Introduction
Lung cancer is one of the leading causes of cancer
death. The overall 5-year survival rate for lung cancer is less
than 15% (1), principally due to
the metastasis, radiation resistance and drug-resistance in the
advanced stage. The main reason for drug treatment failure is the
disturbance of the regulation of cell death (2).
Apoptosis is the primary process of cell death that
is under complex molecular control and can be triggered by various
signals. It is well-documented that apoptosis deficiency in lung
cancer results in the drug-resistance (3). Therefore, it is priority for lung
cancer treatment to find an alternative process that induces lung
cancer death. Recently, many more scientists became interested in
autophagy, a cellular process called type II cell death. Central to
the process is the formation of autophagosomes, double-membrane
vesicles that can be observed by transmission electron microscope
(TEM) (4–6). Already in 1980, it was shown that the
autophagy capacity of lung cancer is lower than that of surrounding
normal cells (7). Furthermore,
nutrient deficiency and high cell density increase autophagy
capacity (8). Therefore, the
decrease of autophagy capacity may be in favor of cancer
survival.
The extract from Tripterygium wilfordii Hook
F, which has been shown to have strong immune inhibition and
anti-inflammation activities, is widely used in China to treat
certain autoimmune diseases (such as rheumatoid arthritis and
kidney diseases) (9).
Tripchlorolide (T4) can be extracted from Tripterygium wilfordii
Hook F, or synthesized from its precursor triptolide after
hydroxyl acylation and subsequently chlorination (10). After structure modification, T4 was
shown to have similar drug activities, but low toxicities compared
to triptolide (11). In the
present study, we examined the role of T4 in regulating
proliferation, apoptosis and autophagy in lung cancer A549.
Materials and methods
Materials
Materials used in this study include
3-Methylamphetamine (3-MA) and PI was purchased from Sigma,
anti-MAP-LC3-II from Santa Cruz, IgG-FITC from LingFei, goat
anti-rabbit secondary antibody from KPL, β-actin antibody from
NeoMarkers, chemiluminescent substrate from KPL, Alex488-conjugated
secondary antibody and Topro-3 from Invitrogen.
Cell culture
Lung cancer cell line A549 was obtained from Cell
Line Bank, Chinese Academy of Sciences, and maintained in F-12K
supplemented with 10% FBA and 100 μg/ml penicillin and streptomycin
at 37°C and 5% CO2. Every two days, cell growth media
was replaced with fresh media.
Drug treatment
A549 cells (around 80% confluence) were treated
with/without drugs as below: i) control, cells were not treated
with drugs; ii) T4 treatment, cells were exposed to T4 as the final
concentrations of 5 nM, 25 nM, 50 nM, 100 nM and 200 nM,
respectively; iii) 3-MA pretreatment plus T4 treatment, cells were
pretreated with 5 mM (final concentration) 3-MA for 1 h, and
followed by T4 treatment with various concentrations as indicated
above. Cells were collected for the following experiments after
they had been treated with/without drugs for 24 h, 48 h or 72 h.
Each experiment was performed in triplicates.
MTT assay
Cells (5×103/well) seeded in a 96-well
plate (3 wells per concentration) were exposed to a series of
concentrations of T4. The control was supplemented with the same
volume of media. Cells were cultured for 0–72 h. MTT (100 μl) (5
g/l) was added to all the wells and the plate was re-incubated at
37°C for 4 h. To each well 90 μl of 10% acid-isopropanol solution
was added. After 12 h, the plates were read using a test wavelength
of 550 nm with a reference wavelength of 630 nm. The mean OD values
of the experiments after calibration were used for calculating the
viability rate and the death rate: The viability rate = A550
sample group/A550 control group, the death rate = 1 -
the viability rate.
Immunofluorescence assay (IFA)
A549 cells were washed with 0.01 M PBS, fixed in 4%
paraformaldehyde solution for 30 min and washed again with PBS.
Then A549 were permeabilized by 0.3% Triton X-100 for 10 min and
blocked in TBS (5% BSA) for 60 min. After adding anti-MAP1-LC3-II
(diluted 1:100), the slides were incubated in a moist chamber at
4°C overnight and subsequently washed again with PBS. The slides
were further incubated with Alex 488-labeled secondary antibodies
for 1 h in a moist chamber at room temperature and washed again.
Topro-3 (diluted 1:100,000 in PBS) was added for nuclear staining
and incubated for 10 min, the slides were washed with PBS, and
anti-fade mounting medium was added, then covered with cover slip,
sealed with nail polish, and examined by fluorescence microscopy.
Micrographs were taken for result assessment. The experiment design
included negative control and blank control.
The examination of autophagosome by
TEM
Cells were fixed in 3% glutaradehyde/1.5%
paraformaldehyde for days or hours at 4°C, and postfixed with 1%
OsO4/1.5% potassiumferrocyanide for 1.5 h. After washing
with PBS, cells were stained with 70% ethanol saturated with uranyl
acetate, followed by gradient dehydration with ethanol-acetone, and
finally embedded in epoxy resin 618 for section. The ultrathin
sections (50 nm) were stained by uranyl acetate and lead citrate
for 5 min, respectively, and examined and photographed with
TEM.
