1. Introduction
Coronavirus disease 2019 (COVID-19) is caused by a
novel coronavirus: Severe acute respiratory syndrome coronavirus 2
(SARS-CoV-2) (1,2). After the first case of COVID-19 was
diagnosed in the city of Wuhan, China, in December, 2019, the
disease spread rapidly and was declared a pandemic by the World
Health Organization (WHO) on March 11, 2020 (3). As the COVID-19 outbreak advances
worldwide, there is a growing need to identify strategies with
which to combat the virus.
Briefly, the cell entry of SARS-CoV-2 occurs after
its spike (S) proteins bind to the human angiotensin-converting
enzyme 2 (ACE2) receptor, which is expressed in several cell types,
such as lung alveolar and nasal epithelial cells, enterocytes and
vascular endothelial cells (4,5).
After binding ACE2, the S proteins are cleaved by SARS-CoV-2
entry-activating proteases from the host cell, including type II
transmembrane protease serine 2 (TMPRSS2), exposing fusion peptide
domains in a process termed S protein priming. Once membrane fusion
occurs, the SARS-CoV-2 RNA genome is released into the host cell,
triggering viral replication and pathogenic events (4,5).
Some nutrients have the ability to modulate the
innate and acquired immune systems, and play important roles in
antiviral resistance. The finding of possible synergistic actions
between nutrients, as well as between nutrients and drugs, may
allow the development of complex formulations that act differently
from the individual components. In this context, certain specific
nutrients, namely zinc, vitamin D and glutamine have been
considered. These nutrients are referenced as potential adjuvants
in the prevention and treatment of viral infections (6-9).
Moreover, emerging evidence provided in the literature has
consistently highlighted the potential role of these nutrients, as
well as the immunotherapeutic agent OncoTherad, in the prevention
or therapy of COVID-19 (10-27). The combination of these nutrients
with OncoTherad aims to provide specific metabolic support that,
when coordinated with their individual antiviral effects, may
represent a putative therapeutic and/or preventive action against
COVID-19.
A common mechanism among these compounds, which
already exhibit an important synergy, is their ability to modulate
the signaling pathways for interferons (IFNs), which are cytokines
recognized for their antiviral properties (8,28-35). This area of metabolic confluence,
dependent on these nutrients, also corresponds to a synthetic
nanocompound known as OncoTherad, developed as an antineoplastic
and immunomodulator used as therapy for non-muscle invasive bladder
cancer (NMIBC) (36-38). With the objective of designing a
joint use, considering a possible enhancement of their
efficiencies, the present review article discusses the common
properties of these compounds (zinc, vitamin D, glutamine and
OncoTherad), considering that when a deficiency occurs, nutrients
may become limiting factors of the action of other nutrients and
drugs (39).
Toll-like receptor (TLR) and interferon
signaling pathways
Once cells are infected by a virus, viral
pathogen-associated molecular patterns (PAMPs) can be recognized by
several intracellular pathogen recognition receptors (PRRs), such
as TLRs. For example, TLR2, TLR3, TLR4, TLR7, TLR8 and TLR9 play
important roles in the recognition and detection of nucleic acids
(RNA and DNA) specific to the viral genome (40). Some viruses can avoid recognition
by TLRs and may be recognized by other intracellular PRRs present
in the cytosol (40).
Despite the different types of PRRs that can be
activated, the antiviral immune response following the PAMP-PRR
interaction involves common signaling pathways. In general, the
recognition of viral pathogens is associated with the production of
several inflammatory cytokines, in particular IFNs, that have
important antiviral and anti-proliferative activity (40).
IFNs are a group of cytokines secreted by infected
or immune cells that perform critical functions in the immune
system, such as the activation of natural killer (NK) cells and
macrophages (40). These
cytokines are the first line of defense against viral infections.
The binding of IFNs to their cell surface receptors leads to the
activation of the Janus-activated kinase (JAK)/signal transducer
and activator of transcription (STAT) signaling pathway, which
induces the expression of several genes that fight viral
replication in the cell itself and in neighboring cells (28,41). Type I IFNs include IFN-α and
IFN-β, while the type II IFN class has IFN-γ as its only member
(42). In turn, type III IFN
(IFN-λ) is a recently discovered distinct class that, despite
activating a complex of different receptors, activates the same
JAK/STAT signaling pathway (43).
In addition to the key role in antiviral immune
responses, IFNs may also exert anti-inflammatory effects associated
with the inhibition of the production of pro-inflammatory
cytokines, such as interleukin (IL)-1, IL-18 and IL-12 and/or
increasing the production of the anti-inflammatory cytokine IL-10
(44-49). Although a rapid and effective
immune response is essential to limit viral spread, this antiviral
response needs to be controlled to limit tissue damage. It is known
that excessively high levels of serum pro-inflammatory cytokines
and chemokines (cytokine storm) produce an uncontrolled systemic
inflammatory response that contributes to the increased severity of
viral infections, such as those caused by SARS-CoV-2, influenza
virus, Middle East respiratory syndrome coronavirus (MERS-CoV) and
severe acute respiratory syndrome coronavirus (SARS-CoV). This is
one of the major mechanisms responsible for acute respiratory
distress syndrome (ARDS), the primary cause of mortality from
COVID-19 (49-55). Thus, IFNs can play a role in the
treatment of COVID-19, not only through their antiviral activity,
but also by counterbalancing the cytokine storm, which is actually
observed in acute infection by the influenza A virus (49). Other mechanisms through which
these compounds could potentially counteract the cytokine storm,
specifically related to their well-known immunomodulatory effects,
and other aspects of COVID-19 not covered in the present review are
further discussed elsewhere (11,13,15,16,19-21,23,25,26).
Although the mechanisms of SARS-CoV-2 are not yet
well known, its genetic (approximately 79%) and structural
similarity with SARS-CoV, as well as the similar pathogenic
mechanism of cell entry via interaction with ACE2, and the
comparable pattern of mortality by age group suggest that previous
findings on SARS may be partially applicable to COVID-19 (56,57). In SARS, there are abnormally low
levels of antiviral cytokines, particularly type I IFNs (58). In addition, in severe respiratory
diseases caused by the coronavirus subtypes SARS-CoV and MERS-CoV,
the reaction to viral infections by type I IFNs is suppressed. Both
types of coronaviruses use varying strategies to evade the
antiviral defense system of IFNs, such as inhibition of the
production of type I IFN by infected cells, interference in the IFN
signaling pathway and increased resistance to IFNs. These
suppression approaches are highly associated with disease severity
and an increased mortality (58-60).
In fact, it has been observed that the reduction in
IFN-γ expression in CD4+ T cells is related to disease
severity in patients with COVID-19 (61). A recent study that used ex
vivo human lung tissue samples found that SARS-CoV-2, although
replicating and infecting more efficiently than SARS-CoV, did not
lead to the expression of type I, II or III IFNs (62).
The use of IFNs for the treatment of viral
infectious diseases has been shown to be effective (63). For example, type I IFNs are part
of the standard treatment for chronic infections by hepatitis B and
C viruses (63). Numerous studies
have also reported the success of the direct administration of IFNs
against coronavirus (64-72). It has been shown that a
combination of type I IFN and IFN-γ synergistically inhibits viral
replication in vitro (73-76). Thus, the induction of IFNs has
immense potential in the defense of organisms against coronavirus
infections. Several clinical trials are currently underway to
investigate the efficacy of IFN treatments in patients with
COVID-19 (NCT04385095, NCT04350671, NCT04465695, NCT04534673,
NCT04480138, NCT04492475, NCT04354259, NC T 0 4 49439 9, NC T 0
4254874, NC T 0 432 4 4 63, NC T 0 4315948, NC T 0 434 4 6 0 0, NC
T 0 4276 688, ChiCTR2000029387 and ChiCTR2000029600), including
cancer patients (NCT04379518, NCT04534725) (77).
OncoTherad
The synthetic nanocompound, Biological Response
Modifier - Inorganic Phosphate Complex 1 (MRB-CFI-1, for its
acronym in Portuguese), registered as OncoTherad, is a nanometric
compound of metallic salts and phosphate and associated with a
glycosidic protein, 420-530 nm in size (36-38,78-80). This novel nano-immunotherapy
stimulates the immune system to eliminate cancer and could become a
promising strategy against SARS-CoV-2. The immunotherapy drug is
being developed by researchers at the University of Campinas in the
state of São Paulo, Brazil, and has patents deposited in Brazil
(no. BR1020170127680) (38), USA
(no. US20200156951A1) (80) and
Europe (no. WO2018227261) (81).
It is being tested in clinical trial (Brazilian Clinical Trials
Registry -RBR-6swqd2, UTN U1111-1226-9096) and is still not
available on the market.
OncoTherad is a novel intravesical therapy with
immunological and antitumor properties for the treatment of NMIBC.
