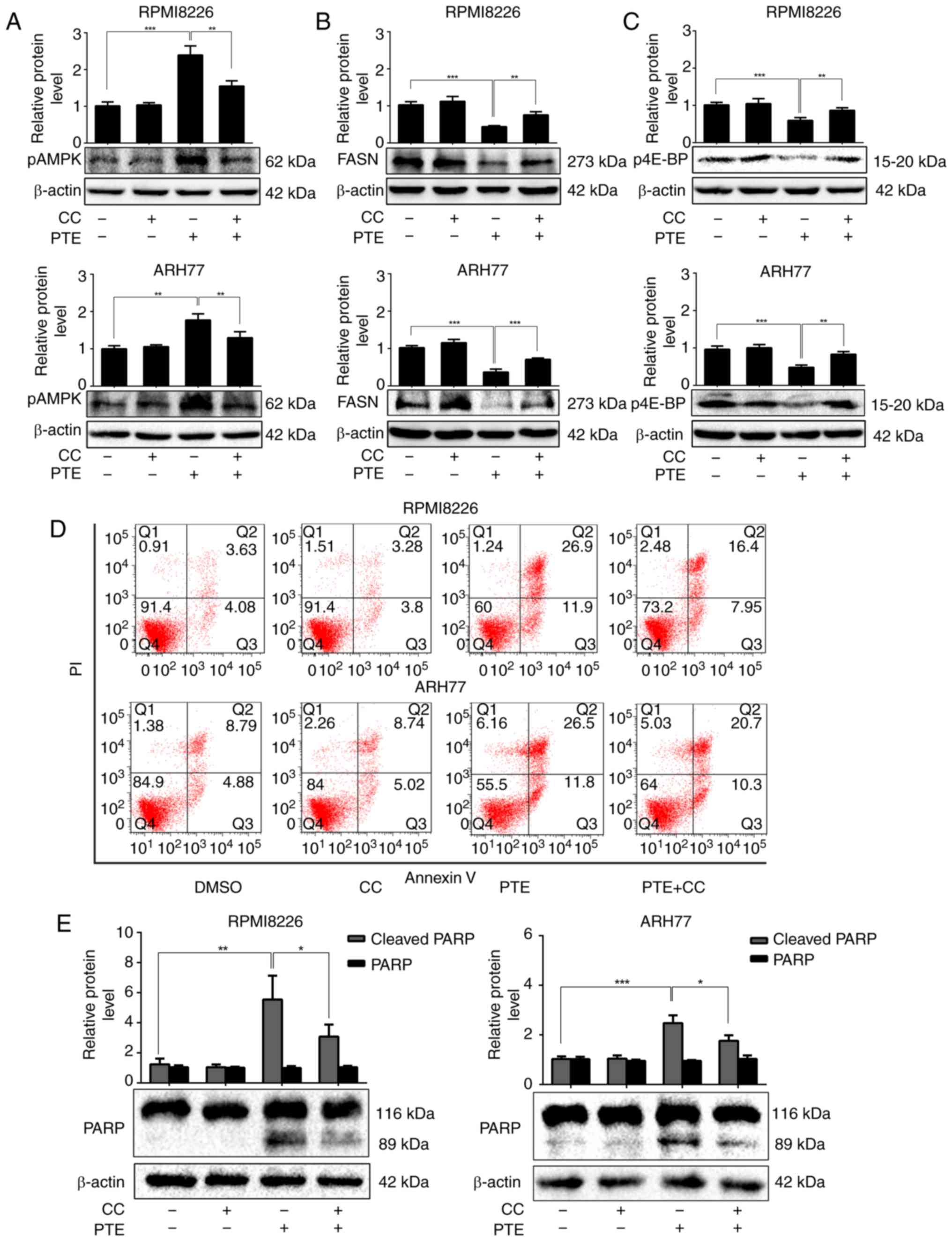 | Figure 3PTE induces MM cells apoptosis in a
AMPK-dependent manner. RPMI-8226 and ARH-77 cells were treated with
PTE in the absence or presence of CC. Western blotting analyzed the
levels of (A) p-AMPK (Thr 172), (B) FASN and (C) p-4E-BP1. β-actin
served as a loading control; (D) Flow cytometric analysis of
apoptotic effects using an Annexin V-FITC/PI kit. (E) Western blot
analysis of the levels of PARP. β-actin served as a loading
control. *P<0.05, **P<0.01,
***P<0.001 vs. PTE-only group. PTE, pterostilbene;
MM, multiple myeloma; AMPK, AMP-activated protein kinase; CC,
compound C; p, phosphorylated; FASN, fatty acid synthase; 4E-BP1,
4E-binding protein 1; FITC, fluorescein isothiocyanate; PARP,
poly(ADP-ribose) polymerase. |
PTE activates autophagic processes in MM
cells
Autophagy is activated when cellular energy is
insufficient. The AMPK/mTOR pathway is involved in the regulation
of autophagy in cancer cells (36,37). Therefore, the effect of PTE on
autophagy was assessed. Using TEM, the gold standard for autophagy
assessment, the majority of the RPMI-8226 cells in the control
group were structurally complete, whereas PTE treatment resulted in
an increase in autophagosomes (Fig.
4A). MDC is a specific label for autophagic vacuoles that was
used to further identify PTE-induced autophagy (38). The PTE-treated RPMI-8226 cells
displayed a greater fluorescence intensity and a greater number of
MDC-labeled particles compared with the untreated control group
(Fig. 4B). Furthermore, the
western blotting analysis of cell lysates revealed the increased
accumulation of autophagy markers, including LC3-II, beclin1 and
ATG5, in a dose-dependent manner at 48 h (Fig. 4C and D). Autophagy can serve
either a pro-survival or pro-death function (39). Its role in the effect of PTE
intervention on MM cells was then demonstrated. The pro-apoptotic
effect of PTE was further enhanced subsequent to blocking
autophagic flux by co-treatment with 3 mM 3MA for 24 h (Fig. 4E), indicating that PTE-induced
autophagy is a cell protective mechanism, as the inhibition of
autophagy aggravated PTE-induced apoptosis. These data confirmed
that PTE induced autophagy in MM cells. The co-presence of PTE and
an autophagy inhibitor improved the anti-MM activity compared with
PTE alone.
