Introduction
Atherosclerosis (AS) is the primary cause of various
cardiovascular and cerebrovascular diseases and has high morbidity
and mortality rates (1). The
dysfunction of endothelial cells (ECs), the infiltration of
inflammatory cells and the formation of vascular smooth muscle
cells and foam cells are considered to be the key mechanisms
involved in the development of AS; however, ECs dysfunction is also
hypothesized to serve a pivotal role in the initiation of AS
(2). In addition, oxidative
stress and mitochondrial dysfunction are responsible for the
development of endothelial dysfunction (3,4).
Oxidative stress leads to a disruption of the capabilities of
intracellular antioxidants, resulting in the presence of a large
number of reactive oxygen species (ROS), thereby causing lipid
peroxidation and biological macromolecular degeneration, which are
considered to be the primary factors leading to vascular
endothelial dysfunction (5,6).
In addition, H2O2 is a critical type of ROS
that has good stability and diffusibility (5,6).
Therefore, in the present study, H2O2 was
used to simulate oxidative stress to construct an EC dysfunction
model.
Sphingomyelin (SM) is a type of sphingolipid, which
are key modulators of a number of physiological and
pathophysiological processes that include the cell cycle,
apoptosis, angiogenesis, and stress and inflammatory responses
(7,8). Sphingomyelin synthase (SMS) is
involved in the biosynthesis of SM. SMS has two isoforms,
sphingomyelin synthase 1 (SMS1) and sphingomyelin synthase 2
(SMS2), which are the key enzymes in the last step of a series of
enzymatic processes involved in SM synthesis (9). In previous years, a number of
studies have demonstrated that SM is involved in AS occurrence and
development (10-12). For example, epidemiological
studies and clinical trial data identified that serum SM content
and AS were positively correlated (11,12). Animal and cellular experiments
have also indicated that SMS may cause AS by inhibiting reverse
cholesterol transport (RCT), leading to lipid deposition (13-15). However, these studies were focused
on the effects of SM on the RCT monocyte-macrophage infiltration
and lipid deposition (13–15).
Although Zhou et al (16)
hypothesized that lovastatin may decrease the levels of
sphingomyelin increased by 27-hydroxycholesterol on cultured
vascular ECs and may attenuate cellular calcification, the role of
SM in endothelial dysfunction remains unclear.
The canonical Wnt/β-catenin signaling pathway has
been established as being involved in a number of cellular
processes, including cell proliferation, differentiation and tissue
patterning, and is also associated with multiple pathological
processes, including metabolic disorders and cancer (17,18). However, several recent studies
have suggested that the Wnt/β-catenin signaling pathway may cause
AS (19,20). Animal experiments and human
endarterectomy samples have demonstrated that the expression of
protein Wnt-5a is increased in AS and is associated with the
severity of atherosclerotic lesions (21,22). In addition, ECs dysfunction is
also associated with the Wnt/β-catenin pathway. For example,
activation of the Wnt/β-catenin pathway may cause endothelial
dysfunction and augment monocyte adhesion to ECs (23,24). Ma et al (25) also suggested that the
Wnt/β-catenin pathway may be activated in the process of ECs
dysfunction induced by oxidized low-density lipoprotein (ox-LDL).
Concomitantly, the Wnt/β-catenin pathway is hypothesized to induce
vascular smooth muscle cell proliferation and migration, which is
one of the hallmarks of the development of atherosclerotic plaques
(26,27).
As aforementioned, the SM and Wnt/β-catenin pathways
are involved in AS. However, the correlation between SM and
Wnt/β-catenin in the development of AS induced by oxidative stress
remains unclear. Therefore, the present study focused on
identifying their association in the process of
H2O2-induced ECs dysfunction. Firstly, an
SMS2 overexpression and SMS-silenced human umbilical vein
endothelial cells (HUVECs) model was constructed. This model was
subsequently treated with H2O2, and the
dysfunction of HUVECs, the expression of SMS2 and the activation of
the Wnt/β-catenin pathway were examined.
Materials and methods
Cell culture
HUVECs were purchased from the Cell Bank of Type
Culture Collection of the Chinese Academy of Sciences (Shanghai,
China) and cultured in Dulbecco’s modified Eagle’s medium (DMEM;
cat.no. 12100-500; Beijing Solarbio Bioscience & Technology
Co., Ltd., Beijing, China) containing penicillin and streptomycin
(100 U/ml and 0.1 mg/ml, respectively) and 10% certified fetal
bovine serum (FBS; Biological Industries Israel Beit Haemek,
KibbutzBeit Haemek, Israel) at 37°C containing 5% CO2.
In addition, THP-1 cells (Cell Bank of Type Culture Collection of
the Chinese Academy of Sciences, Shanghai, China) were grown in
RPMI-1640 (cat. no. 31800; Beijing Solarbio Bioscience &
Technology Co., Ltd.) containing 10% FBS and incubated at 37°C in a
humidified atmosphere containing 5% CO2.
Cell viability assay
Cell proliferation was detected by using an MTT
assay. The untreated HUVECs were cultured in a 96-well plate
(Beaver Nano-Technologies Co., Ltd., Suzhou, China) at a density of
5×104/well for 24 h, and then were treated with
different concentrations of H2O2 (0, 200,
400, 600, 800 and 1,000 µmol/l) for 24 h. Then, 40 µl
MTT (5 mg/ml; cat. no. M8180; Beijing Solarbio Bioscience &
Technology Co., Ltd.) was added to each well and the plates were
incubated for 4 h at 37°C. After 4 h, the medium was removed, and
150 µl dimethyl sulfoxide (cat. no. D8370; Beijing Solarbio
Bioscience & Technology Co., Ltd.) was added to the wells,
which were then agitated on a shaking table for 10 min. Finally,
the absorbance was measured at a wavelength of 490 nm. A total of 6
repeat wells were measured at each time point, and the experiment
was performed in triplicate.
Transfection and grouping
To establish the SMS2 overexpression cell model, a
SMS2-overexpression plasmid [pcDNA3.1(+)], provided by Dr Tingbo
Ding (School of Pharmacy, Fudan University, Shanghai, China) was
used to transfect the HUVECs. The transfection protocol was
performed as described by Luo et al (28). In brief, cells were seeded into
6-well plates (Beaver Nano-Technologies Co., Ltd., Suzhou, China)
containing antibiotic-free RPMI-1640 medium (Biological Industries
Israel Beit Haemek Ltd.). At the time of transfection, the cells
were grown to 60–70% confluency as described previously (27). The plasmid SMS2 (4 µg; SMS2
group) or a control empty plasmid (4 µg; Control group) was
diluted with 50 µl DMEM (FBS-free and antibiotic-free
medium) or 5 µl Entranster™-D-4000 (Engreen Biosystem Ltd.,
Auckland, New Zealand) was also diluted with 50 µl DMEM.
After 5 min, the dilutions were mixed together gently and incubated
at 37°C for 20 min, and the mixture and 2 ml DMEM (10% FBS) were
subsequently added to each well. After 6 h, the medium was replaced
by fresh DMEM (containing 10% FBS and antibiotics) as described
previously (14). The control and
SMS2 group were divided into two groups after 24 h, resulting in
four groups; the control, SMS2, CH
(Control+H2O2) and SH
(SMS2+H2O2) groups, and both the CH and SH
groups were treated with H2O2 (500
µmol/l). After 24 h, to collect all of these cells, HUVECs
were centrifuged at 1,520 × g for 5 min (room temperature) as
appropriate according to the needs of the experiments.
Measurement of lactate dehydrogenase
(LDH) level in medium, intracellular superoxide dismutase (SOD)
activity and malondialdehyde (MDA) content
HUVECs were cultured at a density of
1×105/well in 6-well plates and incubated overnight at
37°C. Following transfection and treatment with
H2O2 (500 µmol/l) for 24 h, the
supernatant were collected, HUVECs were centrifuged (1,520 × g, 5
min, room temperature) and harvested following being digested by
trypsin according to the protocol of the manufacturer. The levels
of LDH (cat. no. A020-1; Nanjing Jiancheng Bioengineering
Institute, Nanjing, China), intracellular MDA content (cat. no.
A003-1; Nanjing Jiancheng Bioengineering Institute) and SOD
activity (cat. no. A001-1-1; Nanjing Jiancheng Bioengineering
Institute) levels were measured using a microplate reader (Thermo
Fisher Scientific, Inc., Waltham, MA, USA). LDH was measured at the
wavelength of 450 nm, however, SOD and MDA were measured at the
wavelength of 560 nm.
Cell adhesion assay
Following transfection of the HUVECs and treatment
with H2O2 (500 µmol/l), THP-1 cells at
a density of 5×103/well were added and incubated for 30
min at 37°C. Next, the medium was discarded, and the cells were
washed gently with PBS twice to remove non-adherent THP-1 cells. As
described previously (29), the
adherent THP-1 cells were counted in a single field under a phase
contrast inverted microscope (Magnification, ×20; Olympus IX71;
Olympus Corporation, Tokyo, Japan). Experiments were repeated three
times to obtain the average number of adhesive monocytes/well.