The primary treatment for NMIBC is based on surgical transurethral
resection followed by intravesical immunotherapy with bacillus
Calmette-Guérin (BCG) to avoid tumor progression and decrease
recurrence. Preclinical, clinical-veterinary and phase I/II
clinical trials have shown that nano-immunotherapy with OncoTherad
leads to the distinct stimulation of the innate immune system
mediated by TLR2 and TLR4, resulting in an increased activation of
the IFN signaling pathway [TLR4, TIR-domain-containing
adapter-inducing IFN-β (TRIF), interferon regulatory factor
(IRF)-3, IFN-α and -γ], which is associated to the superior
efficacy of this nanocompound in the management of NMIBC compared
to the standard treatment with BCG (36-38,82).
More specifically, the OncoTherad-induced
stimulation of the immune system via TLR2 and TLR4 occurs through
the phosphorylation of hydroxylated amino acids, such as tyrosine,
threonine and serine by compounds that contain phosphate salts
(36-38,78-80), resulting in the activation of
stimulator of interferon genes (STING), with a consequent increase
in the production of IFN-α and IFN-γ. The increase in the
production of IFNs mediated by TLRs-2 and 4 promotes the activation
of TCD8+ cells, dendritic cells and M1 macrophages,
culminating in the superior effectiveness of OncoTherad in the
therapy of bladder cancer when compared to BCG (37,38,78,80). Experiments with rodents (36,38,80) and dogs (38,79,80) have demonstrated that OncoTherad is
able to decrease the expression of receptor activator of nuclear
factor-κB (RANK) and receptor activator of nuclear factor-κB ligand
(RANK-L) system and, as a result, prevent the formation of
metastases and/or counteract their progression. In addition, in the
treatment of bladder cancer chemically induced in Fisher 344 rats,
OncoTherad nano-immunotherapy was shown to increase the
immunoreactivities of regulatory proteins, such as phosphatase and
tensin homolog (PTEN) and p21, demonstrating the positive
contribution of this immunotherapy in the regulation of the cell
cycle (80).
A previous study demonstrated that the stimulation
of TLR4 resulted in protective immune response and TRIF-related
adaptor molecule (TRAM) knockout mice were more susceptible to
coronavirus infection and expressed molecular signs similar to
those of SARS-CoV and MERS-CoV with a poor prognosis (83). Furthermore, SARS-CoV may interfere
on IRF-3 signaling, delaying IFN activity (60). Therefore, considering the
immunomodulation promoted by OncoTherad and the role of the IFN
signaling pathway in the control of COVID-19, this compound emerges
as a promising therapeutic option for SARS-CoV-2 infection control
and reduction of lung injury severity.
In 5 patients who developed severe COVID-19 while
undergoing treatment for NMIBC, the administration of OncoTherad
with corticosteroids and antibiotics mitigated the exacerbated
inflammatory response in the lungs, decreased the average hospital
stay from 18 to 10 days, and prevented the need for intubation
(17,84).
A 78-year-old Brazilian male enrolled on the
OncoTherad clinical trial (Brazilian Clinical Trials
Registry-RBR-6swqd2) for the treatment of BCG-refractory or
relapsed NMIBC was the most emblematic case. The patient with a
history of former tabagism (34 years), systemic arterial
hypertension and previous revascularization of myocardium, arrived
at the Hospital Municipal de Paulinia in Brazil, presenting with
COVID-19 symptoms (84). He
reported a dry cough, inappetence, coryza and malaise, which
appeared immediately following a cruise trip. The diagnosis of
SARS-CoV-2 was confirmed with local RT-PCR, IgM/IgG serological
rapid screening and a chest CT scan (84).
Before exhibiting symptoms of COVID-19, the patient
was diagnosed with high-grade papillary urothelial carcinoma. In
January, 2020, due to a refractory tumor, the patient was enrolled
in the OncoTherad clinical trial, developed at the University of
Campinas. Consecutive weekly applications of OncoTherad (one
intravesical and one intramuscularly applied per week, for 4 weeks)
were used to treat the cancer before the patient interrupted the
treatment for 2 weeks for travel purposes. On April 1, 2020,
following 24 h of having flu-like symptoms, the patient's clinical
condition evolved to fever (38.3°C), headache, dyspnea
(pO2, 66.4 mmHg; SatO2, 87%; respiratory
frequency, 21 bpm) and basilar crackling and stertors in the lungs.
A chest CT scan revealed areas of ground-glass opacities on lung
parenchyma with bilateral distribution and predominance on right
upper and middle lobes, pointing to an evolving acute lung
inflammatory process (84).
OncoTherad immunotherapy was resumed to treat the
cancer condition with one intramuscular application. The COVID-19
treatment regimen included oxygen therapy (nasal catheter flux 4
liters/min), Ceftriaxone 2 g (for 6 days, intravenously),
Azithromycin 500 mg (for 6 days, intravenously) and Tamiflu 75 mg
[twice a day (b.i.d.), for 5 days, orally]. At 72 h following
hospitalization, the symptoms of coryza and cough improved, with
the absence of fever. However, the patient presented continued
inappetence and fatigue on minimal exertions. Oxygen therapy was
maintained, and, after 6 days of hospitalization, the patient
exhibited a significant improvement in pulmonary inflammation
(SatO2, 94% at room air), with the withdrawal of oxygen
therapy and significant clinical improvement on the 8th day
(pO2, 96.2 mmHg; SatO2, 98%; decreased
basilar crackling and stertors in the lungs, absence of fever,
attenuated cough and appetite enhancement). The patient was
discharged in a good general condition on the 10th day of
hospitalization (84). Patient
progression and recovery occurred in a shorter time period (10
days) than reported in a previous study (85). As previously demonstrated, 86% of
patients enrolled in a Chinese study were discharged after 16 days
of hospitalization; the majority of individuals exhibited
radiological worsening on the 7th day and improvement on the 14th
day (85).
A new chest CT scan was performed on patient
discharge and a comparative analysis with the chest CT acquired
upon admission revealed areas of periphery pulmonary consolidations
and fibrous stripes. Areas of ground-glass opacities were no longer
observed. Moreover, treatment-related adverse events were Grade 1
or 2, such as pruritus and rash (84).
In addition, as regards the case report of
immunotherapy with OncoTherad, Delafiori et al (84) identified 33 molecules related to
the metabolic state at admission and 39 after improvement of the
clinical condition and discharge. The authors revealed extensive
lipid dysregulation with marked disturbances in the metabolism of
glycerophospholipids, glycerolipids, lipid sterols and fatty acids.
Several fatty acids species were identified in that study under
admission and discharge conditions, including mediators of linoleic
and arachidonic acid metabolism, which play a crucial role in the
response to inflammation (84).
Notably, the other 4 patients subjected to the
clinical protocol of OncoTherad therapy for NMIBC who tested
positive for COVID-19 exhibited a similar outcome to that of the
patient described in the aforementioned case report. Their recovery
was more rapid (10 days) when compared to the observed for patients
subjected to the standard treatment only (17).
Considering the comorbidities and age of the
patients, it is considered that immunotherapy with OncoTherad
exerted a protective effect in this patient, preventing the
progression of infection to the most severe states and promoting
fast recovery without the need for admission to the intensive care
unit (84). A study with 140
participants for the use of OncoTherad immunotherapy associated
with standard clinical treatment for COVID-19 is underway at
Hospital Municipal de Paulinia. The study is expected to last for 1
year (17).
Studies have indicated that some nutrients,
particularly zinc, vitamin D and glutamine, play an important role
in the immune system and also modulate the IFN signaling pathway,
as described below, which may suggest a possible synergistic action
with OncoTherad.
2. Zinc
Several studies have indicated the antiviral
activity of zinc against several viruses, such as SARS-CoV,
rhinovirus, influenza, herpes virus, respiratory syncytial virus,
equine arteritis virus, hepatitis E virus and transmissible
gastroenteritis virus (6,86-90). For example, zinc has been shown to
inhibit viral RNA polymerase activity in vitro and viral
replication in Vero-E6 cells infected with SARS-CoV (89). The antiviral mechanisms of action
of zinc include the modulation of the immune system of the infected
individual, with increased signaling associated with IFNs. These
cytokines also stimulate the influx of zinc into the target cell,
which highlights the importance of this element for IFN signaling
(6).
Zinc induces the production of IFN-α and IFN-γ, and
enhances the antiviral action of IFN-α (29-31). A previous study found that zinc
and its transporter, ZIP8 (SLC39A8), regulated the expression of
IFN-γ in activated human T lymphocytes, which was confirmed by
increased IFN-γ release with ZIP8 overexpression and reduced
release of the cytokine with a reduction in the expression
(knockout) of this zinc transporter (91). The same study demonstrated an
increase in the production of IFN-γ in vitro by activated T
lymphocytes isolated from individuals supplemented with 15 mg/day
zinc orally and an even greater increase after the treatment of
activated T lymphocytes with zinc in vitro (91).
In turn, in human primary lung cells, in the absence
of zinc, treatment with IFN-γ and tumor necrosis factor (TNF)-α and
the activation of Fas receptor signaling may result in cell death
by apoptosis and dysfunction of the pulmonary epithelial barrier,
indicating that zinc is a vital factor in the protection of this
epithelium against acute damage (92).