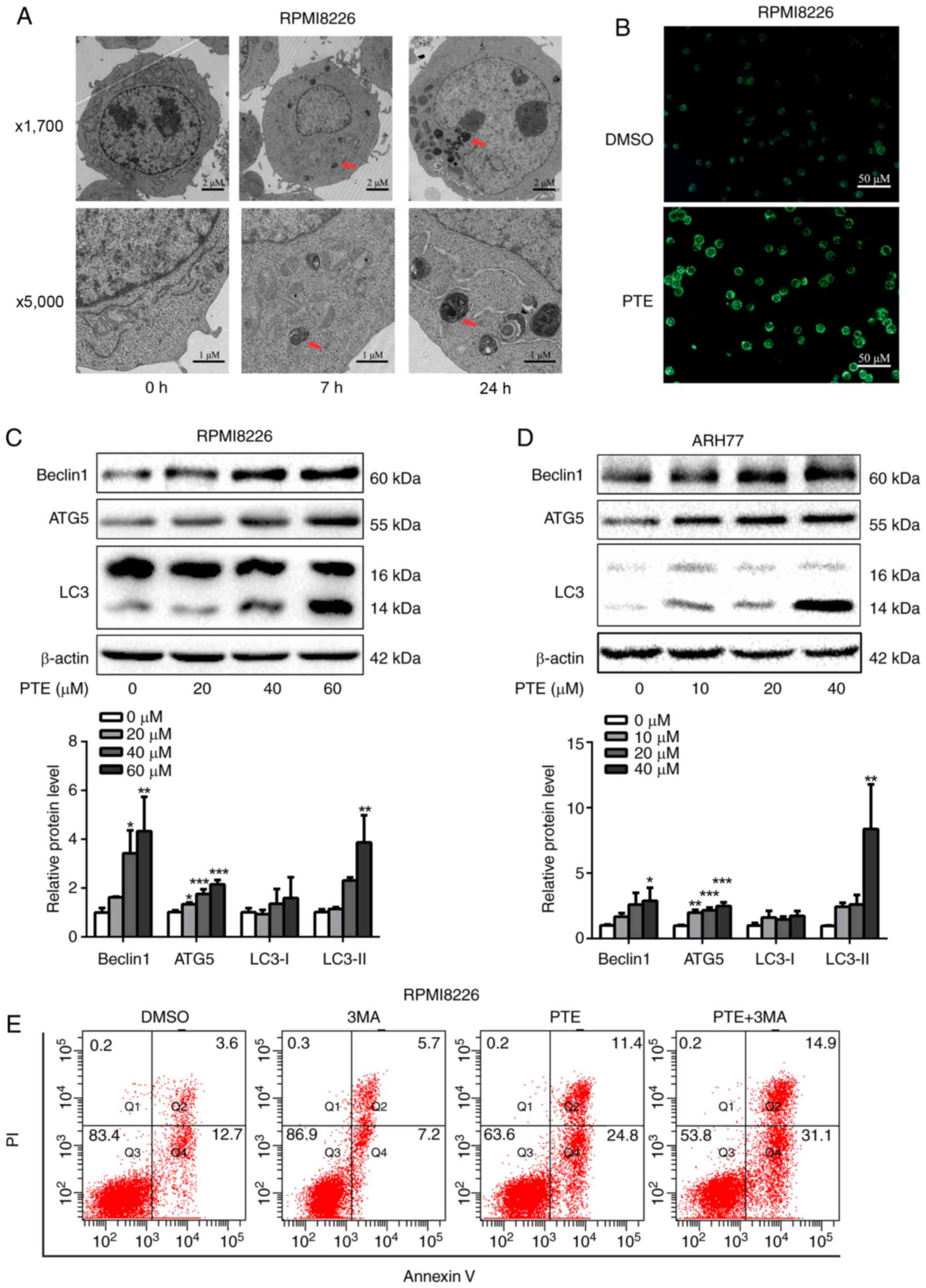 | Figure 4PTE treatment induces autophagy. (A)
Transmission electronic microscopy analyzed the
autophagy-associated morphological change in RPMI-8226 cells
following exposure to PTE. Upper panel, ×1,700 magnification; lower
panel, ×5,000 magnification. (B) Monodansylcadaverine-labeled
vacuoles were observed in RPMI-8226 cells using a fluorescence
microscope. (C) RPMI-8226 and (D) ARH-77 cells were treated with
DMSO (control) or different concentrations of PTE for 48 h, western
blotting was applied to investigate the levels of the autophagy
markers beclin1, ATG5 and LC3. β-actin was used as a loading
control. (E) Effect of PTE treatment on RPMI-8226 cells with or
without 3MA was analyzed by flow cytometry using an Annexin
V-FITC/PI kit. *P<0.05, **P<0.01,
***P<0.001 vs. DMSO group. PTE, pterostilbene; ATG5,
autophagy-related 5; LC3, light chain 3; 3MA, 3-methyladenine;
FITC, fluorescein isothiocyanate; PI, propidium iodide. |
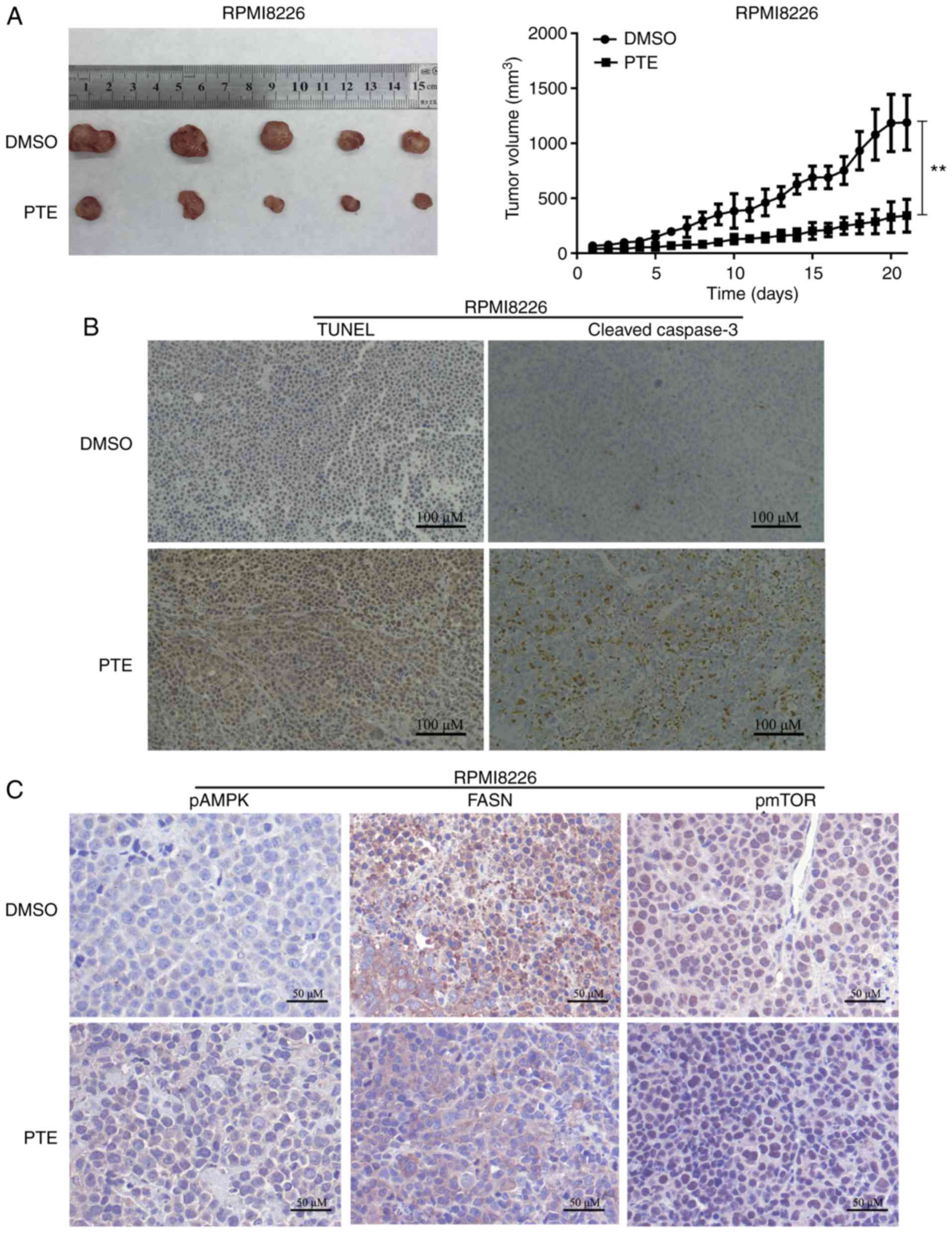
PTE inhibits tumor growth in vivo
The in vivo antitumor effect of PTE was then
examined using a mouse model. RPMI-8226 cells were subcutaneously
injected into 3-week-old female NOD/SCID mice. As demonstrated in
Fig. 5A, the xenograft tumors in
the mice receiving PTE treatment exhibited a reduced tumor volume
compared with the mice receiving DMSO. The tumor xenografts were
harvested following the final treatment, and TUNEL assays and IHC
were performed. From the in situ TUNEL assays, it was
observed that tumors in mice treated with PTE had increased levels
of apoptotic cells compared with control mice (Fig. 5B). IHC was performed to
investigate the molecular mechanisms involved in the
anti-tumorigenic effects of PTE. An increase in cleaved-caspase 3
staining confirmed the induction of apoptosis in the treatment
group (Fig. 5B). Similarly, an
increase in p-AMPK-positive cells and a decrease in FASN- and
p-mTOR-positive cells were observed with IHC in the xenograft
tumors of the PTE treatment group (Fig. 5C). The in vivo data
demonstrated the promising anti-tumor effect of PTE for the
treatment of MM, associated with metabolic dependence, implying
that PTE may be an effective MM treatment.
PTE displays no evident toxicity in
vivo
Mice treated with PTE exhibited no notable change in
body weight, food and water intake, or activity.
Chemotherapy-induced heart disease, liver dysfunction or kidney
malformation are common side effects that may restrict the clinical
application of certain chemotherapeutic agents. Therefore, it was
assessed whether the administration of PTE to tumor-bearing mice
stimulated cardiac, nephritic or hepatic toxicity. Mouse plasma was
collected at sacrifice, and mouse plasma myocardial damage and
liver/kidney disease markers were quantified. The results revealed
that the administration of PTE did not increase liver damage
markers, including ALT, AST, ALP and TBIL (Fig. 6A; P>0.05). No significant
increase in the kidney dysfunction indicators in BUN and Scr was
observed (Fig. 6B; P>0.05).