Flow cytometry analysis
HUVECs were collected by trypsin digestion without
EDTA (trypsin 1:250 from porcine pancreas; cat. no., 9002-07-7;
Sangon Biotech Co., Ltd., Shanghai, China) digestion and washed
twice with PBS. Thereafter, V-fluorescein isothiocyanate (FITC) and
propidium iodide (PI) were added into the HUVECs according to the
protocol of the ANNEXIN V-FITC/PI apoptosis assay kits (cat. no.,
17210J; Beijing Zoman Biotechnology Co., Ltd., Beijing, China). The
apoptosis results were analyzed by flow cytometry (BD Biosciences,
Franklin Lakes, NJ, USA), and FlowJo Software (v.10; Tree Star,
Inc., Ashland, OR, USA) was used to analyze the data as previously
described (28,30).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
After HUVECs were treated by different
concentrations of H2O2 for 24 h, the RNA was
isolated using TRIzol® reagent (Ambion; Thermo Fisher
Scientific, Inc.) according to the manufacturer’s instructions.
Total RNA (2 µg) was used for RT transcription in a total
volume of 20 µl (RevertAid First Strand cDNA; cat. no.
K1622; Thermo Fisher Scientific, Inc.). The primers for β-actin and
SMS2 were synthesized by Shanghai Sangon Biotech Co., Ltd.,
(Shanghai, China) and their sequences were as follows: β-actin
forward, 5′-GATCATTGCTCCTCCTGAGC-3′; β-actin reverse,
5′-ACTCCTGCTTGCTGATCCAC-3′; SMS2 forward,
5′-ATAGCCCTCAGTCATGATT-3′; and SMS2 reverse
5′-CAGCACATGACAACGGTTCA-3′ (30).
Gene expression was measured using qPCR with SYBR-Green (cat. no.
PR820A; Takara Biotechnology Co., Ltd., Dalian, China). The samples
were examined using a StepOnePlus™ Real-Time PCR System, with the
following thermocycler conditions: 95°C for 3 min, then 40 cycles
of 95°C for 10 sec and 60°C for 30 sec. Fluorescence data and
melting curves were subsequently obtained. Experimental data are
presented as the mean ± standard deviation of three biological
replicates. The gene expression was normalized to the expression of
β-actin. The quantification cycle (Cq) value was defined as the
number of cycles required for the fluorescent signal to reach the
threshold. Using the comparative Cq method, the relative expression
levels were calculated using the formula for 2−ΔΔCq
(31).
Western blot analysis
Following treatment with 500 µmol/l
H2O2, only HUVECs that were not used for
measuring the cell adhesion ability to THP-1 were used for western
blot analysis. Total proteins of all groups were extracted by
radioimmunoprecipitation assay buffer (cat. no. ROO20; Beijing
Solarbio Bioscience & Technology Co., Ltd.), and protein
content was measured using a BCA assay kit (cat. no. CW0014;
Beijing Kangwei Century Biotechnology Co., Ltd., Beijing, China).
Equal amounts of protein (~60 µg) were separated by
electrophoresis on 10–12% SDS-PAGE gels and then transferred onto
polyvinylidene fluoride membranes (Immobilon-P; EMD Millipore,
Billerica, MA, USA). Equal transfer was examined by staining with
Ponceau red (cat. no. CW0057S; Beijing Kangwei Century
Biotechnology Co., Ltd.). The membranes were blocked with 10%
skimmed milk or 10% BSA in TBS for 1 h at room temperature and
incubated with primary antibodies in TBS, which included 0.05%
Tween 20, 2% bovine serum albumin (cat. no. A8020; Beijing Solarbio
Science & Technology Co., Ltd.) and 0.05% sodium azide
overnight at 4°C (28,32). The antibodies were used to detect
the expression levels of SMS2 [cat. no AP9740b; dilution, 1:1,000;
Abgent Biotech (Suzhou) Co., Ltd., Suzhou, China],
apoptosis-associated proteins B-cell lymphoma 2 (Bcl-2; cat. no.
BS-0032R; dilution, 1:500; BIOSS, Beijing, China), Bcl-2-associated
X protein (Bax; cat. no. 60267-1-Ig; dilution, 1:2,000; ProteinTech
Group, Inc., Chicago, IL, USA) and adhesion-associated proteins
intracellular adhesion molecule-1 (ICAM-1; cat. no. BS-0608R;
dilution, 1:1,000; BIOSS), vascular cell adhesion molecule-1
(VCAM-1; cat. no. BS-0920R; dilution, 1:500; BIOSS), monocyte
chemoat-tractant protein-1 (MCP-1; cat. no. BS-1101R; dilution,
1:500; BIOSS), and the Wnt/β-catenin signal pathway-associated
proteins β-catenin (cat. no. 66379-1-Ig; dilution, 1:2,000;
ProteinTech Group, Inc.) and phosphorylated β-catenin (cat. no.
DF2989; dilution, 1:1,000; Affinity Biosciences, Cincinnati, OH,
USA). GAPDH (cat. no. HRP-60004; dilution, 1:8,000; ProteinTech
Group, Inc.) was used as the internal control protein to ensure the
equal loading of the proteins in each lane. Following washing of
the membranes three times, horseradish peroxidase-conjugated
anti-rabbit (cat. no. BA1054; dilution, 1:8,000; Boster Biological
Technology, Inc., Wuhan, China) and anti-mouse (cat. no. SA0000I-I;
dilution, 1:8,000; ProteinTech Group, Inc.) antibodies were used as
the secondary antibodies at 1:8,000 in 10% skimmed milk in TBS.
Finally, subsequent to washing the membranes three times, the blots
were visualized using an enhanced chemiluminescence reagent (cat.
no. CW0049M; Beijing Kangwei Century Biotech Co., Ltd.) and an
autoradiography system (Chemiluminescence Imaging System; version
5.1; Bio-Rad Laboratories, Inc., Hercules, CA, USA). Each assay was
repeated three times. The results were analyzed by Image Lab
(version 5.1; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Sphingomyelin synthase enzyme activity
assay
HUVECs were treated with H2O2
for 24 h as aforementioned. HUVECs were treated with different
concentrations of Dy105 (provided by Dr Deyong Ye, School of
Pharmacy, Fudan University; 0, 15, 20 and 25 µmol/l)
(33) for 24 h. NBD-ceramide (0.1
µg/µl, cat. no. 62527; Cayman chemical company, Ann
Arbor, MI, USA) was incubated with treated cells at 37°C to
synthesize sphingomyelin in vitro. After 3 h of incubation,
the cells were harvested by 1,520 × g for 5 min, and the medium was
also collected at room temperature. According to the protein
content in each group, protein levels were adjusted to the volume
of the reaction system (700 µl) to ensure the consistency of
the total protein and medium added. Lipids were extracted in
chloroform: Methanol (2:1), dried under N2 gas, and
separated by thin layer chromatography using chloroform: MeOH:
NH4OH (14:6:1) at room temperature for 10 min. The
chromatography film was scanned after 10 min with an
auto-radiography system (Chemiluminescence Imaging System, Clinx
Science Instruments Co., Ltd., Shanghai, China), and the intensity
of each band was measured using Image-Pro Plus version 6.0 software
(Media Cybernetics, Inc., Rockville, MD, USA) (15).
Treatment with SMS inhibitor and LiCl on
HUVECs
The cells were cultured as aforementioned. The
HUVECs were seeded at a density of 1×106/well into a
6-well plate prior to the addition of 20 µmol/l Dy105 or
lithium chloride (LiCl, 40 µmol/; Shanghai Qingfang Chemical
Technology Co., Ltd., Shanghai, China) treatment for 24 h. Next,
the Dy105 and the LiCl groups were treated with
H2O2 for 24 h, resulting in the following
five groups: Control, CH (Control+H2O2), DH
(Dy105+H2O2), LH
(LiCl+H2O2) and DLH
(Dy105+LiCl+H2O2). The medium and cells were
harvested and analyzed by LDH, MDA and SOD kits and western blot
analysis as aforementioned.
Statistical analysis
Statistical analysis was performed with statistical
software SPSS 22.0 (IBM Corp., Armonk, NY, USA), and the
significant differences between groups were evaluated using an
independent samples (un-paired) t-test. For multiple comparisons, a
one-way analysis of variance with Tukey’s post-hoc analysis was
used. All experiments were repeated at least three times, and
P<0.05 was considered to indicate a statistically significant
difference.
Results
H2O2 inhibits
HUVECs viability in a dose-dependent manner
To investigate whether SMS2 was associated with the
oxidative stress-induced dysfunction, the HUVECs were treated with
H2O2 for 24 h. Firstly, the
results (Fig. 1A) indicated that
H2O2 inhibited HUVECs proliferation, and the
proliferation rate of HUVECs gradually decreased to 25.49% when the
H2O2 dose was increased from 0 to 1,000
µmol/l. Concurrently, the LDH leakage rate into the media of
the HUVECs was also markedly increased in response to
H2O2 (Fig.
1B), which suggested that H2O2 inhibited
the viability of the HUVECs in a dose-dependent manner.
Furthermore, the present study measured the levels of Bax and Bcl-2
by western blot analysis, and as demonstrated in Fig. 1C, the protein expression of Bax
was increased, while the expression of Bcl-2 was decreased.
