Introduction
Keloid formation results from an abnormal wound
healing process that is characterized by excessive extracellular
matrix (ECM) deposition and dermal fibroblast hyperproliferation
(1). Even though several
therapeutic options are available at present, the frustrating
clinical prognosis and the high recurrence rates require solutions,
to improve clinical outcomes (2,3).
Keloids are considered to be benign skin tumors due to the similar
characteristics between keloid fibroblasts (KFs) and tumor cells
(4). Keloids have exhibited a
series of tumor-like pathological features, such as increasing ATP
synthesis, inflammation-induced alteration of the stem cell niche
and histological heteromorphism, in which normal dermal tissue
structures disappear (5-7). Due to the similarities between
keloids and benign skin tumors, a large number of anti-keloid
compounds have been investigated for their therapeutic values from
the perspective of anti-tumor like effects on keloid formation
(1,8-12).
The key pathogenetic events that occur in dermal
fibroblasts during post-trauma keloid formation remain unclear.
Activation of phosphoinositide 3-kinase/RAC-alpha
serine/threonine-protein kinase/mammalian target of rapamycin
(PI3K/Akt/mTOR) pathway was demonstrated to enhance inflammation,
angiogenesis, and deposition of ECM components in keloids (13) and was therefore suggested to be
associated with several fibrous disorders (14). Syed et al (8) described overactivation of mTOR in
keloids compared with corresponding extra-lesional regions.
Inhibitors targeting mTOR have exhibited their potentials in
anti-keloid therapy (11).
Histone deacetylases 2 (HDAC2), a second verified factor that may
participate in the progression of fibrotic development, but no
other type of HDAC, has been confirmed to be overexpressed in
keloid and hypertrophic scars (15). As HDAC inhibitors have been widely
exploited for their anti-fibrosis effects on liver/kidney fibrosis
(16) and their anti-cancer roles
(17), they may also represent
promising therapeutic drugs for keloid disease (9).
Therefore, CUDC-907, a newly-synthesized small
molecular compound that synergistically inhibited both
PI3K/Akt/mTOR pathway and HDACs (18) was selected for examination in the
present study for its potential therapeutic effect on KFs, as it
was previously demonstrated that CUDC-907 exerted a potent
anti-tumor effect in B cell lymphoma cell lines (19) and other tumor types (18). Recently, a phase-I clinical trial
(trial no. NCT01742988) examining the biosafety, tolerability, and
preliminary activity of CUDC-907 demonstrated its potential in
future clinical applications (20).
Compared with normal dermal fibroblasts, KFs exhibit
abnormal cellular behaviors including enhanced cell proliferation,
overproduction and deposition of extracellular matrices like
collagens and growth factors such as transforming growth factor-β
(TGF-β) (21). The present study
investigated the anti-keloid effect of CUDC-907 using in
vitro cell cultures, ex vivo explant cultures and an
in vivo xenograft model, and explored if CUDC-907 was able
to reverse the pathological features of in vitro cultured
KFs and counteract keloid formation ex vivo and in
vivo.
Materials and methods
Tissue samples acquisition
A total of 47 patients without any previous
anti-keloid therapy donated their keloid samples following surgical
excision. Patients were informed that their donated tissues would
be utilized only for scientific research and written informed
consent was provided. These patients included 21 males and 26
females, with ages ranging from 7-72 years old. Patient recruitment
was executed from September 2016 to August 2018 in Shanghai Ninth
People's Hospital, Shanghai Jiao Tong University School of
Medicine. Foreskin samples, which were considered as normal skin
samples, were donated by other 4 male patients during this
recruitment. All patient information is provided in Table SI. Equal amounts of primary KFs
isolated from 3 different patients were mixed as a pooled cell
sample to decrease individual variation among different patients.
Protocols for the handling of human tissues and cells were approved
by the Ethics Committee of Shanghai Ninth People's Hospital.
Chemical reagent preparation
CUDC-907, GDC-0941 and Trichostatin A solid powder
were purchased from Selleck Chemicals and dissolved in dimethyl
sulfoxide (DMSO). They were diluted in Dulbecco's modified eagle
medium (DMEM; Hyclone; GE Healthcare Life Sciences) to produce the
designated drug concentrations with DMSO volumes identical to that
of vehicle control group. The final concentration of DMSO solvent
was controlled under 0.00032% (v/v) during each of the subsequent
assays.
Isolation and culture of keloid and
normal dermal fibroblasts
Isolation and culture of fibroblasts were performed
according to a previously described protocol (12). Keloids or foreskin were treated
with 0.3% (for keloid) or 0.2% (for foreskin) collagenase (SERVA
Electrophoresis GmbH) dissolved in DMEM containing 10% fetal bovine
serum (FBS; Hyclone; GE Healthcare Life Sciences) for 6 h at 37°C
on a shaker. Following digestion, cells were collected and
resuspended in DMEM supplemented with 10% FBS and 100X
Antibiotic-Antimycotic solution (Gibco; Thermo Fisher Scientific,
Inc.). The cells from the secondary culture were used in the
following protocols.
Cell proliferation analysis
The KFs from the secondary culture were seeded onto
96-well plates at a density of 1,000 cells/well. Starved cells were
treated with medium containing the CUDC-907, GDC-0941 and
trichostatin A. Cell proliferation was examined using Cell Counting
Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.) according to
the manufacturer's protocol.
Cell cycle analysis
According to a previously described protocol
(12), KFs treated with CUDC-907
for 5 days were collected. To examine cell cycle profiles, the
washed cells were subsequently fixed in ice-cold 70% ethanol for 24
h. After centrifugation at 37°C and 500 x g for 5 min, cell pellets
were then stained using a cell cycle/apoptosis examination kit
(Shanghai Qihai Futai Biotechnology Co., Ltd.). A flow cytometer
(Beckman Coulter, Inc.) equipped with ModiFit LT v2.0 software was
applied to conduct the flow cytometric analyses.
RNA extraction and reverse transcription
quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol®
reagent (Thermo Fisher Scientific, Inc.) and quantitated using a
DU800 spectrophotometer (Beckman Coulter, Inc.). cDNA was
synthesized using AMV reverse transcriptase (Promega Corporation)
with 1.5 μg total RNA according to the manufacturer's
protocol. qPCR was conducted with Power SYBR Green PCR (2X) master
mix (Applied Biosystems; Thermo Fisher Scientific, Inc.) in a
real-time thermal cycler (Stratagene; Agilent Technologies, Inc.)
according to a previously described protocol (22). Thermocycling conditions used in
present study were as follows: 95°C for 10 min, followed by 40
cycles of 95̊C for 30 sec, 58°C annealing temperature for 30 sec
and 72°C for 45 sec. GAPDH was employed as an internal control. The
primers for this assay are listed in Table SII.
In vitro cell migration and invasion
assays
The scratch assay was performed according to a
previously described protocol (12). The cell monolayer was scratched
using a 200 μl pipette tip and then cultured in a serum-free
medium without or with CUDC-907 for 48 h. Images were captured at
0, 24 and 48 h after scratching, and migration distances were
measured using Image-Pro Plus v6.0 software (Media Cybernetics,
Inc.).
Subsequently, a Transwell system (Corning, Inc.) was
utilized according to a previously described protocol (12). Starved KFs were seeded in the
upper chambers at densities of 3×104 or
6×104/well for migration or invasion assays,
respectively. In the invasion assay, chambers were pre-coated in 80
μl Matrigel (BD Biosciences) at 37°C for 30 min prior to
cell seeding. Traversing cells were fixed in 4% paraformaldehyde
for 24 h and then stained with DAPI (Sigma-Aldrich; Merck KGaA).
The cell numbers in 5 randomly selected fields were counted using
Image-Pro Plus v6.0 software.
Further, an Oris™ cell migration and invasion assay
kit (Platypus Technologies, LLC) were employed as the previously
described protocol (11).
Briefly, 2.5×104 starved KFs were seeded in each well.
In the invasion assay, 40 μl rat tail type I collagen (COL1)
solution was used to pre-coat the inserts and incubated at 37°C for
30 min. At 0 h (for background control), 24 h (for migration) or 48
h (for invasion), cells in the migration zone were stained using
Calcein-AM (2 μM; AAT Bioquest, Inc.) at 37°C for 30 min and
were counted using Image-Pro Plus v6.0 software.
Western blot analysis
Total proteins were extracted from KFs, which had
been treated with or without CUDC-907 for 16 h for the cell cycle
molecules analysis and 72 h for the other experiments, using
radioimmunoprecipitation assay lysis buffer with 1% PMSF and 1%
phosphatase inhibitor (Beyotime Institute of Biotechnology). Cell
lysates were resolved on 10% SDS-PAGE gels. A mass of 40 μg
of protein was loaded in each lane for electrophoresis and then
transferred to PVDF membranes. After transferring, membranes were
blocked using 5% nonfat milk (Bio-Rad Laboratories, Inc.) at 37°C
for 1 h. Immunoblotting was performed according to previously
published procedures (12). The
membranes were incubated with primary antibodies and subsequently
incubated with secondary antibodies conjugated with horseradish
peroxidase. All details regarding the antibodies used are
summarized in Table SIII. An
enhanced chemiluminescence detection system was applied to
visualize the protein bands. ImageJ (version 1.8.0; National
Institutes of Health) was used to perform the densitometric
analysis.
