Introduction
Dengue viruses (DENV) are single-stranded RNA
viruses in the genus Flavivirus (1). They are one of the most widespread
mosquito-borne viruses and are categorized into four serotypes
(DENV 1-4) (2). DENV infection
results in dengue fever and its more severe forms, dengue
hemorrhagic fever and dengue shock syndrome (3,4).
Dengue hemorrhagic fever and dengue shock syndrome are
characterized by increased vascular permeability, plasma leakage
into tissues, thrombocytopenia and hemorrhaging within internal
organs, which are thought to be immune-mediated diseases (5). According to the World Health
Organization, 50-100 million individuals are estimated to develop
dengue fever annually in tropical and subtropical regions of the
world such as South-East Asian and South American countries, and
~20,000 cases of mortality from dengue fever are estimated to occur
each year (6,7). At present, however, there are few
preventative antiviral drugs or vaccines against DENV infection.
Therefore, effective therapeutics and/or prophylaxis against DENV
infection are urgently required (8).
Type I interferons (IFNs) are cytokines that enhance
immunity to viruses or intracellular pathogens (9,10).
By signaling through specific receptors, type I IFNs activate a
janus kinase/signal transducer and activator of
transcription-dependent signaling cascade that increases the
expression of hundreds of IFN-stimulated genes (ISGs) to induce the
antiviral state (11). A number
of reports have demonstrated that DENV is sensitive to type I IFNs,
and that type I and II IFN-receptor-deficient mice are highly
susceptible to DENV infection (12-15). In addition, a number of
IFN-inducible anti-viral genes have been reported to inhibit DENV
infection (16-18).
Plasmacytoid dendritic cells (pDCs) represent a
unique immune cell type that is characterized by the production of
large quantities of type I IFNs in response to viruses or
self-nucleic acids (19,20). pDCs express Toll-like receptor
(TLR) 7 and 9 within their endosomes. TLR7 recognizes RNA viruses,
endogenous RNA and synthetic oligoribonucleotides, and TLR9 senses
non-methylated CpG DNA derived from viruses and bacteria,
endogenous DNA and synthetic CpG oligodeoxyribonucleotides (ODN)
(21-23). Recognition of these nucleic acids
by TLR7 or TLR9 in pDCs results in the production of type I IFNs
through the myeloid differentiation primary response gene 88
(MyD88)-IFN regulatory factor 7 pathway (24).
Lactic acid bacteria (LAB) have been reported to
have immune-modulatory effects on hosts through the activation of
innate immune cells, including macrophages or conventional DCs
(cDCs) (25). Although a number
of pathogenic bacteria have been demonstrated to stimulate pDCs
(26), beneficial bacteria,
including LAB, have been less studied in terms of activating pDCs
(27). It was previously reported
that Lactococcus lactis strain plasma (LC-plasma, also known
Lactococcus lactis subsp. lactis JCM 5805) stimulates
murine pDCs to produce large quantities of type I IFNs in
association with cDCs (28). It
was further demonstrated that the effect of LC-plasma on the
activation of pDCs is dependent on the TLR9-MyD88 signaling
pathway. As pDCs serve a pivotal function in anti-viral immune
responses through type I IFN production, the effect of LC-plasma on
virus infection was verified using virus infection mouse models. It
was demonstrated that murine para-influenza virus-infected mice
that were fed LC-plasma demonstrated a markedly improved survival
rate compared with controls (29). Furthermore, the oral
administration of LC-plasma protected mice against rotavirus
infection (30). Therefore, the
activation of pDCs by LAB administration appeared to defend the
host against viral infection.
In the present study, the effect of a
pDC-stimulating LAB, LC-plasma, on DENV infection in mice was
examined. To verify the effectiveness of LC-plasma against DENV
infection, the virus titer and the expression of inflammatory genes
in DENV-infected tissues when LC-plasma was orally administered
were examined. In addition, to elucidate the mechanism of
protection against DENV infection by LC-plasma administration, the
present study focused on anti-viral gene expression, which is
assumed to be effective for preventing DENV infection (31). Finally, an in vitro study
was performed and examined the influence of type I IFNs induced by
LC-plasma on the enhancement of anti-viral gene expression. To
date, whereas certain types of LAB have been reported to have
immune-modulatory effects on host immunity, it has remained unclear
whether LAB exerts an effect on DENV infection, which is a major
threat to human health. This is the first report, to the best of
our knowledge, to investigate whether the administration of LAB is
effective against DENV infection in vivo.
Materials and methods
Mice
A total of 56 C57BL/6J female mice (6 week-old;
weight, 15-25 g) were purchased from Charles River Laboratories
Japan, Inc. (Kanagawa, Japan). All animals were maintained in a
specific pathogen-free facility under a 12 h light/dark cycle with
ad libitumaccess to water and a basic diet of AIN93G pellets
(Oriental Yeast Co., Ltd., Tokyo, Japan). The temperature in the
facility was maintained at 22-25°C and 40-60% humidity. All animals
were sacrificed using 5% isoflurane exposure until 2-3 min after
the breathing had ceased. All animal experiments were performed in
accordance with the guidelines for the care and use of laboratory
animals of Juntendo University (Tokyo, Japan) and Kirin Company,
Ltd. (Kanagawa, Japan). The study was ethically approved by the
Committee for Animal Experimentation of Juntendo University and
Kirin Company, Ltd. All efforts were made to minimize animal
suffering.
LAB strains
LAB strains tested in this study were purchased from
collections held at the Japan Collection of Microorganisms
(Tsukuba, Japan) and American Type Culture Collection (ATCC;
Manassas, VA, USA). Lactobacillus rhamnosus strain GG
(53103; ATCC) was grown at 37°C for 48 h in MRS broth (Oxoid;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) and LC-Plasma was
grown at 30°C for 48 h in M17 broth (Oxoid; Thermo Fisher
Scientific, Inc.) according to the manufacturer’s protocol.
Cultured LAB strains were washed twice with sterile distilled
water, heat-killed at 100°C, lyophilized and suspended in
phosphate-buffered saline for the in vitro study.
Oral intake of LC-plasma
Six week-old C57BL/6J female mice were acclimatized
for 1 week with ad libitum access to water and a basic diet
of AIN93G pellets. Control groups were fed with 4 g/day AIN93G
pellets and test groups were fed with 4 g/day AIN93G pellets
containing 1 mg heat-killed LC-plasma during the course of the
study.
Virus infection study
A total of 32 mice were divided into two groups
(control; n=16, LC-plasma; n=16) and housed with 4 animals/cage
following LC-plasma oral administration for 2 weeks as described
above. A 5×107 pfu/ml DENV virus solution was prepared,
and mice were intraperitoneally injected with 200 µl
solution. Mice were monitored every day subsequent to viral
infection by monitoring the intake of food and water, the state of
breathing and other abnormal behaviors. As the virus titer peaks at
2-3 days following virus infection, mice were sacrificed 2-3 days
subsequent to infection, and spleens, liver and blood were
harvested. The spleen and liver were homogenized in TRIzol reagent
(Life Technologies; Thermo Fisher Scientific, Inc.) for 1 min at
room temperature followed by RNA extraction. Total RNA was
extracted by phenol/chloroform/isoamyl alcohol extraction method.
Blood samples were collected in EDTA-treated tubes (Terumo Co.,
Tokyo, Japan) that were subjected to centrifugation at 400 × g for
20 min at room temperature. Subsequent to centrifugation, plasma
was obtained from blood samples for NS1 enzyme-linked immunosorbent
assay (ELISA).