PTE treatment also did not result in cardiac damage, as no increase
in cTnT or CK-MB was observed (Fig.
6C; P>0.05). Taken together, these results indicated that
PTE treatment had no evident toxic effects on the heart, liver or
kidneys.
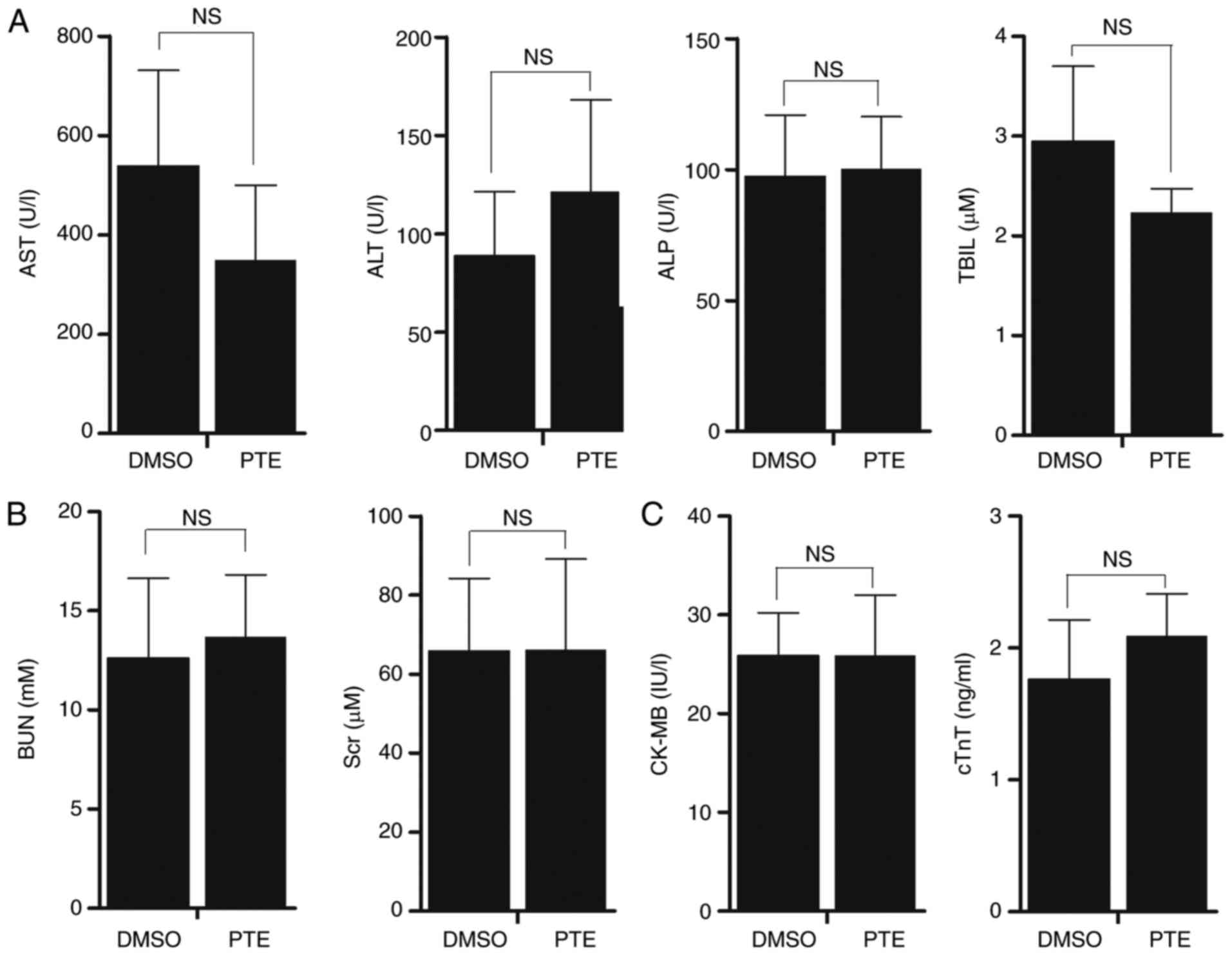 | Figure 6PTE is a safe drug for the treatment
of MM. (A) Liver damage markers, including ALT, AST, ALP and TBIL
were measured in serum samples from NOD/SCID mice treated with 5%
DMSO or PTE with an automatic biochemical analyzer. (B) Kidney
function markers BUN and Scr were measured in serum samples from
NOD/SCID mice treated with 5% DMSO or PTE by an automatic
biochemical analyzer. (C) Cardiac injury markers cTnT and CK-MB
were measured with an ELISA kit. The values are expressed as means
± standard deviation. PTE, pterostilbene; MM, multiple myeloma;
ALT, alanine aminotransferase; AST, aspartate amino transferase;
ALP, alkaline phosphatase; TBIL, total bilirubin; DMSO, dimethyl
sulfoxide; BUN, urea nitrogen; Scr, serum creatinine; cTnT, cardiac
troponins T; CK-MB, creatine kinase-MB. |
Discussion
Multiple myeloma (MM) is an aggressive type of
cancer. Despite notable advances with the advent of proteasome
inhibitors and immunomodulatory agents, management of MM remains
challenging, and relapse of MM and disease progression is common
even following achievement of complete remission (2,40).
Multiple studies have reported that natural products derived from
plants are important sources of drugs that may offer significant
treatment for the majority of cancer types, including colorectal
and oral cancer (41,42). Consistent with previous studies,
in which it was observed that PTE inhibited MM cell viability and
induced cell cycle arrest through regulating the mitogen-activated
protein kinase pathway (43,44), the present study demonstrated that
the administration of PTE could induce MM apoptosis in
vitro, and decrease tumor burden in vivo. Additionally,
to the best of our knowledge, the present study was the first to
report that PTE-induced MM apoptosis was metabolically dependent,
via the activation of AMPK, in vitro and in vivo.
This study suggested that the antitumor effect of PTE may provide a
novel therapeutic option for patients with MM.
Nagao et al (45) reported that the administration of
PTE through diet markedly suppressed abdominal white adipose tissue
accumulation in obese rats, suggesting that PTE enhanced energy
expenditure and/or suppressed lipogenesis in obese rats.
Gomez-Zorita et al (7)
also demonstrated that PTE decreased lipogenesis in adipose tissue,
and increased fatty acid oxidation in the liver. However, the
effect of PTE on the metabolic regulation of cancer has yet to be
reported. Our group previously reported that the fate of MM cells
can be modulated by certain metabolism-associated proteins
(46-48), and that leptin, as secreted by
adipocytes, can promote MM cell proliferation (49). Our group also previously observed
that MM cells have an abnormal de novo lipogenesis, with the
increased expression of lipogenesis genes, including FASN,