Ex vivo explant culture of keloid
tissue
The keloid tissue isolated following epidermal
removal was minced into 3×2×2 mm3 fragments. A total of
20 small pieces were assigned into each group randomly and seeded
onto a culture dish with DMEM (10% FBS) and CUDC-907.
Representative micrographs were captured in the same location for
each culture group, and KFs were collected for counting at day 7
after the treatment, as previously described (12).
Hematoxylin and eosin staining
Tissue samples were fixed in 4% paraformaldehyde at
4°C overnight, embedded in paraffin blocks, and then sectioned to a
5 μm thickness. After complete removal of paraffin, these
sections were stained with a hematoxylin and eosin staining kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's instructions. Staining procedures were conducted at
room temperature. Images were captured via light microscopy at ×200
magnification.
Immunohistochemical staining
This experiment was performed according to a
previously published protocol (12). The primary and secondary
antibodies (Table SIII) were
diluted according to the manufacturer's protocol and target
antigens were visu-alized with a DAB horseradish peroxidase color
development kit (Beyotime Institute of Biotechnology), according to
the manufacturer's instructions, or with the GTVision™ III
detection system/Mo&Rb (Dako; Agilent Technologies, Inc.).
Images were captured through a light microscopy at ×100 or ×200
magnification. Fold changes in integral optical density were
measured using Image-Pro Plus 6.0 and platelet endothelial cell
adhesion molecule-positive (CD31+) and hematopoietic
progenitor cell antigen-positive (CD34+) blood vessel
numbers were counted in 5 randomly selected fields.
In vitro tube formation assay
Human umbilical vein endo-thelial cells (HUVECs),
kindly provided by Shanghai Key Laboratory of Tissue Engineering,
were seeded in each well of a 96 well plate (1.5×104 per
well) where 50 μl Matrigel had been pre-incubated at 37°C
for 1 h. After incubation at 37°C for 16 h, images of newly formed
capillary-like structures were observed via a phase-contrast light
microscope at ×40 magnification and semi-quantitatively analyzed as
previously described (23).
Keloid xenograft model in nude mice
According to a previously published protocol
(24), a total of 12 male nude
mice (6-week-old; 16.00±1.48 g) purchased from Shanghai Laboratory
Animal Centre, Chinese Academy of Sciences were used. The mice were
kept in a specific pathogen-free environment where the temperature
was maintained at 25̊C and humidity was 40-70%, with a light: Dark
cycle of 12: 12 h. Animals were kept in their cages with ad
libitum access to food and water. Anesthesia was induced via
inhalation of 2% isoflurane (0.8 l/min) in air for 30 sec and
maintained using 1.5% isoflurane (0.6 l/min) in air (25-27). Anesthesia was considered
successful when the righting reflex was absent or the mice did not
respond to toe pinches. All surgeries were conducted under aseptic
conditions following the successful induction of anesthesia and all
protocols were approved by the Animal Care and Welfare Committee of
Shanghai Ninth People's Hospital. Epidermis-intact cylindrical
specimens (diameter =8 mm; height=3 mm) were attached to the backs
of the animals, then sutured with 6-0 PDS (Johnson & Johnson).
At 12 days post-surgery, 10 μl normal saline solution
containing 64 nM CUDC-907 or vehicle were injected into keloid
xenografts every 2 days. Animals were sacrificed by cervical
dislocation at the 10th day or the 20th day of drug administration
and the xenografts were harvested for further histological and
immunohistological assessments as aforementioned.
Statistical analysis
All experiments were repeated at least 3 times. All
the data are presented as mean ± standard deviation (SD) or
standard error of the mean (SEM). A one-way analysis of variance
was used to detect significant difference among multiple groups,
followed by a Least Significant Difference post-hoc test using SPSS
v22.0 software (IBM Corp). P<0.05 was considered to indicate a
statistically significant difference.
Results
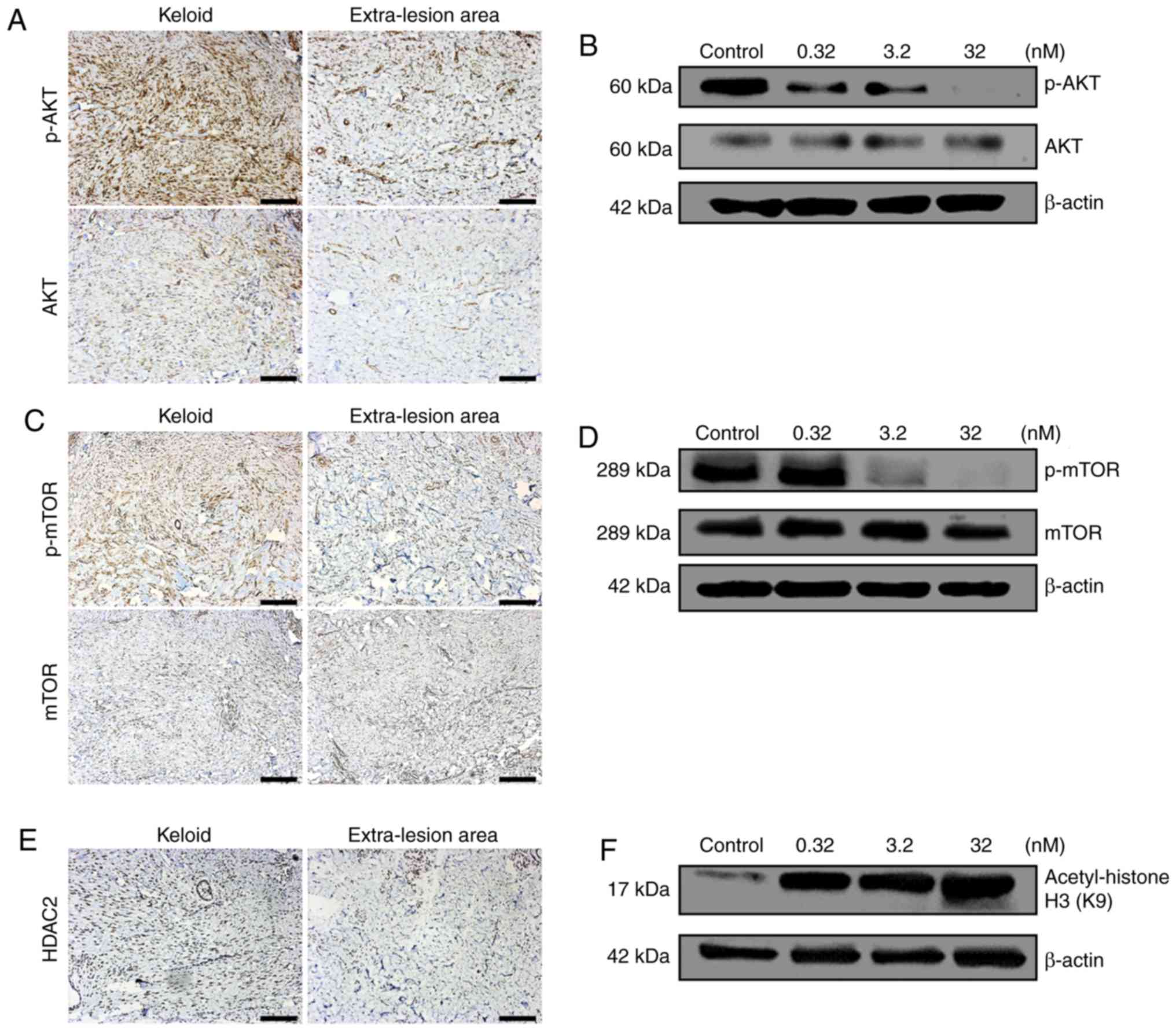
Keloids expresses increased levels of
HDAC2, total and p-Akt and mTOR
Immunohistochemical staining demonstrated that both
total and p-Akt and mTOR were visually identified to be expressed
at increased levels in keloid tissue compared with in the
extra-lesion areas (Fig. 1A and
C). In addition, HDAC2 was also identified visually to be
expressed in keloid tissues at an increased level compared with
that of the extra-lesion area (Fig.
1E).
CUDC-907 inhibits the PI3K/Akt/mTOR
signaling pathway and promotes acetylation of histone H3
Following the retreatment with CUDC-907 for 72 h,
Akt phosphorylation was significantly inhibited, particularly at
the concentrations of 3.2 and 32 nM (Fig. 1B). Similarly, phosphorylation of
mTOR was also significantly inhibited at a concentration of 32 nM
(Fig. 1D). With the increased
concentrations of CUDC-907, the acetylation level of histone H3
gradually increased with a marked dose-dependent pattern (Fig. 1F).
