Introduction
Cervical cancer (CC), which is a common gynecologic
malignancy, has the third highest incidence among female malignant
tumors worldwide (1). The
incidence of CC in China is the highest among female malignant
tumors (2). Radical surgery,
radiotherapy and chemotherapy are main treatment methods for CC,
however, the adverse reactions caused by treatment seriously affect
the quality of life of patients. Therefore, it is necessary to
investigate the mechanism of CC at the molecular level and to
identify effective gene targets for the clinical treatment of
CC.
Micro(mi)RNAs are a group of small, conserved,
non-coding RNAs that can be sequence-specifically bound to
3'untranslated regions (UTRs) on homologous mRNA targets (3). In recent years, studies have found
that miRNAs can regulate the development of tumors and biological
behaviors, such as chemotherapeutic sensitivity, by regulating
oncogenes/tumor suppressor genes (4,5).
miR-217, which is located on chromosome 2P16.1, is an miRNA that
inhibits cell proliferation and serves a key regulatory role in the
growth and development of various cells. It has been shown that
miR-217 is closely related to tumor cell proliferation and
migration (6,7). In human pancreatic ductal gland
tumors, miR-217 suppresses cell proliferation by targeting target
genes, including Tpd52l2, Kras and E2F3 (8-10).
miR-217 also targets dickkopf-1 to regulate the WNT signaling
pathway and therefore affects the properties of stem cell in
hepatocellular carcinoma (11).
However, the effect of miR-217 in CC remains to be fully
elucidated.
It has been demonstrated that the mitogen-activated
protein kinase (MAPK) signaling pathway serves a pivotal biological
role in the progression of CC (12-14). MAPKs, which are a series of
intracellular serine/threonine protein kinases, are highly
conserved in evolution. MAPK1, also known as extracellular
signal-regulated kinase (ERK)2, is located in the cytoplasm prior
to activation and, once activated into the nucleus, it can be
activated or be expressed at a high level in tissues in several
types of tumor, including CC, lung cancer, breast cancer and liver
cancer, by activating target genes (15-18). Therefore, the expression of MAPK1
is closely associated with tumors. The ERK1/2 pathway is a pivotal
signal transduction pathway in the MAPK family and is closely
correlated with the progression of tumorigenesis (19,20).
In the present study, the clinical significance of
the expression levels of miR-217 in CC and the regulatory mechanism
underlying the effect of miR-217 in the progression of CC were
investigated. The study aimed to provide an effective strategy for
treating patients with CC.
Materials and methods
Human-derived CC samples
A total of 60 human-derived CC specimens and
non-cancerous tissues were sampled between 2015 and 2017 at The
Second Affiliated Hospital of Shaanxi University of Traditional
Chinese Medicine (Shaanxi, China). Of the patients involved, 33
were male and 27 were female, aged between 42 and 63 years of age.
The experiments were approved by the Ethics Committee of The Second
Affiliated Hospital of Shaanxi University of Traditional Chinese
Medicine. All tissues were collected prior to the initiation of
radiotherapy or chemotherapy and were frozen immediately and stored
at -80°C until required. Cytological and/or pathological evidence
of a CC diagnosis were available from each subject. Patients
provided written informed consent.
Cell line culture
The Ect1/E6E7 human normal cervical cell line, CC
cell lines of human origin (HeLa, SiHa, Caski, Me180, Ms751 and
C33a) and 293T cells were procured from the American Type Culture
Collection (Rockville, MD, USA). The cells were grown in RPMI 1640
medium or Dulbecco's modified Eagle's medium (DMEM, GE Healthcare
Life Sciences, Logan, UT, USA) supplemented with 10% fetal bovine
serum (FBS; Invitrogen, Thermo Fisher Scientific, Inc., Waltham,
MA, USA), 100 U/ml penicillin and 100 μg/ml streptomycin in
a humidified chamber with 5% CO2 at 37°C.
miRNA and reagents
The miR-217-mimic, miR-217-negative control (NC) and
MAPK1 expression vector were purchased from GenePharma (Shanghai,
China). Lipofectamine® 3000 transfection reagent (Thermo
Fisher Scientific, Inc.) was used to transfect the cells with the
aforementioned miRNAs according to the manufacturer's protocol,
with a transfection incubation time of 6 h. PD98059 was obtained
from Tocris Bioscience (Ellisville, MO, USA; CAS no. 167869-21-8,
HPLC ≥98%).
Cell viability assay
A Cell Counting Kit-8 (CCK-8) assay (Beyotime
Institute of Biotechnology, Beijing, China) was performed to assess
cell viability according to the manufacturer's protocol. Briefly,
the transfected and untransfected cells were transferred to 96-well
plates (3,000 cells/well). Following incubation for 2 h in 10
μl CCK-8 at 37°C, the absorbance in every well was read at
450 nm using a microplate reader (Tecan Infinite M200 Microplate
Reader; LabX, Tecan Group, Ltd., Männedorf, Switzerland).
Bioinformatics prediction
The potential target genes of miR-217 were predicted
using TargetScan 7.2 online software (www.targetscan.org) according to the manufacturer's
instructions. 'miR-217' was inserted and 'human' was selected. The
putative target genes of miR-217 were scanned.
Dual-luciferase reporter assay
Wild-type MAPK1 (MAPK1-WT) and mutated MAPK1
(MAPK1-MUT) were cloned into pMIR-REPORT luciferase vectors
(Ambion; Thermo Fisher Scientific, Inc.). 293T cells
(5×105 cells/well) were seeded into 6-well plates and
then transfected with both vectors using Lipofectamine
3000® (Invitrogen; Thermo Fisher Scientific, Inc.) for
24 h, according to the manufacturer's manual. The
Dual-Luciferase-Reporter 1000 Assay system (Promega Corporation,
Madison, WI, USA) was used to assess luciferase activity. Renilla
activity was used for normalization.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
The total RNA was extracted using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The concentration and purity of RNA were
determined using the NanoDrop2000 spectrophotometer (NanoDrop
Technologies; Thermo Fisher Scientific, Inc.). The total RNA (1
μg) was reverse transcribed using a reverse transcription
cDNA kit (Thermo Fisher Scientific, Inc.) and TaqMan™ MicroRNA
Reverse Transcription kit (Applied Biosystems, Thermo Fisher
Scientific, Inc.) for the synthesis of cDNA (42°C for 60 min, 70°C
for 5 min, preserved at 4°C). SYBR-Green PCR Master mix (Roche
Diagnostics, Basel, Switzerland) and the TaqMan miRNA PCR kit
(Applied Biosystems, Thermo Fisher Scientific, Inc.) were used to
perform qPCR assays for MAPK1 and miRNA-217 using the Opticon
RT-PCR detection system (ABI 7500, Thermo Fisher Scientific, Inc.),
The PCR cycle was set as follows: Pretreatment at 95°C for 10 min,
followed by 40 cycles at 94°C for 15 sec, 60°C for 1 min, 60°C for
1 min and preserved at 4°C. The comparative quantification cycle
(2−ΔΔCq) method (21)
was used to analyze the expression of mRNA. The expression of GAPDH
was used for normalization. The sequences of the primers used were
as follows: MAPK1, forward 5′-GTCGCCATCAAGAAAATCAGC-3′, and reverse
5′-GGAAGGTTTGAGGTCACGGT-3′; GAPDH, forward
5′-AGAAGGCTGGGGCTCATTTG-3′, and reverse 5'-AGGGGCCATCCACAG
TCTTC-3′; miR-217, forward 5′-TACTCAACTCACTACTGCATCAGG A-3′, and
reverse 5′-TATGGTTGTTCTGCTCTCTGTGTC-3′; and U6, forward
5′-CTCGCTTCGGCAGCACA-3′, and reverse 5′-TGGTGTCGTGGAGTCG-3′.