STAT1 is a key mediator of the IFN signaling
pathway. Reduced zinc bioavailability by the zinc chelating agent,
N,N,N′,N′-tetrakis(2-pyridylmethyl)ethylenediamine (TPEN), has been
shown to significantly reduce the gene expression and protein level
of STAT1 in macrophages, which may be associated with increased
susceptibility to infections in zinc deficiency (93). By contrast, increased zinc
concentrations may restrict the replication of hepatitis E virus,
increasing the level and activity of STAT1 and/or inducing the
antiviral response mediated by IFN-stimulated genes via a mechanism
mediated by the nuclear transcription factor-κB (NF-κB) (90).
Interferon-induced transmembrane proteins (IFITMs)
are known for their antiviral functions. They are crucial for the
IFN-mediated inhibition of a number of enveloped viruses, including
influenza A and Ebola virus (94). Similarly, the zinc
metalloprotease, ZMPSTE24, exhibits antiviral activity against
several enveloped viruses, including those already mentioned
(95,96). Both the IFITM3 and ZMPSTE24
proteins are induced by IFN (90), which in turn is induced by zinc
treatment. Given that SARS-CoV-2 is an enveloped virus, it is
assumed that the increase in IFITM and/or ZMPSTE24 induced by zinc
could be effective in exerting antiviral effects against
COVID-19.
Zinc also acts as an important mediator of the
signaling of several immune cell receptors (97). One of these is the TLR4 receptor,
which, similar to other TLR receptors, plays an important role in
virus recognition and antiviral immune response (40,97). Upon activation, TLR4 binds to the
adaptor protein MyD88 and causes an increase in cytoplasmic levels
of free zinc (Zn2+). This zinc signaling is necessary
for the activation of MyD88-dependent NF-κB, resulting in the
expression of genes targeting this transcription factor, such as
the inflammatory cytokines TNF-α, IL-1β and IL-6 (97). These cytokines perform several
important functions in the adaptive immune response, contributing
to antiviral defense (98).
Clinical studies
A clinical study with 21 healthy elderly individuals
in Italy (mean age, 87 years) found that the consumption of
zinc-fortified milk (3 mg of added zinc plus 1 mg of zinc naturally
present in milk) for 2 months, compared to the consumption of
non-fortified milk, increased cellular immunity, which was
concluded by the increased release of the cytokines, IFN-γ and
IL-12p70, and increased endocrine activity in the thymus (active
thymulin, linked to zinc) (99).
In that study, peripheral blood mononuclear cells (PBMCs) isolated
from the elderly individuals who consumed zinc-fortified milk
following inflammatory stimulation with lipopolysaccharide (LPS)
exhibited a significant increase of 25.7% in the release of IFN-γ,
indicating better cellular immune functioning (99).
These data are in agreement with those of a study on
19 healthy elderly individuals supplemented with 10 mg of elemental
zinc (50 mg of zinc aspartate). PBMCs isolated from the elderly
individuals and stimulated with SPEA in vitro were evaluated
before and after zinc supplementation. Supplementation led to a
significant increase in the release of the pro-inflammatory
cytokines, IFN-γ (53.28%) and TNF-α (80.03%) (P<0.006), and the
anti-inflammatory cytokine, IL-10 (31.79%) (P<0.003). These
results indicated that zinc supplementation led to an improvement
in the immune response of T lymphocytes (100).
3. Vitamin D
Despite the traditional name, vitamin D is actually
a hormone, as in addition to being endogenously produced, it is in
involved in the regulation of >200 genes in different cell types
(101-103). Vitamin D is converted into its
active form (calcitriol) through the action of the enzyme
1α-hydroxylase (CYP27B1). The detection of the expression of this
enzyme and of vitamin D receptors in immune cells and the
observation that the expression of these receptors is regulated by
immune signaling provide evidence of the central role played by
vitamin D in the immune system, as well as the clear association
between low serum levels of 25(OH)D3 and autoimmune diseases,
cancer, cardiovascular diseases and respiratory infections
(40). Respiratory epithelial
cells constitutively express CYP27B1, underscoring their local need
for active vitamin D (7).
Several studies have suggested an important
antiviral role of vitamin D, although the mechanisms are complex
and remain poorly understood (7).
The activation of TLRs by PAMPs in immune cells leads to the
increased expression of the vitamin D receptor and of CYP27B1,
increasing the conversion of circulating vitamin D into its active
form. Thus, the metabolism of vitamin D in these cells is linked to
the recognition of pathogens, rendering it an integral part of the
innate immune response (40,104).
For example, the activation of TLR4 by LPS has been
shown to lead to the increased expression of CYP27B1 in monocytes
(105,106). Similarly, in dendritic cells,
the activation of TLR4 or TLR3 induces the expression of CYP27B1,
altering the migratory properties of these cells to allow the
presentation of antigens to CD4+ T lymphocytes (107-110). In human tracheobronchial
epithelial cells, TLR stimulation by poly I:C and live respiratory
syncytial virus increases the expression of CYP27B1, resulting in
the increased conversion of vitamin D into its active form
(calcitriol) and the increased expression of cathelicidins,
antimicrobial peptides with important antiviral action (111). Similar effects have been
reported for TLR8 and TLR2/1 heterodimer (40).
The cytokines, IFN-γ, IL-13, IL-15, IL-4, IL-1, IL-2
and TNF-α, also regulate the expression of CYP27B1 and vitamin D
metabolism. The signaling pathways involved in this process are
probably the same as those involved in regulation induced by the
activation of TLRs (40,112).
Vitamin D is necessary for the antimicrobial immune
response of human macrophages mediated by IFN-γ production against
Mycobacterium tuberculosis as this antimicrobial response
was not observed with macrophages cultured in human serum of
individuals with insufficient levels of this hormone. It has been
suggested that increased serum levels of vitamin D may improve the
IFN-γ-mediated antimicrobial immune response of human macrophages
(32).
In summary, the studies mentioned above indicate
that the extrarenal synthesis of calcitriol is regulated by TLR
activation and cytokine secretion. Thus, in the presence of
sufficient levels of circulating vitamin D, infection by
respiratory viruses, which results in recognition by TLRs and
cytokine production, is able to increase calcitriol levels,
possibly altering the immune response to optimize the fight against
these pathogens (40).
Importantly, as previously mentioned, SARS-CoV-2
binds to ACE2 for host cell entry. Given that it acts as the viral
receptor, it would be reasonable to hypothesize that the higher
expression of ACE2 may be detrimental for the host (113). However, several lines of
evidence indicate that ACE2 plays a critical role in counteracting
ARDS and acute lung injury (113-116). In fact, the viral binding
reduces ACE2 activity, which in turn increases ACE1 activity due to
the disruption of the ACE2/ACE1 activity ratio, causing elevated
pulmonary vasoconstriction and contributing to the severity of
COVID-19 (117,118). Vitamin D has been shown to
increase the expression of ACE2 in the lungs, which prevents the
pulmonary vasoconstriction response in COVID-19 cases and can
reduce lung injury. In this context, targeting ACE2 downregulation
with vitamin D in SARS-CoV-2 infection may be a potential
therapeutic approach to COVID-19 and induced ARDS (113,117-119). Further studies are warranted to
confirm this hypothesis (27).
Clinical studies
Several clinical trials have indicated that vitamin
D3 supplementation (1,000 or 2,000 IU/day) is effective for
enhancing the efficacy of antiviral treatment with pegylated
interferon and ribavirin (PEG-IFN/RBV) in patients with hepatitis C
(120-123). Conversely, low serum
concentrations of the hormone seem to be related to severe hepatic
fibrosis and/or to the lower responsiveness to treatment with
PEG-IFN/RBV (124,125).
In a study with 50 patients with hepatitis C treated
with PEG-IFN/RBV, the authors reported that low serum levels of
vitamin D were associated with negative treatment results, while
supplementation with this hormone (2,000 IU/day for 24 weeks)
significantly improved the viral response, defined as an
undetectable serum level of viral RNA 24 weeks following the
discontinuation of therapy (122). These results were later
corroborated by a clinical trial with 84 patients with hepatitis C
that also concluded that vitamin D supplementation (1,000 IU/day)
from the eighth week until the end of treatment with PEG-IFN/RBV
may increase the effects of antiviral therapy (123).
4. Glutamine
Glutamine is an amino acid used as a nitrogen source
for rapidly-dividing cells, such as lymphocytes, in which it is
critical for nucleotide synthesis and energy production (126).
T lymphocytes are activated rapidly following their
interaction with pathogens, which requires high energy and
biosynthetic demands (35). These
augmented metabolic needs are met by increased absorption and use
of glutamine, which is essential for the proliferation of these
immune cells (33,35,127). In fact, the activation of naive
T lymphocytes is associated with the rapid glutamine uptake, which
requires the amino acid transporter ASCT2 (128). Glutamine transporters are
upregulated during the activation of T lymphocytes, while a
reduction in these transporters impairs the function of these
immune cells (33,129).