Non-virus infection study
A total of 20 mice were divided into two groups
(control, n=10; LC-plasma, n=10) and housed at 1/cage following
LC-plasma oral administration for 2 weeks as described above. After
2 weeks of oral intake of LC-plasma, the mice were sacrificed and
the spleens were collected. The spleen was minced in
Mg2+- and Ca2+-free Hank’s Balanced Salt
solution (Gibco; Thermo Fisher Scientific, Inc.) and digested with
1 mg/ml collagenase (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
for 20 min at 37°C. EDTA (Invitrogen; Thermo Fisher Scientific,
Inc.) was adjusted to 30 mM and the sample was incubated for 10 min
at room temperature. Tissue lysates were filtered through a 100
µm nylon cell strainer and erythrocytes were removed by
exposure to 2 ml of Red Blood Cell Lysis Buffer (Sigma-Aldrich;
Merck KGaA) for 5 min at room temperature. Tissue lysates were
filtered through a 70 µm nylon cell strainer again and
filtered tissue lysates were used for splenocytes.
CD11c+ cells were separated from splenocytes using CD11c
MicroBeads (Miltenyi Biotec, Inc., Cambridge, MA, USA). The
mediated splencytes and CD11c+ cells were suspended in
RNAlater Stabilization solution (Thermo Fisher Scientific,
Inc.).
Viral strains and growth conditions
DENV strain Eden2 was used in the present study.
DENV Eden2 was provided by Dr. Ashley L. St. John of Duke-NUS
Medical School (College Road, Singapore) and C6/36 cells and Baby
hamster kidney 21 cells were provided by Dr. Yoichi Suzuki of Osaka
Medical College (Osaka, Japan). DENV Eden2 was grown in Aedes
albopictusC6/36 cells. C6/36 cells were seeded in plastic
flasks and grown in RPMI-1640 medium (Thermo Fisher Scientific,
Inc.) supplemented with 10% heat-inactivated fetal bovine serum
(Moregate Biotech, Bulimba, Australia) and 100 U/ml
penicillin-streptomycin (Thermo Fisher Scientific, Inc.). Once the
cells were fully confluent, DENV Eden2 was transferred to C6/36
flasks at a multiplicity of infection of 0.01. Viral samples were
incubated for >1 h at 28°C, and subsequently the medium was
substituted using RPMI-1640 medium supplemented with 2% inactivated
fetal bovine serum. Cells were cultured for 5-7 days, and culture
supernatants were collected. Cell debris was removed by
centrifugation at 800 × g for 10 min at 4°C, and supernatants
containing the virus were stored at −80°C. Viral titers were
determined by plaque assays using Baby hamster kidney 21 cells.
Cells were seeded in 24-well plates in RPMI-1640 medium
supplemented with 10% inactivated fetal bovine serum and 100 U/ml
penicillin-streptomycin. Cells were maintained in a humidified
incubator at 5% CO2 and 37°C until they reached
confluence. Then, the medium was removed and the monolayer was
infected with 200 ml of 10-, 102-, 103-,
104-, 105- and 106-fold diluted
virus samples. The plates were incubated for 1 h at 37°C. After 1
h, cells were washed and then 0.5 ml 1% carboximethylcellulose
(Sigma-Aldrich; Merck KGaA) mixed with RPMI-1640 medium
supplemented with 2% inactivated fetal bovine serum and 100 U/ml
penicillin-streptomycin was added to each well. The plates were
maintained in a humidified incubator at 37°C and 5% CO2
for 5-6 days. Subsequent to this period, the cells were fixed with
4% paraformaldehyde phosphate buffer solution (Wako Pure Chemical
Industries, Ltd., Osaka, Japan) at room temperature for 1 h and the
carboxymethylcellulose plugs were removed by washing with water.
Plaques were visualized by staining with a solution of 1% crystal
violet (Wako Pure Chemical Industries, Ltd.) in 20% ethanol for 2-3
min at room temperature.
NS1 ELISA
Concentrations of NS1 antigen in plasma samples were
measured using a commercially available DENV NS1 ELISA kit
purchased from Biocompare (South San Francisco, CA, USA).
In vitro co-culture assays
BM cells were isolated from C57BL/6J mice, and
erythrocytes were removed by exposure to 1 ml of Red Blood Cell
Lysis Buffer (Sigma-Aldrich; Merck KGaA) for 1 min at room
temperature. Cells were cultured at a density of 5×105
cells/ml for 7 days in RPMI-1640 medium supplemented with 1 mM
sodium pyruvate (Life Technologies; Thermo Fisher Scientific,
Inc.), 2.5 mM HEPES (Life Technologies; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin-streptomycin (Life Technologies; Thermo
Fisher Scientific, Inc.), 50 µM 2-mercaptoethanol (Life
Technologies; Thermo Fisher Scientific, Inc.), 10% fetal bovine
serum and 100 ng/ml FMS-like tyrosine kinase 3 ligand (Flt-3L;
R&D Systems, Inc., Minneapolis, MN, USA) at 37°C in a
humidified atmosphere of 5% CO2 and 95% air. After 7
days of culturing, cells were washed with RPMI-1640 medium and then
incubated at a density of 2×105 cells/ml with 1
µM CpG ODN (InvivoGen, Toulouse, France) or 10 µg/ml
LC-plasma or Lactobacillus rhamnosus strain GG (ATCC 53103)
at 37°C for 24 h. To block TLR9 signals, a TLR9 antagonist
(InvivoGen) was added with LC-plasma. To neutralize IFN-α/β,
anti-IFN-α and anti-IFN-β antibody (PBL Assay Science, Piscataway,
NJ, USA) was added with LC-plasma. Subsequent to culturing, the
supernatant was collected and other BMDC or spleno-cytes were
cultured with the collected supernatant for 24 h. Subsequent to
culturing with the collected supernatant, each cell sample was
collected for RNA extraction.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from spleen and liver was extracted using
TRIzol reagent (Ambion; Thermo Fisher Scientific, Inc.). Subsequent
to the extraction of tissue total RNA, a purification step was
performed using an RNeasy mini kit (Qiagen GmbH, Hilden, Germany)
according to the manufacturer’s protocol. Total RNA from
splenocytes, CD11c+ cells or BMDC was extracted using an
RNeasy mini kit. To synthesize complementary DNA, purified total
RNA samples were reverse transcribed using an iScript cDNA
synthesis kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
according to the manufacturer’s protocol. RT-qPCR (qRT-PCR) was
performed using SYBR Premix Ex Taq (Takara Bio, Inc, Otsu, Japan)
and a LightCycler 480 (Roche Diagnostics, Basel, Switzerland). The
PCR conditions were as follows: 2-Step Cycling, 95°C for 10 sec
hold, 45 cycles of 95°C for 5 sec and 60°C for 20 sec. All values
were normalized to the expression of GAPDH and calculated using the
2−ΔΔCq method (32).