Western blot analysis
Total proteins were collected using RIPA (Cell
Signaling Technology, Inc., Danvers, MA, USA). A BCA Protein Assay
kit (Pierce; Thermo Fisher Scientific, Inc.) was used to measure
the concentrations of proteins and adjusted to a concentration of 6
μg/μl using 1X loading and DEPC water. The samples (5
μl) were separated on 10% SDS-PAGE gels and then transferred
onto a polyvinylidene fluoride membrane (EMD Millipore, Billerica,
MA, USA). Following blocking in 5% non-fat milk in PBST [0.1%
Tween-20 in phosphate-buffered saline (PBS)] for 1 h, the membrane
was probed with the primary antibody overnight at 4°C and washed
three times in PBST. The membrane was then incubated with secondary
antibody (horseradish peroxidase-conjugated goat anti-mouse/rabbit
IgG, 1:2,000; cat. no. sc-516102/sc-2357; Santa Cruz Biotechnology,
Inc. Dallas, TX, USA) at room temperature for 2 h. The membrane was
then washed with PBST three times. A developer (EZ-ECL kit;
Biological Industries) was used for development, and the gray
values of the strips were analyzed and counted using ImageJ
software (version 5.0; Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The antibodies used were as follows: Anti-GAPDH (mouse;
1:1,000; cat. no. LS-B1625; LifeSpan BioSciences, Inc.), anti-Bax
(rabbit; 1:1,000; cat. no. ab32503; Abcam), anti-Bcl-2 (rabbit;
1:1,000; cat. no. ab32124; Abcam), anti-cleaved caspase-3 (rabbit;
1:1,000; cat. no. #9661; Cell Signaling Technology, Inc.),
anti-p-ERK1/2 (rabbit; 1:1,000; cat. no. #4370; Cell Signaling
Technology, Inc.), anti-ERK1/2 (mouse; 1:1,000; cat. no. #4696;
Cell Signaling Technology, Inc.) and anti-MAPK1 (rabbit; 1:1,000;
cat. no. #9108; Cell Signaling Technology, Inc.).
Analysis of apoptosis
Flow cytometry was used for the analysis of
apoptosis. The HeLa cells were transfected with an expression
vector or mimics and incubated for 24 h. The supernatant was
collected in a 15-ml centrifuge tube, and the culture flask was
gently washed once by adding 2 ml PBS. Trypsin (1 ml) without
ehylenediaminetetraacetic acid was used to digest the cells by
shaking gently, and the pancreatic enzyme was aspirated when the
wall was wet. The mixture was stood at room temperature for 1 min,
and DMEM (Corning Incorporated) containing 10% FBS was added to
terminate digestion. The cells were centrifuged at 1,000 × g for 3
min at 4°C, and the supernatant was removed. The cells were then
washed twice with pre-cooled PBS and resuspended in 1X Annexin V
binding buffer. According to the Annexin-V-FITC cell apoptosis
detection kit (cat. no. K201-100, BioVision, Inc., Milpitas, CA,
USA), at room temperature, the cells were collected and stained
with Annexin V-FITC and propidium iodide (PI) for 15 min and
counted by flow cytometry (version 10.0, FlowJo, FACS Calibur™, BD
Biosciences, Franklin Lakes, NJ, USA). The flow cytometry scatter
diagrams showed that living cells shown in the lower left quadrant
are mechanically damaged and necrotic cells in the left upper
quadrant are necrotic, whereas advanced apoptotic cells are shown
in the upper right quadrant and early apoptotic cells are shown in
the lower right quadrant.
Flow cytometric cell cycle analysis
Cell cycle analysis was performed by flow cytometry.
To be more specific, 5×105 cells with 70% cold ethanol
were cultured -20°C overnight. The following day, the fixed cells
were centrifuged at 1,200 × g for 1 min at 4°C, and washed twice
with PBS. The cells were treated with 200 μl RNase A (1
mg/ml) for 10 min at 37°C in suspension, following which 300
μl PI (100 μl/ml, BioVision, Inc.) was added to stain
the DNA in cells in the dark. Following incubation at room
temperature for 20 min, the cellular DNA content of the cells was
analyzed in a FACScan flow cytometer (BD Biosciences) using ModFit
LT software V2.0 (BD Biosciences).
Wound-healing assay
Following transfection of the HeLa cells for 24 h, a
gap was created using a 200-μl sterile tip across the middle
of the well. The cells were washed twice with DMEM for smoothing
the edges of the scratch and floating cells were removed. Following
incubation in an incubator (37°C, 5% CO2) for 0 and 24
h, the migration of the cells was observed under a light microscope
(Keyence Corporation, Osaka, Japan), the distance of cell migration
was visualized and images were captured using Image-Pro Plus
Analysis software 6.0 (Media Cybernetics, Inc., Rockville, MD,
USA).
Transwell assay
An 8-μm Transwell chamber (cat. no. 3413,
Corning Incorporated,) was placed on a 24-well plate with a layer
of Matrigel (50 μl; BD Biosciences) coated on the Transwell
chamber. The HeLa cells were cultured in serum-free medium for 12 h
to eliminate the effects of the serum and then resuspended in DMEM
containing bovine serum albumin (BSA; Sigma-Aldrich; Merck KGaA)
with free FBS. The suspended cells (100 μl) were added to
the Transwell chamber, and 400 μl of DMEM containing 20% FBS
was added to the basolateral chamber. The cells were cultured for
24 h at 37°C in an incubator with 5% CO2. The Transwell
chamber was then removed, the culture solution in the Transwell was
discarded and washed twice with calcium-free PBS, and the chamber
was fixed in methanol solution for 30 min and stained with 0.1%
crystal violet for 20 min at room temperature. Subsequently, the
chamber was washed with PBS and the upper chamber liquid was
aspirated. The unmigrated cells in the upper layer were gently
wiped off using a cotton swab. The microporous membrane was
carefully removed using a small pair of tweezers, dried with the
bottom side facing upwards and then transferred onto a glass slide
and sealed using a neutral gum. Images were observed and collected
using an inverted optical microscope (Keyence Corporation).
Statistical analysis
All experimental data are presented as the mean ±
SEM. P<0.05 was considered to indicate a statistically
significant difference. Statistical significance was determined by
one-way analysis of variance between groups, followed by a
Bonferroni test. All statistical analyses were performed using
GraphPad Prism 6 (GraphPad Software, Inc.).