Activated human T lymphocytes also require glutamine
for the production of IFN-γ and IL-2. This compound modulates not
only the proliferation of stimulated PBMCs, but also the secretion
of these cytokines in a concentration-dependent manner (130), and glutamine depletion inhibits
T lymphocyte proliferation, and decreases IL-2 and IFN-γ production
(33,34).
When a naive CD4+ T cell is activated by
antigens, the types of cytokines determine whether this cell will
differentiate into an effector T lymphocyte that performs an immune
response, such as a helper T helper 1 cell (Th1) or a regulatory T
lymphocyte, which suppresses immune responses. Even in the presence
of cytokines that normally promote the generation of Th1 cells,
glutamine depletion favors the regulatory T phenotype rather than
the Th1 phenotype, which is associated with the suspension of IFN-γ
secretion (35). Thus, reduced
glutamine availability alters the balance of the immune response to
become more suppressive (35),
which could impair antiviral immune responses.
Studies have demonstrated that glutamine has
antiviral activity against the herpes virus (8,9).
Following infection with the herpes simplex virus,
glutamine-treated mice (intraperitoneal injection of 8 mg of
glutamine twice daily) were shown to have lower levels of virus,
higher numbers of active CD4 and CD8 cells in the spleen, and
increased levels of IFN-γ in vaginal fluid at 2 and 4 days
post-infection, which was associated with an improvement in the
clinical signs of viral infection (8). By contrast, glutamine depletion
increased the replication of herpes virus (ICP0-null mutant) in
cell culture in vitro (9).
A recent study used advanced bioinformatics to
identify existing therapeutic options for patients with COVID-19
(131). Two computational
approaches were used to identify therapeutic agents already
approved by the Food and Drug Administration (FDA) that could
prevent SARS-CoV-2 from entering cells via ACE2 or TMPRSS2 enzymes
and that could mitigate the gene expression patterns stimulated by
coronavirus. Glutathione and its precursor glutamine were ranked
high by two different methods, suggesting that both justify further
investigation of their potential effects against SARS-CoV-2
(131).
Clinical studies
Low plasma glutamine levels are associated with a
poor clinical outcome in intensive care unit patients (132). Following trauma and surgery,
there is a marked reduction in plasma and muscle glutamine
concentrations and excessive differentiation of T cells towards a
type 2 response. These changes contribute to the state of
immunosuppression and greater susceptibility to infectious
complications, such as pneumonia, sepsis and bacteremia. It has
been observed that the administration of enteral nutrition enriched
with glutamine in patients with severe trauma increases the plasma
concentrations of the amino acid and reduces the incidence of
infectious complications, in addition to improving a variety of
immunological parameters (133-138).
A clinical study investigated the effects of enteral
nutrition enriched with glutamine compared to a control isocaloric
enteral diet on the responses of T lymphocytes during the first 2
weeks after severe traumatic injuries in 28 patients (injury
severity score >20) (138). A
total of 17 healthy volunteers were used as controls. On the first
day, it was observed that T lymphocytes isolated from trauma
patients, compared to healthy individuals, exhibited a low IFN-γ
production following stimulation with phytohemagglutinin (PHA)
in vitro. After 2 weeks, glutamine nutrition significantly
augmented the production of IFN-γ, in addition to preventing the
decrease in IL-4 production by T lymphocytes stimulated in
vitro. In conclusion, trauma caused the suppression of the
cellular immune response, while glutamine supported the restoration
of the lymphocyte response capacity by increasing IFN-γ production
and maintaining normal IL-4 production (138).
Finally, a Cochrane systematic review and
meta-analysis of 57 clinical trials evaluated the effect of
glutamine supplementation in a total of 4,671 adult patients with
serious diseases or undergoing major surgery. The results revealed
that the risk of infectious complications in subjects who received
glutamine was 21% lower (RR, 0.79; 95% CI, 0.71 to 0.87;
P<0.00001) than that in patients who did not receive glutamine.
In addition, glutamine supplementation reduced the mean duration of
hospitalization and the mean duration of mechanical ventilation by
3.46 (95% CI, -4.61 to -2.32; P<0.0001) and 0.69 (95% CI, -1.37
to -0.02; P=0.04) days, respectively (139).
5. Conclusion
With the advancement of the COVID-19 outbreak, there
is an urgent need for the identificsation of strategies with which
to combat SARS-CoV-2 infection. Immunotherapy with OncoTherad has
been found to be safe and effective in the treatment of NMIBC by
increasing the signaling associated with IFNs, cytokines that play
essential functions in the activation of the immune system and
antiviral responses. Taken together, the available evidence
indicates that zinc, vitamin D and glutamine are essential in
antiviral defenses, with an emphasis on the participation of these
nutrients in IFN signaling, suggesting an adjuvant and synergistic
therapeutic potential with OncoTherad in modulating immune function
in the treatment of COVID-19 (Fig.
1). Thus, the possible beneficial role in this viral infection
justifies the need for preclinical and clinical studies to evaluate
the combination of these compounds and standardize the dosages that
could provide greater benefits to patients.
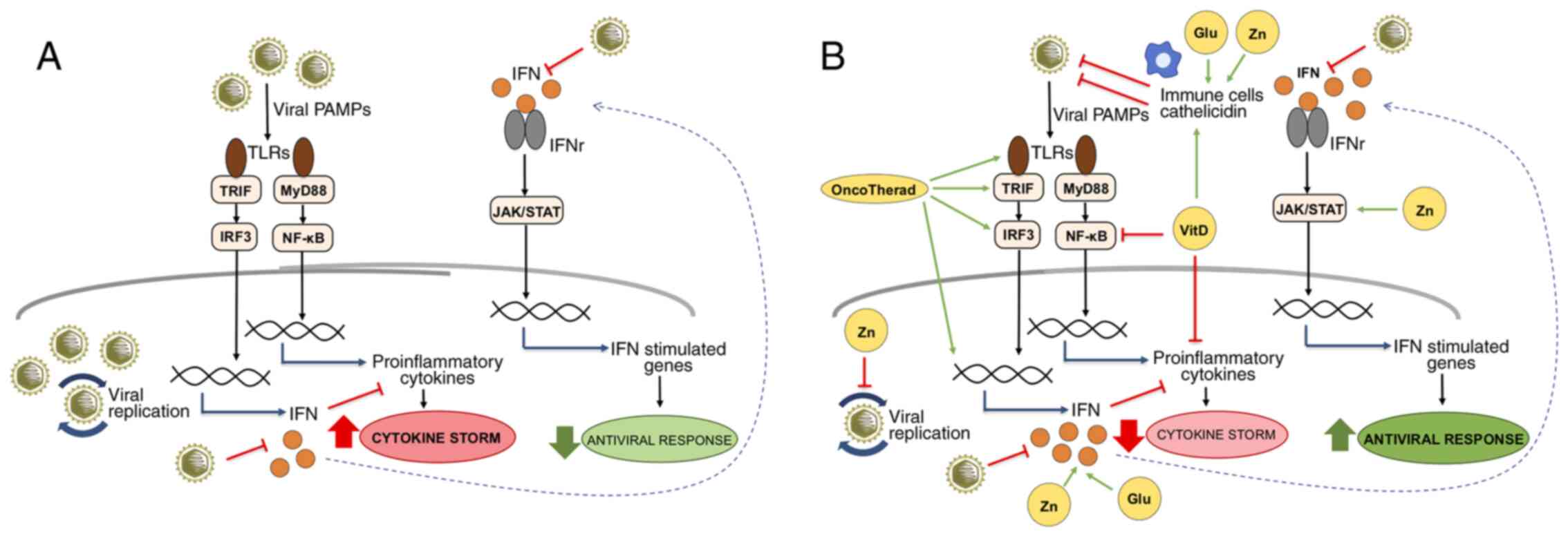 | Figure 1IFN-related signaling pathways
modulated by OncoTherad, zinc, vitamin D and glutamine during viral
infections. (A) Viral PAMPs are recognized by TLRs, triggering
signaling pathways mediated by TRIF and MyD88. The TRIF/IRF3
pathway activates the expression of IFNs that, when interacting
with their receptors, activate the JAK/STAT pathway and the
expression of genes with important antiviral functions. On the
other hand, the MyD88/NF-κB pathway is responsible for the
expression of pro-inflammatory genes, such as cytokines, which,
when in excess, contribute to the cytokine storm characterized by
uncontrolled inflammation. SARS-CoV-2 is known for inducing
cytokine storm associated with severe forms of COVID-19, as well as
for inhibiting the production of IFNs and evading the IFN-related
antiviral defense system. (B) The compounds OncoTherad, zinc,
vitamin D and glutamine can contribute to the organism's antiviral
defenses by several mechanisms: OncoTherad increases the
TLR4/TRIF/IRF3/INF signaling pathway; zinc inhibits viral
replication, activates STAT1, stimulates IFN production and induces
the antiviral responses mediated by IFN-stimulated genes; vitamin D
inhibits NF-κB and the expression of its pro-inflammatory target
genes, as well as stimulates immune cells and the expression of
catelicidin with antiviral activity; and glutamine is essential for
the proliferation of immune cells and the production of IFN.