Notably, as indicated in Fig. 1C,
it was also observed that the protein expression level of SMS2 was
increased concomitant with the dose of H2O2
(P<0.05; n=3), and similar results were also identified for the
mRNA levels (Fig. 1D; P<0.05,
n=3). The expression of SMS1 was also measured, and no significant
differences in its expression levels were observed (data not
shown). Therefore, SMS2 was selected as the target gene. These data
suggest that SMS2 may be involved in
H2O2-induced cellular dysfunction.
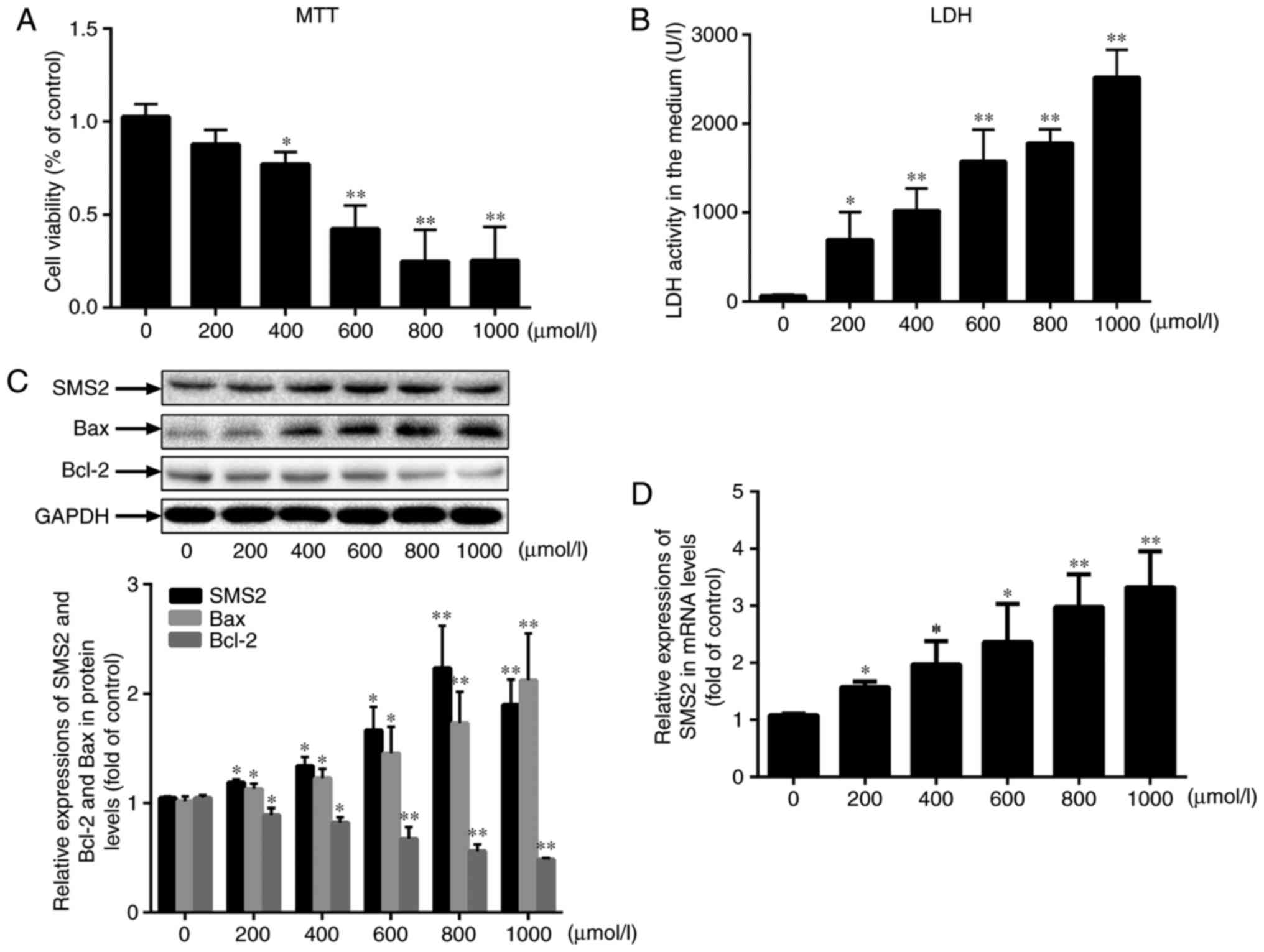 | Figure 1H2O2 inhibits
HUVECs viability in a dose-dependent manner. HUVECs were treated
with H2O2 at different doses (0, 200, 400,
600, 800 and 1,000 µmol/l) for 24 h. (A) Cell viability was
measured by an MTT assay. (B) The activity of LDH in the cellular
supernatant was also analyzed. (C) The protein levels of Bax, Bcl-2
and SMS2 were measured by western blot analysis. (D) The mRNA
expression of SMS2 was measured by reverse transcription
quantitative polymerase chain reaction. Values are presented as
mean ± standard deviation (n=3). *P<0.05 and
**P<0.001 vs. control group. SMS2, sphingomyelin
synthase 2; Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X
protein; LDH, lactate dehydrogenase; HUVECs, human umbilical vein
endothelial cells. |
SMS2 overexpression inhibits
proliferation and increases apoptosis of HUVECs
To additionally analyze the role of SMS2 in the
process of HUVECs dysfunction, an SMS2 overexpression cell model
was constructed. Fig. 2A
indicated that the expression of SMS2 was significantly upregulated
by ~130.7, 231.4 and 320.6% in the SMS2, CH and SH groups,
respectively, compared with the control group (P<0.05; n=3), and
the expression of SMS2 in the SH group was increased compared with
the CH group (P<0.05; n=3). These results demonstrated the
successful establishment of a SMS2 overexpression cell model, and
again, verified that the oxidative stress-induced dysfunction may
upregulate SMS2 in HUVECs. In addition, the results indicated that
the expression of the pro-apoptotic protein Bax was markedly
increased (Fig. 2B; P<0.05;
n=3), while the level of apoptosis inhibitory protein Bcl-2 was
decreased significantly (Fig. 2B;
P<0.05; n=3) in the SMS2, CH and SH groups compared with the
control group; additionally, the expression of Bcl-2 in the SH
group was decreased compared with the CH group (P<0.05;
n=3).
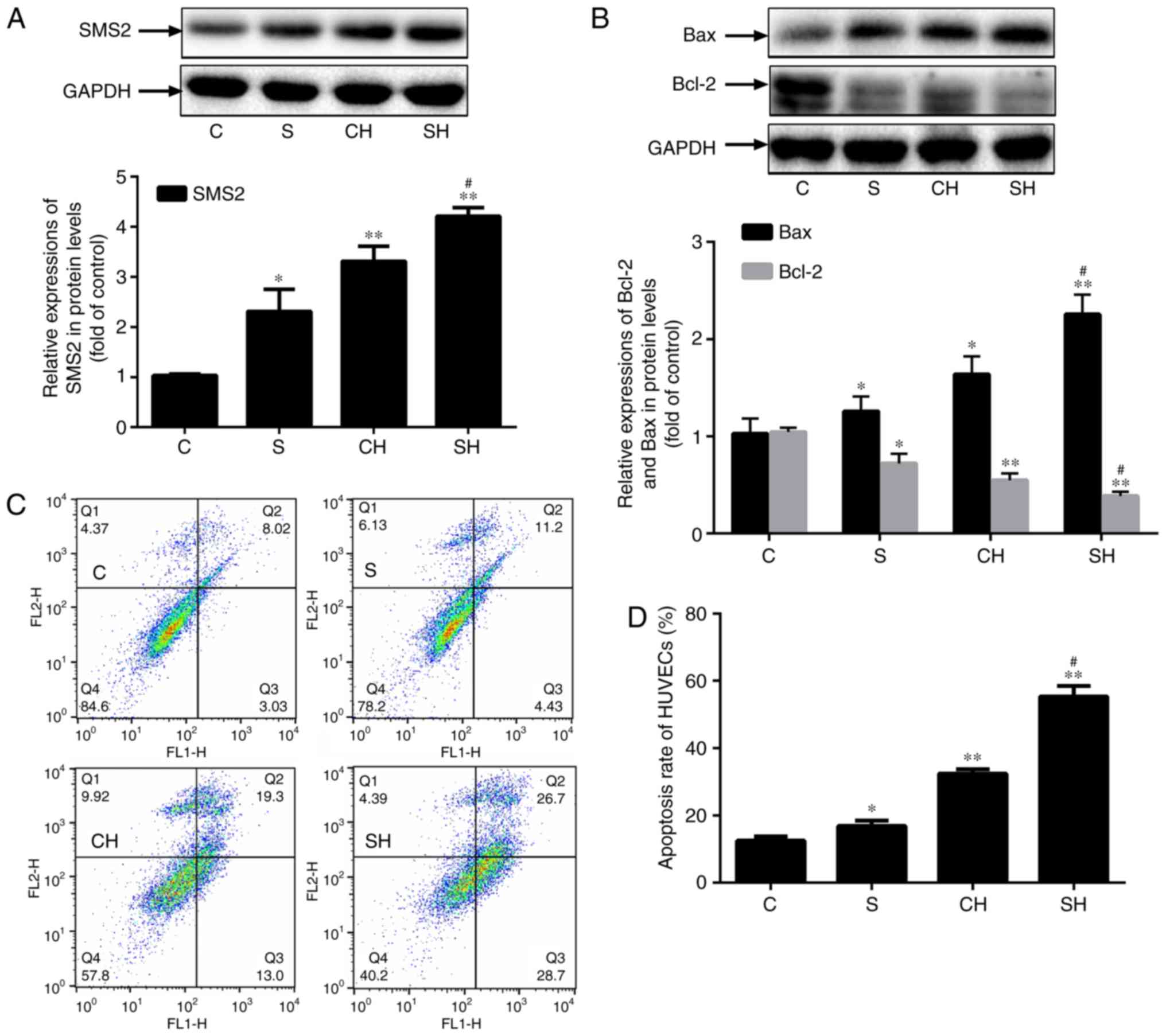 | Figure 2Effect of SMS2 on the proliferation
and apoptosis in HUVECs. Effect of SMS2 on the proliferation and
apoptosis in HUVECs. Cells were transfected with SMS2
overexpression plasmids and treated with or without
H2O2 (500 µmol/l) for 24 h. (A) The
protein levels of (A) SMS2, and (B) Bax and Bcl-2 were analyzed by
western blot analysis. (C) The apoptosis of the C, S, CH and SH
groups were analyzed by flow cytometry. (D) The quantified
apoptosis rate of HUVECs. Values are presented as mean ± standard
deviation (n=3). *P<0.05 and **P<0.001
vs. control group; #P<0.05 and
##P<0.001 vs. CH group. Bcl-2, B-cell lymphoma 2;
Bax, Bcl-2-associated X protein; SMS2, sphingomyelin synthase 2;
HUVECs, human umbilical vein endothelial cells; C, control group;
S, SMS2 overexpression group; CH, control group treated with
H2O2 for 24 h; SH, S group treated with
H2O2 for 24 h. |
The overexpression of SMS2 may inhibit the
proliferation of HUVECs and induce apoptosis. Therefore, to