Results
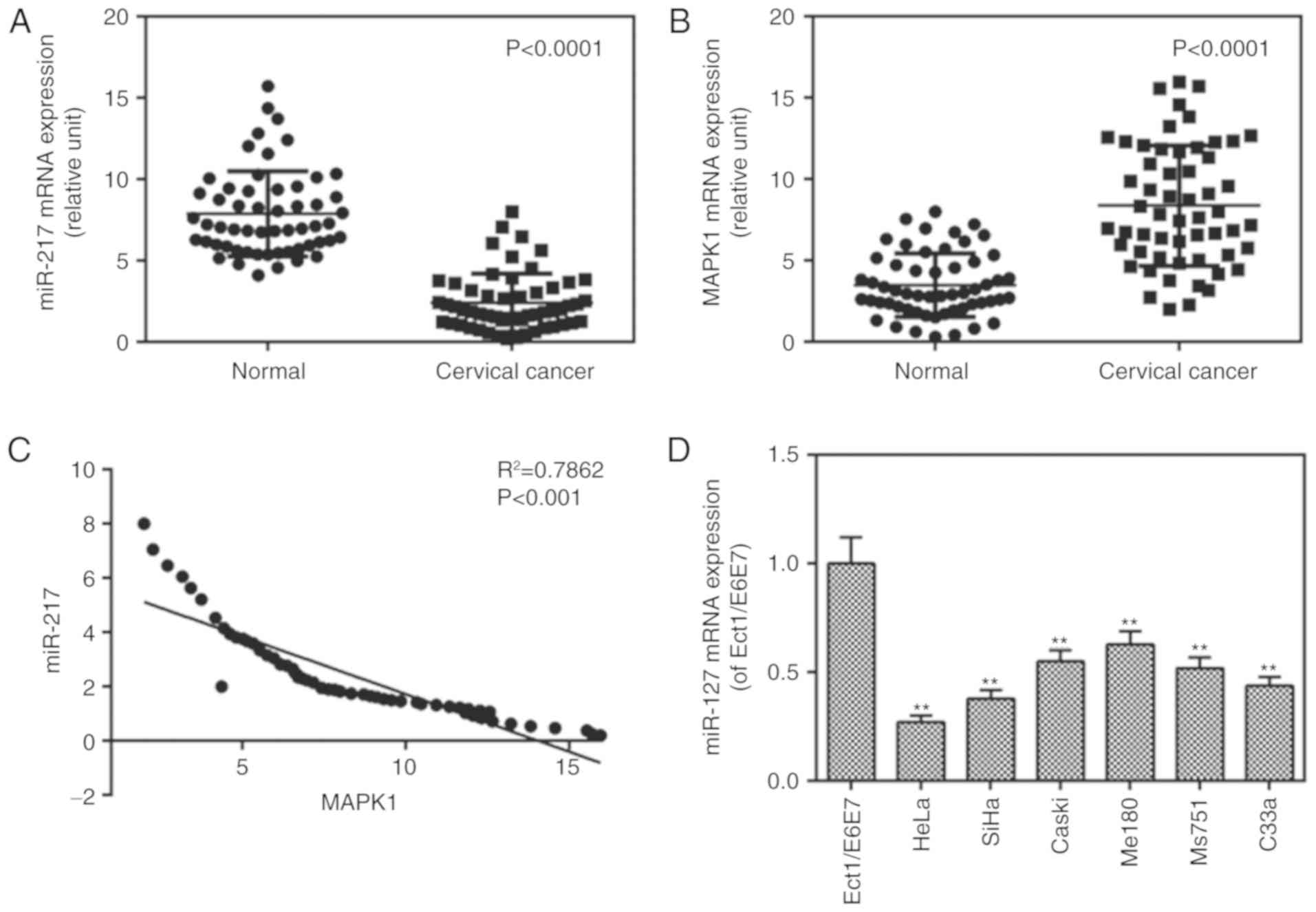
miR-217 is downregulated in tissues and
cells and MAPK1 is expressed in tissues
The mRNA levels of miR-217 in cancer tissues from
patients with CC, CC cell lines and controls were measured, and the
mRNA levels of MAPK1 were determined in CC tissues. The results
showed that the mRNA level of miR-217 was decreased (Fig. 1A) whereas that of MAPK1 (Fig. 1B) was increased in the cancer
tissue group, compared with levels in the normal group. It was also
found that there was a negative correlation between the levels of
miR-217 and MAPK1 (Fig. 1C). It
was also found that the expression of miR-217 was lower in the CC
cell lines than that in the control cells (Fig. 1D). The expression of miR-217
exhibited a greater degree of reduction in the HeLa cells, thus,
the HeLa cells were used in later experiments.
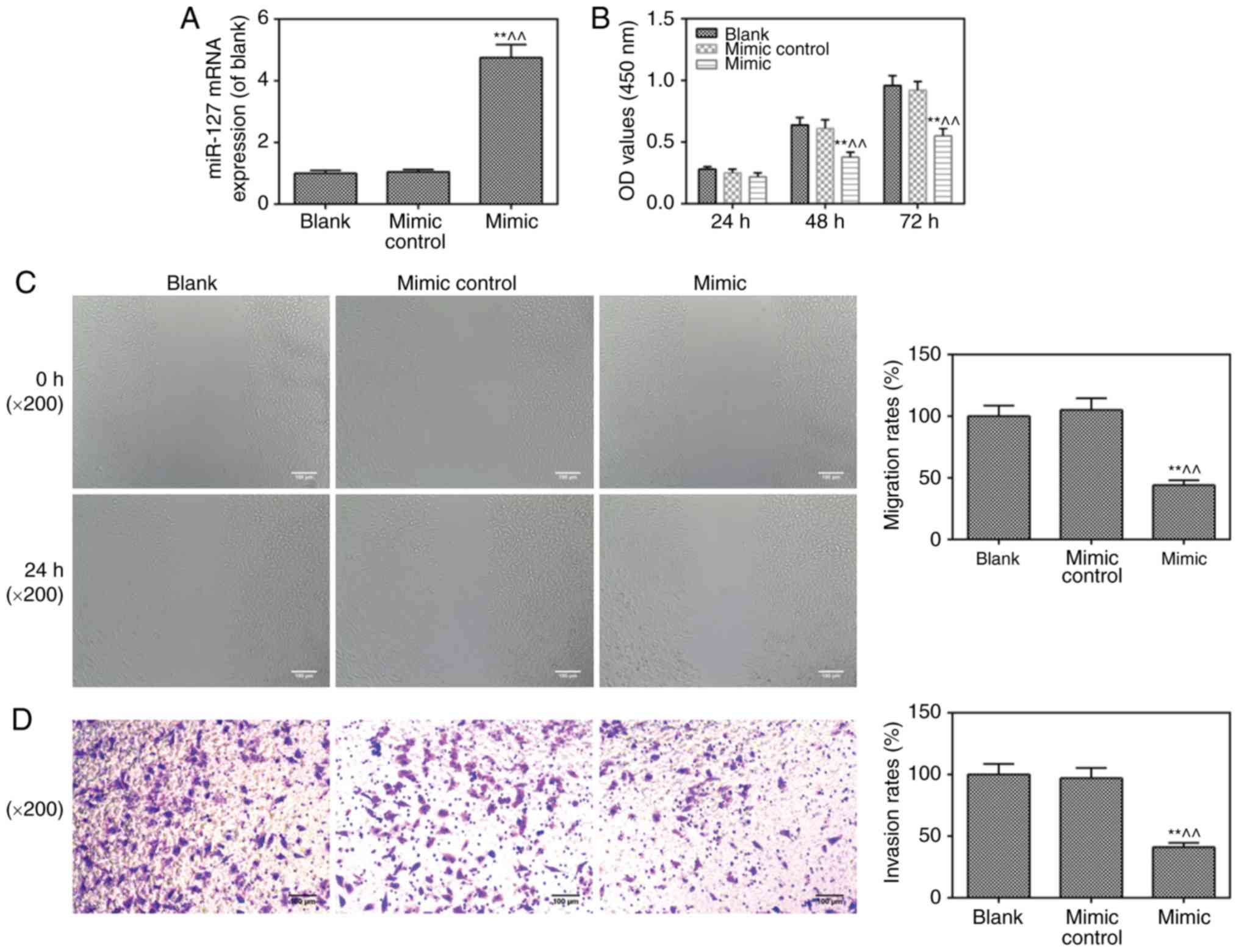
Viability, invasion and migration of
cells are inhibited by miR-217 mimic transfection
To further investigate the role of miR-217 in CC,
the miR-217 mimic was transfected into HeLa cells. As shown in
Fig. 2A, the cells were
successfully transfected with the miR-217 mimic. It was found that
the miR-217 mimic suppressed cell viability (Fig. 2B), migration (Fig. 2C) and invasion (Fig. 2D), compared with levels in the
blank and mimic control groups, respectively.