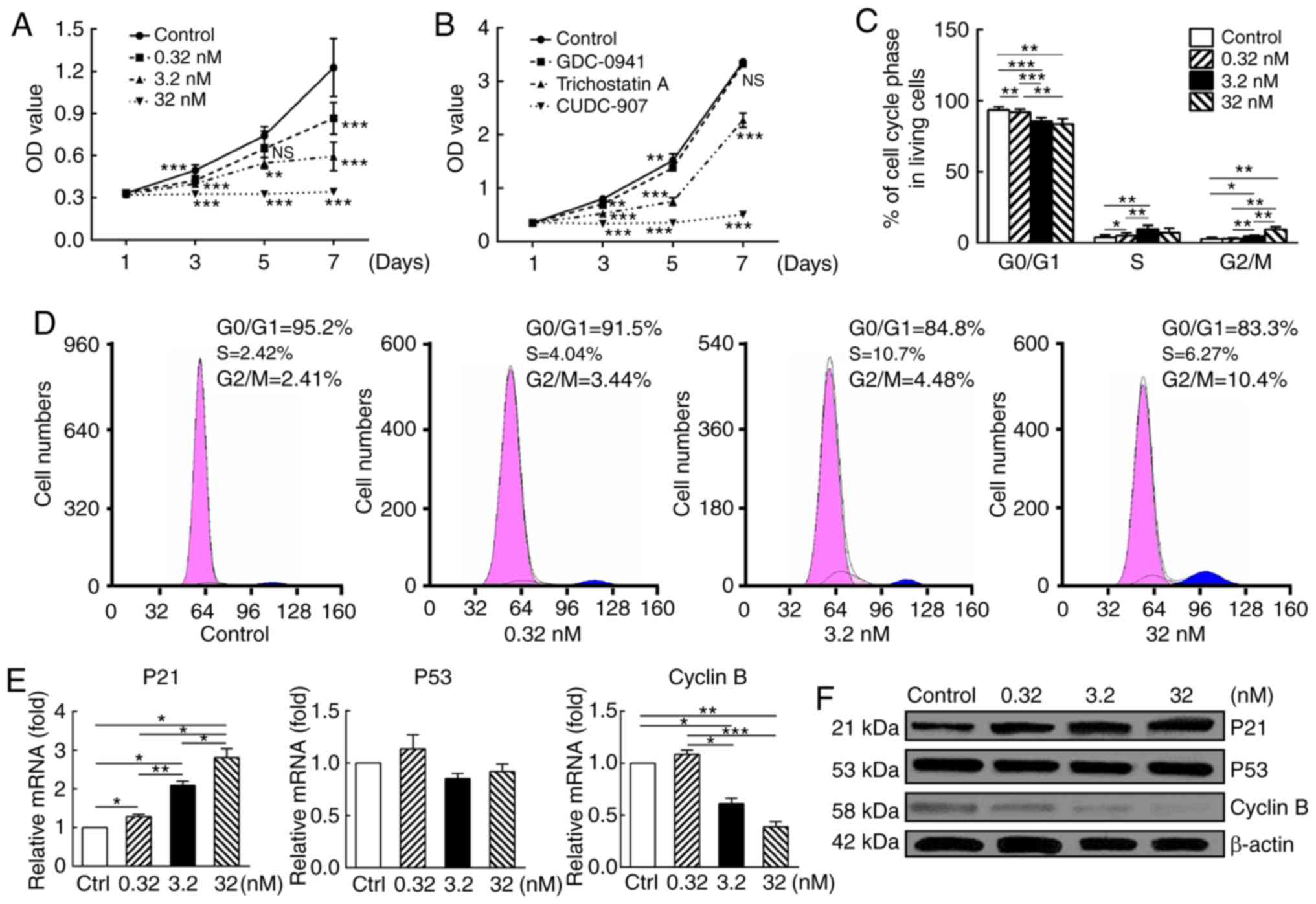
CUDC-907 suppresses KF proliferation and
induces G2-M cell cycle arrest
CUDC-907 significantly decreased the proliferation
of KFs in a dose-dependent manner (Fig. 2A). In addition, the dual inhibitor
CUDC-907 was markedly more potent compared with GDC-0941, a
PI3K/Akt/mTOR inhibitor, and trichostatin A, a HDAC inhibitor,
following drug treatment for 7 days (Fig. 2B).
Cell cycle analysis suggested marked G2/M arrest,
which was expressed as a dose-dependent decrease in the percentage
of G0/G1 phase cells and a corresponding dose-dependent increase in
the percentage of cells in G2/M phase (P<0.05; Fig. 2C and D).
Results of qPCR and western blot analysis further
validated this observation (Fig. 2E
and F). At both mRNA and protein levels, enhanced expression of
cyclin-dependent kinase inhibitor 1 (p21) (P<0.05), steadily
expressed p53 (P>0.05) and attenuated cyclin B expression
(P<0.05) were observed following drug treatment with a
dose-response pattern.
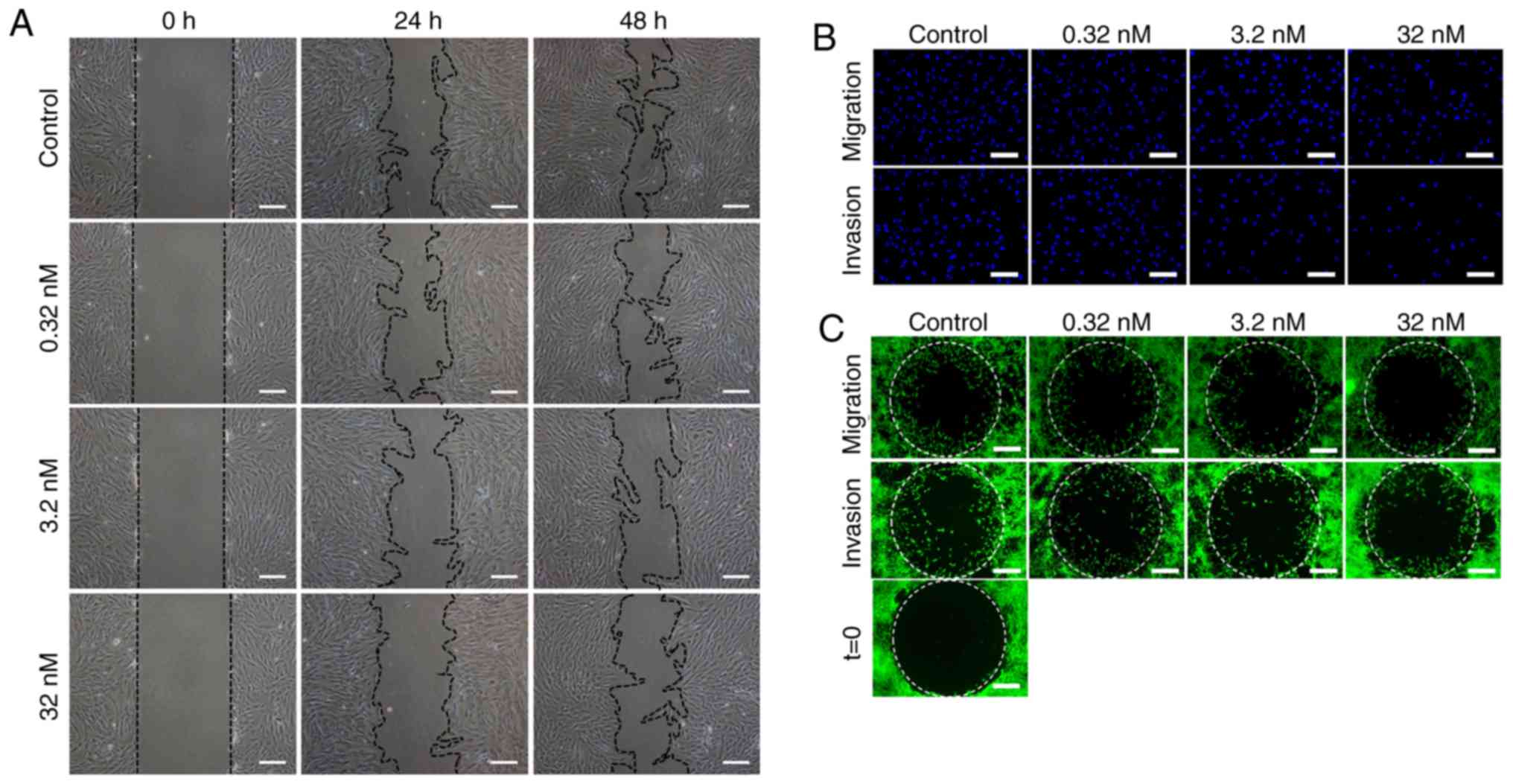
CUDC-907 inhibits migratory capacity of
KFs
At both 24 and 48 h after the scratch assay, a
significant dose-dependent inhibitory effect on migration was
observed (P<0.05: Fig. 3A and
D). In the Transwell migration assay, after 24 h of drug
treatment, the number of migrated cells on the bottom surface of
the top chamber was significantly decreased with the increasing
drug concentrations, in a dose-dependent manner (Fig. 3B). Semi-quantitative analysis
indicated decreased cell numbers with significant differences
between treatment groups (P<0.05; Fig. 3E).