determine the apoptosis rate of HUVECs, the present study analyzed
the apoptosis levels of the HUVECs by flow cytometry. As
demonstrated by Fig. 2C and D, it
was observed that H2O2 led to significantly
increased levels of HUVECs apoptosis compared with the control
group (P<0.001; n=3), and the apoptosis rate was 16.86, 32.43
and 55.40% in the SMS2, CH and SH groups, respectively. The results
also suggested that the apoptosis rates of the SH group were
increased compared with the CH group (P<0.001; n=3), and the
rate in the SH group was increased by ~6.11% compared with the
total increased rate in the CH and SMS2 groups (Fig. 2C and D). Collectively, these data
indicate that H2O2 may induce HUVECs
apoptosis and that SMS2 may increase the level of apoptosis induced
by oxidative stress in HUVECs, and that it may serve an important
role in endothelial dysfunction.
SMS2 overexpression increases the LDH
content of medium, and intracellular MDA content and SOD
activity
H2O2 is an important type of
ROS that may lead to ECs dysfunction through oxidative stress, and
we hypothesized that SMS2 may also aggravate the development of
oxidative stress. As it is well-known, the content of MDA is a
marker that reflects the level of antioxidants in cells; the
content of LDH is considered to be an index for measuring cell
viability and membrane integrity, and SOD activity is hypothesized
to prevent cell dysfunction by eliminating oxygen free radicals
(34). Therefore, the activity of
LDH, and the intracellular MDA and SOD activity levels were
detected. The results indicated that MDA and LDH production in
HUVECs following exposure to H2O2 or SMS2
overexpression was significantly increased compared with the
control group (P<0.05; n=3; Fig.
3A and B), and the levels in the SH group were increased
compared with the CH group (P<0.05; n=3). Conversely, the level
of SOD activity was decreased (Fig.
3C; P<0.05; n=3). Treatment with H2O2
or SMS2 may augment the dysfunction of HUVECs induced by oxidative
stress.
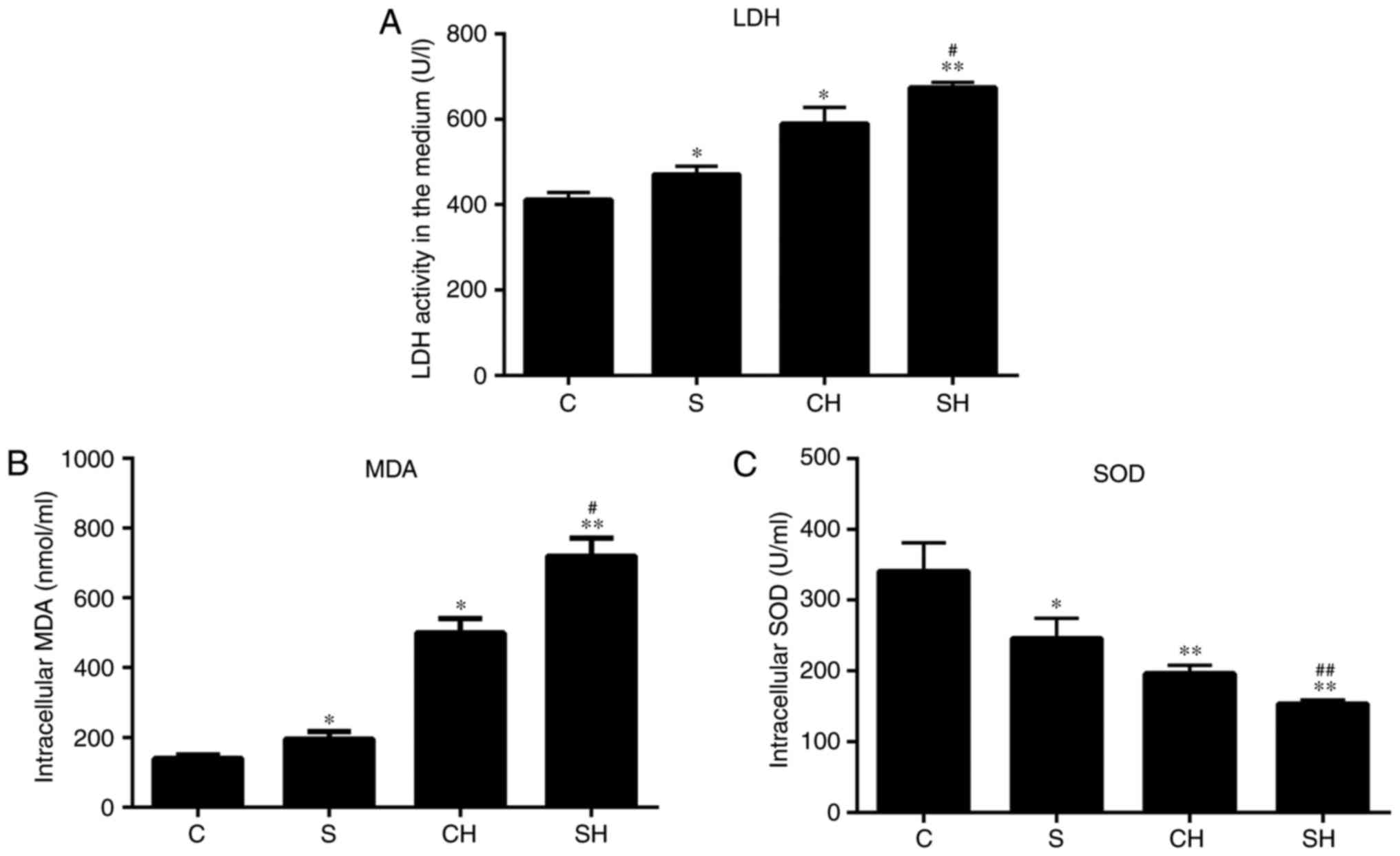 | Figure 3Overexpression of SMS2 increases the
LDH content of cell media, and affects intracellular MDA content
and SOD activity levels. Cells and supernatant were collected, and
the (A) LDH, (B) MDA and (C) SOD levels were measured with assay
kits. Values are presented as mean ± standard deviation (n=3).
*P<0.05 and **P<0.001 vs. control
group; #P<0.05 and ##P<0.001 vs. CH
group. LDH, lactate dehydrogenase; MDA, malondialdehyde; SOD,
superoxide dismutase; SMS2, sphingomyelin synthase 2; C, control
group; S, SMS2 overexpression group; CH, control group treated with
H2O2 for 24 h; SH, S group treated with
H2O2 for 24 h. |
SMS2 overexpression increases monocyte
adhesion in HUVECs
To identify whether SMS2 may affect the adhesion
rate of HUVECs, the present study measured the adhesion ability of
HUVECs to THP-1 cells. As indicated in Fig. 4A, the adhesion ability of the
SMS2, CH and SH groups were significantly increased compared with
the control group (Fig. 4A;
P<0.05; n=3), and the adhesion ability was improved by 29.94,
441.24 and 586.56% in the SMS2, CH and SH groups, respectively.
Therefore, SMS2 may promote the adhesion ability of HUVECs. In
addition, MCP-1, VCAM-1 and ICAM-1 are markers of HUVECs adhesion
ability (35). Compared with the
control group, the SMS2, CH and SH groups exhibited significantly
upregulated levels of the adhesion molecules MCP-1, VCAM-1 and
ICAM-1 (Fig. 4B; P<0.05; n=3).
