Introduction
Cartilage tissues are degenerated and destroyed
during osteoarthritis and cartilage cannot be repaired by itself.
Joint replacement can efficiently relieve the pain symptoms;
however, the implant can become loose and the cost is high
(1). Cartilage tissue engineering
has been suggested as a promising therapy for osteoarthritis
(2). Among the candidates for the
regeneration of articular cartilage, human adipose-derived stem
cells (hADSCs) are a valuable resource. HADSCs are a population of
self-renewing and multipotent cells, which are of clinical
significance in cellular therapies for tissue regeneration
(3,4). HADSCs are capable of differentiating
into several lineages, including chondrocytes (5,6).
As these cells can be readily harvested in abundance through a
relatively non-invasive procedure, they are a valuable resource in
cartilage tissue engineering. However, the regulation of the
chondrogenic differentiation of hADSCs remains to be fully
elucidated. Therefore, investigation of the molecular mechanism
underlying the chondrogenic differentiation of hADSCs is likely to
assist in the treatment of cartilage destruction.
MicroRNAs (miRNAs) are a class of small, non-coding
RNAs, which are ~23 nucleotides in length and usually negatively
regulate target gene expression at the post-transcriptional level
by incomplete complementation to nucleotides within the 3′
untranslated region (3′-UTR) (7).
miRNAs have been found to be involved in diverse biological
processes, including cell proliferation, apoptosis and
differentiation (8). Increasing
evidence shows that miRNAs are involved in the chondrogenic
differentiation of various cell types, particularly in mesenchymal
stem cells from bone marrow (BMSCs) (9). For example, miR-495 was found to be
downregulated during the transforming growth factor
(TGF)-β3-induced chondrogenic differentiation of hMSCs using a
microarray, and further functional investigations have shown that
it inhibits the chondrogenic differentiation of BMSCs by directly
binding and inhibiting the expression of the chondrogenic key
factor sex determining region Y-box (Sox)9 (10). However, miR-410 was found to be
elevated during the TGF-β3-induced chondrogenic differentiation of
MSCs, and it promoted the chondrogenic differentiation of BMSCs by
negatively regulating its target gene, Wnt3a, resulting in
inhibition of the Wnt signaling pathway (11). There is limited data on the
involvement of miRNAs in chondrogenic differentiation hADSCs.
Zhang et al (12) analyzed the miRNA expression
profile of hADSCs during chondrogenic differentiation with miRNA
microarrays and found that 12 miRNAs were differentially expressed.
Our previous study also analyzed the miRNA expression of hADSCs
subjected to chondrogenic induction via microarray, and it was
found that 20 miRNAs were significantly differently expressed. It
was also identified that miR-490 promoted the chondrogenic
differentiation of hADSCs by targeting bone morphogenetic protein
receptor type 2 (BMPR2) (13). A
study by Hou et al (14)
showed that miR-193b inhibited the early chondrogenesis of hADSCs
in vitro, and further mechanistic evaluation showed that it
functioned by targeting TGF-β (TGFB)2 and TGFB receptor 3, and
inhibiting the TGF-β2 signaling pathway. The expression of miR-92a
was elevated in chondrogenic ATDC5 cells and hADSCs, and the
phosphoinositide 3-kinase-Akt, ErbB and focal adhesion kinase
pathways, extracellular matrix-receptor interaction, and mammalian
target of rapamycin signaling pathway may be potential mediators of
the effects of miR-92a on chondrogenesis (15). However, miR-194 was decreased in
chondrogenic differentiation, and it suppressed chondrogenic
differentiation via targeting the transcription factor Sox5
(16). However, evidence of the
role of miR-1307-3p in the chondrogenic differentiation of hADSCs
remains limited.
In our previous study, profiling of the miRNA
expression of hADSCs during chondrogenic differentiation was
performed and the differentially expressed miRNAs verified by
northern blot analysis. Furthermore, the function and mechanism of
miR-490-5p in the chondrogenic differentiation of hADSCs were
characterized (13). Among the
differently expressed miRNAs, the expression of miR-1307-3p was
significantly decreased during chondrogenic differentiation.
However, whether it was involved in chondrogenic differentiation
was unclear. The present study aimed to investigate the potential
role of miR-1307-3p in the chondrogenic differentiation of hADSCs.
The results demonstrated that miR-1307-3p suppressed the
chondrogenic differentiation of hADSCs by inhibiting the
BMPR2-mothers against decapentaplegic (Smad) signaling pathway via
targeting BMPR2, which suggested the potential use of miR-1307-3p
in hADSC-based cartilage tissue engineering.
Materials and methods
Isolation and culture of hADSCs
The hADSCs were isolated from the adipose tissue of
healthy donors during liposuction at Tianjin Haihe Hospital
(Tianjin, China). Written informed consent was obtained from the
donors. The study was approved by the Ethics Committee of Tianjin
Haihe Hospital. Following excision during liposuction, 5 g of
adipose tissue was minced with scissors and digested with type IA
collagenase at 37°C for 30 min with agitation. The digested
supernatants were later centrifuged at 400 x g for 10 min at room
temperature, and the pellet was washed with PBS and subsequently
resuspended with Dulbecco’s modified Eagle’s medium/nutrient
mixture F-12 (DMEM/F12) (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The digestion and collection procedures were
repeated twice, and all of the collected cells were filtered
through a 200-mesh sieve. The cells were later seeded into a
culture dish and cultured in DMEM/F12 medium at 37°C under 5%
CO2. The medium was replaced 24 h later and was replaced
every 3 days thereafter. The cells were passaged at 1:3 when the
confluence reached 80% and cells of the third passage were used for
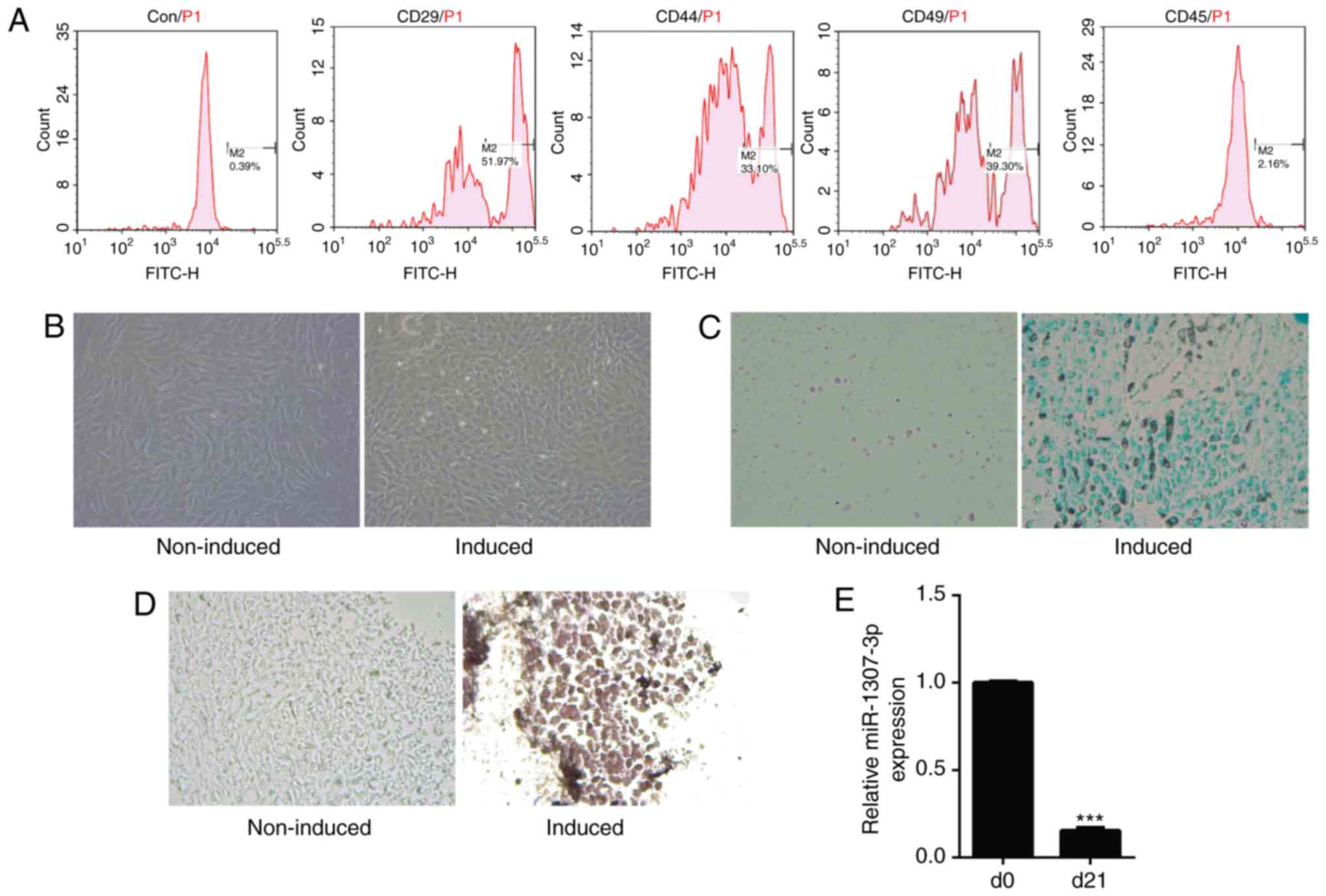
experiments. The characterization of hADSCs was determined by flow
cytometry. The corresponding antibody including rabbit anti-human
antibodies for CD29 (cat. no. 12594-1-AP; 1:100), CD44 (cat. no.