Together, these compounds can help to attenuate inflammation
associated with the cytokine storm and to increase the organism's
antiviral status. The green arrows represent activation, the red
truncated lines indicate inhibition and the blue arrows indicate
activation of gene expression. Glu, glutamine; IFN, interferon;
IFNr, interferon receptor; JAK/STAT, Janus-activated kinase/signal
transducer and activator of transcription proteins; MyD88, myeloid
differentiation factor 88; NF-κB, nuclear factor-κB; PAMP,
pathogen-associated molecular pattern; TLR, Toll-like receptor;
TRIF, Toll/IL-1 receptor domain-containing adaptor inducing IFN-β;
VitD, vitamin D; Zn, zinc. |
Funding
The present study was supported by Coordenação de
Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
(313152/2018-7) to WJF.
Availability of data and materials
Not applicable.
Authors' contributions
JJN contributed to the study concept and critically
reviewed the article. ARV contributed to the design of the
manuscript and figure preparation and edition. JJN, ARV, ACRS and
WJF contributed to literature review and drafted the manuscript.
All authors gave the final approval for all aspects of the work,
agree to be fully accountable for ensuring the integrity and
accuracy of the work, and read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Ms. Paula Mitie
Hirata from Kilyos Nutrition for providing technical assistance
with the editing of the figure.
References
|
1
|
Zhou P, Yang XL, Wang XG, Hu B, Zhang L,
Zhang W, Si HR, Zhu Y, Li B, Huang CL, et al: A pneumonia outbreak
associated with a new coronavirus of probable bat origin. Nature.
579:270–273. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gorbalenya AE, Baker SC, Baric RS, de
Groot RJ, Drosten C, Gulyaeva AA, Haagmans BL, Lauber CL,
Leontovich AM, Neuman BW, et al: Severe acute respiratory
syndrome-related coronavirus: The species and its viruses- A
statement of the Coronavirus Study Group. Microbiology. 2020.
|
|
3
|
Cucinotta D and Vanelli M: WHO declares
COVID-19 a pandemic. Acta Biomed. 91:157–160. 2020.PubMed/NCBI
|
|
4
|
Shang J, Wan Y, Luo C, Ye G, Geng Q,
Auerbach A and Li F: Cell entry mechanisms of SARS-CoV-2. Proc Natl
Acad Sci USA. 117:11727–11734. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nitulescu GM, Paunescu H, Moschos SA,
Petrakis D, Nitulescu G, Ion GND, Spandidos DA, Nikolouzakis TK,
Drakoulis N and Tsatsakis A: Comprehensive analysis of drugs to
treat SARSCoV2 infection: Mechanistic insights into current COVID19
therapies (Review). Int J Mol Med. 46:467–488. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Read SA, Obeid S, Ahlenstiel C and
Ahlenstiel G: The role of zinc in antiviral immunity. Adv Nutr.
10:696–710. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Teymoori-Rad M, Shokri F, Salimi V and
Marashi SM: The interplay between vitamin D and viral infections.
Rev Med Virol. 29:e20322019. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Uyangaa E, Lee HK and Eo SK: Glutamine and
leucine provide enhanced protective immunity against mucosal
infection with herpes simplex virus type 1. Immune Netw.
12:196–206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bringhurst RM, Dominguez AA and Schaffer
PA: Glutamine deprivation causes enhanced plating efficiency of a
herpes simplex virus type 1 ICP0-null mutant. J Virol.
82:11472–11475. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arvinte C, Singh M and Marik PE: Serum
levels of vitamin C and vitamin D in a cohort of critically ill
COVID-19 patients of a North American Community Hospital Intensive
Care Unit in May 2020: A pilot study. Med Drug Discov.
8:1000642020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Alexander J, Tinkov A, Strand TA, Alehagen
U, Skalny A and Aaseth J: Early nutritional interventions with
zinc, selenium and vitamin D for raising anti-viral resistance
against progressive COVID-19. Nutrients. 12:23582020. View Article : Google Scholar :
|
|
12
|
Bauer SR, Kapoor A, Rath M and Thomas SA:
What is the role of supplementation with ascorbic acid, zinc,
vitamin D, or N-acetylcysteine for prevention or treatment of
COVID-19? Cleve Clin J Med. Jun 8–2020.Epub ahead of print.
View Article : Google Scholar
|
|
13
|
Benskin LL: A basic review of the
preliminary evidence that COVID-19 risk and severity is increased
in vitamin D deficiency. Front Public Health. 8:5132020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brewer J, Gomez Marti JL and Brufsky A:
Potential interventions for SARS-CoV-2 infections: Zinc showing
promise. J Med Virol. Sep 17–2020.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Calder PC, Carr AC, Gombart AF and
Eggersdorfer M: Optimal nutritional status for a well-functioning
immune system is an important factor to protect against viral
infections. Nutrients. 12:11812020. View Article : Google Scholar :
|
|
16
|
Chakhtoura M, Napoli N and El Hajj
Fuleihan G: Commentary: Myths and facts on vitamin D amidst the
COVID-19 pandemic. Metabolism. 109:1542762020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Durán N and Fávaro WJ: Immunomodulators
acting on covid-19: Actual knowledge and perspectives. J Appl
Microb Res. 3:37–44. 2020.
|
|
18
|
Ebadi M and Montano-Loza AJ: Perspective:
Improving vitamin D status in the management of COVID-19. Eur J
Clin Nutr. 74:856–859. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ferrara F, De Rosa F and Vitiello A: The
central role of clinical nutrition in COVID-19 patients during and
after hospitalization in intensive care unit. SN Compr Clin Med.
1–5. 2020.
|
|
20
|
Grant WB, Lahore H, McDonnell SL, Baggerly
CA, French CB, Aliano JL and Bhattoa HP: Evidence that vitamin D
supplementation could reduce risk of influenza and COVID-19
infections and deaths. Nutrients. 12:9882020. View Article : Google Scholar :
|
|
21
|
Jovic TH, Ali SR, Ibrahim N, Jessop ZM,
Tarassoli SP, Dobbs TD, Holford P, Thornton CA and Whitaker IS:
Could vitamins help in the fight against COVID-19? Nutrients.
12:25502020. View Article : Google Scholar :
|
|
22
|
Siuka D, Pfeifer M and Pinter B: Vitamin D
supplementation during the COVID-19 pandemic. Mayo Clin Proc.
95:1804–1805. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Skalny AV, Rink L, Ajsuvakova OP, Aschner
M, Gritsenko VA, Alekseenko SI, Svistunov AA, Petrakis D, Spandidos
DA, Aaseth J, et al: Zinc and respiratory tract infections:
Perspectives for COVID19 (Review). Int J Mol Med. 46:17–26.
2020.PubMed/NCBI
|
|
24
|
Xu Y, Baylink DJ, Chen CS, Reeves ME, Xiao
J, Lacy C, Lau E and Cao H: The importance of vitamin d metabolism
as a potential prophylactic, immunoregulatory and neuroprotective
treatment for COVID-19. J Transl Med. 18:3222020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Weir EK, Thenappan T, Bhargava M and Chen
Y: Does vitamin D deficiency increase the severity of COVID-19?
Clin Med (Lond). 20:e107–e108. 2020. View Article : Google Scholar
|
|
26
|
Zhang J, McCullough PA and Tecson KM:
Vitamin D deficiency in association with endothelial dysfunction:
Implications for patients with COVID-19. Rev Cardiovasc Med.
21:339–344. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bergman P: The link between vitamin D and
COVID-19: Distinguishing facts from fiction. J Intern Med. Jul
11–2020.Epub ahead of print.
|
|
28
|
Totura AL and Bavari S: Broad-spectrum
coronavirus antiviral drug discovery. Expert Opin Drug Discov.
14:397–412. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Berg K, Bolt G, Andersen H and Owen TC:
Zinc potentiates the antiviral action of human IFN-alpha tenfold. J
Interferon Cytokine Res. 21:471–474. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Foster M and Samman S: Zinc and regulation
of inflammatory cytokines: Implications for cardiometabolic
disease. Nutrients. 4:676–694. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cakman I, Kirchner H and Rink L: Zinc
supplementation reconstitutes the production of interferon-alpha by
leukocytes from elderly persons. J Interferon Cytokine Res.