Specific forward and reverse primer pairs that were used are listed
as follows: GAPDH forward, 5′-TGT GTC CGT CGT GGA TCT GA-3′ and
reverse, 5′-TTG CTG TTG AAG TCG CAG GAG-3′; interleukin (IL)-6
forward, 5′-CCA CTT CAC AAG TCG GAG GCT TA-3′ and reverse, 5′-TGC
AAG TGC ATC ATC GTT GTTC-3′; tumor necrosis factor α (TNFα)
forward, 5′-ACT CCA GGC GGT GCC TAT GT-3′ and reverse, 5′-GTG AGG
GTC TGG GCC ATA GAA-3′; IL-1β forward, 5′-TCC AGG ATG AGG ACA TGA
GCA C-3′ and reverse, 5′-GAA CGT CAC ACA CCA GCA GGT TA-3′;
monocyte chemoattractant protein 1 (MCP-1) forward, 5′-AGC AGC AGG
TGT CCC AAA GA-3′ and reverse, 5′-GTG CTG AAG ACC TTA GGG CAG A-3′;
IFNγ forward, 5′-CGG CAC AGT CAT TGA AAG CCT A-3′ and reverse,
5′-GTT GCT GAT GGC CTG ATT GTC-3′; interferon-induced GTP-binding
protein Mx1 (Mx1) forward, 5′-AAT GTG GAC ATT GCT ACC ACA GA-3′ and
reverse, 5′-GGT TCC GCA TCA CAT CCA AG-3′; Viperin forward, 5′-CCG
CAG TAA CTC AGC TCA TGG A-3′ and reverse, 5′-CTG AGC ATT AGA CCT
CAT CTG GAC A-3′; 2′-5′-oligoadenylate synthase 1A (Oas1a) forward,
5′-TGC TGC CAG CCT TTG ATG TC-3′ and reverse, 5′-TCC TCG ATG AGG
ATG GCA TAG A-3′; ISG15 forward, 5′-CTG TGA GAG CAA GCA GCC AGA-3′
and reverse, 5′-GAG TTA GTC ACG GAC ACC AGG AA-3′.
In vivo virus quantitation
Methods for total RNA extraction and purification
were performed as described above. RT-qPCR was performed using
TaqMan Fast Virus 1-Step Master Mix (Thermo Fisher Scientific,
Inc.) and a LightCycler 480 (Roche Diagnostics). The PCR conditions
were as follows: 2-Step Cycling, 50°C for 5 min hold, 45 cycles of
95°C for 20 sec and 60°C for 30 sec. All values were normalized to
the expression of GAPDH and calculated using the 2−ΔΔCq
method (32). The specific
forward and reverse primer pairs and probes used are listed as
follows: GAPDH forward, 5′-CTT CAC CAC CAT GGA GAA GGC-3′ and
reverse, 5′-GGC ATG GAC TGT GGT CAT GAG-3′; GAPDH probe,
5′-(6-FAM)-CCT GGC CAA GGT CAT CCA TGA CAA CTT T-(TAMRA)-3′; DENV
Eden2 forward, 5′-CAT ATT GAC GCT GGG AAA GA-3′ and reverse, 5′-AGA
ACC TGT TGA TTC AAC-3′; DENV Eden2 probe, 5′-(6-FAM)-CTG TCT CCT
CAG CAT CAT TCC AGG CA-(TAMRA)-3′.
Statistical analysis
The data of virus titer and NS1 ELISA were presented
as the mean + the value of each individual. All other data were
presented as the mean ± standard error of the mean. Data from the
in vivo study were analyzed using an unpaired Student’s
t-test. Data from the in vitro study were analyzed using
one-way analysis of variance followed by Dunnett’s test. All
statistical analyses were performed using the Excel-Toukei 2012
software program (Social Survey Research Information, Tokyo,
Japan). P<0.05 was considered to indicate a statistically
significant difference.
Results
Oral administration of LC-plasma inhibits
DENV infection
To investigate whether the pDC-stimulative LAB,
LC-plasma, is effective for the prevention of DENV infection in
mice, the mouse-infectious DENV strain Eden2 was used. As DENV
Eden2 has been previously reported to infect the spleen and liver
and the virus infection peaks at 48 or 72 h (33), the relative virus titers were
assessed at 48 and 72 h following DENV infection to examine the
effect of LC-plasma on DENV infection. The study design is
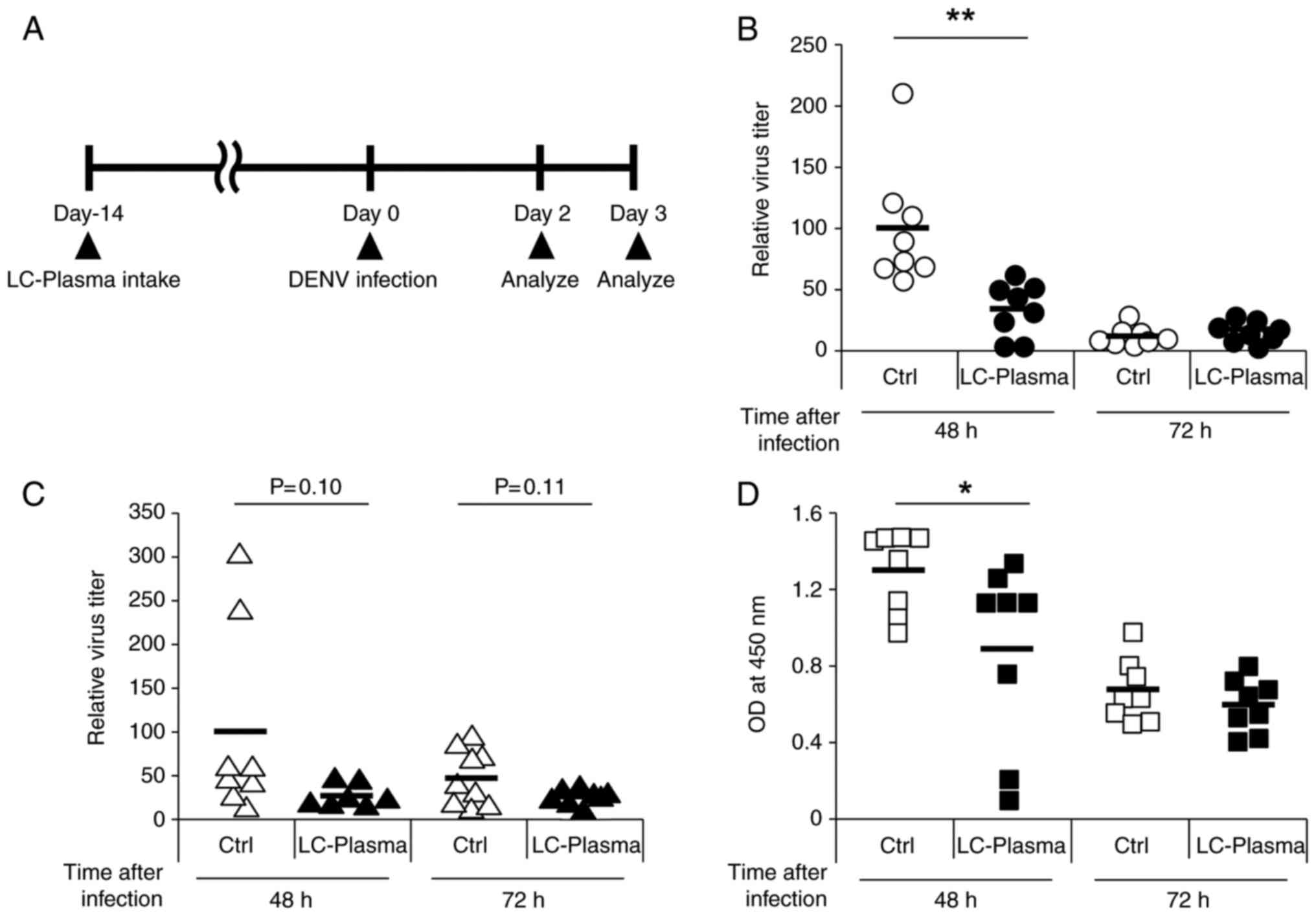
presented in Fig. 1A. The mice
were orally administered LC-plasma for two weeks, and subsequently
were infected with a titer of 1×107 pfu of DENV Eden2 by
intraperitoneal injection. Oral intake of LC-plasma was continued
for 2-3 days subsequent to DENV infection. A total of 2-3 days
following DENV infection, 8 mice from each group were sacrificed,
and the spleens, liver and blood were collected. No animals
succumbed to DENV infection and any behavioral change was not
observed. As presented in Fig.