15675-1-AP; 1:100), CD45 (cat. no. 20103-1-AP; 1:100), CD49 (cat.
no. 21992-1-AP; 1:100), and fluorescein isothiocyanate-conjugated
Affinipure Goat Anti-Rabbit IgG (H+L) (cat. no. SA00003-2; 1:100),
which were purchased from ProteinTech Group, Inc. (Wuhan,
China).
Induction of chondrogenic differentiation
of hADSCs using a pellet culture
The induction of chondrogenic differentiation of
hADSCs was performed as previously described (17). Briefly, 5x105 cells of
the fourth generation were suspended in 1 ml basal medium, which
was added to 15-ml polypropylene conical tubes and centrifuged at
500 x g for 5 min at room temperature to form cell pellets. The
pellets were then resuspended with 3 ml chondrogenic medium
consisting of DMEM/F-12, 5 ng/ml fibroblast growth factor-2, 10
ng/ml TGF-β1, 50 µg/ml vitamin C and 10-7 M
dexamethasone. The cells were cultured at 37°C for ~21 days for
differentiation and the chondrogenic medium was replaced every 2
days.
Small interfering RNA (siRNA), lentiviral
vector construction and cell infection
The BMPR2-specific siRNA and control siRNA were
synthesized by GenePharma (Shanghai, China). The siRNAs were
transfected into cells at a final concentration of 50 nM.
Recombinant lentiviruses expressing miR-1307-3p
(HBLV-h-miR-1307-3p-GFP-PURO), miR-1307-3p inhibitor
(HBLV-h-shmiR-1307-3p-GFP-PURO) or control lentivirus were
purchased from Hanbio (Shanghai, China). Lentiviruses at a dose of
2x108 TU/ml and a multiplicity of infection of 10 were
used to infect hADSCs cells according to the manufacturer’s
protocol. Following incubation with the lentivirus for 48 h, the
medium was replaced with fresh medium and the cells were cultured
for further experiments. The infection efficiency was determined by
microscopic examination of GFP-positive cells.
Alcian blue staining
To assess the deposition of cartilage matrix
proteoglycans, the cell pellets were first fixed with 4%
paraformaldehyde and dehydrated with an ethanol gradient.
Subsequently, the cells were paraffin-embedded and cut into
5-µm sections. The sections were first washed with PBS three
times, following which they were immersed in 0.1 N HCL for 5 min
and stained with Alcian blue overnight. Following being washed with
0.1 N HCL for 5 min three times, the sections were examined using a
Leica DM LB2 upright light microscope (Leica Microsystems GmbH,
Wetzlar, Germany).
Immunocytochemical examination
Following the induction of chondrogenesis for 21
days, the cells pellets were fixed with 4% paraformaldehyde,
dehydrated and paraffin-embedded as described above. The sections
were then incubated with blocking solution [PBS/10% goat serum
(Beijing Biosynthesis Biotechnology Co., Ltd., Beijing, China)/0.1%
Triton X-100] for 30 min, and were then incubated with monoclonal
rabbit anti-human antibodies against collagen type II (ColII; cat.
no. bsm-33129M; 1:200; Beijing Biosynthesis Biotechnology Co.,
Ltd.) for 2 h at room temperature and washed with PBS three times.
Subsequently, the sections were incubated with HRP-conjugated
secondary antibody for 1 h at room temperature (cat. no. SA00001-2;
1:1,000; ProteinTech Group, Inc.) and washed with PBS three times.
Subsequently, the sections were incubated with DAB solution for 30
min at room temperature and washed with deionized water for 5 min.
Finally, the sections were stained with hematoxylin to identify the
nuclei. Images were captured with a Leica DM LB2 upright light
microscope.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was isolated using TRIzol®
reagent (Invitrogen, Thermo Fisher Scientific, Inc., Waltham, MA,
USA) according to the manufacturer’s protocol. cDNA was synthesized
using the PrimeScript RT reagent kit (Takara Biotechnology Co.,
Ltd., Dalian, China). RT-qPCR analysis was performed using the SYBR
Premix Ex Taq kit (Takara Biotechnology Co., Ltd.). The PCR (20
µl) included SYBR Green premix (10 µl), cDNA (2
µl), forward primer (10 µM; 1 µl), reverse
primer (10 µM; 1 µl) and H2O (6
µl). The cycles were as follows: 95°C for 30 sec, followed
by 40 cycles of 95°C for 5 sec, and 60°C for 30 sec. The primers
used are listed in Table I. The
results were normalized to the expression of U6 or GAPDH and were
calculated with the 2−ΔΔCq method (18).
 | Table IPrimers and oligonucleotides used in
the present study. |
Table I
Primers and oligonucleotides used in
the present study.