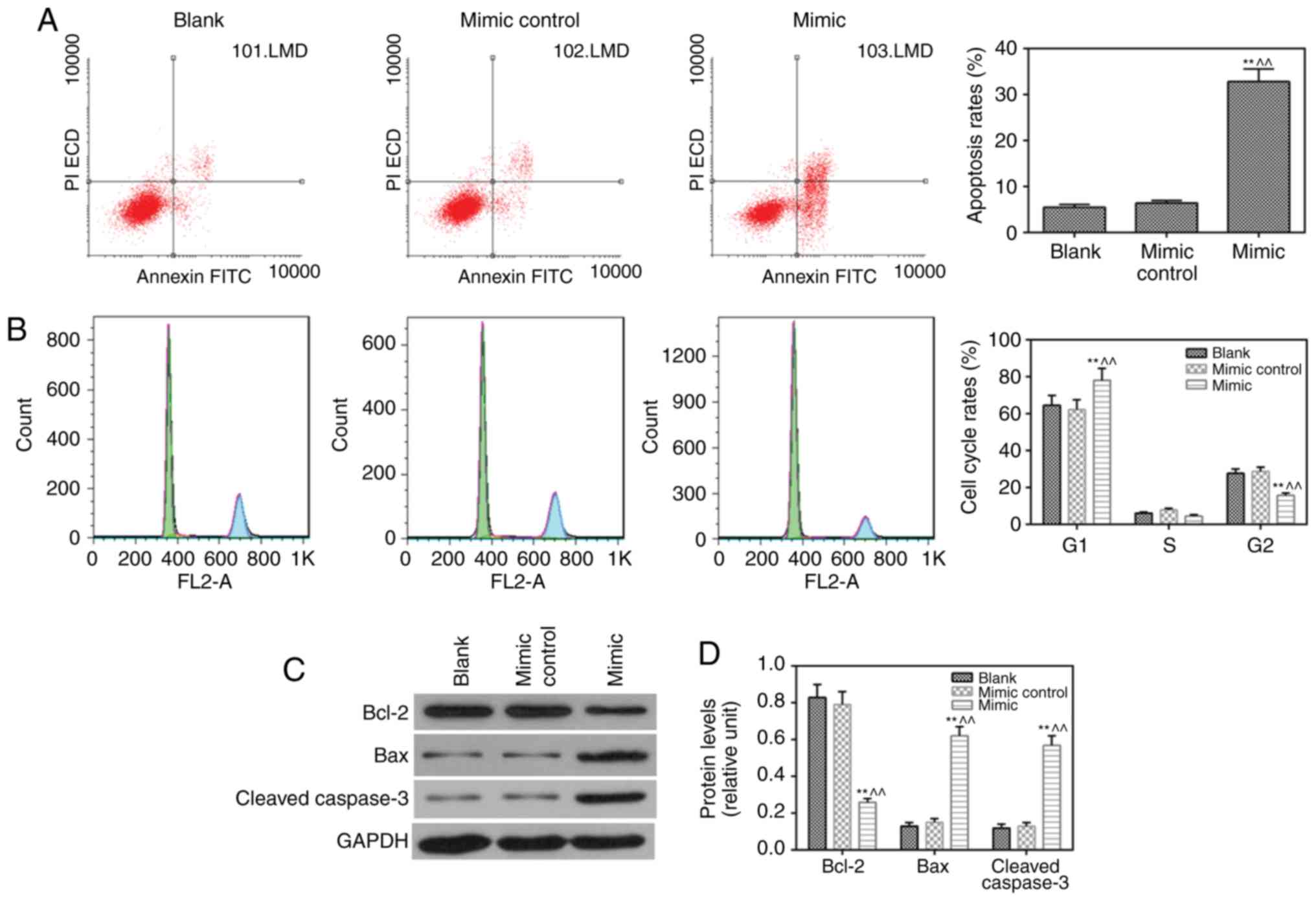
miR-217 mimic induces apoptosis and
inhibits the cell cycle
The results revealed that apoptosis (Fig. 3A) was induced and cell cycle
(Fig. 3B) was suppressed by the
miR-217 mimic, compared with observations in the blank and mimic
control groups, respectively. The protein level of anti-apoptotic
Blc-2 was lower, whereas the expression levels of pro-apoptotic
proteins (Bax and cleaved caspase-3) were higher in the miR-217
mimic group compared with those in the blank and mimic control
groups (Fig. 3C and D).