1B, the relative virus titer in the spleens of the LC-plasma
group 48 h subsequent to infection was significantly lower compared
with that in the control group (P<0.01). On the other hand,
there was no significant difference in the relative virus titers in
the spleens at 72 h after infection between the two groups. The
relative virus titer in the liver is presented in Fig. 1C. In the LC-plasma group, the
relative virus titers at 48 and 72 h following infection were
significantly lower compared with that of the control group (P=0.10
and P=0.11, respectively). NS1 antigen, which is a non-structural
protein of the DENV (34), was
also measured in the blood and the amount of NS1 at 48 h in the
LC-plasma group was significantly lower compared with the control
group (P<0.05). However, there was no difference in the amount
of NS1 between the two groups 72 h after infection (Fig. 1D). These results suggested that
oral administration of LC-plasma is effective in the prevention of
DENV infection.
LC-plasma alleviates DENV-mediated
inflammation
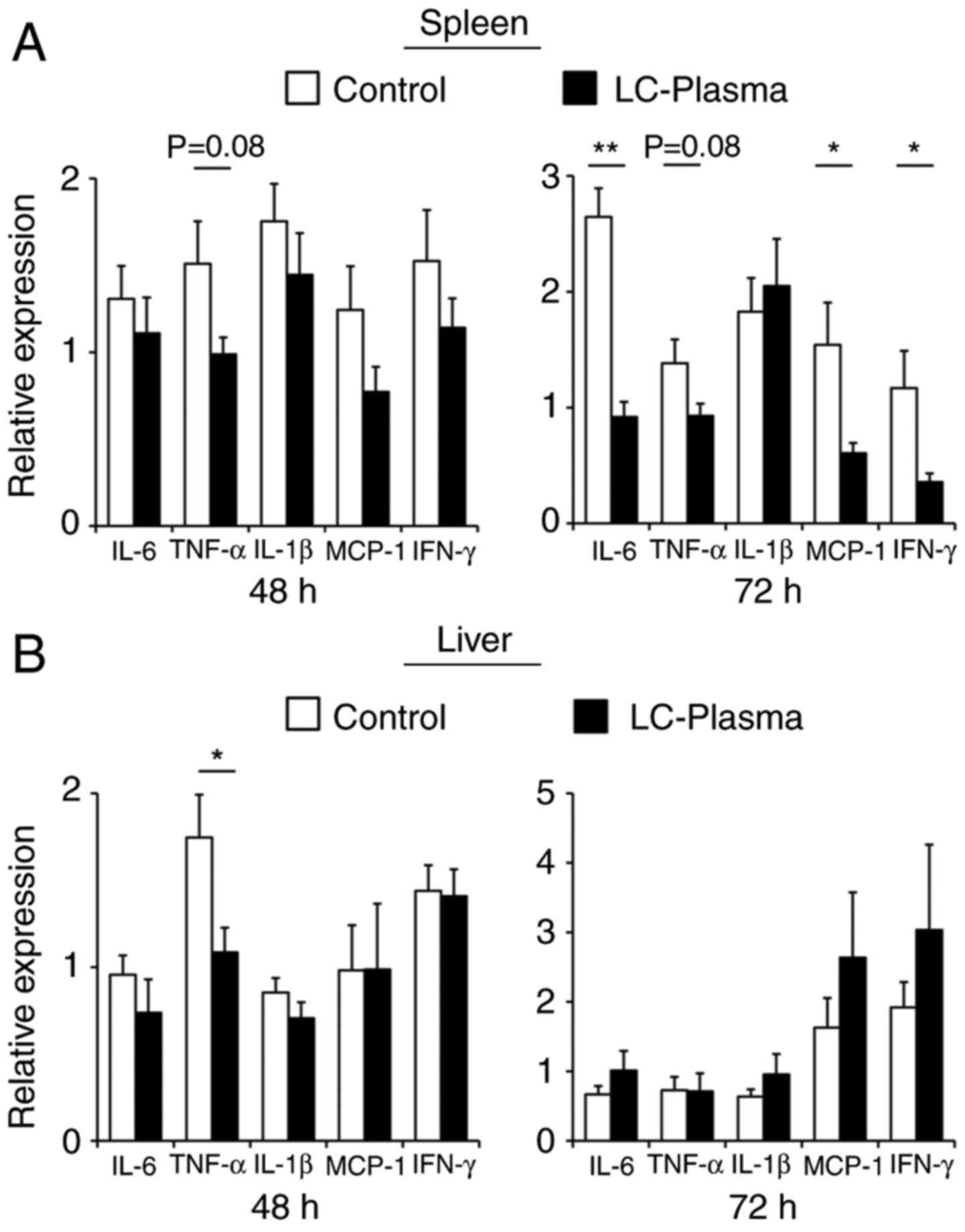
To evaluate the effect of LC-plasma on the
suppression of inflammatory responses induced by DENV infection,
the expression levels of inflammatory cytokine genes in the spleen
and liver at 48 and 72 h subsequent to DENV infection were assessed
by RT-qPCR. As presented in Fig.
2A, in the LC-plasma group, the expression levels of IL-6
(P<0.01), MCP-1 (P<0.05) and IFN-γ (P<0.05) in the spleen
72 h after infection were significantly lower compared with that in
the control group. In addition, TNF-α expression levels in the
spleen at 48 and 72 h after infection were non-significantly lower
compared with that of the control group (P=0.08 and P=0.08,
respectively). However, only TNF-α expression levels in the
liver 48 h after infection were significantly lower compared with
the control group (P<0.05), whereas the expression of other
cytokine genes in the liver remained unchanged. These results
suggest that the oral administration of LC-plasma alleviated
inflammatory responses in DENV infected tissue.