In addition, the increased levels of these proteins in the SH group
was increased by ~75.99, 22.85 and 54.2% compared with the CH
group. These results suggested that the overexpression of SMS2 may
increase MCP-1, VCAM-1 and ICAM-1 expression and adhesion ability
in the HUVECs dysfunction model induced by
H2O2.
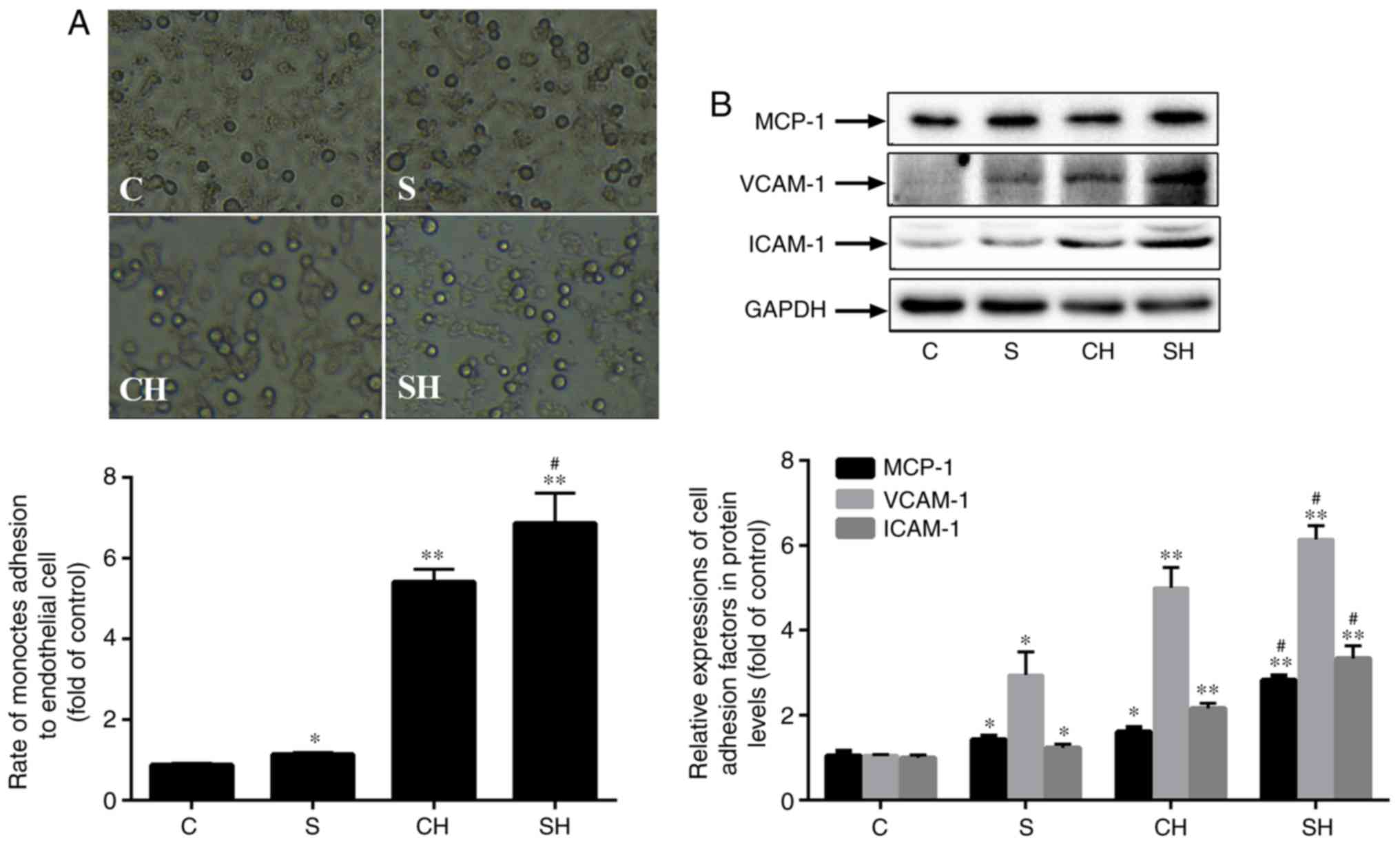 | Figure 4Overexpression of SMS2 increased
monocyte adhesion in HUVECs. The overexpression of SMS2 enhances
monocyte adhesion in HUVECs. (A) The adhesion rate of HUVECs to
THP-1 cells. (B) The protein expression of MCP-1, VCAM-1 and ICAM-1
were detected by western blot analysis. Values are presented as
mean ± standard deviation (n=3). *P<0.05 and
**P<0.001 vs. control group; #P<0.05
vs. CH group. Magnification, ×20; HUVECs, human umbilical vein
endothelial cells; VCAM-1, vascular cell adhesion molecule 1;
ICAM-1, intracellular adhesion molecule 1; MCP-1, monocyte
chemotactic protein 1; SMS2, sphingomyelin synthase 2; C, control
group; S, SMS2 overexpression group; CH, control group treated with
H2O2 for 24 h; SH, S group treated with
H2O2 for 24 h. |
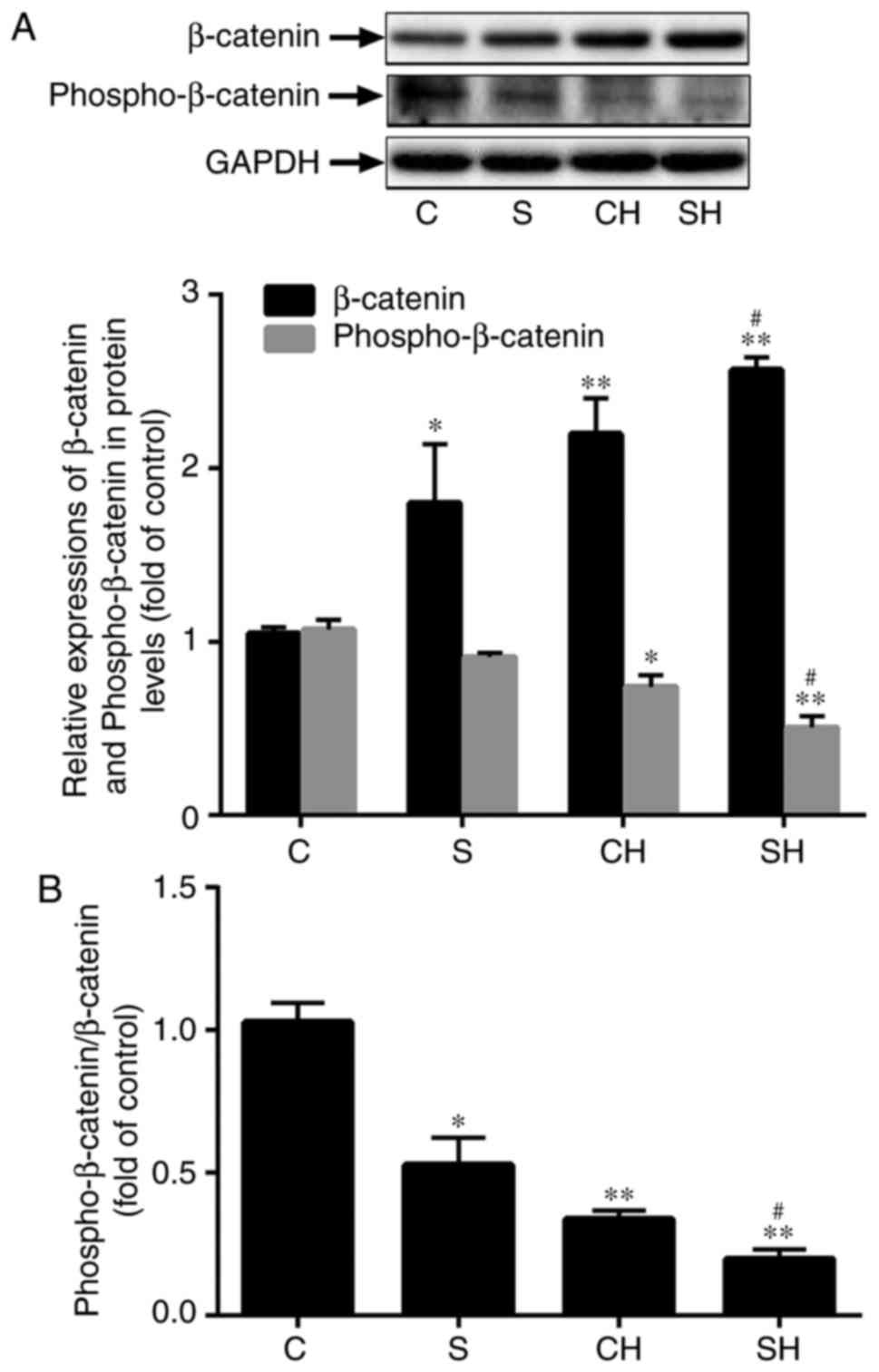
SMS2 overexpression activates the
Wnt/β-catenin signaling pathway
The Wnt/β-catenin signaling pathway regulates a
number of biological and pathological processes. However, the
association between the Wnt/β-catenin signaling pathway and EC
dysfunction remains unclear. To determine whether SMS2
overexpression may increase H2O2-induced
HUVECs dysfunction by the Wnt/β-catenin signaling pathway, the
present study analyzed the expression of β-catenin by western blot
analysis. Fig. 5A indicated that
the protein expression of β-catenin was significantly upregulated
in the CH, SMS2 and SH groups compared with the control group
(Fig. 5A; P<0.05; n=3); in
addition, the SH group exhibited higher expression levels compared
with the CH group (Fig. 5A;
P<0.001; n=3). The levels of phosphorylated β-catenin were also
investigated, and the results suggested that the relative content
of phosphorylated β-catenin in the SMS2, CH and SH groups was
decreased compared with the control group (Fig. 5A; P<0.05; n=3). Additionally,
it was also observed that its expression level in the SH group was
decreased compared with the CH group (Fig. 5A; P<0.05; n=3). As indicated in
Fig. 5B, the ratio of
phosphorylated-β-catenin/ β-catenin was investigated, and the
results demonstrated that this ratio was decreased in the SMS2, CH
and SH groups compared with the control group (P<0.05; n=3). The
ratio in the SH group was decreased compared with the CH group
(Fig. 5B; P<0.05; n=3). All of
these results indicated that H2O2-induced and
SMS2 overexpression may increase the expression of β-catenin and
prevent β-catenin phosphorylation.