| Primer | Sequence
(5′-3′) |
|---|
| miR-1307-3p-RT |
GTCGTATCCAGTGCAGGGTCCGAGGTGCACTGGATACGACCACGACC |
| U6-RT |
AACGCTTCACGAATTTGCGTG |
|
miR-1307-3p-qPCR | Forward:
TGCGGGTCCAGTTTTCCCAGGAA |
| Reverse:
CCAGTGCAGGGTCCGAGGT |
| U6-qPCR | Forward:
GCTCGCTTCGGCAGCACA |
| Reverse:
GAGGTATTCGCACCAGAGGA |
| BMPR2-qPCR | Forward:
TGGGAAAGAAACAAATCTGTGAGC |
| Reverse:
GAGGAGGAAGAATAATCTGGATAAGGAC |
| Smad1-qPCR | Forward:
CACCTGCTTACCTGCCTCCTGAA |
| Reverse:
GCAACCGCCTGAACATCTCCTCT |
| Smad2-qPCR | Forward:
ATGAATTAAATCAGAGGGTTGGAG |
| Reverse:
CTTCTTGTCATTTCTACCGTGGC |
| Smad3-qPCR | Forward:
GTGACCACCAGATGAACCACAGCA |
| Reverse:
CGTAGTAGGAGATGGAGCACCAGAAGG |
| Smad4-qPCR | Forward:
TCCAGCCTCCCATTTCCAATCAT |
| Reverse:
CTCCAGAAGGGTCCACGTATCCA |
| Smad5-qPCR | Forward:
AAAACACTAGGCGACATATTGGA |
| Reverse:
GGAATCTTACAGACAGTGGTGGG |
| Smad6-qPCR | Forward:
AGTGACTGCGAGACGGTGACCTGCTG |
| Reverse:
GCGAGTACGTGACGGTTTTGAGTTCCTG |
| Smad7-qPCR | Forward:
TCTCAGGCATTCCTCGGAAGTCAAG |
| Reverse:
CTAGTTCGCAGAGTCGGCTAAGGTGAT |
| Smad8-qPCR | Forward:
GAGGTGTATGCCGAGTGCGTGAG |
| Reverse:
GCTGAGCGAAGAGCTGGTTGTTG |
| Noggin-qPCR | Forward:
GCCATGCCGAGCGAGATCAAAGGG |
| Reverse:
CGACCACAGCCACATCTGTAACTTCCTCC |
| Chordin-qPCR | Forward:
GATGCCCTGATCCCAGTCCAGACG |
| Reverse:
GCCACCACCTCACTGCTTGTCCCTAC |
| GAPDH-qPCR | Forward:
ATGACATCAAGAAGGTGGTGAAGCAGG |
| Reverse:
GCGTCAAAGGTGGAGGAGTGGGT |
| miRNA-NC
mimics | Forward:
UCACAACCUCCUAGAAAGAGUAGA |
| Reverse:
UCUACUCUUUCUAGGAGGUUGUGA |
| miR-1307-3p
mimics | Forward:
ACUCGGCGUGGCGUCGGUCGUG |
| Reverse:
CACGACCGACGCCACGCCGAGU |
| BMPR2 3′UTR wt | Forward:
CTAGCAGGUUGUUGCAGUGAGCCGAGG |
| Reverse:
TCGACCTCGGCTCACTGCAACAACCTG |
| BMPR2 3′UTR
mut | Forward:
CTAGCAGGUUGUUGCAGUGACGGCTCG |
| Reverse:
TCGACGAGCCGTCACTGCAACAACCTG |
| Si-BMPR2 | Sense:
GCAUGUUUGAUUCCUGAUGTT |
| Antisense:
CGUACAAACUAAGGACUACTT |
| Control siRNA | Sense:
UUCUCCGAACGUGUCACGUTT |
| Antisense:
ACGUGACACGUUCGGAGAATT |
Western blot analysis
The cells were lysed with RIPA buffer, and the
protein concentration was determined with the Bradford assay
(Bio-Rad Laboratories, Hercules, CA, USA). An equal amount of
protein (30 µg) was separated by SDS-PAGE on a 10% gel and
transferred onto a nitrocellulose membrane. Membranes were
incubated with blocking buffer (cat. no. P0023B; Beyotime Institute
of Biotechnology, Haimen, China) at room temperature for 2 h. The
membranes were incubated with primary antibodies against BMPR2
(cat. no. bs-4237R; 1:500), COL2A1 (cat. no. bs-11929R; 1:500),
SOX9 (cat. no. bs-10725R; 1:500), Aggrecan (cat. no. bs-1223R;
1:1,000), Smad1 (cat. no. bs-16376R; 1:500), phosphorylated
(p)Smad1 (cat. no. bs-3417R; 1:500), Smad5 (cat. no. bs-13890R;
1:1,000), pSmad5 (cat. no. bs-19918R; 1:500), Smad8 (sc-293413;
1:1,000), pSmad8 (cat. no. 9511L; 1:500) and GAPDH (cat. no.
bs-10900R; 1:1,000) at room temperature for 2 h, followed by
secondary horseradish peroxidase-conjugated antibodies (cat. no.
bs-0295G-HRP; 1:1,000) at room temperature for 2 h thereafter.
Smad8 antibody was purchased from Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA and pSmad8 antibody was purchased from Cell
Signaling Technology, Inc., Danvers, MA, USA; the other antibodies
were purchased from Beijing Biosynthesis Biotechnology Co., Ltd.,
(Beijing, China). GAPDH was used as an internal control. The
protein were visualized by enhanced chemiluminescence (Applygen
Technologies Inc., Beijing, China). Densitometric analysis was
performed using LabWorks 4.0 image acquisition and analysis
software (UVP, LLC, Upland, CA, USA).
Luciferase reporter assay
Prediction of miR-1307-3p targets was performed
using web-based bioinformatics tools including TargetScan
(http://www.targetscan.org/vert_72/),
and PicTar (www.pictar.org). The BMPR2 3′ UTR
harboring the miR-1307-3p target sequence and the seed sequence
mutated type (BMPR2 3′UTR mut) were synthesized by GenePharma
(Shanghai, China) and were then ligated following the luciferase
ORF into the pmirGLO vector (Promega Corporation, Madison, WI,
USA). The cells were seeded into 24-well plates at a confluence of
~80% the day before transfection, thus ensuring that the cells were
at a confluence of 80% on the day of transfection. miR-1307-3p
mimics were cotransfected with BMPR2 3′UTR or BMPR2 3′UTR mut
reporter. Transfection was performed using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc. Waltham, MA, USA). At 48 h post-transfection, the
cells were lysed according to the manufacturer’s protocol (Promega
Corporation), and the luciferase activities were determined using
the Dual-Glo luciferase reporter assay system (Promega
Corporation).
Statistical analysis
The data are expressed as the mean ± standard error
of the mean. Statistical analyses were performed with SPSS
statistical software version 16.0 (SPSS, Inc., Chicago, IL, USA)
using Student’s t-test and one-way analysis of variance with
Bonferroni post hoc analysis. P<0.05 was considered to indicate
a statistically significant difference.
Results
miR-1307-3p is downregulated during
chondrogenic differentiation
To determine the expression levels of miR-1307-3p
during the chondrogenic differentiation of hADSCs, hADSCs were
cultured in chondrogenic media using the pellet culture system for
21 days. First, the characterization of hADSCs was analyzed by flow
cytometry. The results showed that the cells were positive for
CD29, CD44 and CD49 and negative for CD45 (Fig. 1A). The chondrogenic
differentiation of hADSCs was confirmed by Alcian blue staining and
immunocytochemical examination of the chondrogenic marker, ColII.