17:469–472. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fabri M, Stenger S, Shin DM, Yuk JM, Liu
PT, Realegeno S, Lee HM, Krutzik SR, Schenk M, Sieling PA, et al:
Vitamin D is required for IFN-gamma-mediated antimicrobial activity
of human macrophages. Sci Transl Med. 3:104ra1022011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Carr EL, Kelman A, Wu GS, Gopaul R,
Senkevitch E, Aghvanyan A, Turay AM and Frauwirth KA: Glutamine
uptake and metabolism are coordinately regulated by ERK/MAPK during
T lymphocyte activation. J Immunol. 185:1037–1044. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hörig H, Spagnoli GC, Filgueira L, Babst
R, Gallati H, Harder F, Juretic A and Heberer M: Exogenous
glutamine requirement is confined to late events of T cell
activation. J Cell Biochem. 53:343–351. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Klysz D, Tai X, Robert PA, Craveiro M,
Cretenet G, Oburoglu L, Mongellaz C, Floess S, Fritz V, Matias MI,
et al: Glutamine-dependent alpha-ketoglutarate production regulates
the balance between T helper 1 cell and regulatory T cell
generation. Sci Signal. 8:ra972015. View Article : Google Scholar
|
|
36
|
Fávaro WJ, Iantas SR, Gonçalves JM, Dias
QC, Reis IB, Billis A, Duran N and Alonso JC: Role of OncoTherad
immunotherapy in the regulation of toll-like receptors-mediated
immune system and RANK/RANKL signaling: New therapeutic perspective
for non-muscle invasive bladder cancer. J Clin Oncol.
37:e160042019. View Article : Google Scholar
|
|
37
|
Durán N, Dias QC and Fávaro WJ:
OncoTherad: A new nanobiological response modifier, its
toxicological and anticancer activities. J Phys Conf Ser. Oct
2–2019.Epub ahead of print. View Article : Google Scholar
|
|
38
|
Fávaro W and Durán N: Process of obtaining
a nanostructured complex (CFI-1), associated to nanostructured
CFI-1 with a protein (MRB-CFI-1) and its use. Patent
BR1020170127680. June 14–2017
|
|
39
|
Challem JJ: Toward a new definition of
essential nutrients: Is it now time for a third 'vitamin' paradigm?
Med Hypotheses. 52:417–422. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Greiller CL and Martineau AR: Modulation
of the immune response to respiratory viruses by vitamin D.
Nutrients. 7:4240–4270. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Platanias LC: Mechanisms of type-I- and
type-II-interferon-mediated signalling. Nat Rev Immunol. 5:375–386.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Le Page C, Génin P, Baines MG and Hiscott
J: Interferon activation and innate immunity. Rev Immunogenet.
2:374–386. 2000.
|
|
43
|
O'Brien TR, Thomas DL, Jackson SS,
Prokunina-Olsson L, Donnelly RP and Hartmann R: Weak induction of
interferon expression by SARS-CoV-2 supports clinical trials of
interferon lambda to treat early COVID-19. Clin Infect Dis.
71:1410–1412. 2020.PubMed/NCBI
|
|
44
|
Benveniste EN and Qin H: Type I
interferons as anti-inflammatory mediators. Sci STKE.
2007:pe702007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Karimi Y, Giles EC, Vahedi F, Chew MV,
Nham T, Loukov D, Lee AJ, Bowdish DM and Ashkar AA: IFN-beta
signalling regulates RAW 264.7 macrophage activation, cytokine
production, and killing activity. Innate Immun. 26:172–182. 2020.
View Article : Google Scholar
|
|
46
|
Billiau A: Anti-inflammatory properties of
Type I interferons. Antiviral Res. 71:108–116. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
González-Navajas JM, Lee J, David M and
Raz E: Immunomodulatory functions of type I interferons. Nat Rev
Immunol. 12:125–135. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Guarda G, Braun M, Staehli F, Tardivel A,
Mattmann C, Förster I, Farlik M, Decker T, Du Pasquier RA, Romero P
and Tschopp J: Type I interferon inhibits interleukin-1 production
and inflammasome activation. Immunity. 34:213–223. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Arimori Y, Nakamura R, Yamada H, Shibata
K, Maeda N, Kase T and Yoshikai Y: Type I interferon limits
influenza virus-induced acute lung injury by regulation of
excessive inflammation in mice. Antiviral Res. 99:230–237. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Doherty PC, Turner SJ, Webby RG and Thomas
PG: Influenza and the challenge for immunology. Nat Immunol.
7:449–455. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
51
|
Maines TR, Szretter KJ, Perrone L, Belser
JA, Bright RA, Zeng H, Tumpey TM and Katz JM: Pathogenesis of
emerging avian influenza viruses in mammals and the host innate
immune response. Immunol Rev. 225:68–84. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Taubenberger JK and Morens DM: The
pathology of influenza virus infections. Annu Rev Pathol.
3:499–522. 2008. View Article : Google Scholar :
|
|
53
|
Li X, Geng M, Peng Y, Meng L and Lu S:
Molecular immune pathogenesis and diagnosis of COVID-19. J Pharm
Anal. 10:102–108. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Huang C, Wang Y, Li X, Ren L, Zhao J, Hu
Y, Zhang L, Fan G, Xu J, Gu X, et al: Clinical features of patients
infected with 2019 novel coronavirus in Wuhan, China. Lancet.
395:497–506. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Xu Z, Shi L, Wang Y, Zhang J, Huang L,
Zhang C, Liu S, Zhao P, Liu H, Zhu L, et al: Pathological findings
of COVID-19 associated with acute respiratory distress syndrome.
Lancet Respir Med. 8:420–422. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Gralinski LE and Menachery VD: Return of
the coronavirus: 2019-nCoV. Viruses. 12:1352020. View Article : Google Scholar :
|
|
57
|
Xu J, Zhao S, Teng T, Abdalla AE, Zhu W,
Xie L, Wang Y and Guo X: Systematic comparison of two
animal-to-human transmitted human coronaviruses: SARS-CoV-2 and
SARS-CoV. Viruses. 12:2442020. View Article : Google Scholar :
|
|
58
|
Sarzi-Puttini P, Giorgi V, Sirotti S,
Marotto D, Ardizzone S, Rizzardini G, Antinori S and Galli M:
COVID-19, cytokines and immunosuppression: What can we learn from
severe acute respiratory syndrome? Clin Exp Rheumatol. 38:337–342.
2020.PubMed/NCBI
|
|
59
|
Kindler E, Thiel V and Weber F:
Interaction of SARS and MERS coronaviruses with the antiviral
interferon response. Adv Virus Res. 96:219–243. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Perlman S and Dandekar AA:
Immunopathogenesis of coronavirus infections: Implications for
SARS. Nat Rev Immunol. 5:917–927. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Pedersen SF and Ho YC: SARS-CoV-2: A storm
is raging. J Clin Invest. 130:2202–2205. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chu H, Chan JF, Wang Y, Yuen TT, Chai Y,
Hou Y, Shuai H, Yang D, Hu B, Huang X, et al: Comparative
replication and immune activation profiles of SARS-CoV-2 and
SARS-CoV in human lungs: An ex vivo study with implications for the
pathogenesis of COVID-19. Clin Infect Dis. 71:1400–1409. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li SF, Gong MJ, Zhao FR, Shao JJ, Xie YL,
Zhang YG and Chang HY: Type I interferons: Distinct biological
activities and current applications for viral infection. Cell
Physiol Biochem. 51:2377–2396. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Haagmans BL, Kuiken T, Martina BE,
Fouchier RA, Rimmelzwaan GF, van Amerongen G, van Riel D, de Jong
T, Itamura S, Chan KH, et al: Pegylated interferon-alpha protects
type 1 pneumocytes against SARS coronavirus infection in macaques.
Nat Med. 10:290–293. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cinatl J, Morgenstern B, Bauer G, Chandra
P, Rabenau H and Doerr HW: Treatment of SARS with human
interferons. Lancet. 362:293–294. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Falzarano D, de Wit E, Rasmussen AL,
Feldmann F, Okumura A, Scott DP, Brining D, Bushmaker T, Martellaro
C, Baseler L, et al: Treatment with interferon-α2b and ribavirin
improves outcome in MERS-CoV-infected rhesus macaques. Nat Med.
19:1313–1317. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Ströher U, DiCaro A, Li Y, Strong JE, Aoki
F, Plummer F, Jones SM and Feldmann H: Severe acute respiratory
syndrome-related coronavirus is inhibited by interferon-alpha. J
Infect Dis. 189:1164–1167. 2004. View
Article : Google Scholar
|
|
68
|
Channappanavar R, Fehr AR, Zheng J,
Wohlford-Lenane C, Abrahante JE, Mack M, Sompallae R, McCray PB Jr,
Meyerholz DK and Perlman S: IFN-I response timing relative to virus
replication determines MERS coronavirus infection outcomes. J Clin
Invest. 130:3625–3639. 2019. View Article : Google Scholar
|
|
69
|
Hensley LE, Fritz LE, Jahrling PB, Karp
CL, Huggins JW and Geisbert TW: Interferon-beta 1a and SARS
coronavirus replication. Emerg Infect Dis. 10:317–319. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Turner RB, Felton A, Kosak K, Kelsey DK
and Meschievitz CK: Prevention of experimental coronavirus colds
with intranasal alpha-2b interferon. J Infect Dis. 154:443–447.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Falzarano D, de Wit E, Martellaro C,
Callison J, Munster VJ and Feldmann H: Inhibition of novel beta
coronavirus replication by a combination of interferon-alpha2b and
ribavirin. Sci Rep. 3:16862013. View Article : Google Scholar
|
|
72
|
Thiel V and Weber F: Interferon and
cytokine responses to SARS-coronavirus infection. Cytokine Growth
Factor Rev. 19:121–132. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Sainz B Jr, Mossel EC, Peters CJ and Garry
RF: Interferon-beta and interferon-gamma synergistically inhibit
the replication of severe acute respiratory syndrome-associated
coronavirus (SARS-CoV). Virology. 329:11–17. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Mossel EC, Sainz B Jr, Garry RF and Peters
CJ: Synergistic inhibition of SARS-coronavirus replication by type
I and type II IFN. Adv Exp Med Biol. 581:503–506. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Yoshikawa T, Hill TE, Yoshikawa N, Popov
VL, Galindo CL, Garner HR, Peters CJ and Tseng CT: Dynamic innate
immune responses of human bronchial epithelial cells to severe
acute respiratory syndrome-associated coronavirus infection. PLoS
One. 5:e87292010. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Larkin J, Jin L, Farmen M, Venable D,
Huang Y, Tan SL and Glass JI: Synergistic antiviral activity of
human interferon combinations in the hepatitis C virus replicon
system. J Interferon Cytokine Res. 23:247–257. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Li G and De Clercq E: Therapeutic options
for the 2019 novel coronavirus (2019-nCoV). Nat Rev Drug Discov.