stearoyl-CoA desaturase 1, and ACC, in MM cell lines and MM patient
tissue, consistent with a number of other cancer cell types
(10,50). Therefore, we hypothesized that PTE
induces MM cell apoptosis by modulating aberrant lipid metabolism.
In the present study, p-AMPK was upregulated by PTE treatment in a
dose-dependent manner. AMPK activation regulates signaling cascades
involved in metabolic events, including lipogenic activity
(35). Following PTE treatment,
the activation of AMPK increased the phosphorylation of ACC and
decreased FASN protein expression, attenuating the expression
levels/activity of these key enzymes in de novo
lipogenesis.
Previous studies have highlighted the emerging role
of AMPK/mTOR in cancer, and the best understood role of mTOR in
mammalian cells is the control of mRNA translation through the
phosphorylation of 4E-BP1 (51,52). In addition to lipogenic
suppression, it was observed that PTE may also act as a
translation-suppressing mimetic. In the present study, it was
observed that PTE treatment downregulated the phosphorylation of
mTOR and, consequently, the dephosphorylation of 4E-BP1. The
regulation of these key steps to translation initiation following
PTE treatment may suppress general mRNA translation in MM cells. It
is established that cancer cells are sensitive to nutrient shortage
(16,34). Once PTE treatment drives MM cells
into a low-nutrient state, the cells may have been killed by the
depletion of intracellular lipids and proteins. The effect of PTE
on metabolic suppression and apoptosis was reversed by the
pharmacological AMPK inhibitor compound C, further confirming the
central role of AMPK in mediating the nutrient suppression and
antineoplastic effects of PTE on MM cells.
Autophagy occurs at physiological levels to
facilitate macromolecular turnover; it is upregulated when cell
homeo-stasis is disrupted through nutrient shortage (53). In the present study, it was
observed at the molecular and cellular levels that PTE evidently
induced autophagy in MM cells. It was then demonstrated that the
pro-apoptotic effect of PTE was enhanced following co-treatment
with autophagy inhibitors, revealing that autophagy exerts a
protective role against PTE-induced apoptosis. Therefore, PTE in
combination with autophagy inhibitors may be a novel alternative
for the treatment of MM. As the AMPK/mTOR pathway is also involved
in the regulation of autophagy in cancer cells (37), we hypothesized that treatment with
PTE drives cells into a low-nutrient state, and that autophagy may
be adaptively upregulated to maintain cell homeostasis through
promoting the lysosomal degradation of damaged or redundant cell
constituents. As a self-protective step in nutrient shortage,
autophagy may promote the survival of MM cells. However, despite
these observations, the underlying mechanism for the effect of
autophagy on survival and apoptosis in conditions of nutrient
shortage due to PTE treatment remains uncharacterized and should be
the subject of further investigation.