Morphological observation showed the morphology of cells was
altered following induction and it consisted of polygon-shaped
cells compared with those in the control group (Fig. 1B). The Alcian blue staining showed
the deposition of cartilage matrix proteoglycans was promoted in
hADSCs subjected to chondrogenic induction (Fig. 1C). In addition, the level of Co1II
was increased in hADSCs following induction (Fig. 1D). These results confirmed the
chondrogenic differentiation of hADSCs exposed to the chondrogenic
induction medium. Subsequently, RT-qPCR analysis was used to
examine the expression of miR-1307-3p. As shown in Fig. 1E, the expression of miR-1307-3p
was significantly decreased following 21 days of induction, which
was consistent with previous microarray and northern blot
results.
miR-1307-3p inhibits chondrogenic
differentiation of hADSCs
To investigate the function of miR-1307-3p in the
chondrogenic differentiation of hADSCs, the expression of
miR-1307-3p was overexpressed or knocked down with a GFP-expressing
lentivirus system. The hADSCs were infected with recombinant
lentivirus expressing miR-1307-3p or miR-1307-3p inhibitor or
control and were then subjected to chondrogenic differentiation.
First, microscopic examination of GFP-positive cells was performed
to determine the infection efficiency of the recombinant
lentivirus. As shown in Fig. 2A,
the infection efficiency was high as shown by the ~80% GFP-positive
cells in all groups. Subsequently, an RT-qPCR assay was used to
determine the expression efficiency of the recombinant lentivirus.
Infection with the miR-1307-3p-expressing lentivirus resulted in a
significant increase of miR-1307-3p levels, whereas infection with
the miR-1307-3p inhibitor lentivirus led to a decrease in its
levels (Fig. 2B). The effect of
miR-1307-3p on chondrogenic differentiation of hADSCs was
determined by examining the deposition of cartilage matrix
proteoglycans and the levels of three cartilage-related markers
COL2A1, SOX9 and Aggrecan. Alcian blue staining showed the
deposition of cartilage matrix proteoglycans was suppressed when
miR-1307-3p was overexpressed but was promoted when miR-1307-3p was
knocked down (Fig, 2C). The
RT-qPCR results showed that the mRNA levels of COL2A1, SOX9 and
Aggrecan were markedly decreased following miR-1307-3p
overexpression, whereas they were increased in the
miR-1307-3p-knockdown group (Fig.
2D). The western blot assay showed that the proteins levels of
COL2A1, SOX9 and Aggrecan were significantly decreased in the
miR-1307-3p-overexpressing cells, but increased following
miR-1307-3p knockdown (Fig. 2E),
which suggested that miR-1307-3p suppressed chondrogenic
differentiation.
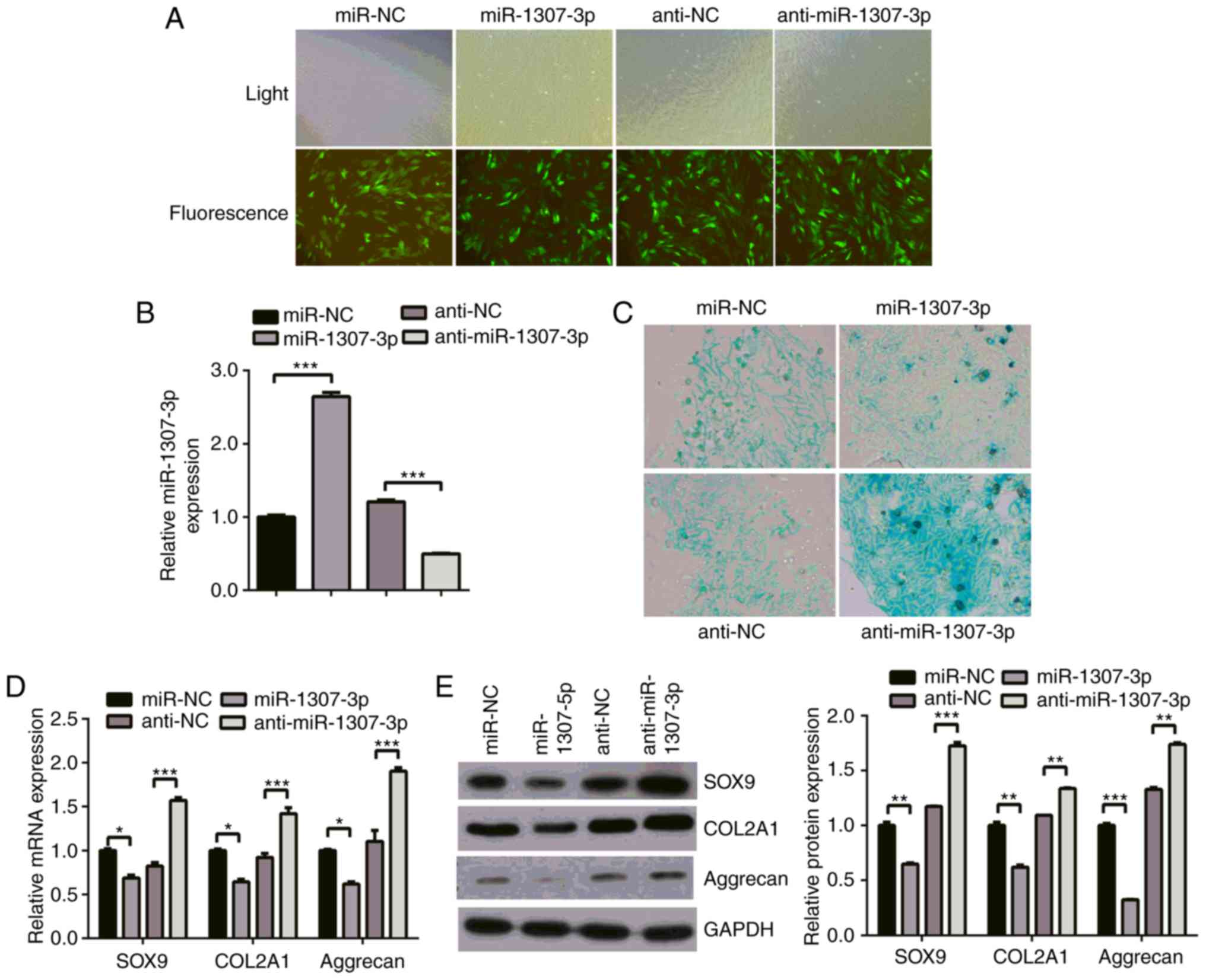 | Figure 2miR-1307-3p inhibits the chondrogenic
differentiation of hADSCs. hADSCs were infected with recombinant
lentivirus expressing miR-1307-3p or miR-1307-3p inhibitor or
control. Following infection, chondrogenic differentiation was
induced for 21 days. (A) Infection efficiency was determined by
microscopic examination of the GFP-expressing cells (magnification,
x10). (B) expression of miR-1307-3p was determined with reverse
transcription-quantitative polymerase chain reaction analysis. (C)
Deposition of cartilage matrix proteoglycans was determined by
Alcian blue staining (magnification, x40). (D) mRNA levels of
chondrogenic genes (SOX9, COL2A1 and aggrecan) were analyzed by
RT-qPCR analysis. (E) Protein levels of SOX9, COL2A1 and aggrecan
were detected with western blot analysis. *P<0.05,
**P<0.01 and ***P<0.001. miR, microRNA;
hADSCs, human adipose-derived stem cells; SOX9, sex determining
region Y-box 9; COL2A1, collagen type II α1 chain; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; NC, negative
control. |
BMPR2 is a direct target of
miR-1307-3p
To examine the underlying mechanism by which
miR-1307-3p suppresses chondrogenic differentiation, potential
target genes were searched for using TargetScan and PicTar. Among
the predicted targets, BMPR2 was identified, which was suggestive
of its involvement in chondrogenic differentiation. To confirm the
involvement of BMPR2 in the chondrogenic differentiation of hADSCs,
its expression during differentiation was exam-ined. The RT-qPCR
and western blot analyses showed that the mRNA and protein levels
of BMPR2 were increased following 21 days of induction (Fig. 3A and B), which was in accordance
with the downregulation of miR-1307-3p (Fig. 1D). Bioinformatics analysis showed
that there was one putative binding site in the 3′UTR of BMPR2 mRNA
(Fig. 3C). First, a dual
luciferase assay was used to determine whether miR-1307-3p directly
targeted BMPR2; as shown in Fig.