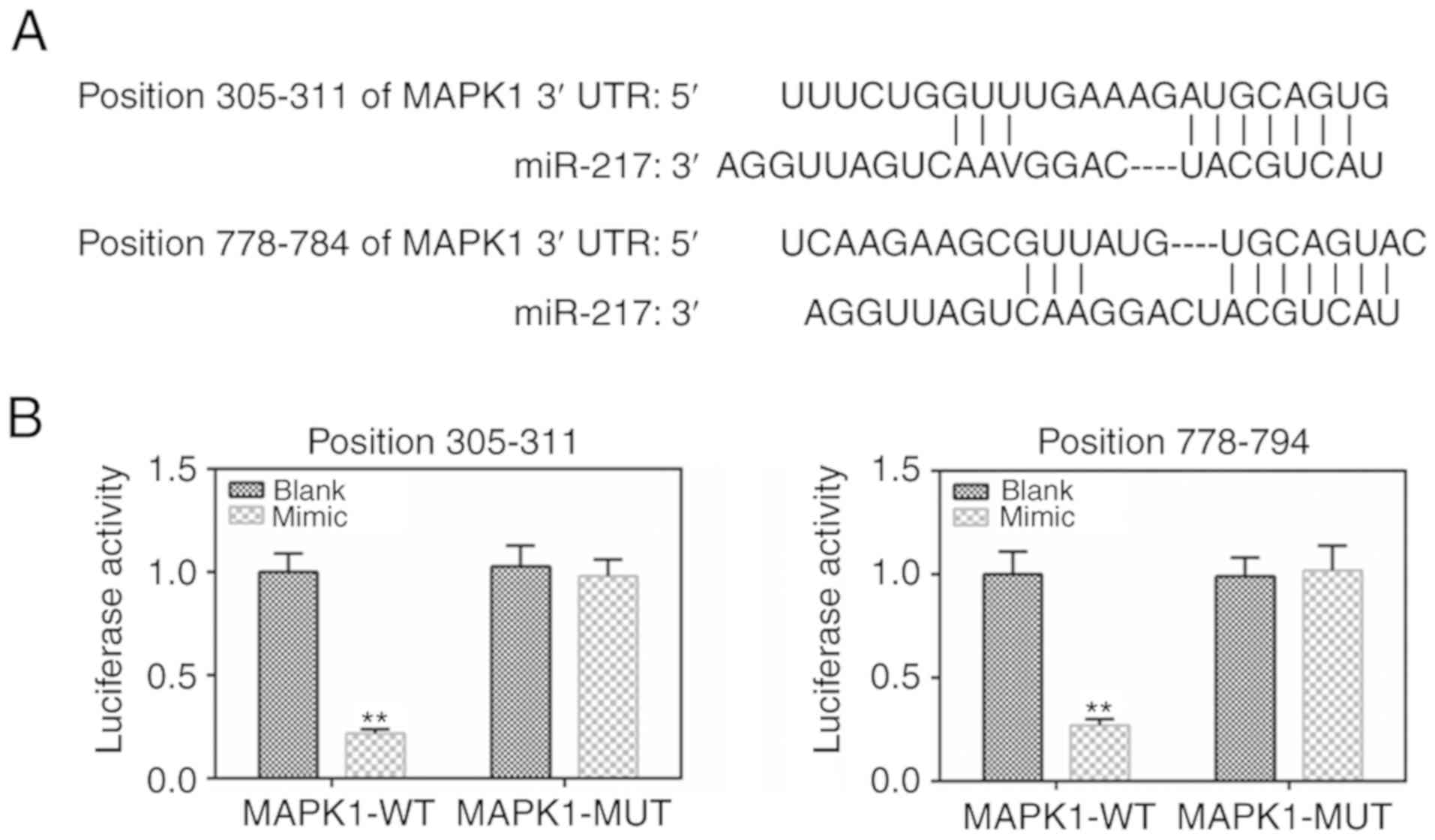
MAPK1 is the target gene of miR-217
To determine the binding capacity of miR-217 to
MAPK1, the TargetScan 7.2 website was used and MAPK1 was identified
as a potential target of miR-217 (Fig. 4A). The dual-luciferase reporter
gene assay data demonstrated that the luciferase activity was lower
in the MAPK1-WT mimic group than that in the MAPK1-WT blank group
(Fig. 4B), with no significant
changes shown in the MAPK1-MUT group.
Inhibitory effects of miR-217 mimic on
cells are reversed by MAPK1
The role of MAPK1 in CC was investigated in
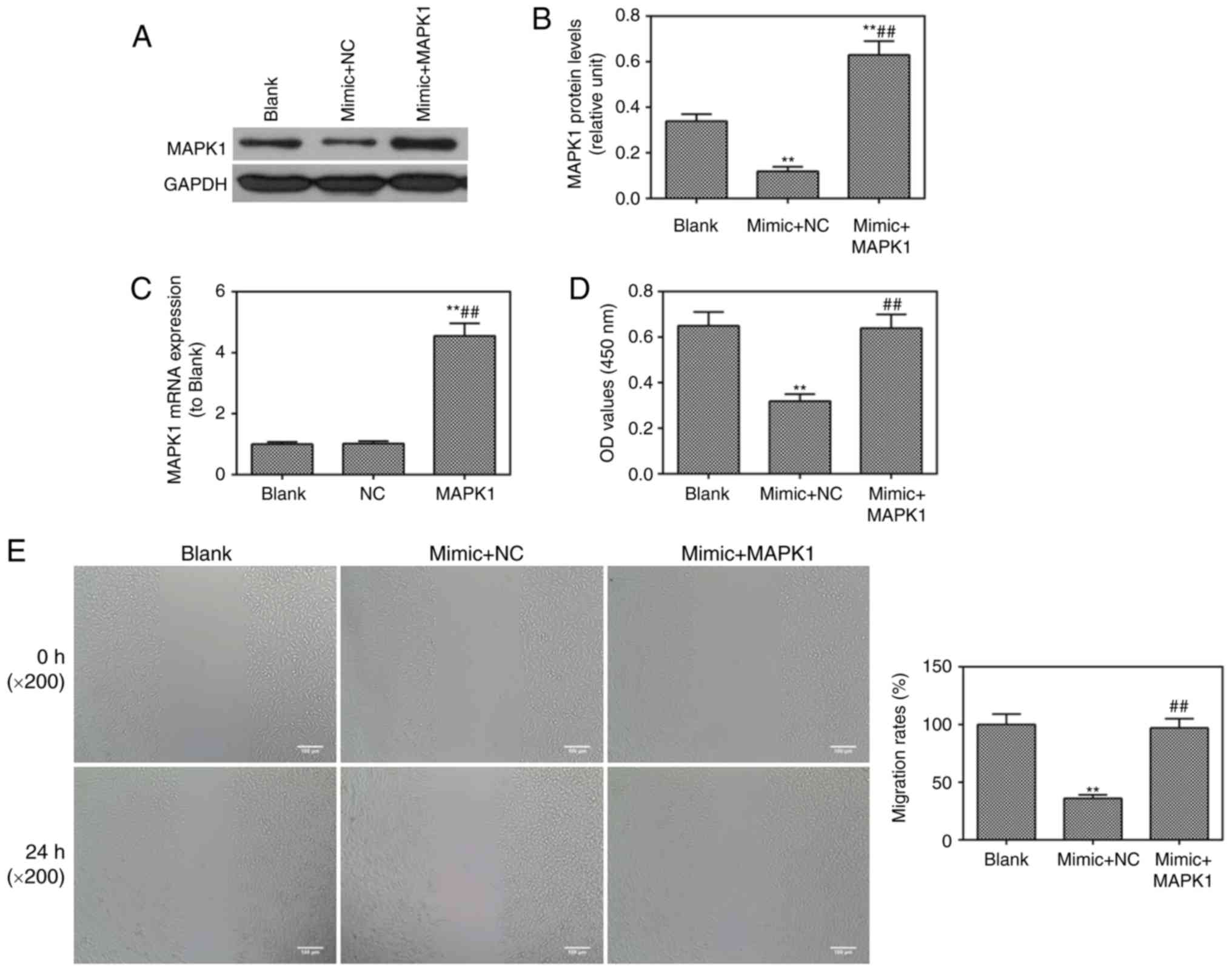
vitro. It was found that MAPK1 was successfully transfected
into cells through observation at the protein level (Fig. 5A and B). The mRNA expression of
MAPK1 was enhanced and confirmed by RT-qPCR analysis (Fig. 5C). The miR-217 mimic was found to
inhibit cell viability (Fig. 5D),
migration (Fig. 5E) and invasion
(Fig. 5F), which were reversed by
MAPK1. The miR-217 mimic-induced cell apoptosis was ameliorated by
MAPK1 (Fig. 5G). The protein
level of p-ERK1/2 was lower in the mimic group than that in the
blank group, which was also reversed by MAPK1 (Fig. 5H and I).