The majority of existing treatments for MM are
highly toxic. The classical chemotherapy drugs, including melphalan
and cyclophosphamide, and newly emerging targeted therapy drugs are
all associated with side effects, including heart, lung and liver
disease. In the present study, mice treated with PTE exhibited no
evident signs of toxicity based on body weight, food and water
intake, activity and general examinations. Furthermore, biochemical
analysis demonstrated that PTE treatment did not induce hepatic,
renal or cardiac toxicity, demonstrating that PTE is not only an
effective anti-MM compound, but also a safe drug with minimal
toxicity in vivo. Recent clinical trials investigating the
safety of PTE concluded that it was generally safe at doses of up
to 250 mg/day in humans (54).
Previous study reported that mice tolerated PTE at doses of 250
mg/kg five times per week for 21 continuous weeks presented no
measurable toxicity (55). In the
present study, the PTE dose used for mice was 50 mg/kg, which is
well within the reported safety range. This demonstrated that PTE
not only possesses great anti-MM potential, but that it also may be
well-tolerated and safe for patients with MM.
In recent decades, the abnormal metabolic profiles
of cancer cells have been extensively studied, with the goal of
identifying clinically targeted therapies. Glycolysis inhibitor,
2-deoxy-glucose, has reached phase II clinical trials (56). Statins, a class of drugs used to
lower cholesterol levels, has been proven to inhibit cancer
development and progression (57). Numerous data suggested that use of
the standard antidiabetic agent metformin decreases cancer
incidence and mortality, predominately through AMPK signal
activation, including in MM cells (58). The data from the present study
demonstrated that PTE could be a naturally derived AMPK activator
in MM cells, thereby disrupting abnormal metabolic processes and
further inducing the apoptosis of MM cells. The effectiveness and
safety of PTE indicates its potential for clinical use.
Notably, articles published in the 1990s have
indicated that, although established from cells taken from a
patient with a plasma cell leukemia (PCL), ARH-77 is actually an
EBV-transformed B lymphoblastoid cell line (59,60). However, ARH-77 cells have
continued to be extensively used as a myeloma model or a PCL model
over the past two decades, and has proven valuable in understanding
the progression of MM, as well as for the testing of novel
therapeutic strategies (61-64). Due to the controversy, attention
should be paid when interpreting data from ARH-77 cells.
In conclusion, to the best of our knowledge, the
present study was the first to demonstrate that PTE decreases lipid
and protein synthesis through the activation of the AMPK pathway,
which directly leads to the apoptosis of MM cells. Following PTE
treatment, autophagy is adaptively upregulated, and co-treatment
with an autophagy inhibitor may further potentiate the anti-MM
efficacy of PTE (Fig. 7). Taken
together, the data provided an improved understanding of the
association between nutrient metabolism, autophagy and apoptosis in
PTE intervention. The present study indicated that the
antineoplastic activity of PTE on MM cells could provide a novel
therapeutic option for patients with MM.
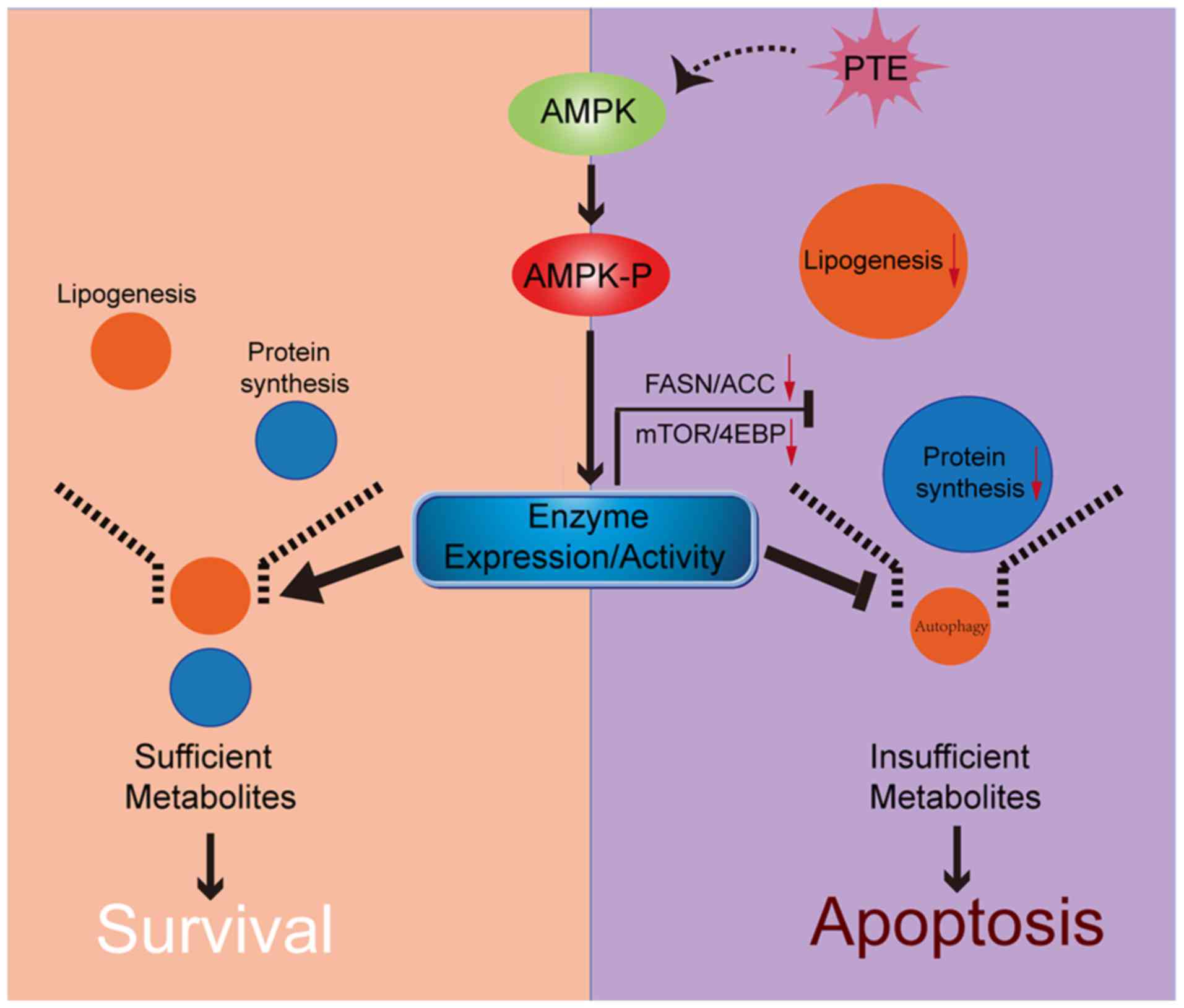 | Figure 7Schematic of the association between
AMPK, nutrient metabolism, autophagy and apoptosis. PTE treatment
results in the phosphorylation and thus, activation, of AMPK.