2D, the overexpression of miR-1307-3p significantly decreased
the luciferase activity of the BMPR2 3′UTR (~60%) but had no effect
on that of the BMPR2 3′UTR mut. Furthermore, the effect of
miR-1307-3p on the endogenous levels of BMPR2 was examined. The
RT-qPCR results showed that the mRNA level of BMPR2 was decreased
by ~40% following miR-1307-3p overexpression, whereas the knockdown
of miR-1307-3p led to the opposite results (Fig. 3E). The western blot analysis also
showed that the overexpression of miR-1307-3p significantly
decreased the protein levels of BMPR2, whereas miR-1307-3p
knockdown increased the protein levels of BMPR2 (Fig. 3F). These results indicated that
miR-1307-3p bound directly and negatively regulated the expression
of BMPR2.
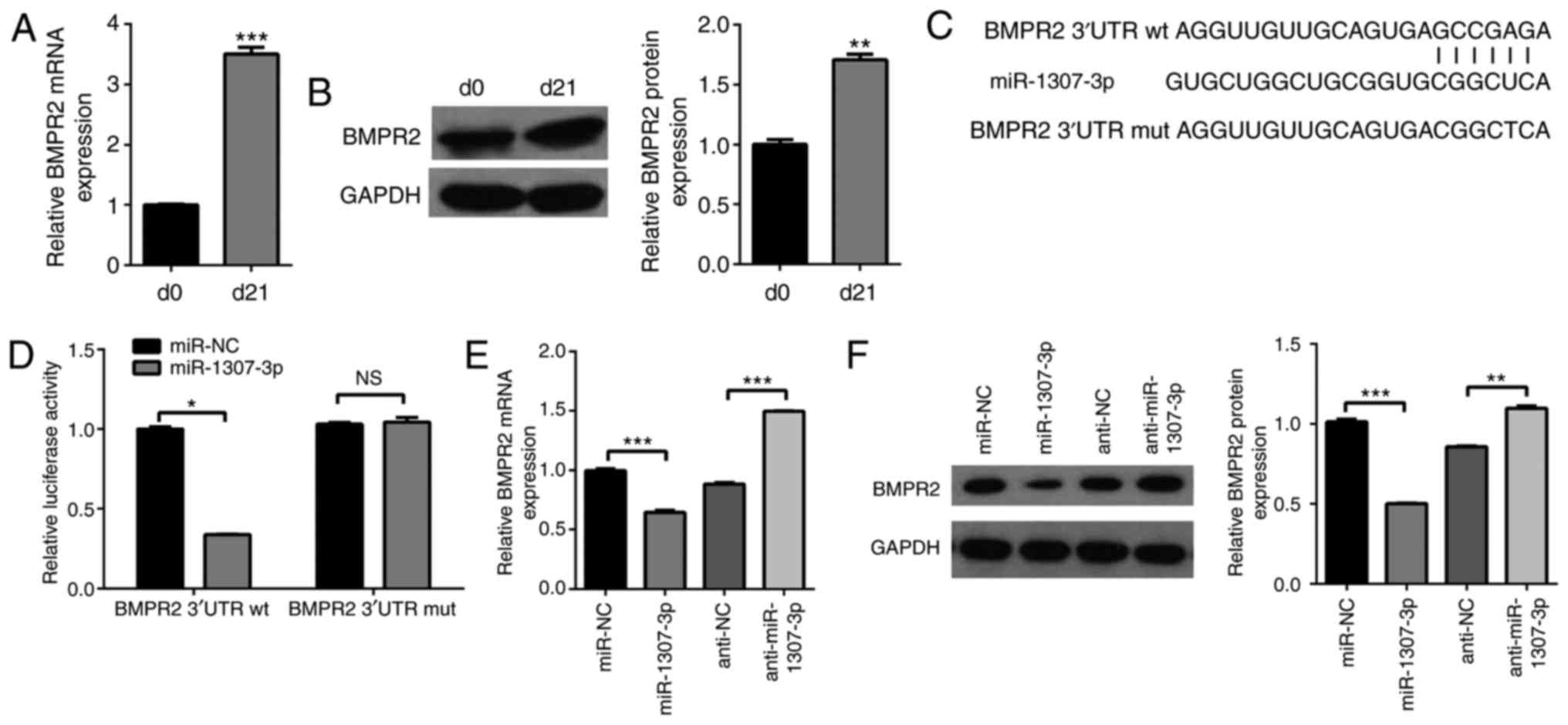 | Figure 3BMPR2 is a direct target of
miR-1307-3p. The expression of BMPR2 during chondrogenic
differentiation was examined by (A) RT-qPCR and (B) western blot
analyses. (C) Predicted binding site of miR-1307-3p in the 3′UTR of
BMPR2 is shown. (D) hADSCs were transfected with the indicated
plasmids, and the luciferase activities were determined using a
dual luciferase assay. hADSCs were infected with the indicated
recombinant lentivirus, and (E) mRNA and (F) protein levels of
BMPR2 were analyzed by RT-qPCR and western blot analyses,
respectively. *P<0.05, **P<0.01 and
***P<0.001. miR, microRNA; hADSCs, human
adipose-derived stem cells; BMPR2, bone morphogenetic protein
receptor type 2; 3′UTR, 3′ untranslated region; wt, wild-type; mut,
mutant; NC, negative control; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; NS, not
significant; d, day. |
BMPR2 mediates the suppression of
chondrogenic differentiation of hADSCs by miR-1307-3p
To determine whether BMPR2 mediated the effects of
miR-1307-3p on the chondrogenic differentiation of hADSCs, si-BMPR2
was transfected into hADSCs that were infected with the miR-1307-3p
inhibitor lentivirus. The RT-qPCR and western blot analyses showed
that the depletion of miR-1307-3p led to an increase in the mRNA
and protein levels of BMPR2, whereas the knockdown of BMPR2
attenuated the promoting effect of miR-1307 depletion on the
expression of BMPR2 (Fig. 4A and
B). Furthermore, the knockdown of BMPR2 prevented the promoting
effect of miR-1307 inhibition on the expression of cartilage
related markers COL2A1, SOX9 and Aggrecan at the mRNA and protein
levels (Fig. 4C and D). These
results demonstrated that BMPR2 was a functional target of
miR-1307-3p.