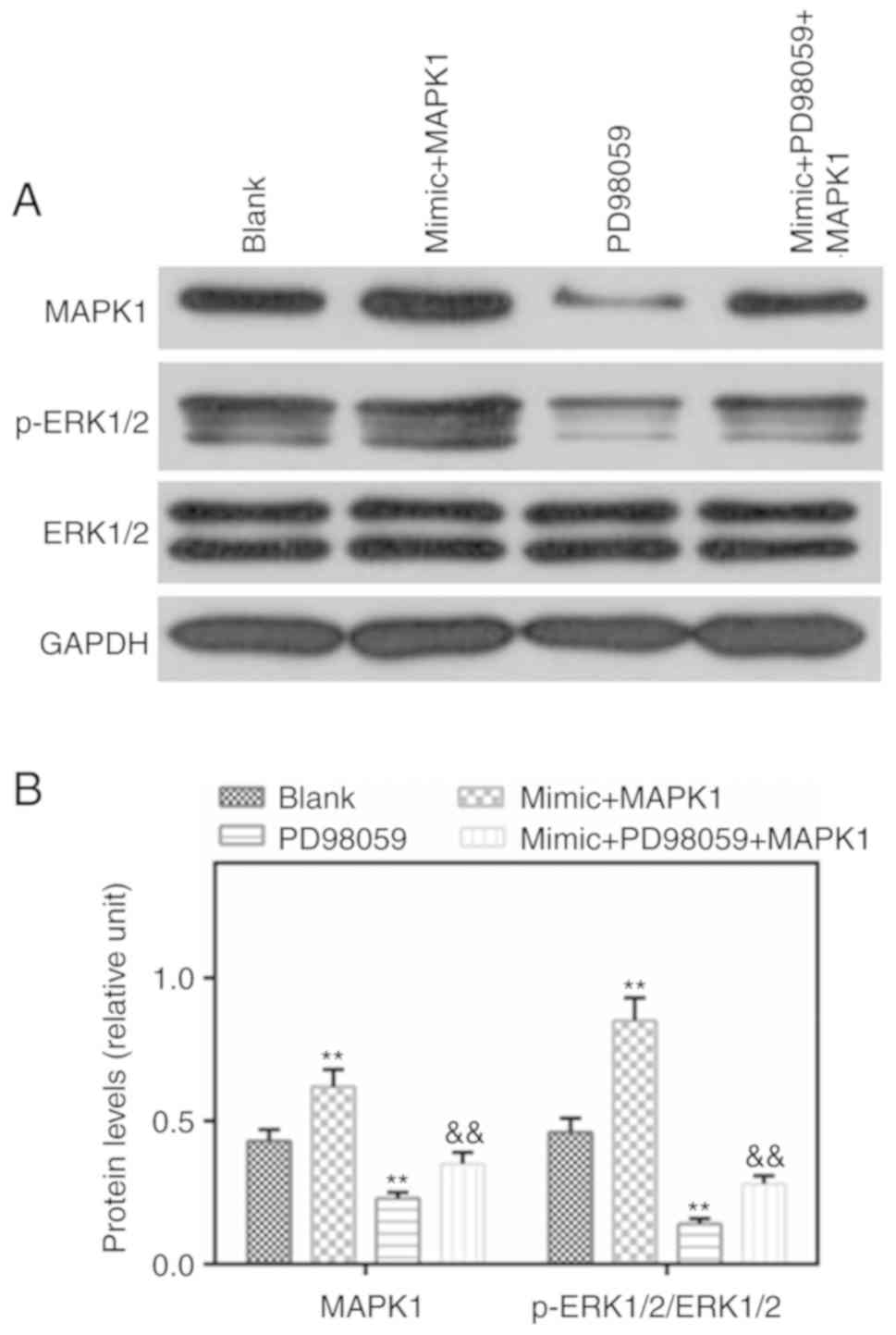
Protein levels of MAPK1 and p-ERK1/2 are
induced by MAPK1, which are reduced by PD98059
p-ERK1/2 is an important component of the MAPK
signaling pathway. In the present study, western blotting was
performed to detect the levels of p-ERK1/2 and MAPK1, and it was
found that the levels of MAPK1 and p-ERK1/2 were higher in the
mimic MAPK1 group compared with those in the Blank group. However,
PD98059 reversed the effects of MAPK1 on the levels of MAPK1 and
p-ERK1/2 (Fig. 6A and B).
Discussion
CC is one of the most important malignant tumors in
women worldwide and a major cause of cancer-related mortality.
Currently, the majority of patients with CC have no obvious
symptoms at an early stage, therefore, metastasis is often present
in patients with CC when they are diagnosed with the disease. In
the last few decades, progress has been made in the treatment of CC
(22,23). However, increasing the survival
rates of patients with CC remains a challenge.
miRNAs can act as tumor suppressors or promoters in
different types of cancer, as they target a variety of target genes
(24). According to reports,
miRNAs serve a pivotal role in the progression and metastasis of CC
by regulating cancer cell proliferation, apoptosis and cell cycle
(25). As an miRNA, miR-217 is
closely linked to tumor progression and poor prognosis (26-28). Previous studies have reported that
miR-217 bound to its target mRNA to inhibit the formation and
progression of tumors, including gastric cancer (29) and liver cancer (30). However, the role of miR-217 in CC
remains poorly understood.
The purpose of the present study was to examine the
role of mir-217 in the development of CC. The expression of miR-217
in CC tissues and cells was detected and confirmed to be
significantly reduced in CC tissues compared with that in normal
adjacent tissues, and similar results were observed in CC cell
lines. These data suggested that miR-217 may act as a tumor
suppressor gene in CC.
The biological functions of normal cells experience
major changes during carcinogenesis to promote the progression of
cancer, including invasion and metastasis (31,32). Various tissue cells in an organism
maintain a quantitative balance through proliferation and
apoptosis, however, disturbing this balance leads to diseases such
as cancer (33,34). Apoptosis is the main mechanism of
cell death induced by various anticancer drugs, thus, the role of
apoptosis in cancer therapy has become a focus in antitumor
research (35,36). miR-217 has previously been found
to be expressed at low levels in CC tissues and cells. The present
study aimed to determine the effect of a high expression of miR-217
on CC cells, therefore, an miR-217 mimic and a mimic control were
transfected into HeLa cells. It was found that cell viability,
metastasis, invasion and cell cycle were inhibited and apoptosis
was increased by the miR-217 mimic. In addition, the expression of
pro-apoptotic proteins was increased but that of anti-apoptotic
proteins was decreased in the miR-217 mimic group. These data
indicated that miR-217 may be a tumor suppressor gene in the
progression of CC.