In addition, as demonstrated in Fig. 3C, decreased numbers of the cells
in the migration zone were observed following increasing
concentrations of CUDC-907, in a dose-dependent pattern after 24 h
drug treatment (P<0.05; Fig. 3C
and F).
CUDC-907 inhibits the invasive
capabilities of KFs
The aforementioned Transwell system was also used to
measure the invasive capabilities of KFs. As indicated in Fig. 3B and E, KFs in the control group
successfully passed through the Matrigel to the bottom surface of
the top chamber. Conversely, relatively fewer cells in the CUDC-907
treatment groups successfully invaded through Matrigel (P<0.05).
Analogous results were also observed in an assay conducted with an
Oris™ invasion kit, but with the inclusion of a collagen gel. As
demonstrated in Fig. 3C and F, a
decrease in cell numbers in the detection zone was observed with
the increasing drug concentrations (P<0.05). In addition, matrix
metalloproteinases (MMPs) and tissue inhibitor of
metalloproteinases (TIMPs) serve an important role in invasion
ability of tumor cells (28).
Decreased MMP/TIMP1 gene expression ratios, concomitant with
increased drug concentrations, were observed in MMP1/TIMP1,
MMP2/TIMP1, MMP3/TIMP1 and MMP13/TIMP1 (P<0.05; Fig. 3G).
CUDC-907 attenuates collagen expression
and production of cultured KFs in vitro and keloid tissue ex
vivo
As demonstrated in Fig. 4A, at an mRNA level, CUDC-907
treatment significantly downregulated the expression of collagen I
(COL1) and collagen III (COL3) (P<0.05) at the concentrations of
3.2 and 32 nM, but not at 0.32 nM (P>0.05). Expression levels of
these genes in the KFs treated with GDC-0941, a PI3K/Akt/mTOR
inhibitor, or trichostatin A, a HDAC inhibitor, at 32 nM were not
identified to be significantly different from those of control
(P>0.05; Fig. 4B), but were
significantly increased compared with that of the CUDC-907-treated
cells (P<0.05). In contrast to the KFs, the normal dermal
fibroblasts were identified to be less sensitive to CUDC-907
treatment, indicating a cell type-specific inhibitory effect of
CUDC-907 (Fig. 4C).
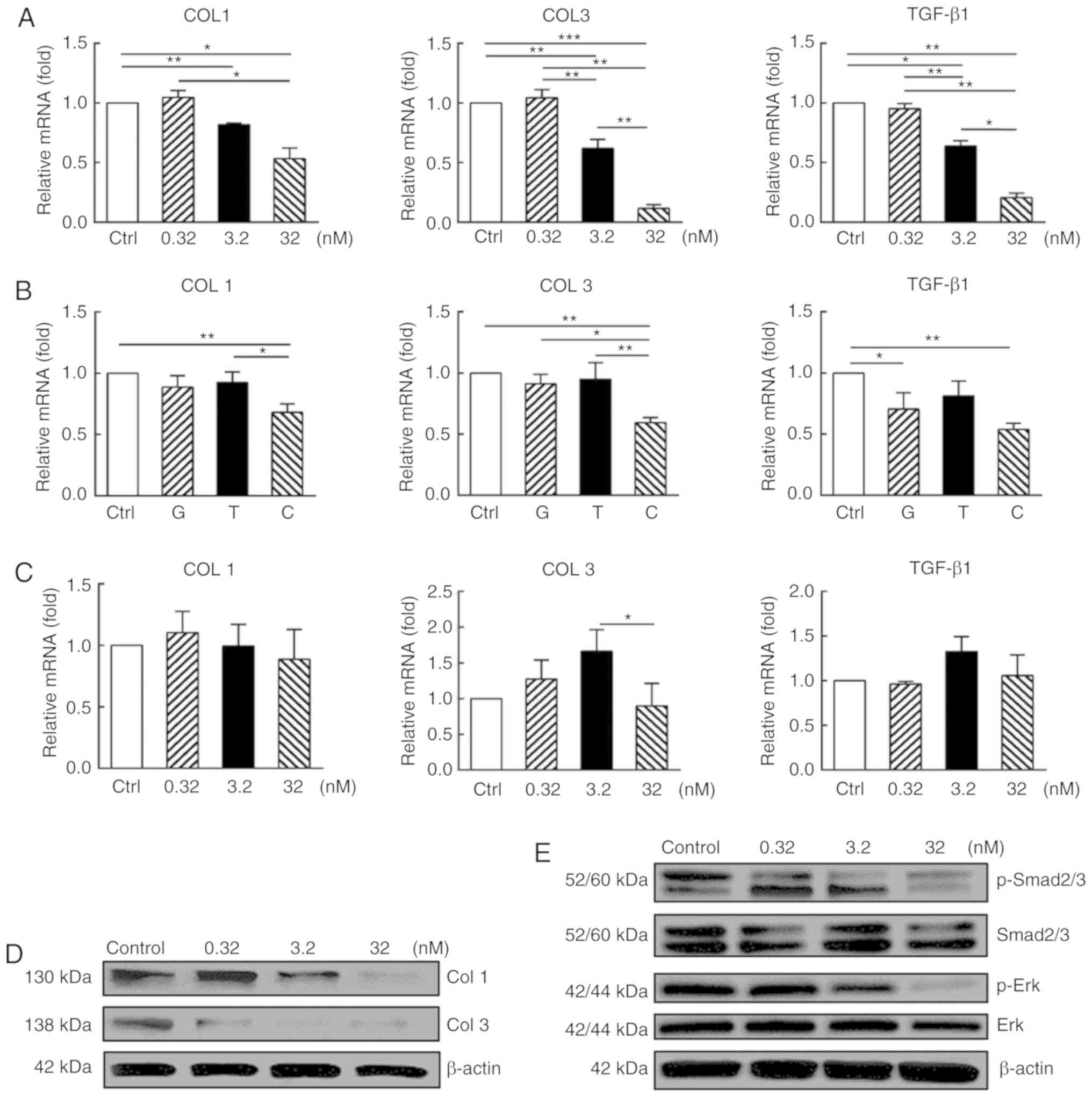 | Figure 4CUDC-907 decreases collagen
expression or production of in vitro cultured keloid
fibroblasts along with attenuated Smad and Erk phosphorylation. (A)
After 72 h CUDC-907 treatment, KFs expressed decreased levels of
COL1, COL3 and TGF-β1 at transcriptional level with significant
differences among the treatment groups. (B) Inhibitory effects of
GDC-0941, trichostatin A and CUDC-907 on the gene expression levels
of COL1, COL3 and TGF-β1 at 32 nM for 72 h. (C) No significant
inhibitory effect of CUDC-907 on COL1, COL3 and TGF-β1 gene
expression levels of normal dermal fibroblasts was observed after
72 h from the treatment. (D) CUDC-907 treatment decreased the
production of COL1 and COL3 at 72 h post-treatment, as demonstrated
by western blot analysis. (E) CUDC-907 also attenuated the
phosphorylation levels of Smad 2/3 and Erk of the keloid
fibroblasts treated with various concentrations of CUDC-907 at 72 h
post-treatment. The assays were repeated 3 times with 3
independently pooled cell samples. Data are presented as the mean ±
standard error of the mean. *P<0.05,
**P<0.01 and ***P<0.001. COL1, collagen
I; COL3, collagen III; TGF-β1, transforming growth factor β1; p,
phosphorylated; G, GDC-0941; T, trichostatin A; C, CUDC-907. |
Concomitantly, the results from the western blot
analysis also suggested significantly decreased protein production
of COL1 and COL3 following CUDC-907 treatment (Fig. 4D). Furthermore,
immunohistochemical staining of the keloid tissue explant also
exhibited decreased COL1 and COL3 production, due to the treatment
of CUDC-907 (Fig. 5A),
particularly at 32 nM (P<0.05; Fig. 5B).
CUDC-907 attenuates TGF-β1 production and
signaling
At the mRNA level, CUDC-907 treatment for 72 h
significantly downregulated the expression of TGF-β1 at the
concentrations of 3.2 and 32 nM (P<0.05) but not at 0.32 nM
(P>0.05) (Fig 4A). Western
blot analysis indicated that CUDC-907 treatments at 3.2 and 32 nM
significantly inhibited the phosphorylation of Smad2/3, whereas the
reduction of Erk phosphorylation was only observed at the
concentration of 32 nM, as demonstrated in Fig. 4E.