Activated AMPK directly suppresses the expression levels and
activity of lipogenesis and mRNA translation-related enzymes,
inducing apoptosis in MM cells. Additionally, autophagy is
adaptively upregulated, to protect MM cells during nutrient
shortage. AMPK, AMP-activated protein kinase; PTE, pterostilbene;
MM, multiple myeloma. |
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
TG and HuM designed the project. HuM, YX and QW
performed the experiments. HuM, YX and DC analyzed the data. HuM
and BF prepared all the figures. HuM, YX and BF wrote the
manuscript. HM, TG and YH guided and supervised the study. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Committee on
Animals Handling of Huazhong University of Science and Technology
(Wuhan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
Acknowledgments
The authors would like to thank the members at the
Collaborative Innovation Center of Hematology, China for providing
assistance with statistical analysis, and the Wuhan Institute of
Virology, Chinese Academy of Sciences for providing assistance with
TEM.
References
|
1
|
Rajkumar SV, Dimopoulos MA, Palumbo A,
Blade J, Merlini G, Mateos MV, Kumar S, Hillengass J, Kastritis E,
Richardson P, et al: International myeloma working group updated
criteria for the diagnosis of multiple myeloma. Lancet Oncol.
15:e538-e5482014. View Article : Google Scholar
|
|
2
|
Dimopoulos MA, Richardson PG, Moreau P and
Anderson KC: Current treatment landscape for relapsed and/or
refractory multiple myeloma. Nat Rev Clin Oncol. 12:42–54. 2015.
View Article : Google Scholar
|
|
3
|
Estrela JM, Ortega A, Mena S, Rodriguez ML
and Asensi M: Pterostilbene: Biomedical applications. Crit Rev Clin
Lab Sci. 50:65–78. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang P and Sang S: Metabolism and
pharmacokinetics of resveratrol and pterostilbene. Biofactors.
44:16–25. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kosuru R, Rai U, Prakash S, Singh A and
Singh S: Promising therapeutic potential of pterostilbene and its
mechanistic insight based on preclinical evidence. Eur J Pharmacol.
789:229–243. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wakimoto R, Ono M, Takeshima M, Higuchi T
and Nakano S: Differential anticancer activity of pterostilbene
against three subtypes of human breast cancer cells. Anticancer
Res. 37:6153–6159. 2017.PubMed/NCBI
|
|
7
|
Gomez-Zorita S, Fernandez-Quintela A, Lasa
A, Aguirre L, Rimando AM and Portillo MP: Pterostilbene, a dimethyl
ether derivative of resveratrol, reduces fat accumulation in rats
fed an obesogenic diet. J Agric Food Chem. 62:8371–8378. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kosuru R and Singh S: Pterostilbene
ameliorates insulin sensitivity, glycemic control and oxidative
stress in fructose-fed diabetic rats. Life Sci. 182:112–121. 2017.
View Article : Google Scholar
|
|
9
|
Lin VC, Tsai YC, Lin JN, Fan LL, Pan MH,
Ho CT, Wu JY and Way TD: Activation of AMPK by pterostilbene
suppresses lipogenesis and cell-cycle progression in p53 positive
and negative human prostate cancer cells. J Agric Food Chem.
60:6399–6407. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang MD, Wu H, Fu GB, Zhang HL, Zhou X,
Tang L, Dong LW, Qin CJ, Huang S, Zhao LH, et al: Acetyl-coenzyme A
carboxylase alpha promotion of glucose-mediated fatty acid
synthesis enhances survival of hepatocellular carcinoma in mice and
patients. Hepatology. 63:1272–1286. 2016. View Article : Google Scholar
|
|
11
|
Okawa Y, Hideshima T, Ikeda H, Raje N,
Vallet S, Kiziltepe T, Yasui H, Enatsu S, Pozzi S, Breitkreutz I,
et al: Fatty acid synthase is a novel therapeutic target in
multiple myeloma. Br J Haematol. 141:659–671. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Flaveny CA, Griffett K, El-Gendy Bel D,
Kazantzis M, Sengupta M, Amelio AL, Chatterjee A, Walker J, Solt
LA, Kamenecka TM and Burris TP: Broad anti-tumor activity of a
small molecule that selectively targets the warburg effect and
lipogenesis. Cancer Cell. 28:42–56. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zaytseva YY, Rychahou PG, Gulhati P,
Elliott VA, Mustain WC, O'Connor K, Morris AJ, Sunkara M, Weiss HL,
Lee EY and Evers BM: Inhibition of fatty acid synthase attenuates
CD44-associated signaling and reduces metastasis in colorectal
cancer. Cancer Res. 72:1504–1517. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li L, Pilo GM, Li X, Cigliano A, Latte G,
Che L, Joseph C, Mela M, Wang C, Jiang L, et al: Inactivation of
fatty acid synthase impairs hepatocarcinogenesis driven by AKT in
mice and humans. J Hepatol. 64:333–341. 2016. View Article : Google Scholar :
|
|
15
|
Svensson RU, Parker SJ, Eichner LJ, Kolar
MJ, Wallace M, Brun SN, Lombardo PS, Van Nostrand JL, Hutchins A,
Vera L, et al: Inhibition of acetyl-CoA carboxylase suppresses
fatty acid synthesis and tumor growth of non-small-cell lung cancer
in preclinical models. Nat Med. 22:1108–1119. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Medina EA, Oberheu K, Polusani SR, Ortega
V, Velagaleti GV and Oyajobi BO: PKA/AMPK signaling in relation to
adiponectin's antiproliferative effect on multiple myeloma cells.