19:149–150. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Fávaro WJ, Iantas SR, Gonçalves JM, Socca
EAR, Durán N and Billis A: Single-arm phase I/II study of the
safety and efficacy of OncoTherad immunomodulator in patients
BCG-refractory or relapsed non-muscle invasive bladder cancer. J
Clin Oncol. 37:e160002019. View Article : Google Scholar
|
|
79
|
Böckelmann PK, Tizziani SH, Durán N and
Fávaro WJ: New therapeutic perspective for bladder cancer in dogs:
Toxicological and clinical effects of oncotherad nanostructured
immunotherapy. J Phys Conf Ser. 1323:0120222019. View Article : Google Scholar
|
|
80
|
Fávaro WJ and Caballero NE: A method for
producing a nanostructured complex (cfi-1), a protein-associated
nanostructured complex (mrb-cfi-1) and use. US Patent 20200156951.
June 14–2018, issued May 21, 2020.
|
|
81
|
Fávaro WJ and Caballero NED: A method for
producing a nanostructured complex (cfi-1), a protein-associated
nanostructured complex (mrb-cfi-1) And use. Patent WO2018227261.
Filed June 13, 2018; issued December 19, 2018.
|
|
82
|
Alonso JCC, Reis IB, Gonçalves JM, Sasaki
BR, Cintra AA, Duran N, Billis A and Fávaro WJ: Oncotherad
immunotherapy elicits promising responses in Bacillus
Calmette-Guérin-unresponsive non-muscle invasive bladder cancer:
Results from phase I/II study. J Clin Oncol. 38:e170482020.
View Article : Google Scholar
|
|
83
|
Totura AL, Whitmore A, Agnihothram S,
Schäfer A, Katze MG, Heise MT and Baric RS: Toll-like receptor 3
signaling via TRIF contributes to a protective innate immune
response to severe acute respiratory syndrome coronavirus
infection. mBio. 6:e00638–00615. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Delafiori J, Alonso JCC, Santos LA,
Oliveira DN, Navarro LC, Brandt Busanello EN, Sales GM, Oliveira
AN, Rocha AR, Durán N, et al: A 78-year old urothelial cancer
patient with faster recovery from COVID-19: Potential benefit from
adjuvant active immunotherapy. SSRN. Jun 4–2020.Epub ahead of
print. View Article : Google Scholar
|
|
85
|
Chen J, Qi T, Liu L, Ling Y, Qian Z, Li T,
Li F, Xu Q, Zhang Y, Xu S, et al: Clinical progression of patients
with COVID-19 in Shanghai, China. J Infect. 80:e1–e6. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Wei Z, Burwinkel M, Palissa C, Ephraim E
and Schmidt MF: Antiviral activity of zinc salts against
transmissible gastroenteritis virus in vitro. Vet Microbiol.
160:468–472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Korant BD, Kauer JC and Butterworth BE:
Zinc ions inhibit replication of rhinoviruses. Nature. 248:588–590.
1974. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Suara RO and Crowe JE Jr: Effect of zinc
salts on respiratory syncytial virus replication. Antimicrob Agents
Chemother. 48:783–790. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
te Velthuis AJ, van den Worm SH, Sims AC,
Baric RS, Snijder EJ and van Hemert MJ: Zn(2+) inhibits coronavirus
and arterivirus RNA polymerase activity in vitro and zinc
ionophores block the replication of these viruses in cell culture.
PLoS Pathog. 6:e10011762010. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kaushik N, Anang S, Ganti KP and Surjit M:
Zinc: A potential antiviral against Hepatitis E virus infection?
DNA Cell Biol. 37:593–599. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Aydemir TB, Liuzzi JP, McClellan S and
Cousins RJ: Zinc transporter ZIP8 (SLC39A8) and zinc influence
IFN-gamma expression in activated human T cells. J Leukoc Biol.
86:337–348. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Bao S and Knoell DL: Zinc modulates
cytokine-induced lung epithelial cell barrier permeability. Am J
Physiol Lung Cell Mol Physiol. 291:L1132–L1141. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Reiber C, Brieger A, Engelhardt G, Hebel
S, Rink L and Haase H: Zinc chelation decreases IFN-β-induced STAT1
upregulation and iNOS expression in RAW 264.7 macrophages. J Trace
Elem Med Biol. 44:76–82. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Huang IC, Bailey CC, Weyer JL, Radoshitzky
SR, Becker MM, Chiang JJ, Brass AL, Ahmed AA, Chi X, Dong L, et al:
Distinct patterns of IFITM-mediated restriction of filoviruses,
SARS coronavirus, and influenza A virus. PLoS Pathog.
7:e10012582011. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Fu B, Wang L, Li S and Dorf ME: ZMPSTE24
defends against influenza and other pathogenic viruses. J Exp Med.
214:919–929. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Li S, Fu B, Wang L and Dorf ME: ZMPSTE24
is downstream effector of interferon-induced transmembrane
antiviral activity. DNA Cell Biol. 36:513–517. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Brieger A, Rink L and Haase H:
Differential regulation of TLR-dependent MyD88 and TRIF signaling
pathways by free zinc ions. J Immunol. 191:1808–1817. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Newton K and Dixit VM: Signaling in innate
immunity and inflammation. Cold Spring Harb Perspect Biol.
4:a0060492012. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Costarelli L, Giacconi R, Malavolta M,
Basso A, Piacenza F, DeMartiis M, Giannandrea E, Renieri C, Busco
F, Galeazzi R and Mocchegiani E: Effects of zinc-fortified drinking
skim milk (as functional food) on cytokine release and thymic
hormone activity in very old persons: A pilot study. Age (Dordr).
36:96562014. View Article : Google Scholar
|
|
100
|
Kahmann L, Uciechowski P, Warmuth S,
Plümäkers B, Gressner AM, Malavolta M, Mocchegiani E and Rink L:
Zinc supplementation in the elderly reduces spontaneous
inflammatory cytokine release and restores T cell functions.
Rejuvenation Res. 11:227–237. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
ILSI Brasil International Life Sciences
Institute do Brasil: Vitamina D. Funções Plenamente Reconhecidas de
Nutrientes. 2. Brazil: pp. 432018
|
|
102
|
Pludowski P, Holick MF, Pilz S, Wagner CL,
Hollis BW, Grant WB, Shoenfeld Y, Lerchbaum E, Llewellyn DJ,
Kienreich K and Soni M: Vitamin D effects on musculoskeletal
health, immunity, autoimmunity, cardiovascular disease, cancer,
fertility, pregnancy, dementia and mortality-a review of recent
evidence. Autoimmun Rev. 12:976–989. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Catarino AM, Claro C and Viana I: Vitamin
D-current perspectives. J Portug Soc Dermatol Venereol. 74:345–353.
2016. View Article : Google Scholar
|
|
104
|
White JH: Regulation of intracrine
production of 1,25-dihydroxyvitamin D and its role in innate immune
defense against infection. Arch Biochem Biophys. 523:58–63. 2012.
View Article : Google Scholar
|
|
105
|
Evans KN, Taylor H, Zehnder D, Kilby MD,
Bulmer JN, Shah F, Adams JS and Hewison M: Increased expression of
25-hydroxyvitamin D-1alpha-hydroxylase in dysgerminomas: A novel
form of humoral hypercalcemia of malignancy. Am J Pathol.