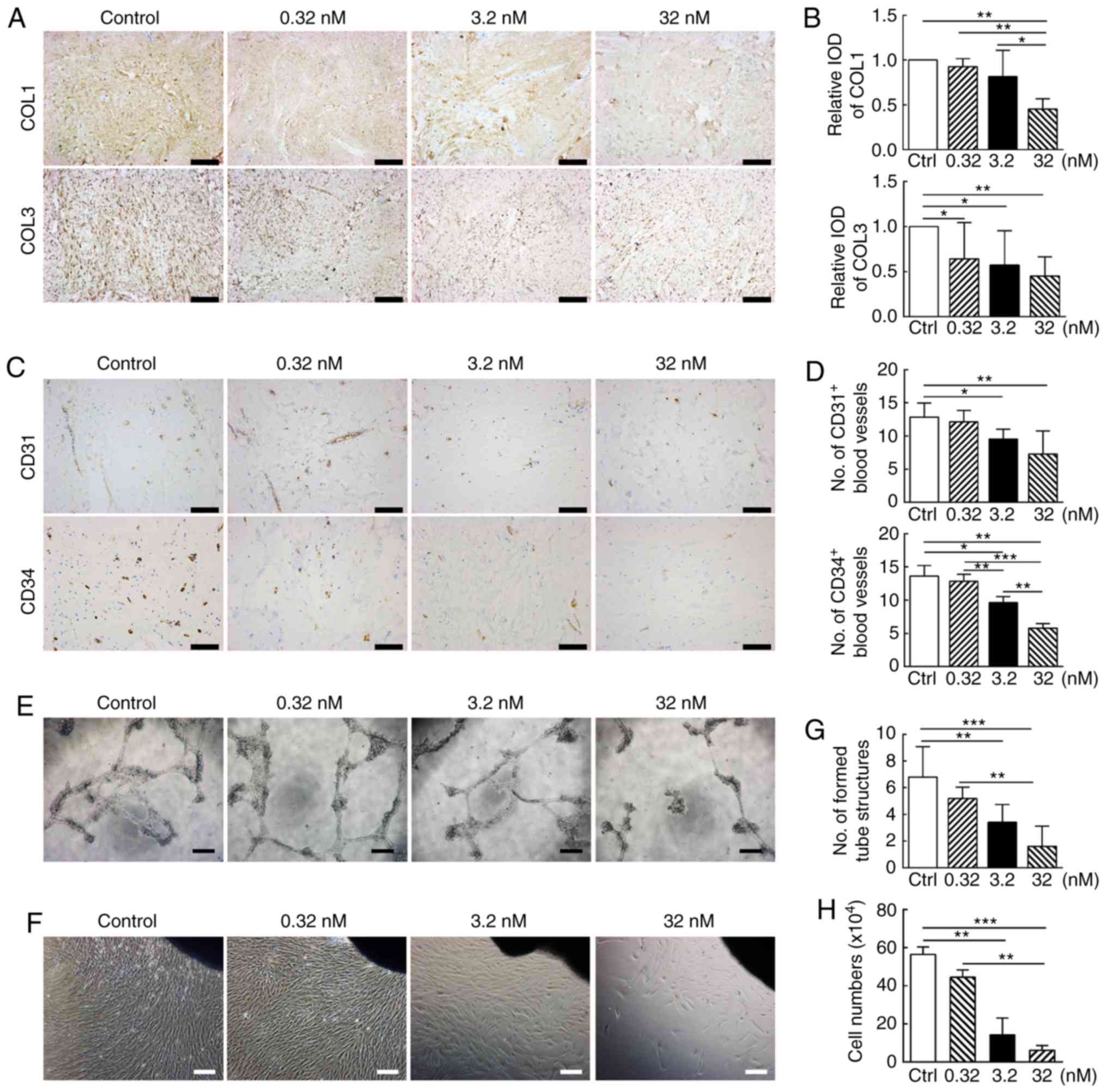
CUDC-907 inhibits angiogenesis in keloid
tissue explant ex vivo
Excessive vascularization following trauma is
another important pathogenic factor underlying keloid formation
(29). As indicated in Fig. 5C, the number of blood vessels
marked with CD31 and CD34, two common immunohistochemical markers
of endothelial cells (30),
gradually decreased as the concentration of CUDC-907 increased
(P<0.05; Fig. 5D). As a
confirmation of this observation, CUDC-907 treatment significantly
disrupted vessel tube formation mediated by HUVECs in vitro
(Fig. 5E and G).
CUDC-907 suppresses KF migration and
proliferation in cultured keloid explants ex vivo
As demonstrated in Figs. 5F and S1, KFs with a spindle shape migrated
out from the edges of the explanted tissue and gradually filled the
Petri dish over 1 week via cell migration and proliferation.
CUDC-907 treatment for 7 days significantly decreased the number of
cells migrating from the keloid explants. The inhibitory effect
became more significant with the increasing drug concentrations
(P<0.05; Fig. 5H).
CUDC-907 attenuates collagen deposition
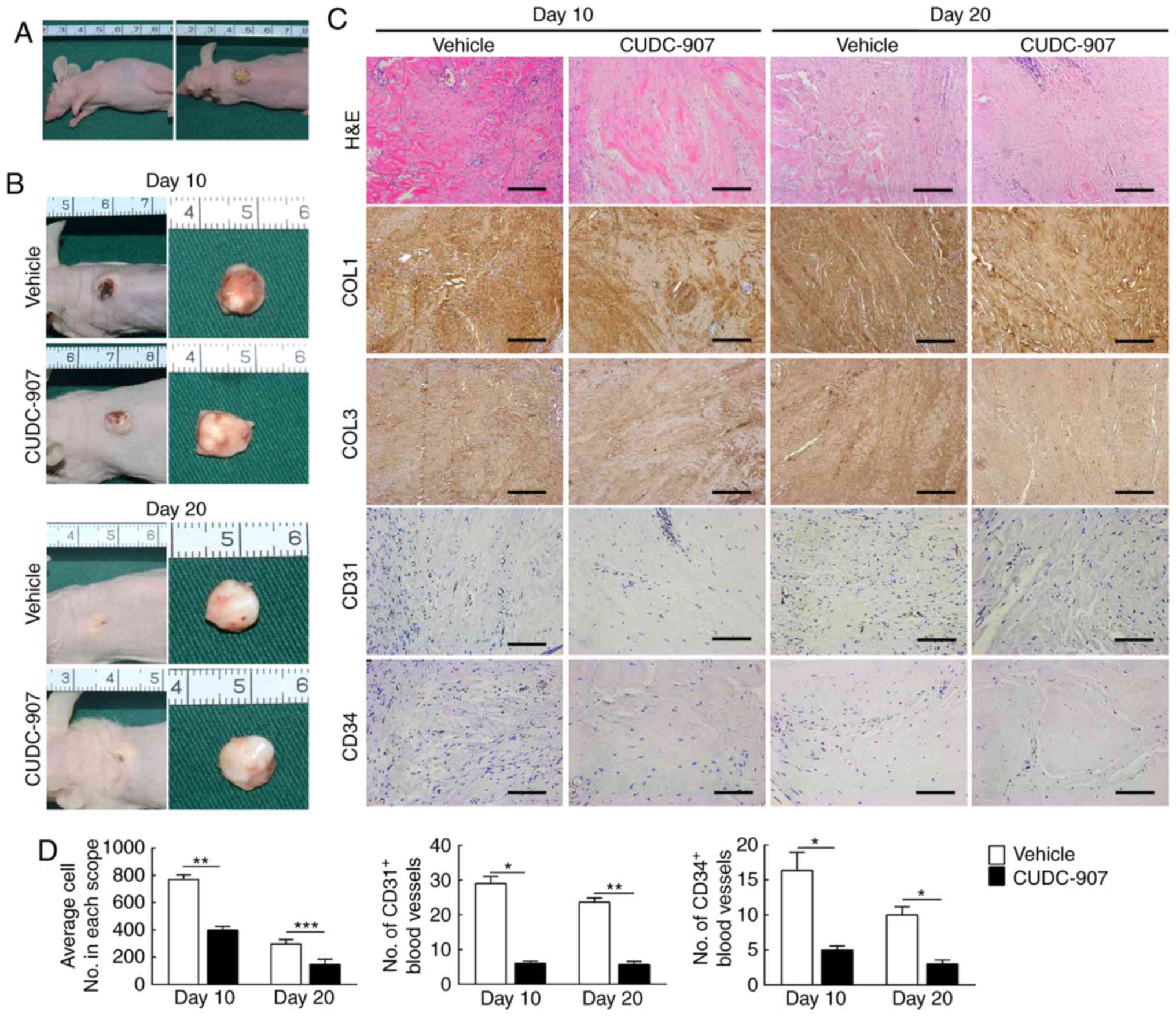
and angiogenesis in a keloid xenograft model
At the 10th and the 20th day of CUDC-907
administration, xenografts on the backs of nude mice had not
decreased nor increased in size consistently (Fig. 6A and B). However, at the
histological level, decreased cell density, COL1 and COL3
deposition, CD31+ and CD34+ blood vessels
were observed in the keloid xenograft by hematoxylin and eosin or
immunohistochemical staining, which suggested that CUDC-907 was
able to exert its anti-fibrosis and anti-angiogenesis effects in
vivo (Fig. 6C and D).