Oral administration of LC-plasma enhances
anti-viral factors in the spleen and CD11c+ cells in the
absence of the virus
It was demonstrated that oral intake of LC-plasma is
effective against DENV infection and the associated inflammatory
response. To elucidate how orally administered LC-plasma exerts an
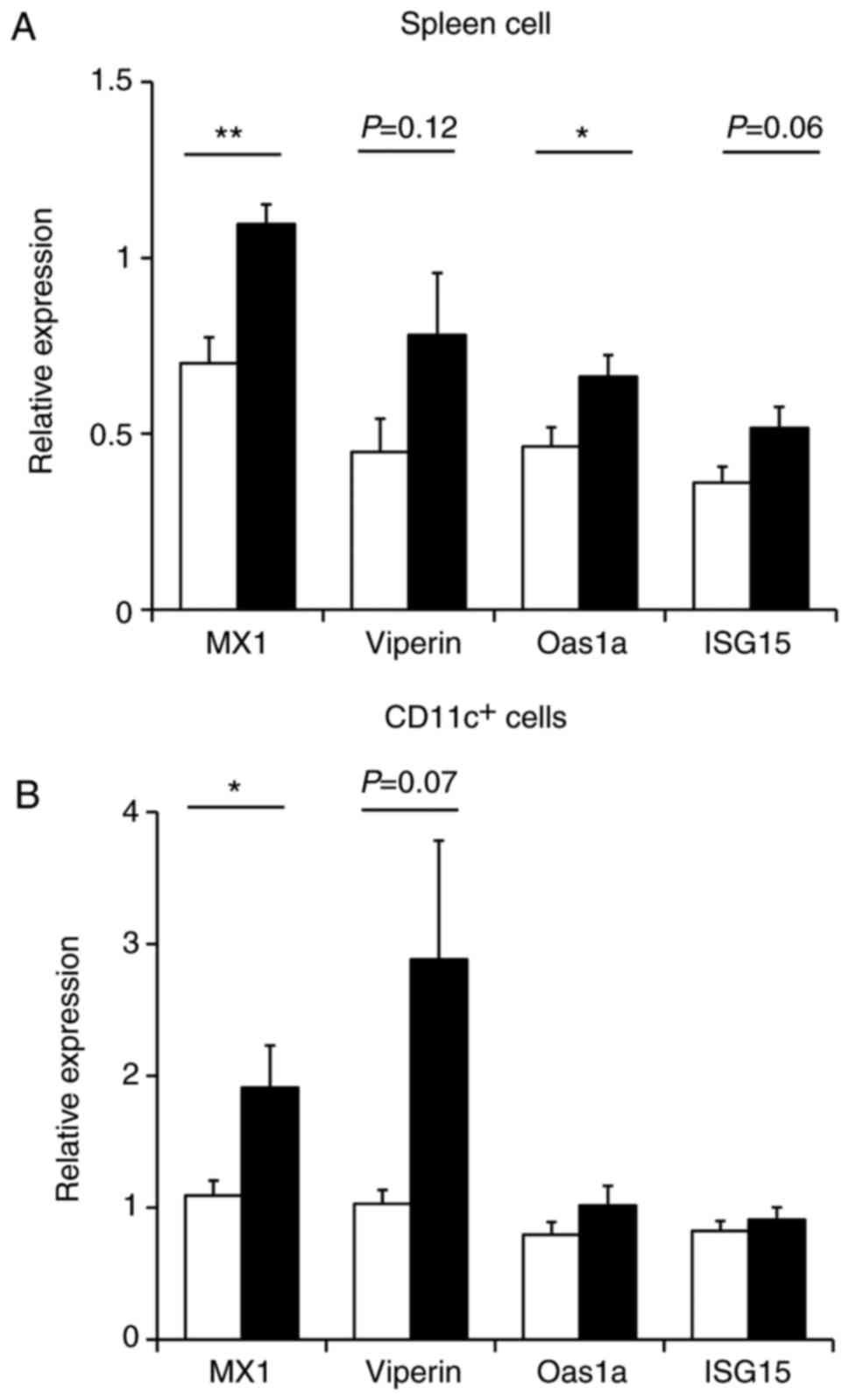
inhibitory effect on DENV, the expression of anti-viral factor
genes in the spleen were evaluated, which is a primary site of DENV
infection (33). Subsequent to
two weeks of LC-plasma intake, 10 mice from each group were
sacrificed, the spleens were harvested and the expression of
anti-viral factor genes in the spleen were measured using RT-qPCR.
As presented in Fig. 3A, oral
administration of LC-plasma significantly enhanced the expression
levels of Mx1 (P<0.01) and Oas1a (P<0.05) compared with the
control group. In addition, LC-plasma non-significantly increased
the expression levels of Viperin and ISG15 compared with the
control group (P=0.12 and P=0.06, respectively). Thus, LC-plasma
administration caused the induction of anti-viral factor gene
expression in the spleen. Among the various immune cells in the
spleen, CD11c+ cells are the major target cells for DENV
infection (33). Therefore,
anti-viral gene expression in CD11c+ cells was also
measured, which were magnetically separated from the spleen tissue.
As presented in Fig. 3B,
LC-plasma administration significantly induced the expression
levels of Mx1 (P<0.05) and also non-significantly enhanced
Viperin expression levels (P=0.07) in CD11c+ cells from
the spleen compared with the control group. On the other hand,
Oas1a and ISG15 expression levels in CD11c+ cells were
not influenced by LC-plasma administration. These results
demonstrate that LC-plasma may alleviate DENV infection through an
increase of anti-viral factor gene expression in infected
tissue.
LC-plasma enhances anti-viral factors
through type I INF in vitro
In vivo studies revealed that the oral
treatment of LC-plasma enhanced the anti-viral factors in DENV
infected tissue, which may be one mechanism used by LC-plasma to
alleviate DENV infection. As LC-plasma strongly induces type I IFNs
through pDC activation, it was investigated whether the effect of
LC-plasma on the enhancement of anti-viral factors was dependent on
type I IFNs through the use of in vitro studies. To assess
this hypothesis, Flt-3L-induced bone-marrow derived DC (BMDC) were
used, which includes pDC and cDC (35). BMDC were stimulated with the TLR9
agonist CpG ODN, or with the LAB strains of LC-plasma or
Lactobacillus rhamnosus strain GG (ATCC 53103). After 24 h
of stimulation, BMDC were collected and the expression levels of
anti-viral genes were analyzed. As presented in Fig. 4A, stimulation of BMDC with CpG ODN
or LC-plasma significantly increased the expression of the
anti-viral factor genes compared with the control group (P<0.05;
Fig. 4A). In contrast, ATCC 53103
did not influence the expression of the anti-viral genes. To
investigate whether this LC-plasma effect is dependent on soluble
factors including interleukins or interferons produced by BMDC, the
supernatants were collected and another sample of BMDC was
re-stimulated with these supernatants. As presented in Fig. 4B, the supernatants from BMDC
stimulated with LC-plasma also significantly enhanced anti-viral
gene expression in another BMDC sample (P<0.01). However, this
effect was completely abolished by TLR9 antagonist treatment.
Furthermore, anti-IFNα/β antibody treatment with BMDC stimulated
with LC-plasma also blocked this effect. To investigate whether
this LC-plasma effect also occurs in the spleen, which is the major
DENV infected tissue, supernatants collected from BMDC stimulated
with LC-plasma were added to splenocytes. As presented in Fig. 4C, LC-plasma supernatants also
significantly enhanced anti-viral gene expression levels compared
with the control group (P<0.01) and this effect was completely
abolished by TLR9 antagonist or anti-IFNα/β antibody treatment.
Based on these results, it appears that LC-plasma may attenuate
DENV infection through type I IFN production by pDCs.