To investigate the molecular mechanism underlying
the tumor suppressor effects of miR-217 on CC, bioinformatics
software (TargetScan 7.2) was used to identify the target of
miR-217 in CC cells. MAPK1 3'UTRs were found to have two binding
sequences for miR-217 at the positions 305-311 and 778-784. The
luciferase activity assay further confirmed that MAPK1 was a target
gene of miR-217.
The MAPK pathway, which is considered to be an
important protein cascade in cells, transfers signals from
receptors on the cell surface to the nucleus (37). The important signaling molecules
in this pathway are ERK and its upstream kinase (MEK) (20,38). The MAPK pathway is considered as a
potential target for cancer therapeutic intervention, as it is
effective in the regulation of cancer cell proliferation, invasion
and survival. Zhang et al (39) demonstrated that the inhibition of
MAPK1 inhibited tumorigenicity and metastasis in prostate cancer.
Zhang et al (40) found
that miR-217 suppressed tumor growth and apoptosis by targeting
MAPK1 in colorectal cancer. Hu et al (41) also showed that miR-585 directly
targeted MAPK1 to inhibit gastric cancer proliferation. These data
demonstrate that MAPK1 is involved in the development of cancer. In
the present study, it was found that the co-transfection of MAPK1
and the miR-217 mimic in CC cells restored the effects of the
miR-217 mimic in CC cells. The level of p-ERK1/2 was inhibited by
miR-217 mimic, which was reversed by MAPK1. To validate the role of
the MAPK signaling pathway in CC, PD98095, which is a potent
inhibitor of MEK, and MAPK1 (miR-217 mimic) were used to co-treat
the cells, and it was found that PD98095 increased the expression
levels of p-ERK1/2 and MAPK1. The above experimental data indicated
that miR-217 inhibited the occurrence and development of CC and
such a function may be correlated with the MAPK signaling
pathway.
Although the present study found that MAPK1 is a
target of miR-217 regulation in CC and effectively inhibited the
deterioration of CC, there are limitations in the study. First, the
function of miR-217 in protecting CC by modulating MAPK1 was only
supported by in vitro experiments. Additionally, the
observed significant regulation by miR-217 regulating MAPK1 in CC
is unclear and requires further investigation.
In conclusion, the findings of the present study
contribute to our understanding of the role of dysregulated miR-217
in the progression of CC by targeting MAPK1. The results provide an
effective therapeutic target to improve the survival rates of
patients with CC.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant. no. 81072940).
Availability of data and materials
The analyzed datasets generated during this study
are available from the corresponding author on reasonable
request.
Authors' contributions
LZ and JW made substantial contributions to
conception and design; SY, JW and LZ contributed to data
acquisition, data analysis and interpretation; SY and JW
contributed to drafting and critically revising the manuscript for
important intellectual content. All authors provided final approval
of the version to be published. All authors agreed to be
accountable for all aspects of the study, ensuring that questions
relating to its accuracy or integrity are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All procedures performed in experiments involving
human participants were in accordance with the Ethics Committee of
The Second Affiliated Hospital of Shaanxi University of Traditional
Chinese Medicine and with the 1964 Helsinki declaration and its
later amendments or comparable ethical standards.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Arbyn M, Castellsagué X, de Sanjosé S,
Bruni L, Saraiya M, Bray F and Ferlay J: Worldwide burden of
cervical cancer in 2008. Ann Oncol. 22:2675–2686. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cheng L, Guo Y, Zhan S and Xia P:
Association between HLA-DP gene polymorphisms and cervical cancer
risk: A meta-analysis. Biomed Res Int. 2018.7301595:2018.
|
|
3
|
Guo Z, Shu Y, Zhou H and Zhang W:
Identification of diagnostic and prognostic biomarkers for cancer:
Focusing on genetic variations in microRNA regulatory pathways
(Review). Mol Med Rep. 13:1943–1952. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kwok GT, Zhao JT, Weiss J, Mugridge N,
Brahmbhatt H, MacDiarmid JA, Robinson BG and Sidhu SB:
Translational applications of microRNAs in cancer, and therapeutic
implications. Noncoding RNA Res. 2:143–150. 2017. View Article : Google Scholar
|
|
5
|
Rizzo FM and Meyer T: Liquid biopsies for
neuroendocrine tumors: Circulating tumor cells, DNA, and MicroRNAs.
Endocrinol Metab Clin North Am. 47:471–483. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Su J, Wang Q, Liu Y and Zhong M: miR-217
inhibits invasion of hepatocellular carcinoma cells through direct
suppression of E2F3. Mol Cell Biochem. 392:289–296. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
de Yebenes VG, Bartolomé-Izquierdo N,
Nogales-Cadenas R, Pérez-Durán P, Mur SM, Martínez N, Di Lisio L,
Robbiani DF, Pascual-Montano A, Cañamero M, et al: miR-217 is an
oncogene that enhances the germinal center reaction. Blood.
124:229–239. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen Q, Wang P, Fu Y, Liu X, Xu W, Wei J,
Gao W, Jiang K, Wu J and Miao Y: MicroRNA-217 inhibits cell
proliferation, invasion and migration by targeting Tpd52l2 in human
pancreatic adeno-carcinoma. Oncol Rep. 38:3567–3573.
2017.PubMed/NCBI
|
|
9
|
Liu P, Yang H, Zhang J, Peng X, Lu Z, Tong
W and Chen J: The lncRNA MALAT1 acts as a competing endogenous RNA
to regulate KRAS expression by sponging miR-217 in pancreatic
ductal adenocarcinoma. Sci Rep. 7:51862017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang J, Zhang HF and Qin CF: MicroRNA-217
functions as a prognosis predictor and inhibits pancreatic cancer
cell proliferation and invasion via targeting E2F3. Eur Rev Med
Pharmacol Sci. 21:4050–4057. 2017.PubMed/NCBI
|
|
11
|
Jiang C, Yu M, Xie X, Huang G, Peng Y, Ren
D, Lin M, Liu B, Liu M, Wang W and Kuang M: miR-217 targeting DKK1
promotes cancer stem cell properties via activation of the Wnt
signaling pathway in hepatocellular carcinoma. Oncol Rep.
38:2351–2359. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li ZH, Li L, Kang LP and Wang Y:
MicroRNA-92a promotes tumor growth and suppresses immune function
through activation of MAPK/ERK signaling pathway by inhibiting PTEN
in mice bearing U14 cervical cancer. Cancer Med. May 11;2018Epub
ahead of print.
|
|
13
|
Zheng HY, Shen FJ, Tong YQ and Li Y: PP2A
inhibits cervical cancer cell migration by dephosphorylation of
p-JNK, p-p38 and the p-ERK/MAPK signaling pathway. Curr Med Sci.