Discussion
Keloid is a tumor-like skin disorder unique to
humans, which is commonly observed in those of African and Asian
descent, with neoplasms growing beyond the wound boundary but never
metastasizing to distant sites (31). Different from normal scars, an
excessive inflammatory response, unrestricted angiogenesis and
collagen accumulation are present during the whole process during
keloid formation, which may lead to intolerable pruritus, pain and
even disfigurement (21). Due to
the high recurrence rate following keloid excision, a classic
anti-keloid drug, 5-Fluorouracil, was generally utilized as an
auxiliary chemotherapy after surgical procedures (32). Although a diverse range of small
molecule compounds have been developed in order to improve the
clinical outcome of keloid therapy, their low efficacy and multiple
adverse side effects demand more advanced solutions and the
identification of a compound that precisely targets the vital
elements of keloid growth (33).
The occurrence of keloids cannot be attributed to
any known single predisposing factor. In fact, ethnicity or genetic
background, immune dysfunction, abnormal fibroblasts and other
extrinsic stimuli like tension and infection may all be associated
with keloid formation (31,34). Among multiple pathological
elements, abnormality of KFs serves a significant role in the
pathogenesis, including hyperproliferation, aberrant capacity of
migration and invasion, excessive production and deposition of
extracellular matrices (35).
Notably, certain small chemical compounds developed for anti-tumor
treatment were demonstrated to have similar mechanism of action to
keloid chemotherapy (36,37). Studies investigating the
mechanisms of these compounds may also contribute to the
understanding of keloid pathogenesis and subsequently to the
development of anti-keloid compounds.
The involvement of PI3K/AKT/mTOR signaling and HDAC
in the abnormal functions of keloid cells has been previously
described (9,11,14). Single-target inhibitors of either
the PI3K/Akt/mTOR pathway or HDACs would be good drug candidates
for keloid therapy, but their demonstrated low efficacies have
limited their potential in clinical application. Inhibition of the
PI3K pathway alone did not work as well as expected due to adverse
activation of other tumor-associated signaling pathways or
drug-resistance (38,39). Similarly, trichostatin A, a
previously described anti-keloid HDAC inhibitor, effectively
inhibited the proliferation and collagen production of KFs at
relatively higher concentrations (9), leading to increasing potential risk
in its further clinical application. To lower this potential risk,
a feasible solution would be to identify a compound that has
multiple targets and may achieve a satisfactory therapeutic effect
with minimal doses via a synergistic effect across the multiple
targets (40).
It has been demonstrated that drugs that
simultaneously suppressed both HDAC and PI3K exhibited improved
function compared with single HDAC inhibitors for the treatment of
cutaneous T-cell lymphoma or other types of cancer (41,42). In addition, it is difficult for a
hybrid molecule like CUDC-907 to escape from cells following
conversion into pharmacologically active acid form, easily
increasing the levels of intracellular accumulation (43). In the present study, 3
concentrations of 0.32, 3.2 and 32 nM were selected based on
extensive pre-experimental screening and previous data (18). It should be noted that the
concentrations used in the present study were much lower compared
with the single-target inhibitors investigated in previous studies
(8,9,11).
The inhibitory effect of CUDC-907 on the hyperproliferation of KFs
was observed even at the lowest concentration (0.32 nM). At higher
concentrations (3.2 and 32 nM), the drug potently suppressed the
migratory and invasive capabilities of KFs, decreased the excessive
production of collagen, and prevented excessive angiogenesis. In
addition, by comparing the inhibitory effect of CUDC-907 with
GDC-0941 and trichostatin A at 32 nM, it was confirmed that the
CUDC-907-mediated inhibition of cell proliferation and collagen
production in KFs was improved compared with GDC-0941 and
trichostatin A. The present study demonstrated that CUDC-907
exhibited a more potent inhibitory effect compared with either
trichostatin A or GDC-904 alone on cell proliferation and collagen
gene expression, indicating that CUDC-907 exerted the synergistic
effect of dual signaling inhibition and therefore may be a more
potent targeted drug for keloid therapy.
Such phenomena were also described in previous
studies (44). Different
anti-proliferative mechanisms of mTOR or HDAC inhibitors were
demonstrated (11,45-48), but another previous study revealed
that CUDC-907 induced a G2/M cell cycle arrest of H460, a human
tumor cell line (18), indicating
that CUDC-907 may interfere with the cell mitosis process. In fact,
G2/M arrest in other tumor cell lines resulting from the treatment
with several kinds of HDAC inhibitors has been observed (49). Although no significant differences
were detected in p53 expression levels at the concentrations used
in the present study, a significant increase in p21 expression
levels [also identified previously (18)], has been established to be a
definite result of inhibition of either the PI3K/Akt/mTOR or HDACs
pathways (50,51). In addition, inhibitors targeting
any one of these signaling pathways may also downregulate the
levels of cyclin B, resulting in a G2/M arrest (52,53). The HDAC inhibitor valproic acid
downregulated the levels of cyclin B and even demonstrated a
sensibilization effect on rapamycin, a classic mTOR inhibitor,
indicating the synergistic effect between inhibition of the
PI3K/Akt/mTOR pathway and HDACs (53).
The attenuated ability of cell migration and
invasion in KFs is also associated with the inhibition of the PI3K
pathway and HDACs (49,54). To be specific, the HDAC1
inhibitor, largazole, was previously confirmed to be capable of
suppressing the migration of a human microvascular endo-thelial
cell line (55). Bian et
al (56) identified that the
multiple anticancer effects of HDAC inhibitors were achieved
partially by upregulating miR-200c and thereby decreasing Crk-like
protein. However, the vital mechanism has not been fully understood
yet. Furthermore, the drug treatment also altered MMP expression
levels in the present study; measuring the activities of MMP will
provide a more accurate reflection of the cellular functions.
Functional alterations following the abrogation of
exogenous TGF-β1 by inhibition of the PI3K/Akt/mTOR pathway or
HDACs have been investigated previously (57,58), but the present study provided
evidence that the dual inhibition of the PI3K/Akt/mTOR pathway and
HDACs decreased the expression levels of endogenous TGF-β1. The
decrease of gene expression of TGF-β1 and the inhibition of
its downstream cascade may provide an explanation why CUDC-907
exhibited such potent anti-fibrotic capacity (59,60). Although the activation of PI3K/Akt
pathway alone was insufficient to upregulate collagen, type I,
alpha 2 gene transcription, its activation triggered by TGF-β1 may
enhance Smad3 transcriptional activity, leading to increased COL1
expression (36). Decreased
phosphorylation of Erk, which may be activated simultaneously along
with the PI3K pathway in KFs, also provided an explanation of how
CUDC-907 attenuated collagen production (61). A previous study suggested that a
HDAC inhibitor was able to counteract exogenous TGF-β1 and thereby
suppress collagen deposition in KFs (8), which was in concordance with the
results of the present study.
To further investigate the potential effects of
CUDC-907 treatment in a clinical setting, an in vivo
xenograft model was employed in the present study. The implanted
xenograft did not grow following implantation, and therefore no
significant differences in tissue volume were observed. However,
significant differences were identified in matrix deposition and in
microvessel structure integrity. Therefore, the in vivo
model also supports the conclusions of the in vitro
assays.
In summary, the present study confirmed that the
efficient inhibition of both the PI3K/Akt/mTOR pathway and HDACs
activity by CUDC-907 led to significant inhibition of pathological
cell proliferation, collagen production and ECM deposition, and the
cellular migratory and invasive capabilities, in addition to the
angiogenic process of keloids. As all of these factors are the key
contributors to keloid formation, the results of the present study
described the great therapeutic potential of CUDC-907 for keloid
clinical therapy in the future.
Supplementary Data
Acknowledgements
The authors would like to acknowledge the technical
support of Dr Wanyao Xia, Dr Lijuan Zong, Dr Juanjuan Wu and Dr
Junhong Lu from Shanghai Key Laboratory of Tissue Engineering. In
addition, the authors would like to thank Mr. Haibo Li of Xiangya
School of Medicine, Central South University for assistance with
image acquisition.