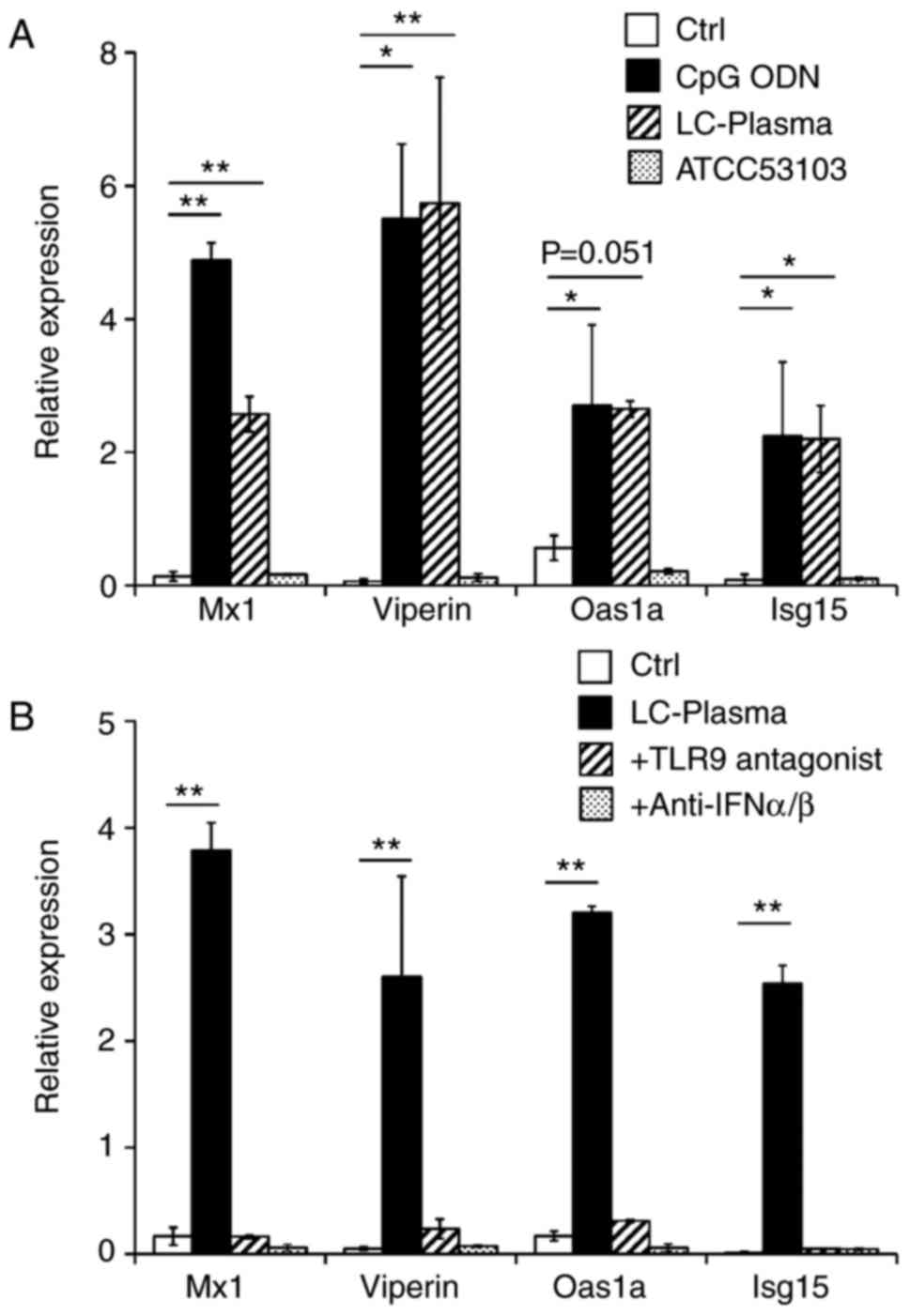 | Figure 4Enhancement of anti-viral factor gene
expression mediated by LC-plasma via type I IFNs in vitro.
(A) Flt-3L induced BMDC were cultured with CpG ODN, LC-plasma or
ATCC 53103 for 24 h. Subsequent to culturing, BMDC were collected
for RNA extraction. (B) Flt-3L-induced BMDC were cultured with
LC-plasma in the presence or absence of a TLR9 antagonist or
anti-IFN-α/β antibody for 24 h. After culturing, the supernatant
was collected and another sample of Flt-3L-induced BMDC was
cultured with the collected supernatant for 24 h. Subsequent to
culturing with the supernatant, BMDC were collected for RNA
extraction. (C) Flt-3L-induced BMDC were cultured with LC-plasma in
the presence or absence of TLR9 antagonist or anti-IFN-α/β antibody
for 24 h. Subsequent to culturing, the supernatant was collected
and the splenocytes were cultured with the collected supernatant
for 24 h. After culturing with the supernatant, splenocytes were
collected for RNA extraction. Total RNA was extracted from each
cell culture to analyze Mx1, Viperin, Oas1a and ISG15 expression by
reverse transcription-quantitative polymerase chain reaction
normalized to GAPDH. Data are presented as the mean ± standard
error of the mean. *P<0.05 and **P<0.01
with comparisons shown by lines. LC-plasma, Lactococcus
lactis strain plasma; IFN, interferon; Flt-3L, FMS-like
tyrosine kinase 3 ligand; BMDC, bone-marrow derived dendritic
cells; ODN, oligodeoxyribonucleotides; TLR9, Toll-like receptor 9;
Mx1, interferon-induced GTP-binding protein Mx1; Oas1a,
2′-5′-oligoadenylate synthase 1A; ISG15, IFN-stimulated gene
15. |
Discussion
To date, despite major efforts, there exists no
effective treatment or vaccine for DENV infection. It has been
previously reported that the specific LAB, LC-plasma, stimulates
pDCs to induce the production of a large quantity of type I IFNs
(28). As pDCs and type I IFNs
serve a pivotal role in anti-viral immune responses, it was
predicted that pDC-stimulative LAB would be effective against DENV.
In the present study, the effect of LC-plasma on DENV infection was
examined using a mouse model.
A major technical barrier in developing therapeutics
or vaccines against DENV infection is the absence of a suitable
animal model that mimics dengue disease. The first established
animal models used immune-competent mice in which a high dose of
DENV was administered intracranially (36). Other animal models included
immunocompromised mice lacking type I and/or type II IFNs receptors
(37,38). However, neither of these models is
suitable to evaluate treatment or vaccines for DENV as the route of
infection is different from the usual DENV infection route in one
model, and the other uses an immunocompromised mouse lacking the
relevant immune receptor(s) (39). Previously, a DENV strain capable
infecting mice, DENV Eden2, was reported (33,40). This strain was used to infect
wild-type mice by intraperitoneal injection and it was revealed be
capable of replicating in the spleen and liver. These organs are
also DENV-target organs in humans, so this strain is appropriate
for evaluating the effectiveness of therapeutics for DENV.
Therefore, the present study used the DENV Eden2 strain to evaluate
the effect of LC-plasma on DENV infection in vivo, and
revealed that LC-plasma is effective against DENV infection by
measuring the virus titer in the spleen and the liver and the NS1
expression level in blood.
It was also revealed that inflammatory gene
expression in DENV infected tissue was inhibited by oral
administration of LC-plasma. DENV infection in children and adults
usually results in dengue fever accompanied by a combination of
symptoms that includes headache, retro-orbital pain, myalgia and
hemorrhagic manifestations (41,42). Certain patients including newborns
and elderly people occasionally have an onset of dengue hemorrhagic
fever, the severest form of dengue disease. Patients with dengue
hemorrhagic fever manifest a cytokine storm, with high levels of
circulating pro-inflammatory cytokines and chemokines, which
results in endothelial damage and vascular leakage with
hemorrhaging and shock (6,42).
Therefore, excessive inflammation induced by DENV infection results
in severe dengue disease and the inhibition of inflammatory
cytokines or chemokines may result in a reduction of
DENV-associated inflammatory symptoms (43,44). Thus, LC-plasma administration may
contribute to the suppression of DENV pathogenesis.
A number of reports have demonstrated that type I
IFNs serve a principal role in inhibiting DENV replication
(12-15). One of the essential roles of type
I IFNs is the induction of anti-viral factors, which are involved
in the suppression of viral replication and release (16-18). Thus, it was predicted that
LC-plasma administration may inhibit DENV replication through the
enhancement of anti-viral factors in DENV infected tissues. To
reveal the mechanism of the LC-plasma effect against DENV
infection, it was investigated whether LC-plasma enhances the
expression of anti-viral genes in DENV infected tissues and cells.