Inhibition of SMS2 activity may decrease
EC dysfunction and inactivate the Wnt/β-catenin signaling
pathway
To confirm that SMS2 may regulate the Wnt/β-catenin
signaling pathway, the present study simultaneously inhibited SMS
activity and activated the Wnt/β-catenin signaling pathway by using
an SMS inhibitor (Dy105) and LiCl, respectively. Firstly, HUVECs
were treated with different concentrations of Dy105 (0, 15, 20 and
25 µmol/l) for 24 h. Then, the expression levels of SMS2 and
β-catenin were analyzed by western blot analysis; Fig. 6A indicated that the levels of SMS2
and β-catenin were most significantly decreased by 20 µmol/l
Dy105 treatment (P<0.001; n=3). Therefore, the treatment
concentration of Dy105 selected for subsequent experiments was 20
µmol/l. From the data presented in Fig. 6B, it was identified that the SMS
enzyme activity was markedly decreased compared with the control
group; it was decreased by 50.09% compared with control group
(P<0.001; n=3). Subsequently, the levels of LDH, MDA and SOD
intracellularly and in the culture media were measured, and it was
demonstrated that the inhibition of SMS enzyme activity by Dy105
decreased the levels of LDH and MDA, and increased the level of SOD
activity (Fig. 6C-E; P<0.05;
n=3). However, LiCl reversed these changes (Fig, 6C-E; P<0.001; n=3). These
results suggested that the inhibition of SMS enzyme activity may
attenuate EC dysfunction. In addition, Fig. 6F indicated that the protein
expression of β-catenin was significantly upregulated in the CH,
DH, LH and DLH group compared with the control group (P<0.05;
n=3), and the content of β-catenin in the LH and DLH groups was
increased by 122.03 and 103.05%, respectively, compared with the DH
group (Fig. 6F; P<0.05; n=3).
In particular, the content of β-catenin was markedly increased in
the LH group compared with the DLH group. Concomitantly, the levels
of phosphorylated β-catenin were also investigated, and the results
were the inverse of those for β-catenin levels (Fig. 6F; P<0.05; n=3). The ratio of
phosphorylated-β-catenin/β-catenin (Fig. 6G) was also investigated; the
results indicated that the changes in this ratio were coincident
with Fig. 6F. These results
indicated that Dy105 may decrease the expression of β-catenin to
inactivate the Wnt/β-catenin signaling pathway; conversely, LiCl
may re-activate it.
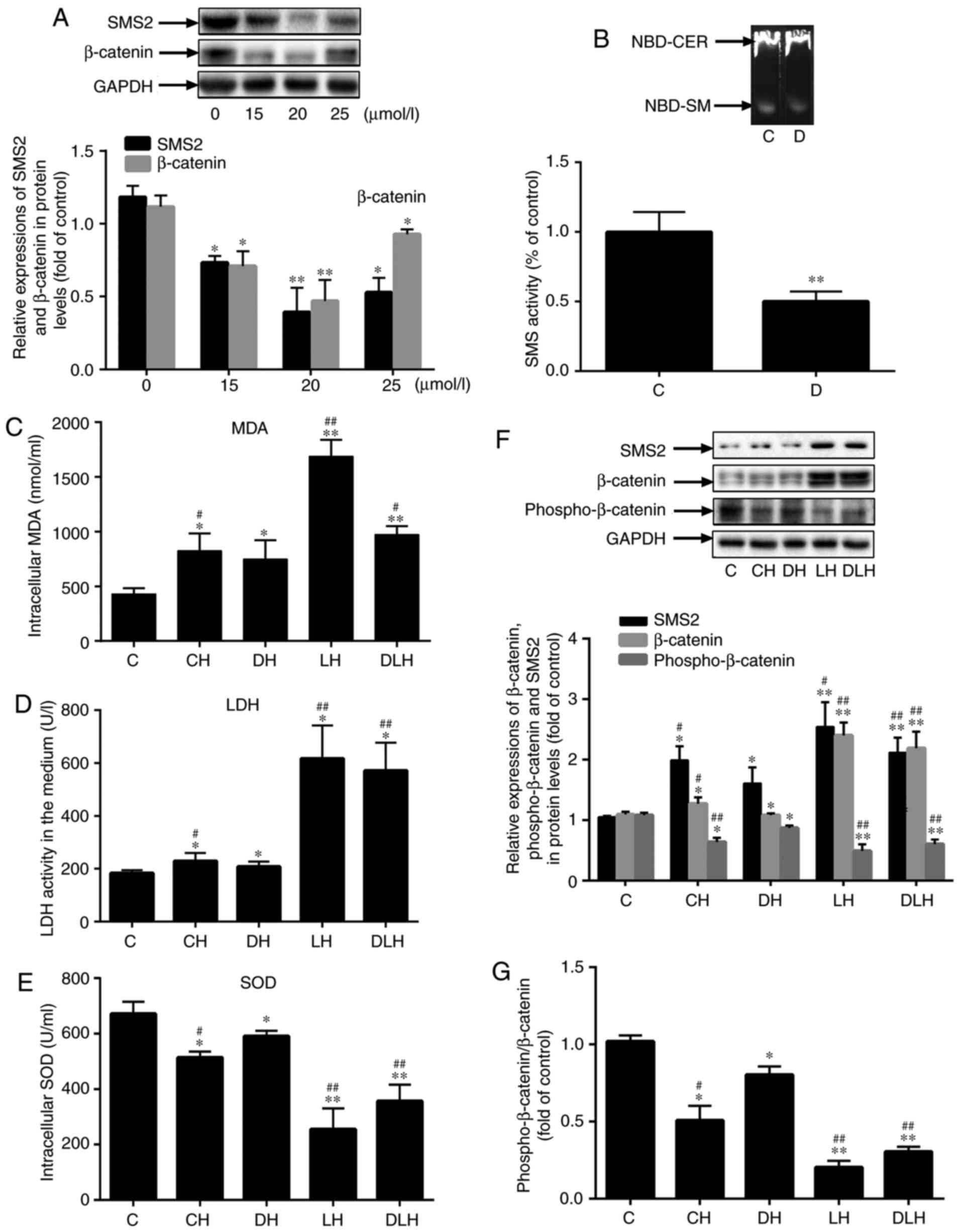 | Figure 6Inhibition of the SMS activity may
decrease endothelial cells dysfunction and inactivate the
Wnt/β-catenin signaling pathway. (A) The expression of SMS2 and
β-catenin was measured by western blot analysis in human umbilical
vein endothelial cells treated with Dy105 at different dosages. (B)
SMS activity was measured by thin-layer chromatography. Values are
presented as the mean ± standard deviation (n=3). (C) The level of
MDA. (D) The activity of LDH. (E) The levels of SOD. (F) The
protein expression of SMS2, β-catenin and phosphorylation of
β-catenin were determined by western blot analysis. (G)
Phospho-β-catenin/β-catenin ratio. Values are presented as mean ±
standard deviation (n=3). *P<0.05 and
**P<0.001 vs. control group; #P<0.05
and ##P<0.001 vs. DH group. SMS2, sphingomyelin
synthase 2; LDH, lactate dehydrogenase; MDA, malondialdehyde; SOD,
superoxide dismutase; D, Dy105 treatment, NBD-CER, NBD-ceramide,
NBD-SM, NBD-sphingomyelin; C, control group, CH, control group
treated by H2O2 for 24 h; DH,
Dy105+H2O2 group, LH,
LiCl+H2O2 group, DLH,
Dy105+LiCl+H2O2 group. |
Discussion
The results of the present study indicated that
H2O2 may upregulate the levels of SMS2 in a
dose-dependent manner, and that the oxidative stress-induced
dysfunction of HUVECs was associated with the activation of
β-catenin. In addition, SMS2 may increase the expression of
β-catenin and attenuate the phosphorylation of β-catenin, which may
augment the oxidative stress-induced dysfunction of HUVECs induced
by H2O2. Conversely, when the SMS activity
was inhibited by Dy105, the Wnt/β-catenin signaling pathway was
suppressed and ECs dysfunction was also attenuated; however, LiCl
reversed these changes.