Leukemia. 28:2080–2089. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zismanov V, Attar-Schneider O, Lishner M,
Heffez Aizenfeld R, Tartakover Matalon S and Drucker L: Multiple
myeloma proteostasis can be targeted via translation initiation
factor eIF4E. Int J Oncol. 46:860–870. 2015. View Article : Google Scholar
|
|
18
|
Agnelli L, Fabris S, Bicciato S, Basso D,
Baldini L, Morabito F, Verdelli D, Todoerti K,
Lambertenghi-Deliliers G, Lombardi L and Neri A: Upregulation of
translational machinery and distinct genetic subgroups characterise
hyperdiploidy in multiple myeloma. Br J Haematol. 136:565–573.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Holcik M and Sonenberg N: Translational
control in stress and apoptosis. Nat Rev Mol Cell Biol. 6:318–327.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Moschetta M, Reale A, Marasco C, Vacca A
and Carratu MR: Therapeutic targeting of the mTOR-signalling
pathway in cancer: Benefits and limitations. Br J Pharmacol.
171:3801–3813. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Morotomi-Yano K, Oyadomari S, Akiyama H
and Yano K: Nanosecond pulsed electric fields act as a novel
cellular stress that induces translational suppression accompanied
by eIF2alpha phosphorylation and 4E BP1 dephosphorylation. Exp Cell
Res. 318:1733–1744. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mihaylova MM and Shaw RJ: The AMPK
signalling pathway coordinates cell growth, autophagy and
metabolism. Nat Cell Biol. 13:1016–1023. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Garcia D and Shaw RJ: AMPK: Mechanisms of
cellular energy sensing and restoration of metabolic balance. Mol
Cell. 66:789–800. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Duan W, Chen K, Jiang Z, Chen X, Sun L, Li
J, Lei J, Xu Q, Ma J, Li X, et al: Desmoplasia suppression by
metformin-mediated AMPK activation inhibits pancreatic cancer
progression. Cancer Lett. 385:225–233. 2017. View Article : Google Scholar
|
|
25
|
Zadra G, Photopoulos C, Tyekucheva S,
Heidari P, Weng QP, Fedele G, Liu H, Scaglia N, Priolo C, Sicinska
E, et al: A novel direct activator of AMPK inhibits prostate cancer
growth by blocking lipogenesis. EMBO Mol Med. 6:519–538. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dong GZ, Lee YI, Jeong JH, Zhao HY, Jeon
R, Lee HJ and Ryu JH: Stilbenoids from rheum undulatum protect
hepatocytes against oxidative stress through AMPK activation.
Phytother Res. 29:1605–1609. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang L, Cui L, Zhou G, Jing H, Guo Y and
Sun W: Pterostilbene, a natural small-molecular compound, promotes
cytoprotective macroautophagy in vascular endothelial cells. J Nutr
Biochem. 24:903–911. 2013. View Article : Google Scholar
|
|
28
|
Dhar S, Kumar A, Zhang L, Rimando AM, Lage
JM, Lewin JR, Atfi A, Zhang X and Levenson AS: Dietary
pterostilbene is a novel MTA1-targeted chemopreventive and
therapeutic agent in prostate cancer. Oncotarget. 7:18469–18484.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
McCormack DE, Mannal P, McDonald D, Tighe
S, Hanson J and McFadden D: Genomic analysis of pterostilbene
predicts its antiproliferative effects against pancreatic cancer in
vitro and in vivo. J Gastrointest Surg. 16:1136–1143. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wawszczyk J, Kapral M, Hollek A and
Weglarz L: In vitro evaluation of antiproliferative and cytotoxic
properties of pterostilbene against human colon cancer cells. Acta
Pol Pharm. 71:1051–1055. 2014.
|
|
31
|
Liu Y, Wang L, Wu Y, Lv C, Li X, Cao X,
Yang M, Feng D and Luo Z: Pterostilbene exerts antitumor activity
against human osteosarcoma cells by inhibiting the JAK2/STAT3
signaling pathway. Toxicology. 304:120–131. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Maiso P, Huynh D, Moschetta M, Sacco A,
Aljawai Y, Mishima Y, Asara JM, Roccaro AM, Kimmelman AC and
Ghobrial IM: Metabolic signature identifies novel targets for drug
resistance in multiple myeloma. Cancer Res. 75:2071–2082. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ma J, Duan W, Han S, Lei J, Xu Q, Chen X,
Jiang Z, Nan L, Li J, Chen K, et al: Ginkgolic acid suppresses the
development of pancreatic cancer by inhibiting pathways driving
lipogenesis. Oncotarget. 6:20993–21003. 2015.PubMed/NCBI
|
|
34
|
Fernando RC, de Carvalho F, Mazzotti DR,
Evangelista AF, Braga WMT, de Lourdes Chauffaille M, Leme AFP and
Colleoni GWB: Multiple myeloma cell lines and primary tumors
proteoma: Protein biosynthesis and immune system as potential
therapeutic targets. Genes Cancer. 6:462–471. 2015.
|
|
35
|
Steinberg GR and Kemp BE: AMPK in health
and disease. Physiol Rev. 89:1025–1078. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lv S, Xu Q, Sun E, Zhang J and Wu D:
Impaired cellular energy metabolism contributes to
bluetongue-virus-induced autophagy. Arch Virol. 161:2807–2811.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jing K, Song KS, Shin S, Kim N, Jeong S,
Oh HR, Park JH, Seo KS, Heo JY, Han J, et al: Docosahexaenoic acid
induces autophagy through p53/AMPK/mTOR signaling and promotes
apoptosis in human cancer cells harboring wild-type p53. Autophagy.
7:1348–1358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang Y, Han C, Lu L, Magliato S and Wu T:
Hedgehog signaling pathway .regulates autophagy in human
hepatocellular carcinoma cells. Hepatology. 58:995–1010. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Levy JMM, Towers CG and Thorburn A:
Targeting autophagy in cancer. Nat Rev Cancer. 17:528–542. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chim CS, Kumar SK, Orlowski RZ, Cook G,
Richardson PG, Gertz MA, Giralt S, Mateos MV, Leleu X and Anderson
KC: Management of relapsed and refractory multiple myeloma: Novel
agents, antibodies, immunotherapies and beyond. Leukemia.
32:252–262. 2018. View Article : Google Scholar :
|
|
41
|
Yao Z, Xie F, Li M, Liang Z, Xu W, Yang J,
Liu C, Li H, Zhou H and Qu LH: Oridonin induces autophagy via
inhibition of glucose metabolism in p53-mutated colorectal cancer
cells. Cell Death Dis. 8:e26332017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chang CH, Lee CY, Lu CC, Tsai FJ, Hsu YM,
Tsao JW, Juan YN, Chiu HY, Yang JS and Wang CC: Resveratrol-induced
autophagy and apoptosis in cisplatin-resistant human oral cancer
CAR cells: A key role of AMPK and Akt/mTOR signaling. Int J Oncol.