Abbreviations:
|
PI3K
|
phosphoinositide 3-kinase
|
|
Akt
|
RAC-alpha serine/threonine-protein
kinase
|
|
mTOR
|
mammalian target of rapamycin
|
|
HDAC2
|
histone deacetylases 2
|
|
KFs
|
keloid fibroblasts
|
|
ECM
|
extracellular matrix
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
Funding
The present study was financially supported by China
National Nature Science Foundation (grant no. 81671921).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TT drafted experiment design and conducted the
experiments. JH and ML performed the primary keloid fibroblasts
culture and tissue explant handling. ZG and XW provided tissue
samples and contributed to critical evaluation of samples, and
provided guidance for the handling of the keloid tissues. WZ and GZ
revised the experimental design and provided technical support. WW
and WL are responsible for revising the manuscript critically for
important intellectual content and manuscript editing. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Protocols for the handling of human tissues and
cells were approved by the Ethics Committee of Shanghai Ninth
People's Hospital. All protocols concerning animal experiments were
approved by the Animal Care and Welfare Committee of Shanghai Ninth
People's Hospital. All involved patients provided written informed
consent.
Patient consent for publication
All patients provided written informed consent.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ghazawi FM, Zargham R, Gilardino MS,
Sasseville D and Jafarian F: Insights into the pathophysiology of
hypertrophic scars and keloids: How do they differ? Adv Skin Wound
Care. 31:582–595. 2018. View Article : Google Scholar
|
|
2
|
Morelli Coppola M, Salzillo R, Segreto F
and Persichetti P: Triamcinolone acetonide intralesional injection
for the treatment of keloid scars: Patient selection and
perspectives. Clin Cosmet Investig Dermatol. 11:387–396. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Forbat E, Ali FR and Al-Niaimi F:
Treatment of keloid scars using light-, laser- and energy-based
devices: A contemporary review of the literature. Lasers Med Sci.
32:2145–2154. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Russell SB, Trupin KM, Rodriguez-Eaton S,
Russell JD and Trupin JS: Reduced growth-factor requirement of
keloid-derived fibroblasts may account for tumor growth. Proc Natl
Acad Sci USA. 85:587–591. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vincent AS, Phan TT, Mukhopadhyay A, Lim
HY, Halliwell B and Wong KP: Human skin keloid fibroblasts display
bioener-getics of cancer cells. J Invest Dermatol. 128:702–709.
2008. View Article : Google Scholar
|
|
6
|
Zhang Q, Yamaza T, Kelly AP, Shi S, Wang
S, Brown J, Wang L, French SW, Shi S and Le AD: Tumor-Like stem
cells derived from human keloid are governed by the inflammatory
niche driven by IL-17/IL-6 axis. PLoS One. 4:e77982009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jumper N, Paus R and Bayat A: Functional
histopathology of keloid disease. Histol Histopathol. 30:1033–1057.
2015.PubMed/NCBI
|
|
8
|
Syed F, Sherris D, Paus R, Varmeh S,
Pandolfi PP and Bayat A: Keloid disease can be inhibited by
antagonizing excessive mTOR signaling with a novel dual TORC1/2
inhibitor. Am J Pathol. 181:1642–1658. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Diao J, Xia W, Yi C, Wang Y, Li B, Xia W,
Liu B, Guo S and Sun X: Trichostatin A inhibits collagen synthesis
and induces apoptosis in keloid fibroblasts. Arch Dermatol Res.
303:573–580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yi D, Bihl J, Newman MS, Chen Y and Simman
R: The preliminary study of effects of tolfenamic acid on cell
proliferation, cell apoptosis, and intracellular collagen
deposition in keloid fibroblasts in vitro. Dermatol Res Pract.
2014.1–8. 2014. View Article : Google Scholar
|
|
11
|
Syed F, Sanganee HJ, Bahl A and Bayat A:
Potent dual inhibitors of TORC1 and TORC2 complexes (KU-0063794 and
KU-0068650) demonstrate in vitro and ex vivo Anti-Keloid scar
activity. J Invest Dermatol. 133:1340–1350. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang W, Qu M, Xu L, Wu X, Gao Z, Gu T,
Zhang W, Ding X, Liu W and Chen Y: Sorafenib exerts an anti-keloid
activity by antagonizing TGF-β/Smad and MAPK/ERK signaling
pathways. J Mol Med (Berl). 94:1181–1194. 2016. View Article : Google Scholar
|
|
13
|
Wong VW, You F, Januszyk M, Gurtner GC and
Kuang AA: Transcriptional profiling of rapamycin-treated
fibroblasts from hypertrophic and keloid scars. Ann Plast Surg.
72:711–719. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ong CT, Khoo YT, Mukhopadhyay A, Do DV,
Lim IJ, Aalami O and Phan TT: mTOR as a potential therapeutic
target for treatment of keloids and excessive scars. Exp Dermatol.
16:394–404. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fitzgerald O'Connor EJ, Badshah II, Addae
LY, Kundasamy P, Thanabalasingam S, Abioye D, Soldin M and Shaw TJ:
Histone deacetylase 2 is upregulated in normal and keloid scars. J
Invest Dermatol. 132:1293–1296. 2012. View Article : Google Scholar
|
|
16
|
Van Beneden K, Mannaerts I, Pauwels M, Van
den Branden C and Van Grunsven LA: HDAC inhibitors in experimental
liver and kidney fibrosis. Fibrogen Tissue Repair. 6:12013.
View Article : Google Scholar
|
|
17
|
Miller TA, Witter DJ and Belvedere S:
Histone deacetylase inhibitors. J Med Chem. 46:5097–5116. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qian C, Lai CJ, Bao R, Wang DG, Wang J, Xu
GX, Atoyan R, Qu H, Yin L, Samson M, et al: Cancer network
disruption by a single molecule inhibitor targeting both histone
deacetylase activity and phosphatidylinositol 3-kinase signaling.
Clin Cancer Res. 18:4104–4113. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mondello P, Derenzini E, Asgari Z, Philip
J, Brea EJ, Seshan V, Hendrickson RC, de Stanchina E, Scheinberg DA
and Younes A: Dual inhibition of histone deacetylases and
phosphoinositide 3-kinase enhances therapeutic activity against B
cell lymphoma. Oncotarget. 8:14017–14028. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Younes A, Berdeja JG, Patel MR, Flinn I,
Gerecitano JF, Neelapu SS, Kelly KR, Copeland AR, Akins A, Clancy
MS, et al: Safety, tolerability, and preliminary activity of
CUDC-907, a first-in-class, oral, dual inhibitor of HDAC and PI3K,
in patients with relapsed or refractory lymphoma or multiple
myeloma: An open-label, dose-escalation, phase 1 trial. Lancet
Oncol. 17:622–631. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Seifert O and Mrowietz U: Keloid scarring:
Bench and bedside. Arch Dermatol Res. 301:259–272. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang W, Li J, Wang K, Zhang Z, Zhang W,
Zhou G, Cao Y, Ye M, Zou H and Liu W: Induction of predominant
tenogenic phenotype in human dermal fibroblasts via synergistic
effect of TGF-β and elongated cell shape. Am J Physiol Cell
Physiol. 310:C357–C372. 2016. View Article : Google Scholar
|
|
23
|
Arnaoutova I and Kleinman HK: In vitro
angiogenesis: Endothelial cell tube formation on gelled basement
membrane extract. Nat Protoc. 5:628–635. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ward Kischer C, Sheridan D and Pindur J:
Use of nude (athymic) mice for the study of hypertrophic scars and
keloids: Vascular continuity between mouse and implants. Anat Rec.
225:189–196. 1989. View Article : Google Scholar
|
|
25
|
Ni C, Li C, Dong Y, Guo X, Zhang Y and Xie
Z: Anesthetic isoflurane induces DNA damage through oxidative
stress and p53 pathway. Mol Neurobiol. 54:3591–3605. 2017.
View Article : Google Scholar :
|
|
26
|
Koutsogiannaki S, Zha H and Yuki K:
Volatile anesthetic isoflurane attenuates liver injury in
experimental polymicrobial sepsis model. Transl Perioper Pain Med.
5:63–74. 2018.PubMed/NCBI
|
|
27
|
Borowiak R, Reichardt W, Kurzhunov D,
Schuch C, Leupold J, Krafft AJ, Reisert M, Lange T, Fischer E and
Bock M: Initial investigation of glucose metabolism in mouse brain
using enriched 17O-glucose and dynamic
17O-MRS. NMR Biomed. 30:2017. View Article : Google Scholar
|
|
28
|
Johansson N, Ahonen M and Kähäri VM:
Matrix metalloproteinases in tumor invasion. Cell Mol Life Sci.
57:5–15. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mogili NS, Krishnaswamy VR, Jayaraman M,
Rajaram R, Venkatraman A and Korrapati PS: Altered angiogenic
balance in keloids: A key to therapeutic intervention. Transl Res.
159:182–189. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pisacane AM, Picciotto F and Risio M: CD31
and CD34 expression as immunohistochemical markers of endothelial
transdifferentiation in human cutaneous melanoma. Cell Oncol.