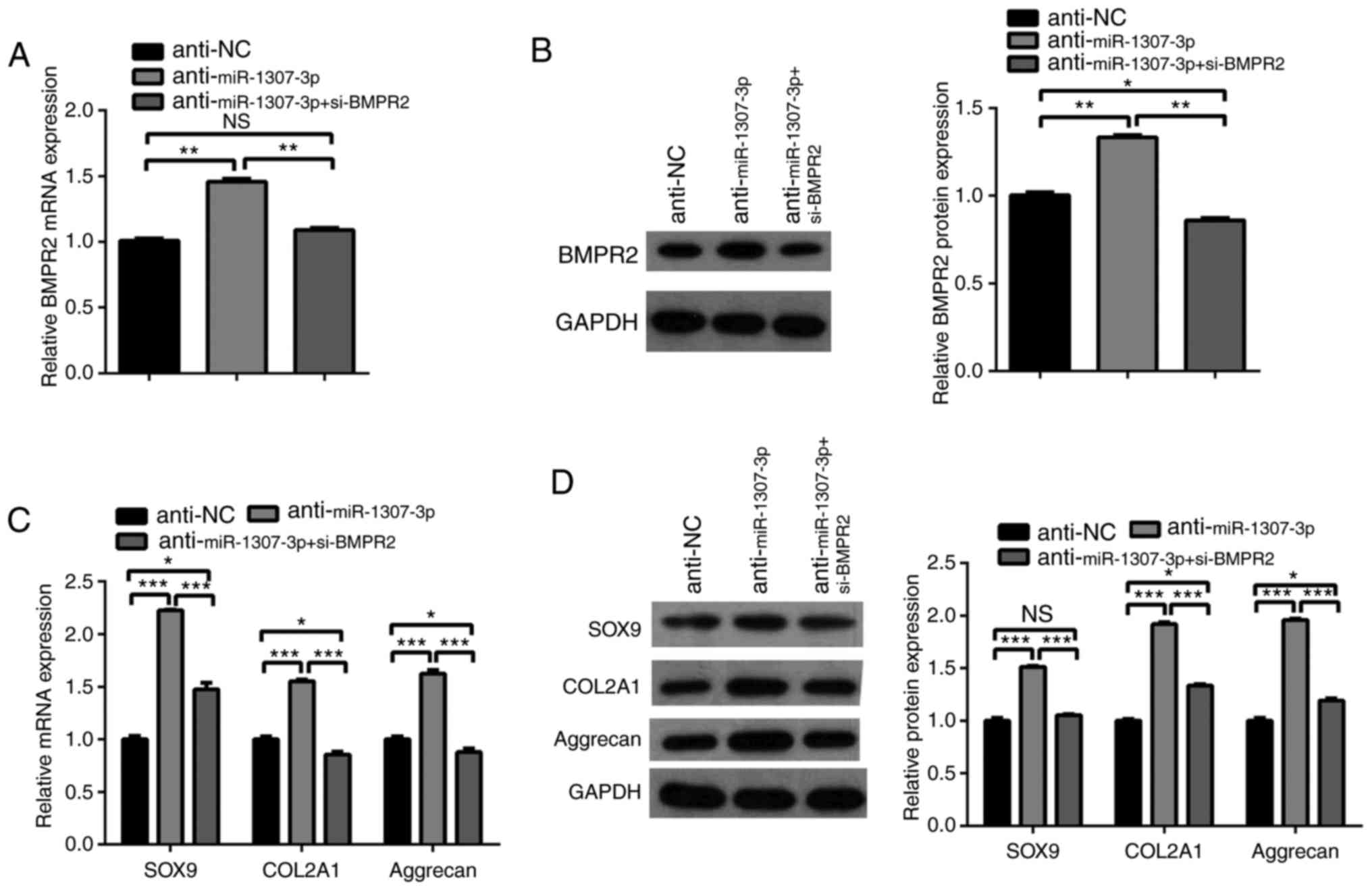 | Figure 4BMPR2 mediates the inhibitory effect
of miR-1307-3p on the chondrogenic differentiation of hADSCs.
hADSCs were infected or transfected with the indicated lentivirus
or siRNA. (A) mRNA and (B) protein levels of BMPR2 were determined
by RT-qPCR and western blot analyses, respectively. (C) mRNA and
(D) protein levels of SOX9, COL2A1 and Aggrecan were determined by
RT-qPCR and western blot analyses, respectively.
*P<0.05, **P<0.01 and
***P<0.001. miR, microRNA; hADSCs, human
adipose-derived stem cells; BMPR2, bone morphogenetic protein
receptor type 2; si-, small interfering RNA; SOX9, sex determining
region Y-box 9; COL2A1, collagen type II α1 chain; NC, negative
control; RT-qPCR, reverse transcription-quantitative polymerase
chain reaction; NS, not significant. |
miR-1307-3p inhibits chondrogenic
differentiation via suppression of BMPR2 downstream pathways
Considering the key role of BMPR2 in the BMPR2-Smad
signaling pathway, the present study investigated whether
miR-1307-3p inhibited the chondrogenic differentiation of hADSCs
through suppressing the BMPR2-Smad signaling pathway. The protein
levels of BMPR2 downstream molecules, including Smad1, 5, and 8,
were examined. Western blot analysis showed that there were no
significant changes in the total levels of Smad1, 5, and 8 when
miR-1307-3p was overexpressed or knocked down. However, the
phosphorylation of the three Smad proteins was markedly decreased
when miR-1307-3p was overexpressed, whereas their phosphorylation
was increased when miR-1307-3p was knocked down (Fig. 5A and B). Furthermore, BMPR2
knockdown prevented the promoting effect of miR-1307-3p inhibition
on the phosphorylation of the three Smad proteins (Fig. 5A and B). These results suggested
that miR-1307-3p suppressed chondrogenic differentiation via
inhibiting BMPR2 downstream pathways.