50:873–882. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xie B, Xu Z, Hu L, Chen G, Wei R, Yang G,
Li B, Chang G, Sun X, Wu H, et al: Pterostilbene inhibits human
multiple myeloma cells via ERK1/2 and JNK pathway in vitro and in
vivo. Int J Mol Sci. 17:E19272016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen G, Xu Z, Chang G, Hou J, Hu L, Zhang
Y, Yu D, Li B, Chang S, Xie Y, et al: The blueberry component
pterostilbene has potent anti-myeloma activity in
bortezomib-resistant cells. Oncol Rep. 38:488–496. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nagao K, Jinnouchi T, Kai S and Yanagita
T: Pterostilbene, a dimethylated analog of resveratrol, promotes
energy metabolism in obese rats. J Nutr Biochem. 43:151–155. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cao D, Zhou H, Zhao J, Jin L, Yu W, Yan H,
Hu Y and Guo T: PGC-1α integrates glucose metabolism and
angiogenesis in multiple myeloma cells by regulating VEGF and
GLUT-4. Oncol Rep. 31:1205–1210. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yu W, Cao D, Zhou H, Hu Y and Guo T:
PGC-1α is responsible for survival of multiple myeloma cells under
hyperglycemia and chemotherapy. Oncol Rep. 33:2086–2092. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cao D, Jin L, Zhou H, Yu W, Hu Y and Guo
T: Inhibition of PGC-1α after chemotherapy-mediated insult confines
multiple myeloma cell survival by affecting ROS accumulation. Oncol
Rep. 33:899–904. 2015. View Article : Google Scholar
|
|
49
|
Yu W, Cao DD, Li QB, Mei HL, Hu Y and Guo
T: Adipocytes secreted leptin is a pro-tumor factor for survival of
multiple myeloma under chemotherapy. Oncotarget. 7:86075–86086.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Pandey PR, Xing F, Sharma S, Watabe M, Pai
SK, Iiizumi-Gairani M, Fukuda K, Hirota S, Mo YY and Watabe K:
Elevated lipogenesis in epithelial stem-like cell confers survival
advantage in ductal carcinoma in situ of breast cancer. Oncogene.
32:5111–5122. 2013. View Article : Google Scholar
|
|
51
|
Inoki K, Kim J and Guan KL: AMPK and mTOR
in cellular energy homeostasis and drug targets. Annu Rev Pharmacol
Toxicol. 52:381–400. 2012. View Article : Google Scholar
|
|
52
|
Shieh JM, Chen YC, Lin YC, Lin JN, Chen
WC, Chen YY, Ho CT and Way TD: Demethoxycurcumin inhibits energy
metabolic and oncogenic signaling pathways through AMPK activation
in triple-negative breast cancer cells. J Agric Food Chem.
61:6366–6375. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ravikumar B, Sarkar S, Davies JE, Futter
M, Garcia-Arencibia M, Green-Thompson ZW, Jimenez-Sanchez M,
Korolchuk VI, Lichtenberg M, Luo S, et al: Regulation of mammalian
autophagy in physiology and pathophysiology. Physiol Rev.
90:1383–1435. 2010. View Article : Google Scholar
|
|
54
|
Riche DM, McEwen CL, Riche KD, Sherman JJ,
Wofford MR, Deschamp D and Griswold M: Analysis of safety from a
human clinical trial with pterostilbene. J Toxicol.
2013:4635952013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen RJ, Tsai SJ, Ho CT, Pan MH, Ho YS, Wu
CH and Wang YJ: Chemopreventive effects of pterostilbene on
urethane-induced lung carcinogenesis in mice via the inhibition of
EGFR-mediated pathways and the induction of apoptosis and
autophagy. J Agric Food Chem. 60:11533–11541. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mohanti BK, Rath GK, Anantha N, Kannan V,
Das BS, Chandramouli BA, Banerjee AK, Das S, Jena A, Ravichandran
R, et al: Improving cancer radiotherapy with 2-deoxy-D-glucose:
Phase I/II clinical trials on human cerebral gliomas. Int J Radiat
Oncol Biol Phys. 35:103–111. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bjarnadottir O, Romero Q, Bendahl PO,
Jirstrom K, Ryden L, Loman N, Uhlen M, Johannesson H, Rose C,
Grabau D and Borgquist S: Targeting HMG-CoA reductase with statins
in a window-of-opportunity breast cancer trial. Breast Cancer Res
Treat. 138:499–508. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wang Y, Xu W, Yan Z, Zhao W, Mi J, Li J
and Yan H: Metformin induces autophagy and G0/G1 phase cell cycle
arrest in myeloma by targeting the AMPK/mTORC1 and mTORC2 pathways.
J Exp Clin Cancer Res. 37:632018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Pellat-Deceunynk C, Amiot M, Bataille R,
Van Riet I, Van Camp B, Omede P and Boccadoro M: Human myeloma cell
lines as a tool for studying the biology of multiple myeloma: A
reappraisal 18 years after. Blood. 86:4001–4002. 1995.
|
|
60
|
Drexler HG, Dirks WG and MacLeod RA: False
human hematopoietic cell lines: Cross-contaminations and
misinterpretations. Leukemia. 13:1601–1607. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Farnoushi Y, Cipok M, Kay S, Jan H, Ohana
A, Naparstek E, Goldstein RS and Deutsch VR: Rapid in vivo testing
of drug response in multiple myeloma made possible by xenograft to
turkey embryos. Br J Cancer. 105:1708–1718. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Lin L, Benson DM Jr, DeAngelis S, Bakan
CE, Li PK, Li C and Lin J: A small molecule, LLL12 inhibits
constitutive STAT3 and IL-6-induced STAT3 signaling and exhibits
potent growth suppressive activity in human multiple myeloma cells.
Int J Cancer. 130:1459–1469. 2012. View Article : Google Scholar
|
|
63
|
Yan H, Wu QL, Sun CY, Ai LS, Deng J, Zhang
L, Chen L, Chu ZB, Tang B, Wang K, et al: piRNA-823 contributes to
tumorigenesis by regulating de novo DNA methylation and
angiogenesis in multiple myeloma. Leukemia. 29:196–206. 2015.
View Article : Google Scholar
|
|
64
|
Besse A, Stolze SC, Rasche L, Weinhold N,
Morgan GJ, Kraus M, Bader J, Overkleeft HS, Besse L and Driessen C:
Carfilzomib resistance due to ABCB1/MDR1 overexpression is overcome
by nelfinavir and lopinavir in multiple myeloma. Leukemia.
32:391–401. 2018. View Article : Google Scholar :
|