165:807–813. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Stoffels K, Overbergh L, Giulietti A,
Verlinden L, Bouillon R and Mathieu C: Immune regulation of
25-hydroxyvitamin-D3-1alpha-hydroxylase in human monocytes. J Bone
Miner Res. 21:37–47. 2006. View Article : Google Scholar
|
|
107
|
Hewison M, Freeman L, Hughes SV, Evans KN,
Bland R, Eliopoulos AG, Kilby MD, Moss PA and Chakraverty R:
Differential regulation of vitamin D receptor and its ligand in
human monocyte-derived dendritic cells. J Immunol. 170:5382–5390.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Fritsche J, Mondal K, Ehrnsperger A,
Andreesen R and Kreutz M: Regulation of 25-hydroxyvitamin D3-1
alpha-hydroxylase and production of 1 alpha,25-dihydroxyvitamin D3
by human dendritic cells. Blood. 102:3314–3316. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Enioutina EY, Bareyan D and Daynes RA:
TLR-induced local metabolism of vitamin D3 plays an important role
in the diversification of adaptive immune responses. J Immunol.
182:4296–4305. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Enioutina EY, Bareyan D and Daynes RA: TLR
ligands that stimulate the metabolism of vitamin D3 in activated
murine dendritic cells can function as effective mucosal adjuvants
to subcutaneously administered vaccines. Vaccine. 26:601–613. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Hansdottir S, Monick MM, Hinde SL, Lovan
N, Look DC and Hunninghake GW: Respiratory epithelial cells convert
inactive vitamin D to its active form: Potential effects on host
defense. J Immunol. 181:7090–7099. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Overbergh L, Stoffels K, Waer M, Verstuyf
A, Bouillon R and Mathieu C: Immune regulation of 25-hydroxyvitamin
D-1alpha-hydroxylase in human monocytic THP1 cells: Mechanisms of
interferon-gamma-mediated induction. J Clin Endocrinol Metab.
91:3566–3574. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Rhodes JM, Subramanian S, Laird E, Griffin
G and Kenny RA: Perspective: Vitamin D deficiency and COVID-19
severity-plausibly linked by latitude, ethnicity, impacts on
cytokines, ACE2 and thrombosis. J Intern Med. Jul 2–2020.Epub ahead
of print.
|
|
114
|
Yan T, Xiao R and Lin G:
Angiotensin-converting enzyme 2 in severe acute respiratory
syndrome coronavirus and SARS-CoV-2: A double-edged sword? FASEB J.
34:6017–6026. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Imai Y, Kuba K, Rao S, Huan Y, Guo F, Guan
B, Yang P, Sarao R, Wada T, Leong-Poi H, et al:
Angiotensin-converting enzyme 2 protects from severe acute lung
failure. Nature. 436:112–116. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Kuba K, Imai Y, Rao S, Gao H, Guo F, Guan
B, Huan Y, Yang P, Zhang Y, Deng W, et al: A crucial role of
angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced
lung injury. Nat Med. 11:875–879. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Kumar D, Gupta P and Banerjee D: Letter:
Does vitamin D have a potential role against COVID-19? Aliment
Pharmacol Ther. 52:409–411. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Musavi H, Abazari O, Barartabar Z,
Kalaki-Jouybari F, Hemmati-Dinarvand M, Esmaeili P and Mahjoub S:
The benefits of vitamin D in the COVID-19 pandemic: Biochemical and
immunological mechanisms. Arch Physiol Biochem:. Oct 8–2020.Epub
ahead of print. View Article : Google Scholar
|
|
119
|
Malek Mahdavi A: A brief review of
interplay between vitamin D and angiotensin-converting enzyme 2:
Implications for a potential treatment for COVID-19. Rev Med Virol.
30:e21192020. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Omori-Mizuno Y, Nakayama N, Inao M, Funyu
J, Asabe S, Tomita K, Nishikawa K, Hosoda Y, Tanaka M, Hashimoto Y,
et al: Randomized study comparing vitamin D3 and
1alpha-Hydroxyvitamin D3 in combination with pegylated
interferon/ribavirin therapy for chronic hepatitis C. J
Gastroenterol Hepatol. 30:1384–1390. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Abu-Mouch S, Fireman Z, Jarchovsky J,
Zeina AR and Assy N: Vitamin D supplementation improves sustained
virologic response in chronic hepatitis C (genotype 1)-naive
patients. World J Gastroenterol. 17:5184–5190. 2011. View Article : Google Scholar
|
|
122
|
Nimer A and Mouch A: Vitamin D improves
viral response in hepatitis C genotype 2-3 naive patients. World J
Gastroenterol. 18:800–805. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Yokoyama S, Takahashi S, Kawakami Y, Hayes
CN, Kohno H, Kohno H, Tsuji K, Aisaka Y, Kira S, Yamashina K, et
al: Effect of vitamin D supplementation on pegylated
interferon/ribavirin therapy for chronic hepatitis C genotype 1b: A
randomized controlled trial. J Viral Hepat. 21:348–356. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Petta S, Cammà C, Scazzone C, Tripodo C,
Di Marco V, Bono A, Cabibi D, Licata G, Porcasi R, Marchesini G and
Craxí A: Low vitamin D serum level is related to severe fibrosis
and low responsiveness to interferon-based therapy in genotype 1
chronic hepatitis C. Hepatology. 51:1158–1167. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Behera MK, Shukla SK, Dixit VK, Nath P,
Abhilash VB, Asati PK and Jain AK: Effect of vitamin D
supplementation on sustained virological response in genotype 1/4
chronic hepatitis C treatment-naive patients from India. Indian J
Med Res. 148:200–206. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Wang K, Hoshino Y, Dowdell K, Bosch-Marce
M, Myers TG, Sarmiento M, Pesnicak L, Krause PR and Cohen JI:
Glutamine supplementation suppresses herpes simplex virus
reactivation. J Clin Invest. 127:2626–2630. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Wang R, Dillon CP, Shi LZ, Milasta S,
Carter R, Finkelstein D, McCormick LL, Fitzgerald P, Chi H, Munger
J and Green DR: The transcription factor Myc controls metabolic
reprogramming upon T lymphocyte activation. Immunity. 35:871–882.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Nakaya M, Xiao Y, Zhou X, Chang JH, Chang
M, Cheng X, Blonska M, Lin X and Sun SC: Inflammatory T cell
responses rely on amino acid transporter ASCT2 facilitation of
glutamine uptake and mTORC1 kinase activation. Immunity.
40:692–705. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Sinclair LV, Rolf J, Emslie E, Shi YB,
Taylor PM and Cantrell DA: Control of amino-acid transport by
antigen receptors coordinates the metabolic reprogramming essential
for T cell differentiation. Nat Immunol. 14:500–508. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Chang WK, Yang KD and Shaio MF: Effect of
glutamine on Th1 and Th2 cytokine responses of human peripheral
blood mononuclear cells. Clin Immunol. 93:294–301. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Kim J, Zhang J, Cha Y, Kolitz S, Funt J,
Escalante Chong R, Barrett S, Kusko R, Zeskind B and Kaufman H:
Advanced bioinformatics rapidly identifies existing therapeutics
for patients with coronavirus disease-2019 (COVID-19). J Transl
Med. 18:2572020. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Oudemans-van Straaten HM, Bosman RJ,
Treskes M, van der Spoel HJ and Zandstra DF: Plasma glutamine
depletion and patient outcome in acute ICU admissions. Intensive
Care Med. 27:84–90. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
O'Mahony JB, Palder SB, Wood JJ, McIrvine
A, Rodrick ML, Demling RH and Mannick JA: Depression of cellular
immunity after multiple trauma in the absence of sepsis. J Trauma.
24:869–875. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
O'Sullivan ST, Lederer JA, Horgan AF, Chin
DH, Mannick JA and Rodrick ML: Major injury leads to predominance
of the T helper-2 lymphocyte phenotype and diminished
interleukin-12 production associated with decreased resistance to
infection. Ann Surg. 222:482–490; discussion 490-492. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Fläring UB, Rooyackers OE, Wernerman J and
Hammarqvist F: Glutamine attenuates post-traumatic glutathione
depletion in human muscle. Clin Sci (Lond). 104:275–282. 2003.
View Article : Google Scholar
|
|
136
|
Mittendorfer B, Gore DC, Herndon DN and
Wolfe RR: Accelerated glutamine synthesis in critically ill
patients cannot maintain normal intramuscular free glutamine
concentration. JPEN J Parenter Enteral Nutr. 23:243–252. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Houdijk AP, Rijnsburger ER, Jansen J,
Wesdorp RI, Weiss JK, McCamish MA, Teerlink T, Meuwissen SG,
Haarman HJ, Thijs LG and van Leeuwen PA: Randomised trial of
glutamine-enriched enteral nutrition on infectius morbidity in
patients with multiple trauma. Lancet. 352:772–776. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Boelens PG, Houdijk AP, Fonk JC, Puyana
JC, Haarman HJ, von Blomberg-van der Flier ME and van Leeuwen PA:
Glutamine-enriched enteral nutrition increases in vitro
interferon-gamma production but does not influence the in vivo
specific antibody response to KLH after severe trauma. A
prospective, double blind, randomized clinical study. Clin Nutr.
23:391–400. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Tao KM, Li XQ, Yang LQ, Yu WF, Lu ZJ, Sun
YM and Wu FX: Glutamine supplementation for critically ill adults.
Cochrane Database Syst Rev. 2014:CD0100502014.
|