The expression of four important anti-viral genes, Mx1, Viperin,
Oas1a and Isg15, were assessed. Mx1 is an interferon-induced
GTP-binding protein that prevents transcription of DENV by
interacting with viral polymerase (45). Viperin is a multifunctional
antiviral factor that inhibits the replication of DNA and RNA
viruses including hepatitis C virus, cytomegalovirus, influenza
virus and DENV (46). Isg15 is an
ubiquitin-like interferon-stimulated protein that conjugates viral
proteins, resulting in the suppression of viral release (47). Oas1a is a 2′,5′-oligoadenylate
synthase that synthesizes higher oligomers of 2′-5′-oligoadenylates
from ATP. The oligoadenylates activate RNase L, which degrades
viral RNAs (48). Oral
administration of LC-plasma significantly enhanced Mx1 and Oas1a
and non-significantly enhanced Viperin and Isg15 in the spleen.
Furthermore, LC-plasma induced Mx1 and Viperin in CD11c+
cells, which is one of the cell types that DENV infects. The
results of the present study indicate that LC-plasma may suppress
DENV replication by inducing the expression of these anti-viral
genes.
The in vitro experiments in the present study
clearly demonstrated that LC-plasma increases the expression of
anti-viral genes; and this effect was completely abolished by TLR9
antagonists or anti-type I IFN antibody treatment. However, another
strain of LAB, ATCC 53103, which does not induce type I IFNs, did
not have the ability to enhance anti-viral gene expression. These
results demonstrate that pDC-stimulative LAB strongly inhibit viral
infection through type I IFN production.
To date, various strains of LAB have been reported
to have immune-modulatory effects by affecting immune cells,
resulting in anti-pathogenic effects or improving allergy symptoms
(49-51). In the present study, it was
revealed for the first time that the pDC-stimulating LAB,
LC-plasma, is effective against DENV infection. As LC-plasma also
has this effect on different types of viruses (29,30), pDC-stimulating LAB may
additionally be effective against other viruses.
Acknowledgments
The authors would like to thank Dr. Ashley L. St.
John in Duke-NUS Medical School (College Road, Singapore) for
providing Dengue virus strain Eden2 and Dr. Yoichi Suzuki in Osaka
Medical College (Osaka, Japan) for providing C6/36 cells and Baby
hamster kidney 21 cells.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors’ contributions
HS and NoY designed the experiments. HS, RT, MS and
NoY performed the experiments and analyzed the data. NaY, NoY and
OK supervised the experiments. HS wrote the paper and all authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were performed in strict
accordance with the guidelines for the care and use of laboratory
animals of Juntendo University (Tokyo, Japan) and Kirin Company
(Kanagawa, Japan). The studies were approved by the Committee for
Animal Experimentation of Juntendo University and Kirin
Company.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
DENV
|
dengue virus
|
|
IFNs
|
interferons
|
|
ISGs
|
IFN-stimulated genes
|
|
LAB
|
lactic acid bacteria
|
|
LC-plasma
|
Lactococcus lactis strain plasma
|
|
pDCs
|
plasmacytoid dendritic cells
|
|
cDCs
|
conventional dentritic cells
|
|
TLR
|
toll-like receptors
|
|
ODN
|
oligodeoxyribonucleotides
|
|
MyD88
|
myeloid differentiation primary
response 88
|
|
BMDC
|
bone-marrow derived dendritic
cells
|
References
|
1
|
Diamond MS and Pierson TC: Molecular
Insight into dengue virus pathogenesis and its implications for
disease control. Cell. 162:488–492. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Holmes EC and Burch SS: The causes and
consequences of genetic variation in dengue virus. Trends
Microbiol. 8:74–77. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Halstead SB: Dengue. Lancet.
370:1644–1652. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gubler DJ: Dengue/dengue haemorrhagic
fever: History and current status. Novartis Found Symp. 277:3–16.
16–22. 71–73. 251–253. 2006.
|
|
5
|
Srikiatkhachorn A, Mathew A and Rothman
AL: Immune- mediated cytokine storm and its role in severe dengue.
Semin Immunopath. 39:563–574. 2017. View Article : Google Scholar
|
|
6
|
Guzman MG and Kouri G: Dengue: An update.
Lancet Infect Dis. 2:33–42. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kalayanarooj S, Vaughn DW, Nimmannitya S,
Green S, Suntayakorn S, Kunentrasai N, Viramitrachai W,
Ratanachu-eke S, Kiatpolpoj S, Innis BL, et al: Early clinical and
laboratory indicators of acute dengue illness. J Infect Dis.
176:313–321. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fagundes CT, Costa VV, Cisalpino D, Souza
DG and Teixeira MM: Therapeutic opportunities in dengue infection.
Drug Devel Res. 72:480–500. 2011. View Article : Google Scholar
|
|
9
|
Perry AK, Chen G, Zheng D, Tang H and
Cheng G: The host type I interferon response to viral and bacterial
infections. Cell Res. 15:407–422. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Stetson DB and Medzhitov R: Type I
interferons in host defense. Immunity. 25:373–381. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fleming SB: Viral inhibition of the
IFN-induced JAK/STAT signalling pathway: Development of live
attenuated vaccines by mutation of viral-encoded IFN-antagonists.
Vaccines. 4:pii: E232016. View Article : Google Scholar
|
|
12
|
Schoggins JW, Dorner M, Feulner M, Imanaka
N, Murphy MY, Ploss A and Rice CM: Dengue reporter viruses reveal
viral dynamics in interferon receptor-deficient mice and
sensitivity to interferon effectors in vitro. Proc Natl Acad Sci
USA. 109:14610–14615. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zellweger RM, Prestwood TR and Shresta S:
Enhanced infection of liver sinusoidal endothelial cells in a mouse
model of antibody-induced severe dengue disease. Cell Host Microbe.
7:128–139. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zust R, Toh YX, Valdes I, Cerny D,
Heinrich J, Hermida L, Marcos E, Guillén G, Kalinke U, Shi PY, et
al: Type I interferon signals in macrophages and dendritic cells
control dengue virus infection: Implications for a new mouse model
to test dengue vaccines. J Virol. 88:7276–7285. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sarathy VV, White M, Li L, Gorder SR,
Pyles RB, Campbell GA, Milligan GN, Bourne N and Barrett AD: A
lethal murine infection model for dengue virus 3 in AG129 mice
deficient in type I and II interferon receptors leads to systemic
disease. J Virol. 89:1254–1266. 2015. View Article : Google Scholar :
|
|
16
|
Diamond MS and Harris E: Interferon
inhibits dengue virus infection by preventing translation of viral
RNA through a PKR-independent mechanism. Virology. 289:297–311.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Helbig KJ, Carr JM, Calvert JK, Wati S,
Clarke JN, Eyre NS, Narayana SK, Fiches GN, McCartney EM and Beard
MR: Viperin is induced following dengue virus type-2 (DENV-2)
infection and has anti-viral actions requiring the C-terminal end
of viperin. PLoS Negl Trop Dis. 7:e21782013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hishiki T, Han Q, Arimoto K, Shimotohno K,
Igarashi T, Vasudevan SG, Suzuki Y and Yamamoto N:
Interferon-mediated ISG15 conjugation restricts dengue virus 2
replication. Biochem Biophys Res Commun. 448:95–100. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Siegal FP, Kadowaki N, Shodell M,
Fitzgerald-Bocarsly PA, Shah K, Ho S, Antonenko S and Liu YJ: The
nature of the principal type 1 interferon-producing cells in human
blood. Science. 284:1835–1837. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cella M, Jarrossay D, Facchetti F,
Alebardi O, Nakajima H, Lanzavecchia A and Colonna M: Plasmacytoid
monocytes migrate to inflamed lymph nodes and produce large amounts
of type I interferon. Nat Med. 5:919–923. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lund JM, Alexopoulou L, Sato A, Karow M,
Adams NC, Gale NW, Iwasaki A and Flavell RA: Recognition of
single-stranded RNA viruses by Toll-like receptor 7. Proc Natl Acad
Sci USA. 101:5598–5603. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hemmi H, Takeuchi O, Kawai T, Kaisho T,
Sato S, Sanjo H, Matsumoto M, Hoshino K, Wagner H, Takeda K, et al:
A Toll-like receptor recognizes bacterial DNA. Nature. 408:740–745.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bauer S, Kirschning CJ, Häcker H, Redecke
V, Hausmann S, Akira S, Wagner H and Lipford GB: Human TLR9 confers
responsiveness to bacterial DNA via species-specific CpG motif
recognition. Proc Natl Acad Sci USA. 98:9237–9242. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gilliet M, Cao W and Liu YJ: Plasmacytoid
dendritic cells: Sensing nucleic acids in viral infection and
autoimmune diseases. Nat Rev Immunol. 8:594–606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Perdigon G, Fuller R and Raya R: Lactic
acid bacteria and their effect on the immune system. Current Issues
Intest Microbiol. 2:27–42. 2001.