38:115–123. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiao M, Feng Y, Cao G, Liu C and Zhang Z:
A novel MtHSP70-FPR1 fusion protein enhances cytotoxic T lymphocyte
responses to cervical cancer cells by activating human
monocyte-derived dendritic cells via the p38 MAPK signaling
pathway. Biochem Biophys Res Commun. 503:2108–2116. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang D, Li X, Yao Z, Wei C, Ning N and Li
J: GABAergic signaling facilitates breast cancer metastasis by
promoting ERK1/2-dependent phosphorylation. Cancer Lett.
348:100–108. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shilo A, Ben Hur V, Denichenko P, Stein I,
Pikarsky E, Rauch J, Kolch W, Zender L and Karni R: Splicing factor
hnRNP A2 activates the Ras-MAPK-ERK pathway by controlling A-Raf
splicing in hepatocellular carcinoma development. Rna. 20:505–515.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang C, Jin H, Gao D, Lieftink C, Evers B,
Jin G, Xue Z, Wang L, Beijersbergen RL, Qin W and Bernards R:
Phospho-ERK is a biomarker of response to a synthetic lethal drug
combination of sorafenib and MEK inhibition in liver cancer. J
Hepatol. 69:1057–1065. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li Q, Feng Y, Chao X, Shi S, Liang M, Qiao
Y, Wang B, Wang P and Zhu Z: HOTAIR contributes to cell
proliferation and metastasis of cervical cancer via targetting
miR-23b/MAPK1 axis. Biosci Rep. 38:BSR201715632018. View Article : Google Scholar :
|
|
19
|
Tasioudi KE, Saetta AA, Sakellariou S,
Levidou G, Michalopoulos NV, Theodorou D, Patsouris E and
Korkolopoulou P: pERK activation in esophageal carcinomas:
Clinicopathological associations. Pathol Res Pract. 208:398–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
McCubrey JA, Steelman LS, Chappell WH,
Abrams SL, Wong EW, Chang F, Lehmann B, Terrian DM, Milella M,
Tafuri A, et al: Roles of the Raf/MEK/ERK pathway in cell growth,
malignant transformation and drug resistance. Biochim Biophys Acta.
1773:1263–1284. 2007. View Article : Google Scholar
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Burd EM: Human papillomavirus and cervical
cancer. Clin Microbiol Rev. 16:1–17. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nour NM: Cervical cancer: A preventable
death. Rev Obstet Gynecol. 2:240–244. 2009.
|
|
24
|
Zhou W, Song F, Wu Q, Liu R, Wang L, Liu
C, Peng Y, Mao S, Feng J and Chen C: miR-217 inhibits
triple-negative breast cancer cell growth, migration, and invasion
through targeting KLF5. PLoS One. 12:e01763952017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tian YW, Shen Q, Jiang QF, Wang YX, Li K
and Xue HZ: Decreased levels of miR-34a and miR-217 act as
predictor biomarkers of aggressive progression and poor prognosis
in hepatocellular carcinoma. Minerva Med. 108:108–113. 2017.
|
|
27
|
Azam AT, Bahador R, Hesarikia H, Shakeri M
and Yeganeh A: Downregulation of microRNA-217 and microRNA-646 acts
as potential predictor biomarkers in progression, metastasis, and
unfavorable prognosis of human osteosarcoma. Tumour Biol.
37:5769–5773. 2016. View Article : Google Scholar
|
|
28
|
Zhang Q, Yuan Y, Cui J, Xiao T and Jiang
D: MiR-217 promotes tumor proliferation in breast cancer via
targeting DACH1. J Cancer. 6:184–191. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu YP, Sun XH, Cao XL, Jiang WW, Wang XX,
Zhang YF and Wang JL: MicroRNA-217 suppressed
epithelial-to-mesenchymal transition in gastric cancer metastasis
through targeting PTPN14. Eur Rev Med Pharmacol Sci. 21:1759–1767.
2017.PubMed/NCBI
|
|
30
|
Wang LP, Wang JP and Wang XP: HOTAIR
contributes to the growth of liver cancer via targeting miR-217.
Oncol Lett. 15:7963–7972. 2018.PubMed/NCBI
|
|
31
|
Salehi B, Zucca P, Sharifi-Rad M, Pezzani
R, Rajabi S, Setzer WN, Varoni EM, Iriti M, Kobarfard F and
Sharifi-Rad J: Phytotherapeutics in cancer invasion and metastasis.
Phytother Res. 32:1425–1449. 2018. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cascio S and Finn OJ: Intra- and
extra-cellular events related to altered glycosylation of MUC1
promote chronic inflammation, tumor progression, invasion, and
metastasis. Biomolecules. 6:E392016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee CH, Lin YF, Chen YC, Wong SM, Juan SH
and Huang HM: MPT0B169 and MPT0B002, new tubulin inhibitors, induce
growth inhibition, G2/M cell cycle arrest, and apoptosis in human
colorectal cancer cells. Pharmacology. 102:262–271. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tian X, Han Z, Zhu Q, Tan J, Liu W, Wang
Y, Chen W, Zou Y, Cai Y, Huang S, et al: Silencing of cadherin-17
enhances apoptosis and inhibits autophagy in colorectal cancer
cells. Biomed Pharmacother. 108:331–337. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qian W, Lv S, Li J, Chen K, Jiang Z, Cheng
L, Zhou C, Yan B, Cao J, Ma Q and Duan W: Norepinephrine enhances
cell viability and invasion, and inhibits apoptosis of pancreatic
cancer cells in a Notch-1-dependent manner. Oncol Rep.
40:3015–3023. 2018.PubMed/NCBI
|
|
36
|
Soleimani A, Bahreyni A, Roshan MK,
Soltani A, Ryzhikov M, Shafiee M, Soukhtanloo M, Jaafari MR,
Mashkani B and Hassanian SM: Therapeutic potency of pharmacological
adenosine receptors agonist/antagonist on cancer cell apoptosis in
tumor microenvironment, current status, and perspectives. J Cell
Physiol. 234:2329–2336. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Orton RJ, Sturm OE, Vyshemirsky V, Calder
M, Gilbert DR and Kolch W: Computational modelling of the
receptor-tyrosine-kinase-activated MAPK pathway. Biochem J.
392:249–261. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kyriakis JM: Making the connection:
Coupling of stress-activated ERK/MAPK
(extracellular-signal-regulated kinase/mitogen-activated protein
kinase) core signalling modules to extracellular stimuli and
biological responses. Biochem Soc Symp. 64:29–48. 1999.PubMed/NCBI
|
|
39
|
Zhang Y, Meng L, Xiao L, Liu R, Li Z and
Wang YL: The RNA-binding protein PCBP1 functions as a tumor
suppressor in prostate cancer by inhibiting mitogen activated
protein kinase 1. Cell Physiol Biochem. 48:1747–1754. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang N, Lu C and Chen L: miR-217
regulates tumor growth and apoptosis by targeting the MAPK
signaling pathway in colorectal cancer. Oncol Lett. 12:4589–4597.
2016. View Article : Google Scholar
|
|
41
|
Hu L, Wu H, Wan X, Liu L, He Y, Zhu L, Liu
S, Yao H and Zhu Z: MicroRNA-585 suppresses tumor proliferation and
migration in gastric cancer by directly targeting MAPK1. Biochem
Biophys Res Commun. 499:52–58. 2018. View Article : Google Scholar : PubMed/NCBI
|