29:59–66. 2007.PubMed/NCBI
|
|
31
|
Unahabhokha T, Sucontphunt A, Nimmannit U,
Chanvorachote P, Yongsanguanchai N and Pongrakhananon V: Molecular
signalings in keloid disease and current therapeutic approaches
from natural based compounds. Pharm Biol. 53:457–463. 2015.
View Article : Google Scholar
|
|
32
|
Bijlard E, Steltenpool S and Niessen FB:
Intralesional 5-Fluorouracil in keloid treatment: A systematic
review. Acta Derm Venerol. 95:778–782. 2015.PubMed/NCBI
|
|
33
|
Ud-Din S and Bayat A: New insights on
keloids, hypertrophic scars, and striae. Dermatol Clin. 32:193–209.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mari W, Alsabri SG, Tabal N, Younes S,
Sherif A and Simman R: Novel insights on understanding of keloid
scar: Article review. J Am Coll Clin Wound Spec. 7:1–7. 2016.
|
|
35
|
Andrews JP, Marttala J, Macarak E,
Rosenbloom J and Uitto J: Keloids: The paradigm of skin
fibrosis-pathomechanisms and treatment. Matrix Biol. 51:37–46.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Runyan CE, Schnaper HW and Poncelet AC:
The phosphati-dylinositol 3-kinase/Akt pathway enhances
Smad3-stimulated mesangial cell collagen I expression in response
to transforming growth factor-beta1. J Biol Chem. 279:2632–2639.
2004. View Article : Google Scholar
|
|
37
|
Lopiccolo J, Blumenthal GM, Bernstein WB
and Dennis PA: Targeting the PI3K/Akt/mTOR pathway: Effective
combinations and clinical considerations. Drug Resist Update.
11:32–50. 2008. View Article : Google Scholar
|
|
38
|
Carracedo A, Ma L, Teruya-Feldstein J,
Rojo F, Salmena L, Alimonti A, Egia A, Sasaki AT, Thomas G, Kozma
SC, et al: Inhibition of mTORC1 leads to MAPK pathway activation
through a PI3K-dependent feedback loop in human cancer. J Clin
Invest. 118:3065–3074. 2008.PubMed/NCBI
|
|
39
|
Wee S, Jagani Z, Xiang KX, Loo A, Dorsch
M, Yao YM, Sellers WR, Lengauer C and Stegmeier F: PI3K pathway
activation mediates resistance to MEK inhibitors in KRAS mutant
cancers. Cancer Res. 69:4286–4293. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jia J, Zhu F, Ma X, Cao ZW, Li YX and Chen
YZ: Mechanisms of drug combinations: Interaction and network
perspectives. Nat Rev Drug Discov. 8:111–128. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pei Y, Liu KW, Wang J, Garancher A, Tao R,
Esparza LA, Maier DL, Udaka YT, Murad N, Morrissy S, et al: HDAC
and PI3K antagonists cooperate to inhibit growth of MYC-driven
medulloblastoma. Cancer Cell. 29:311–323. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wozniak MB, Villuendas R, Bischoff JR,
Aparicio CB, Martínez Leal JF, de La Cueva P, Rodriguez ME,
Herreros B, Martin-Perez D, Longo MI, et al: Vorinostat interferes
with the signaling transduction pathway of T-cell receptor and
synergizes with phosphoinositide-3 kinase inhibitors in cutaneous
T-cell lymphoma. Haematologica. 95:613–621. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Delcuve GP, Khan DH and Davie JR:
Targeting class I histone deacetylases in cancer therapy. Expert
Opin Ther Tar. 17:29–41. 2013. View Article : Google Scholar
|
|
44
|
Kawada J, Ito Y, Iwata S, Suzuki M, Kawano
Y, Kanazawa T, Siddiquey MN and Kimura H: mTOR inhibitors induce
cell-cycle arrest and inhibit tumor growth in Epstein-Barr
virus-associated T and natural killer cell lymphoma cells. Clin
Cancer Res. 20:5412–5422. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cuyas E, Corominas-Faja B, Joven J and
Menendez JA: Cell cycle regulation by the nutrient-sensing
mammalian target of rapamycin (mTOR) pathway. Methods Mol Biol.
1170:113–144. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Loos C, Syrovets T, Musyanovych A,
Mailänder V, Landfester K and Simmet T: Amino-functionalized
nanoparticles as inhibitors of mTOR and inducers of cell cycle
arrest in leukemia cells. Biomaterials. 35:1944–1953. 2014.
View Article : Google Scholar
|
|
47
|
Du L, Risinger AL, King JB, Powell DR and
Cichewicz RH: A potent HDAC inhibitor, 1-Alaninechlamydocin, from a
Tolypocladium sp. induces G2/M cell cycle arrest and apoptosis in
MIA PaCa-2 cells. J Nat Prod. 77:1753–1757. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Feng W, Cai D, Zhang B, Lou G and Zou X:
Combination of HDAC inhibitor TSA and silibinin induces cell cycle
arrest and apoptosis by targeting survivin and cyclinB1/cdk1 in
pancreatic cancer cells. Biomed Pharmacother. 74:257–264. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ramakrishnan S, Ku S, Ciamporcero E, Miles
KM, Attwood K, Chintala S, Shen L, Ellis L, Sotomayor P, Swetzig W,
et al: HDAC 1 and 6 modulate cell invasion and migration in clear
cell renal cell carcinoma. BMC Cancer. 16:6172016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tian H, Zhang Y, Zhang Q, Li S, Liu Y and
Han X: Effects of BENC-511, a novel PI3K inhibitor, on the
proliferation and apoptosis of A549 human lung adenocarcinoma
cells. Biosci Trends. 13:40–48. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Eto S, Saeki K, Yoshitake R, Yoshimoto S,
Shinada M, Ikeda N, Kamoto S, Tanaka Y, Kato D, Maeda S, et al:
Anti-tumor effects of the histone deacetylase inhibitor vorinostat
on canine urothelial carcinoma cells. PLoS One. 14:e02183822019.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Huang WW, Tsai SC, Peng SF, Lin MW, Chiang
JH, Chiu YJ, Fushiya S, Tseng MT and Yang JS: Kaempferol induces
autophagy through AMPK and AKT signaling molecules and causes G2/M
arrest via downregulation of CDK1/cyclin B in SK-HEP-1 human
hepatic cancer cells. Int J Oncol. 42:2069–2077. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Makarević J, Rutz J, Juengel E, Maxeiner
S, Tsaur I, Chun FK, Bereiter-Hahn J and Blaheta RA: Influence of
the HDAC inhibitor valproic acid on the growth and proliferation of
temsirolimus-resistant prostate cancer cells in vitro. Cancers
(Basel). 11. pp. E5662019, View Article : Google Scholar
|
|
54
|
Sun Z, Cao B and Wu J: Protease-activated
receptor 2 enhances renal cell carcinoma cell invasion and
migration via PI3K/AKT signaling pathway. Exp Mol Pathol.
98:382–389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhou H, Jiang S, Chen J, Ren X, Jin J and
Su SB: Largazole, an inhibitor of class I histone deacetylases,
attenuates inflammatory corneal neovascularization. Eur J
Pharmacol. 740:619–626. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bian X, Liang Z, Feng A, Salgado E and
Shim H: HDAC inhibitor suppresses proliferation and invasion of
breast cancer cells through regulation of miR-200c targeting CRKL.
Biochem Pharmacol. 147:30–37. 2018. View Article : Google Scholar
|
|
57
|
Luo K: Signaling cross talk between
TGF-β/Smad and other signaling pathways. Cold Spring Harb Perspect
Biol. 9:a0221372017. View Article : Google Scholar
|
|
58
|
Guo W, Shan B, Klingsberg RC, Qin X and
Lasky JA: Abrogation of TGF-beta1-induced fibroblast-myofibroblast
differentiation by histone deacetylase inhibition. Am J Physiol
Lung Cell Mol Physiol. 297:L864–L870. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Bettinger DA, Yager DR, Diegelmann RF and
Cohen KI: The effect of TGF-beta on keloid fibroblast proliferation
and collagen synthesis. Plast Reconstr Surg. 98:827–833. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Peltonen J, Hsiao LL, Jaakkola S, Sollberg
S, Aumailley M, Timpl R, Chu M and Uitto J: Activation of collagen
gene expression in keloids: Co-localization of type I and VI
collagen and transforming growth factor-beta1 mRNA. J Invest
Dermatol. 97:240–248. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Lim IJ, Phan TT, Tan EK, Nguyen TT, Tran
E, Longaker MT, Song C, Lee ST and Huynh HT: Synchronous activation
of ERK and phosphatidylinositol 3-kinase pathways is required for
collagen and extracellular matrix production in keloids. J Biol
Chem. 278:40851–40858. 2003. View Article : Google Scholar : PubMed/NCBI
|