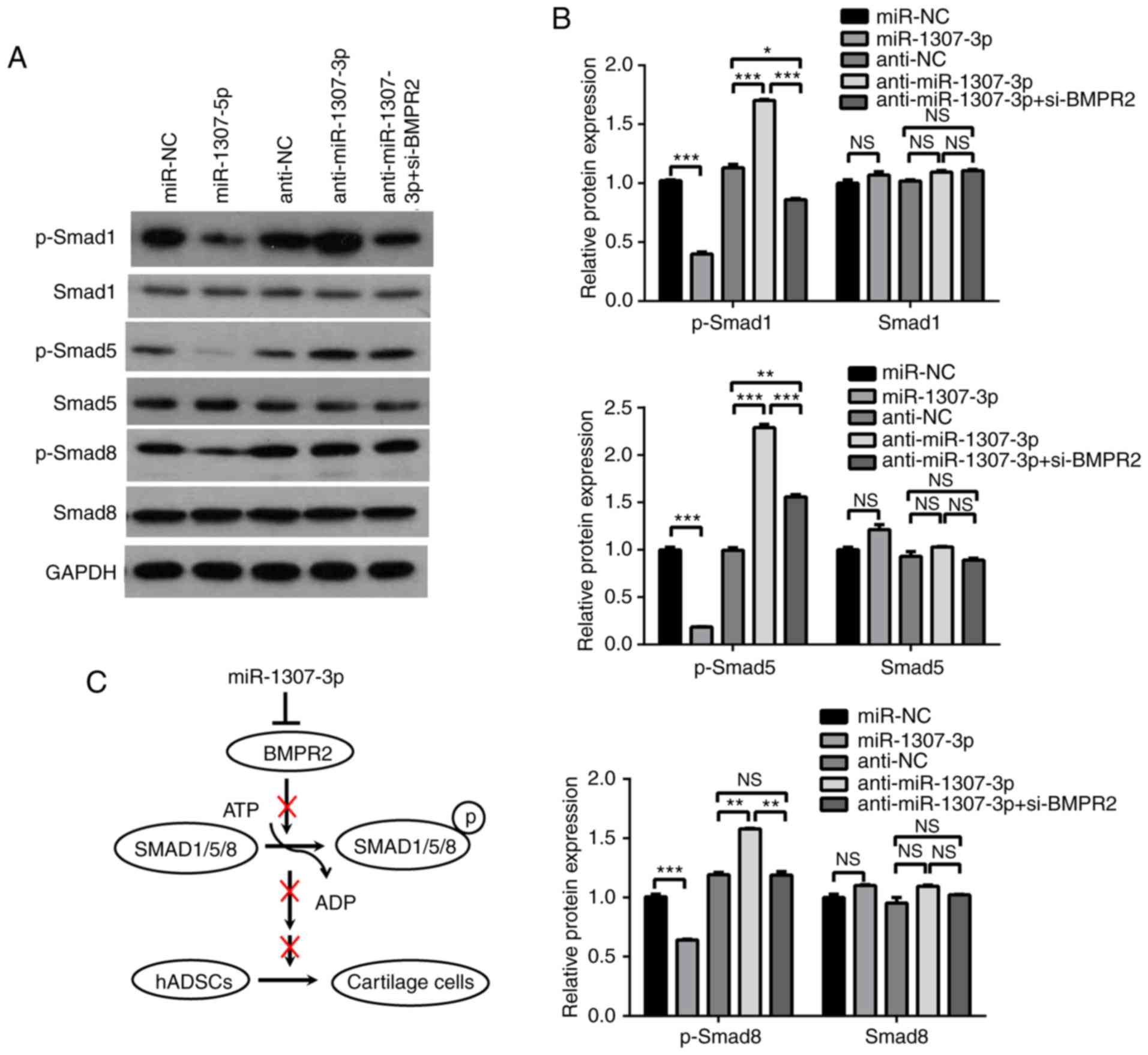 | Figure 5miR-1307-3p inhibits the BMPR2-SMAD
pathway. (A) hADSCs were infected with the indicated recombinant
lentivirus, and the protein levels of Smad1, 5, and 8, and the
phosphorylation of Smad1, 5, and 8 were analyzed by western blot
analysis. (B) Graphs show quantification of western blot results
relative to β-actin. (C) Proposed model for the suppression of
miR-1307-3p on chondrogenic differentiation. *P<0.05,
**P<0.01 and ***P<0.001. miR, microRNA;
hADSCs, human adipose-derived stem cells; BMPR2, bone morphogenetic
protein receptor type 2; si-, small interfering RNA; SMAD, mothers
against decapentaplegic; p-, phosphorylated; NC, negative cotrol;
NS, not significant. |
Discussion
Emerging data has shown that miRNAs may exert key
functions in the chondrogenic differentiation of hADSCs (9). According to the miRNA microarray and
northern blot analysis in our previous study, miR-1307-3p was
significantly downregulated during the chondrogenic differentiation
of hADSCs (13). In the present
study, its decreased expression was verified with RT-qPCR analysis.
Previous data show that it may be used as a valuable marker for the
diagnosis and treatment of certain diseases, however, the function
of miR-1307-3p has not been determined (19-22). Johnson et al (19) found that the combinations of five
miRNAs, including miR-1307-3p, in the saliva of children can
identify patients with prolonged symptoms on a logistic regression.
The polymorphism rs7911488 T>C in pre-miR-1307p was closely
associated with the efficacy of capecitabine chemotherapy in
patients with colon cancer (20).
García-Donas et al (21)
found that the expression of miR-1307-03p was increased in tumor
tissues from cases of metastatic renal cell carcinoma uniformly
treated with tyrosine kinase inhibitors by deep sequencing of miRNA
expression profiles. Its expression was significantly associated
with progression-free survival and overall survival rates, which
suggest it may be a useful biomarker for metastatic renal cell
carcinoma treatment. Shimomura et al (22) found that the level of miR-1307 was
increased in breast cancer, and its combination with the four other
miRNAs was a useful marker for the diagnosis of breast cancer. In
the present study, the results demonstrated that miR-1307-3p
suppressed the chondrogenic differentiation of hADSCs by targeting
BMPR2 and inhibited the BMPR2 downstream signaling pathway.
miRNAs function at post-transcriptional levels by
degrading target mRNAs or inhibiting the translation of target
mRNAs. To identify targets of miR-1307-3p, the present study used
two online software tools, TargetScan and PicTar, to predict its
targets and found that BMPR2 is a potential target of miR-1307-3p.
Its expression during the chondrogenic differentiation of hADSCs
was increased, which was negatively correlated with the decreased
levels of miR-1307-3p. A luciferase assay showed that BMPR2 was the
target of miR-1307-3p. RT-qPCR and western blot analyses showed
that miR-1307-3p suppressed the endogenous expression of BMPR2. The
rescue experiments indicated that the inhibitory function of
miR-1307-3p on the chondrogenic differentiation of hADSCs was
mediated by BMPR2. As miRNAs can often target more than one target,
whether other targets are involved in the effect of miR-1307-3p on
the chondrogenic differentiation of hADSCs requires further
investigation.
BMPR2, a member of the BMP receptor family of
transmembrane serine/threonine kinases, is involved in the
differentiation of mesenchymal progenitor cells (23). It promotes osteogenic or
chondrogenic differentiation, and can be targeted and negatively
regulated by miR-99a, miR-153, miR-490 and miR-100, respectively
(13,24-26). BMPR2 is a key molecule in BMP
signaling. On binding to BMP, BMPR2 is phosphorylated and activates
BMPR1, which in turn leads to the phosphorylation of intracellular
Smad1, 5, and 8. Subsequently, the common mediator Smad4 binds to
phosphorylated Smad1, 5, and 8 and is translated into the nucleus,
which can activate the transcription of BMPR target genes (27,28). In the present study, it was found
that miR-1307-3p suppressed the chondrogenic differentiation of
hADSCs by targeting and inhibiting the expression of BMPR2. To
further examine the specific mechanism, the expression of molecules
of the BMP downstream signaling pathway was assessed. It was found
that the overexpression of miR-1307-3p decreased the
phosphorylation of Smad1, 5, and 8, whereas the inhibition of
miR-1307-3p had the opposite effects. However, the knockdown of
BMPR2 attenuated the increased phosphorylation of Smad1, 5, and 8
by miR-1307-3p. These results indicated that the suppression of
chondrogenic differentiation by miR-1307-3p was due to the
inhibition of the BMPR2-Smad signaling pathway.
In conclusion, the results of the present study
demonstrated that miR-1307-3p suppressed the chondrogenic
differentiation of hADSCs by targeting BMPR2 and subsequently
inhibiting the BMPR2-Smad signaling pathway. These findings assist
in understanding the molecular mechanisms underlying chondrogenic
differentiation, which may provide novel therapeutic approaches for
hADSC-based cartilage tissue engineering.
Funding
This study was supported by the Youth Fund of
Guizhou Provincial People’s Hospital [grant no. GZSYQN(2015)06],
the National Natural Science Foundation of China (grant nos.
31660265, 81060145 and 81560356), the Subsidy Foundation of
National Natural Science Foundation of Guizhou Provincial People’s
Hospital [Guizhou Science and Technology Platform (grant no.
(2017)5724], and the Science and Technology Foundation of Guizhou
Province [Guizhou Science and Technology J Word (2015)2096].