It has been established that HUVECs serve a pivotal
role in AS occurrence and development (2). However, oxidative stress is one of
the major factors causing HUVECs dysfunction when antioxidant
mechanisms are overwhelmed by ROS (5,6).
H2O2 is the primary source of endogenous ROS
and has been extensively used to establish in vitro models
(5,36,37). In the present study, different
doses of H2O2 were used to treat HUVECs to
induce dysfunction, and it was identified that the expression of
SMS2 also increased with increasing H2O2
concentrations, which suggested that SMS2 expression is closely
associated with HUVECs dysfunction. Subsequently, additional data
from the present study validated that SMS2 was involved in the
oxidative stress-induced dysfunction of HUVECs, as its expression
was induced by H2O2, and its overexpression
augmented HUVECs dysfunction. ECs dysfunction and the upregulation
of MCP-1, VCAM-1 and ICAM-1 are critical events in AS (2,36).
The results from the present study also indicated that SMS2
overexpression increased the expression of MCP-1, VCAM-1 and
ICAM-1, and the adhesion ability of THP-1 cells, which may assist
in recruiting macrophages into the intima and contribute to the
initiation of AS (38,39). All of these results suggested that
SMS2 is involved in AS occurrence and development via promoting
HUVECs dysfunction and adhesion ability to THP-1 cells. Although
previous studies have verified that SMS contributes to AS
occurrence and development through the inhibition of RCT and may
lead to lipid deposition (13–15,40,33), to the best of our knowledge, the
present study is the first to suggest that SMS2 may promote HUVEC
dysfunction and adhesion ability to THP-1 cells, and assist in AS
occurrence and development.
It has been previously established that the
canonical Wnt/β-catenin signaling pathway is critical in a number
of cellular processes (17,18). In these cellular processes, when
the Wnt/β-catenin signaling pathways are activated, cytoplasmic
β-catenin, the central molecule of the Wnt signaling pathway, is
not able to be phosphorylated and is degraded by the action of the
destruction complex (axis inhibition protein 1, APC, WNT signaling
pathway regulator, casein kinase 1 alpha 1 and glycogen synthase
kinase 3β). This effect results in the accumulation of cytoplasmic
β-catenin and translocation into the nucleus, where it promotes the
expression of a number of key genes involved in different cellular
functions via activating the transcription regulation activity of
T-cell factors (17). However,
the results from the present study and the data from several other
studies have suggested that the Wnt/β-catenin signal pathway is
associated with the cell dysfunction and apoptosis induced by
H2O2 (41,42). For example, in cardiomyocytes,
H2O2 may upregulate dishevelled-1 (Dvl-1),
β-catenin and Myc proto-oncogene protein, promote β-catenin nuclear
activity, activate Wnt/β-catenin signaling and finally facilitate
DNA damage and tumor protein 53-mediated apoptosis (41,42), whereas when secreted frizzled
related protein 1 suppressed the expression of Dvl-1 and β-catenin
and the activity of the Wnt/frizzled pathway, the
H2O2-induced apoptosis of cardiomyocytes was
decreased (43). However, in ECs,
oxidative stress is regarded as a critical pathogenic factor for
dysfunction, and the accumulation of ROS may result in EC apoptosis
(25,44). Therefore, ox-LDL may serve a
critical role in the progression and development of AS via
increasing oxidative stress (45). Additional studies have
demonstrated that in this process, the Wnt/β-catenin signaling
pathway is activated; this observation resulted in the
identification of certain genes with expression patterns associated
with HUVECs apoptosis (25,44). In the present study, when HUVECs
were treated with H2O2, the expression of
β-catenin was increased, and the phosphorylation of β-catenin was
decreased. It is clear that the Wnt/β-catenin signaling pathway is
involved in the H2O2-induced dysfunction of
HUVECs. In addition, the present study identified that the
overexpression of SMS2 improved the dysfunction of HUVECs and their
adhesion ability to THP-1 cells. The possible mechanism may be that
SMS2 overexpression results in the accumulation of cytoplasmic
β-catenin and translocation to the nucleus through attenuating
phosphorylation of β-catenin, while the inhibition of SMS activity
may increase the phosphorylation of β-catenin and decreased the
nuclear translocation of β-catenin.
Although SMS has two isozymes, no significant
difference was observed in the expression levels of SMS1, but SMS2
expression was altered following treatment with
H2O2 in the present study. Therefore, the
overexpression of SMS2 may promote the dysfunction of HUVECs
through activating the Wnt/β-catenin signal pathway. In addition,
it has been established that SMS2 is a key enzyme involved in the
biosynthesis of SM, and that it is also involved in the production
of diacylglycerol (DAG) (46).
Yang et al (47)
hypothesized that DAG may activate β-catenin, where osteoclasts
were stimulated by colony-stimulating factor 1. Therefore, we
hypothesize that the overexpression of SMS2 increases the content
of DAG, which may activate β-catenin and aggravate the dysfunction
of HUVECs. However, the detailed mechanism by which SMS2 affects
the Wnt/β-catenin signaling pathway requires additional
investigation.
Taken together, the results from the present study
indicate the following: Oxidative stress induced by
H2O2 may cause dysfunction in HUVECs via
increasing the expression of SMS2 and activation of the
Wnt/β-catenin signaling pathway; and Dy105 may inhibit
H2O2-induced dysfunction of HUVECs through
inactivating the Wnt/β-catenin signaling pathway.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81560151)
and the Jiangxi Provincial Department of Science and Technology
(grant no. 20181BAB205022).
Availability of data and materials
The analyzed datasets generated during the study
are available from the corresponding author on reasonable
request.
Authors’ contributions
PZ, LH and HH performed the experiments and were
major contributors in writing the manuscript. XD and ZH analyzed
and interpreted the data and were responsible for the study design
and drafting of the manuscript. ML was responsible for statistical
analysis. XH prepared the reagents and revised the manuscript. NY
designed all of the study and provided funding. All of the authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Luscher TF, Landmesser U, von Eckardstein
A and Fogelman AM: High-density lipoprotein: Vascular protective
effects, dysfunction, and potential astherapeutic target. Circ Res.
114:171–182. 2014. View Article : Google Scholar
|
|
2
|
Otsuka F, Finn AV, Yazdani SK, Nakano M,
Kolodgie FD and Virmani R: The importance of the endothelium in
atherothrombosis and coronary stenting. Nat Rev Cardiol. 9:439–453.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Minuz P, Fava C, Vattemi G, Arcaro G,
Riccadonna M, Tonin P, Meneguzzi A, Degan M, Guglielmi V, Lechi A
and Tomelleri G: Endothelial dysfunction and increased oxidative
stress in mitochondrial diseases. Clin Sci (Lond). 122:289–297.
2012. View Article : Google Scholar
|
|
4
|
Zhang M, Pan H, Xu Y, Wang X, Qiu Z and
Jiang L: Allicin decreases lipopolysaccharide-induced oxidative
stress and inflammation in human umbilical vein endothelial cells
through suppression of mitochondrial dysfunction and activation of
nrf2. Cell Physiol Biochem. 41:2255–2267. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu D, Li D, Liu Z, Liu X, Zhou S and Duan
H: Role and underlying mechanism ofSPATA12in oxidative damage.
Oncol Lett. 15:3676–3684. 2018.PubMed/NCBI
|
|
6
|
Nohl H, Kozlov AV, Gille L and Staniek K:
Cell respiration and formation of reactive oxygen species: Facts
and artefacts. Biochem Soc Trans. 31:1308–1311. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dong L, Watanabe K, Itoh M, Huan CR, Tong
XP, Nakamura T, Miki M, Iwao H, Nakajima A, Sakai T, et al:
CD4+ T-cell dysfunctions through the impaired lipid
rafts ameliorate concanavalin A-induced hepatitis in sphingomyelin
synthase 1-knockout mice. Int Immunol. 24:327–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Adada M, Luberto C and Canals D:
Inhibitors of the sphingomyelin cycle: Sphingomyelin synthases and
sphingomyelinases. Chem Phys Lipids. 197:45–59. 2016. View Article : Google Scholar
|
|
9
|
Yeang C, Ding T, Chirico WJ and Jiang XC:
Subcellular targeting domains of sphingomyelin synthase 1–2. Nutr
Metab. 8:892011. View Article : Google Scholar
|
|
10
|
Jiang XC, Paultre F, Pearson TA, Reed RG,
Francis CK, Lin M, Berglund L and Tall AR: Plasma sphingomyelin
level as a risk factor for coronary artery disease. Arterioscler
Thromb Vasc Biol. 20:2614–2618. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schlitt A, Blankenberg S, Yan D, Von
Gizycki H, Buerke M, Werdan K, Bickel C, Lackner KJ, Meyer J,
Rupprecht HJ and Jiang XC: Further evaluation of plasma
sphingomyelin levels as a risk factor for coronary artery disease.