|
|
26
|
Parcina M, Wendt C, Goetz F, Zawatzky R,
Zähringer U, Heeg K and Bekeredjian-Ding I: Staphylococcus
aureus-induced plasmacytoid dendritic cell activation is based on
an IgG-mediated memory response. J Immunol. 181:3823–3833. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Piccioli D, Sammicheli C, Tavarini S, Nuti
S, Frigimelica E, Manetti AG, Nuccitelli A, Aprea S, Valentini S,
Borgogni E, et al: Human plasmacytoid dendritic cells are
unresponsive to bacterial stimulation and require a novel type of
cooperation with myeloid dendritic cells for maturation. Blood.
113:4232–4239. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jounai K, Ikado K, Sugimura T, Ano Y,
Braun J and Fujiwara D: Spherical lactic acid bacteria activate
plasmacytoid dendritic cells immunomodulatory function via
TLR9-dependent crosstalk with myeloid dendritic cells. PLoS One.
7:e325882012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jounai K, Sugimura T, Ohshio K and
Fujiwara D: Oral administration of Lactococcus lactis subsp lactis
JCM5805 enhances lung immune response resulting in protection from
murine para-influenza virus infection. PLoS One. 10:e01190552015.
View Article : Google Scholar
|
|
30
|
Jounai K, Sugimura T, Morita Y, Ohshio K
and Fujiwara D: Administration of Lactococcus lactis strain Plasma
induces maturation of plasmacytoid dendritic cells and protection
from rotavirus infection in suckling mice. Int Immunopharmacol.
56:205–211. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Castillo Ramirez JA and Urcuqui-Inchima S:
Dengue virus control of type I IFN responses: A history of
manipulation and control. J Interferon Cytokine Res. 35:421–430.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods.
25:402–408. 2001. View Article : Google Scholar
|
|
33
|
St John AL, Rathore AP, Raghavan B, Ng ML
and Abraham SN: Contributions of mast cells and vasoactive
products, leukotrienes and chymase, to dengue virus-induced
vascular leakage. Elife. 2:e004812013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
da Costa VG, Marques-Silva AC and Moreli
ML: A meta-analysis of the diagnostic accuracy of two commercial
NS1 antigen ELISA tests for early dengue virus detection. PLoS One.
9:e946552014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Brawand P, Fitzpatrick DR, Greenfield BW,
Brasel K, Maliszewski CR and De Smedt T: Murine plasmacytoid
pre-dendritic cells generated from Flt3 ligand-supplemented bone
marrow cultures are immature APCs. J Immunol. 169:6711–6719. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Raut CG, Deolankar RP, Kolhapure RM and
Goverdhan MK: Susceptibility of laboratory-bred rodents to the
experimental infection with dengue virus type 2. Acta Virol.
40:143–146. 1996.PubMed/NCBI
|
|
37
|
Johnson AJ and Roehrig JT: New mouse model
for dengue virus vaccine testing. J Virol. 73:783–786. 1999.
|
|
38
|
Orozco S, Schmid MA, Parameswaran P,
Lachica R, Henn MR, Beatty R and Harris E: Characterization of a
model of lethal dengue virus 2 infection in C57BL/6 mice deficient
in the alpha/beta interferon receptor. J Gen Virol. 93:2152–2157.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zellweger RM and Shresta S: Mouse models
to study dengue virus immunology and pathogenesis. Front Immunol.
5:1512014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Syenina A, Jagaraj CJ, Aman SA, Sridharan
A and St John AL: Dengue vascular leakage is augmented by mast cell
degranulation mediated by immunoglobulin Fcγ receptors. Elife. 4:
View Article : Google Scholar : 2015.
|
|
41
|
Gubler DJ: Dengue and dengue hemorrhagic
fever. Clin Microbiol Rev. 11:480–496. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Green S and Rothman A: Immunopathological
mechanisms in dengue and dengue hemorrhagic fever. Curr Opin Infect
Dis. 19:429–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Srikiatkhachorn A: Plasma leakage in
dengue haemorrhagic fever. Thromb Haemost. 102:1042–1049. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chaturvedi UC, Agarwal R, Elbishbishi EA
and Mustafa AS: Cytokine cascade in dengue hemorrhagic fever:
Implications for pathogenesis. FEMS Immunol Med Microbiol.
28:183–188. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Verhelst J, Hulpiau P and Saelens X: Mx
proteins: Antiviral gatekeepers that restrain the uninvited.
Microbiol Mol Biol Rev. 77:551–566. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fitzgerald KA: The interferon inducible
gene: Viperin. J Interferon Cytokine Res. 31:131–135. 2011.
View Article : Google Scholar :
|
|
47
|
Zhao C, Collins MN, Hsiang TY and Krug RM:
Interferon- induced ISG15 pathway: An ongoing virus-host battle.
Trends Microbiol. 21:181–186. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Elkhateeb E, Tag-El-Din-Hassan HT, Sasaki
N, Torigoe D, Morimatsu M and Agui T: The role of mouse
2′,5′-oligoadenylate synthetase 1 paralogs. Infect Genet Evol.
45:393–401. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Takeda S, Takeshita M, Kikuchi Y, Dashnyam
B, Kawahara S, Yoshida H, Watanabe W, Muguruma M and Kurokawa M:
Efficacy of oral administration of heat-killed probiotics from
Mongolian dairy products against influenza infection in mice:
Alleviation of influenza infection by its immunomodulatory activity
through intestinal immunity. Int Immunopharmacol. 11:1976–1983.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fujiwara D, Inoue S, Wakabayashi H and
Fujii T: The anti-allergic effects of lactic acid bacteria are
strain dependent and mediated by effects on both Th1/Th2 cytokine
expression and balance. Int Arch Allergy Immunol. 135:205–215.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wakabayashi H, Nariai C, Takemura F, Nakao
W and Fujiwara D: Dietary supplementation with lactic acid bacteria
attenuates the development of atopic-dermatitis-like skin lesions
in NC/Nga mice in a strain-dependent manner. Int Arch Allergy
Immunol. 145:141–151. 2008. View Article : Google Scholar
|