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors’ contributions
BL and XBT were involved in the study design and
revision of the manuscript. ZY and RL wrote the manuscript. ZY, RL,
JA, QDW and YZ performed the experiments. ZY, LC, JW, BC and WP
analysed and interpreted the data. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Tianjin Haihe Hospital (Tianjin, China). Written informed consent
was obtained from the donors.
Patient consent for publication
Written informed consent was obtained from the
donors.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
BMPR2
|
bone morphogenetic protein receptor
type 2
|
|
BMSCs
|
bone marrow mesenchymal stem cells
|
|
COL2A1
|
collagen type II α1 chain
|
|
hADSCs
|
human adipose-derived stem cells
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
miRNAs
|
microRNAs
|
|
SOX9
|
sex determining region Y-box 9
|
|
TGF
|
transforming growth factor
|
|
3′-UTR
|
3′ untranslated region
|
References
|
1
|
Ge Z, Hu Y, Heng BC, Yang Z, Ouyang H, Lee
EH and Cao T: Osteoarthritis and therapy. Arthritis Rheum.
55:493–500. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang J and Chen J: Bone tissue
regeneration-Application of mesenchymal stem cells and cellular and
molecular mechanisms. Curr Stem Cell Res Ther. 12:357–364. 2017.
View Article : Google Scholar
|
|
3
|
Vallée M, Côté JF and Fradette J:
Adipose-tissue engineering: Taking advantage of the properties of
human adipose-derived stem/stromal cells. Pathol Biol (Paris).
57:309–317. 2009. View Article : Google Scholar
|
|
4
|
Wang S, Qu X and Zhao RC: Mesenchymal stem
cells hold promise for regenerative medicine. Front Med. 5:372–378.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zuk PA, Zhu M, Ashjian P, De Ugarte DA,
Huang JI, Mizuno H, Alfonso ZC, Fraser JK, Benhaim P and Hedrick
MH: Human adipose tissue is a source of multipotent stem cells. Mol
Biol Cell. 13:4279–4295. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gimble J and Guilak F: Adipose-derived
adult stem cells: Isolation, characterization, and differentiation
potential. Cytotherapy. 5:362–369. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dong H, Lei J, Ding L, Wen Y, Ju H and
Zhang X: MicroRNA: Function, detection, and bioanalysis. Chem Rev.
113:6207–6233. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu C, Tian B, Qu X, Liu F, Tang T, Qin A,
Zhu Z and Dai K: MicroRNAs play a role in chondrogenesis and
osteoarthritis (Review). Int J Mol Med Jul. 34:13–23. 2014.
View Article : Google Scholar
|
|
10
|
Lee S, Yoon DS, Paik S, Lee KM, Jang Y and
Lee JW: microRNA-495 inhibits chondrogenic differentiation in human
mesenchymal stem cells by targeting Sox9. Stem Cells Dev.
23:1798–1808. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Y, Huang X and Yuan Y: MicroRNA-410
promotes chondrogenic differentiation of human bone marrow
mesenchymal stem cells through downregulating Wnt3a. Am J Transl
Res. 9:136–145. 2017.
|
|
12
|
Zhang Z, Kang Y, Zhang Z, Zhang H, Duan X,
Liu J, Li X and Liao W: Expression of microRNAs during
chondrogenesis of human adipose-derived stem cells. Osteoarthritis
Cartilage. 20:1638–1646. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang Z, Hao J and Hu ZM: MicroRNA
expression profiles in human adipose-derived stem cells during
chondrogenic differentiation. Int J Mol Med. 35:579–586. 2015.
View Article : Google Scholar :
|
|
14
|
Hou C, Yang Z, Kang Y, Zhang Z, Fu M, He
A, Zhang Z and Liao W: MiR-193b regulates early chondrogenesis by
inhibiting the TGF-beta2 signaling pathway. FEBS Lett.
589:1040–1047. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hou C, Zhang Z, Zhang Z, Wu P, Zhao X, Fu
M, Sheng P, Kang Y and Liao W: Presence and function of
microRNA-92a in chondrogenic ATDC5 and adipose-derived mesenchymal
stem cells. Mol Med Rep. 12:4877–4886. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu J, Kang Y, Liao WM and Yu L: MiR-194
regulates chondrogenic differentiation of human adipose-derived
stem cells by targeting Sox5. PLoS One. 7:e318612012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu SC, Hsiao HF, Ho ML, Hung YL, Chang JK,
Wang GJ and Wang CZ: Suppression of discoidin domain receptor 1
expression enhances the chondrogenesis of adipose-derived stem
cells. Am J Physiol Cell Physiol. 308:C685–C896. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
19
|
Johnson JJ, Loeffert AC, Stokes J, Olympia
RP, Bramley H and Hicks SD: Association of salivary microRNA
changes with prolonged concussion symptoms. JAMA Pediatr.
172:65–73. 2018. View Article : Google Scholar
|
|
20
|
Chen Q, Mao Y, Meng F, Wang L, Zhang H,
Wang W and Hua D: Rs7911488 modified the efficacy of
capecitabine-based therapy in colon cancer through altering
miR-1307p and TYMS expression. Oncotarget. 8:74312–74319.
2017.PubMed/NCBI
|
|
21
|
García-Donas J, Beuselinck B,
Inglada-Pérez L, Graña O, Schöffski P, Wozniak A, Bechter O,
Apellániz-Ruiz M, Leandro-García LJ, Esteban E, et al: Deep
sequencing reveals microRNAs predictive of antiangiogenic drug
response. JCI Insight. 1:e860512016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shimomura A, Shiino S, Kawauchi J,
Takizawa S, Sakamoto H, Matsuzaki J, Ono M, Takeshita F, Niida S,
Shimizu C, et al: Novel combination of serum microRNA for detecting
breast cancer in the early stage. Cancer Sci. 107:326–334. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kang Q, Song WX, Luo Q, Tang N, Luo J, Luo
X, Chen J, Bi Y, He BC, Park JK, et al: A comprehensive analysis of
the dual roles of BMPs in regulating adipogenic andosteogenic
differentiation of mesenchymal progenitor cells. Stem Cells Dev.
18:545–559. 2009. View Article : Google Scholar
|
|
24
|
Zhou X, Wang J, Sun H, Qi Y, Xu W, Luo D,
Jin X, Li C, Chen W, Lin Z, et al: MicroRNA-99a regulates early
chondrogenic differentiation of rat mesenchymal stem cells by
targeting the BMPR2 gene. Cell Tissue Res. 366:143–53. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cao Y, LV Q and LV C: MicroRNA-153
suppresses the osteogenic differentiation of human mesenchymal stem
cells by targeting bone morphogenetic protein receptor type II. Int
J Mol Med. 36:760–766. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zeng Y, Qu X, Li H, Huang S, Wang S, Xu Q,
Lin R, Han Q, Li J and Zhao RC: MicroRNA-100 regulates osteogenic
differentiation of human adipose-derived mesenchymal stem cells by
targeting BMPR2. FEBS Lett. 586:2375–2381. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu F, Ventura F, Doody J and Massagué J:
Human type II receptor for bone morphogenic proteins (BMPs):
Extension of the two-kinase receptor model to the BMPs. Mol Cell
Bio. 15:3479–3486. 1995. View Article : Google Scholar
|
|
28
|
Miyazono K, Maeda S and Imamura T: BMP
receptor signaling: transcriptional targets, regulation of signals,
and signaling cross-talk. Cytokine Growth Factor Rev. 16:251–263.
2005. View Article : Google Scholar : PubMed/NCBI
|