Nutr Metab (Lond). 3:52006. View Article : Google Scholar
|
|
12
|
Park TS, Panek RL, Rekhter MD, Mueller SB,
Rosebury WS, Robertson A, Hanselman JC, Kindt E, Homan R and
Karathanasis SK: Modulation of lipoprotein metabolism by inhibition
of sphingomyelin synthesis in apoe knockout mice. Atherosclerosis.
189:264–272. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mou D, Yang H, Qu C, Chen J and Zhang C:
Pharmacological activation of peroxisome proliferator-activated
receptor {delta increases sphingomyelin synthase activity in THP-1
macrophage-derived foam cell. Inflammation. 39:1538–1546. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dong J, Liu J, Lou B, Li Z, Ye X, Wu M and
Jiang XC: Adenovirus-mediated overexpression of sphingomyelin
synthases 1 and 2 increases the atherogenic potential in mice. J
Lipid Res. 47:1307–1314. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yan N, Ding T, Dong J, Li Y and Wu M:
Sphingomyelin synthase overexpression increases cholesterol
accumulation and decreases cholesterol secretion in liver cells.
Lipids Health Dis. 10:462011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou Q, Luo A and Kummerow FA: Lovastatin
reversed the enhanced sphingomyelin caused by 27-hydroxycholesterol
in cultured vascular endothelial cells. Biochem Biophys Rep.
5:127–133. 2015.PubMed/NCBI
|
|
17
|
de Jaime-Soguero A, Abreu de Oliveira WA
and Lluis F: The pleiotropic effects of the canonical wnt pathway
in early development and pluripotency. Genes Bales. 9:932018.
View Article : Google Scholar
|
|
18
|
Nusse R and Clevers H: Wnt/β-catenin
signaling, disease, and emerging therapeutic modalities. Cell.
169:985–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Foulquier S, Daskalopoulos EP, Lluri G,
Hermans KCM, Deb A and Blankesteijn WM: WNT signaling in cardiac
and vascular disease. Pharmacol Rev. 70:68–141. 2018. View Article : Google Scholar
|
|
20
|
Badimon L and Borrell-Pages M: Wnt
signaling in the vessel wall. Curr Opin Hematol. 24:230–239. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Christman MA II, Goetz DJ, Dickerson E,
McCall KD, Lewis CJ, Benencia F, Silver MJ, Kohn LD and Malgor R:
Wnt5a is expressed in murine and human atherosclerotic lesions. Am
J Physiol Heart Circ Physiol. 294:H2864–H2870. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Malgor R, Bhatt PM, Connolly BA, Jacoby
DL, Feldmann KJ, Silver MJ, Nakazawa M, McCall KD and Goetz DJ:
Wnt5a, TLR2 and TLR4 are elevated in advanced human atherosclerotic
lesions. Inflamm Res. 63:277–285. 2014. View Article : Google Scholar :
|
|
23
|
Tsaousi A, Mill C and George SJ: The Wnt
pathways in vascular disease: Lessons from vascular development.
Curr Opin Lipidol. 22:350–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee DK, Nathan Grantham R, Trachte AL,
Mannion JD and Wilson CL: Activation of the canonical
Wnt/beta-Catenin pathway enhances monocyte adhesion to endothelial
cells. Biochem Biophys Res Commun. 347:109–116. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ma S, Yao S, Tian H, Jiao P, Yang N, Zhu P
and Qin S: Pigment epithelium-derived factor alleviates endothelial
injury by inhibiting Wnt/β-Catenin pathway. Lipids Health Dis.
16:312017. View Article : Google Scholar
|
|
26
|
Quasnichka H, Slater SC, Beeching CA,
Boehm M, Sala-Newby GB and George SJ: Regulation of smooth muscle
cell proliferation by beta-catenin/T-cell factor signaling involves
modulation of cyclin D1 and p21 expression. Circ Res. 99:1329–1337.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsaousi A, Williams H, Lyon CA, Taylor V,
Swain A, Johnson JL and George SJ: Wnt4/β-Catenin signaling induces
VSMC proliferation and is associated with intimal thickening. Circ
Res. 108:427–436. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Luo S, Pan Z, Liu S, Yuan S and Yan N:
Sphingomyelin synthase 2 overexpression promotes cisplatin-induced
apoptosis of HepG2 cells. Oncol Lett. 15:483–488. 2018.PubMed/NCBI
|
|
29
|
Chengye Z, Daixing Z, Qiang Z and Shusheng
L: PGC-1-related coactivator (PRC) negatively regulates endothelial
adhesion of monocytes via inhibition of NF κB activity. Biochem
Biophys Res Commun. 439:121–125. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Song Z, Liu Y, Hao B, Yu S, Zhang H, Liu
D, Zhou B, Wu L, Wang M, Xiong Z, et al: Ginsenoside Rb1 prevents
H2O2-induced HUVEC senescence by stimulating sirtuin-1 pathway.
PLoS One. 9:e1126992014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
32
|
Zhang D, Wang Y, Dai Y, Wang J, Suo T, Pan
H and Liu H, Shen S and Liu H: Downregulation of RIP140 in
hepatocellular carcinoma promoted the growth and migration of the
cancer cells. Tumour Biol. 36:2077–2085. 2015. View Article : Google Scholar
|
|
33
|
Lou B, Dong J, Li Y, Ding T, Bi T, Li Y,
Deng X, Ye D and Jiang XC: Pharmacologic inhibition of
sphingomyelin synthase (SMS) activity reduces apolipoprotein-B
secretion from hepatocytes and attenuates endotoxin-mediated
macrophage inflammation. PLoS One. 9:e1026412014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Duan J, Yu Y, Li Y, Yu Y, Li Y, Zhou X,
Huang P and Sun Z: Toxic effect of silica nanoparticles on
endothelial cells through DNA damage response via chk1-dependent
G2/M checkpoint. PLoS One. 8:e620872013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Arlian LG, Elder BL and Morgan MS: House
dust mite extracts activate cultured human dermal endothelial cells
to express adhesion molecules and secrete cytokines. J Med Entomol.
46:595–604. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Han R, Tang F, Lu M, Xu C, Hu J, Mei M and
Wang H: Astragalus polysaccharide ameliorates
H2O2-induced human umbilical vein endothelial
cell injury. Mol Med Rep. 15:4027–4034. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang YK, Hong YJ, Wei M, Wu Y, Huang ZQ,
Chen RZ and Chen HZ: Curculigoside attenuates human umbilical vein
endothelial cell injury induced by H2O2. J
Ethnopharmacol. 132:233–239. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu M, Yu P, Jiang H, Yang X, Zhao J, Zou
Y and Ge J: The essential role of Pin1 via NF-κB signaling in
vascular inflammation and atherosclerosis in ApoE−/−
mice. Int J Mol Sci. 18:E6442017. View Article : Google Scholar
|
|
39
|
Liu M, Yu Y, Jiang H, Zhang L, Zhang PP,
Yu P, Jia JG, Chen RZ, Zou YZ and Ge JB: Simvastatin suppresses
vascular inflammation and atherosclerosis in ApoE−/−
mice by downregulating the HMGB1-RAGE axis. Acta Pharmacol Sin.
34:830–836. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen Y and Cao Y: The sphingomyelin
synthase family: Proteins, diseases and inhibitors. Biol Chem.
398:1319–1325. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang N, Huo R, Cai B, Lu Y, Ye B, Li X, Li
F and Xu H: Activation of Wnt/β-Catenin signaling by hydrogen
peroxide transcriptionally inhibits NaV1.5 expression. Free Radic
Biol Med. 96:34–44. 2017. View Article : Google Scholar
|
|
42
|
Liu P, Su J, Song X and Wang S: Activation
of nuclear β-Catenin/c-Myc axis promotes oxidative stress injury in
strep-tozotocin-induced diabetic cardiomyopathy. Biochem Biophys
Res Commun. 493:1573–1580. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tao J, Chen BD, Ma YT, Yang YN, Li XM, Ma
X, Yu ZX, Liu F, Xiang Y and Chen Y: FrzA gene protects
cardiomyocytes from H2O2-induced oxidative
stress through restraining the Wnt/frizzled pathway. Lipids Health
Dis. 14:902015. View Article : Google Scholar
|
|
44
|
Thakkar S, Wang X, Khaidakov M, Dai Y,
Gokulan K, Mehta JL and Varughese KI: Structure-based design
targeted at LOX-1, a receptor for oxidized low-density lipoprotein.
Sci Rep. 5:167402015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yao Y, Wang Y, Zhang Y and Liu C: Klotho
ameliorates oxidized low density lipoprotein (ox-LDL)-induced
oxidative stress via regulating LOX-1 and PI3K/Akt/eNOS pathways.
Lipids Health Dis. 16:772017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Luberto C and Hannun YA: Sphingomyelin
synthase, a potential regulator of intracellular levels of ceramide
and diacylglycerol during SV40 transformation. Does sphingomyelin
synthase account for the putative phosphatidylcholine-specific
phospholipase C. J Biol Chem. 273:14550–14559. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang Z, Kim S, Mahajan S, Zamani A and
Faccio R: Phospholipase cγ1 (PLCγ1) controls osteoclast numbers via
colony-stimulating factor 1 (csf-1)-dependent
diacylglycerol/β-catenin/cyclind1 pathway. J Biol Chem.
292:1178–1186. 2017. View Article : Google Scholar
|