Western blotting
Cells were lysed by lysate buffer (10 mmol/l Tris,
pH 7.4, 100 mmol/l NaCl, 1 mmol/l EDTA, 1 mmol/l EGTA, 1 mmol/l
NaF, 20 mmol/l Na4P2O7, 2 mmol/l
Na3VO4, 0.1% SDS, 0.5% sodium deoxycholate,
1% Triton-X 100, 1 mmol/l PMSF, 60 μg/ml aprotinin, 10 μg/ml
leupeptin, 1 μg/ml pepstatin) on ice for 30 min, centrifuged at
12,000 g for 30 min, and collected the supernatant. The supernatant
was diluted to 2 mg/ml (protein concentration), mixed 1:1 with
loading buffer (62.5 mM Tris-HCl, pH 6.8, 10% glycerol, 2% SDS, and
0.1% bromophenol blue), and boiled for 5 min. After cooling down,
samples (30 μg/lane) were loaded and run on a SDS-PAGE (10%) gel.
The separated proteins were transferred to PVDF membrane. Membrane
was blocked in TBS supplemented with 5% non-fat milk and 0.2%
Tween-20 for 1 h, and incubated with primary antibody (diluted
1:500) at 4°C overnight. Following incubation with secondary
antibody (coat anti-rabbit HRPO, diluted 1:4,000) for 2 h the
membrane was exposed to film. The images of bands were assessed by
software from Scion Co. β-actin was used as loading control.
Statistical analysis
Data are expressed as the mean ± SD (standard
deviation) of triplicates. The significance of the differences
among the treatments was assessed by one-way analysis of variance
(ANOVA). All analyses were conducted with SPSS statistical package.
P<0.05 was considered statistically significant.
Results
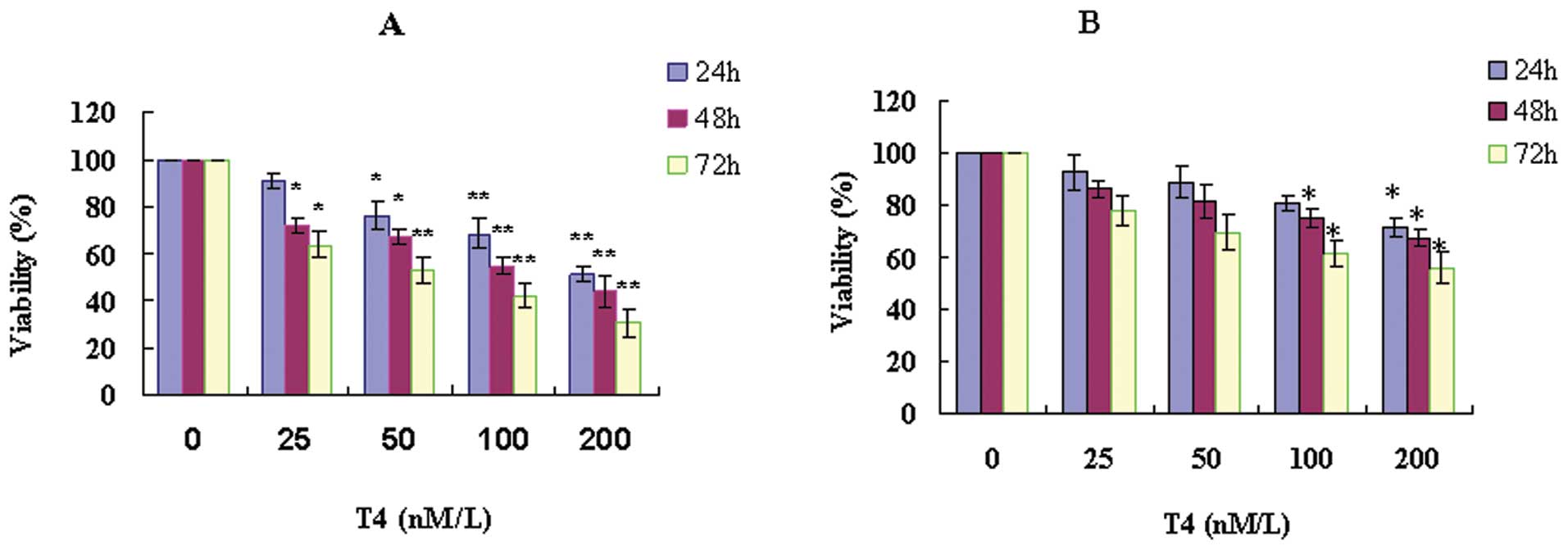
The toxicity of T4
A549 cells were treated with a series of
concentrations of T4 (0–200 nM) for different periods (24–72 h).
The viability of A549 was reduced with increasing T4 concentration
and longer incubation time (Fig.
1). After exposure to 200 nM T4 for 24 h, the cells viability
declined to around 50%. These results suggested that T4 suppressed
the proliferation of A549 in a dose- and time-dependent manner.
When the cells were pretreated with 3-MA, the inhibitor of
autophagy, T4 toxicity was dramatically reduced. Compared to the
controls, the 3-MA pretreated cells followed by T4 treatment were
not shown to have significance difference of the cell viability,
except those exposed to 100 nM T4 for 48–72 h or 200 nM T4 for
24–72 h.
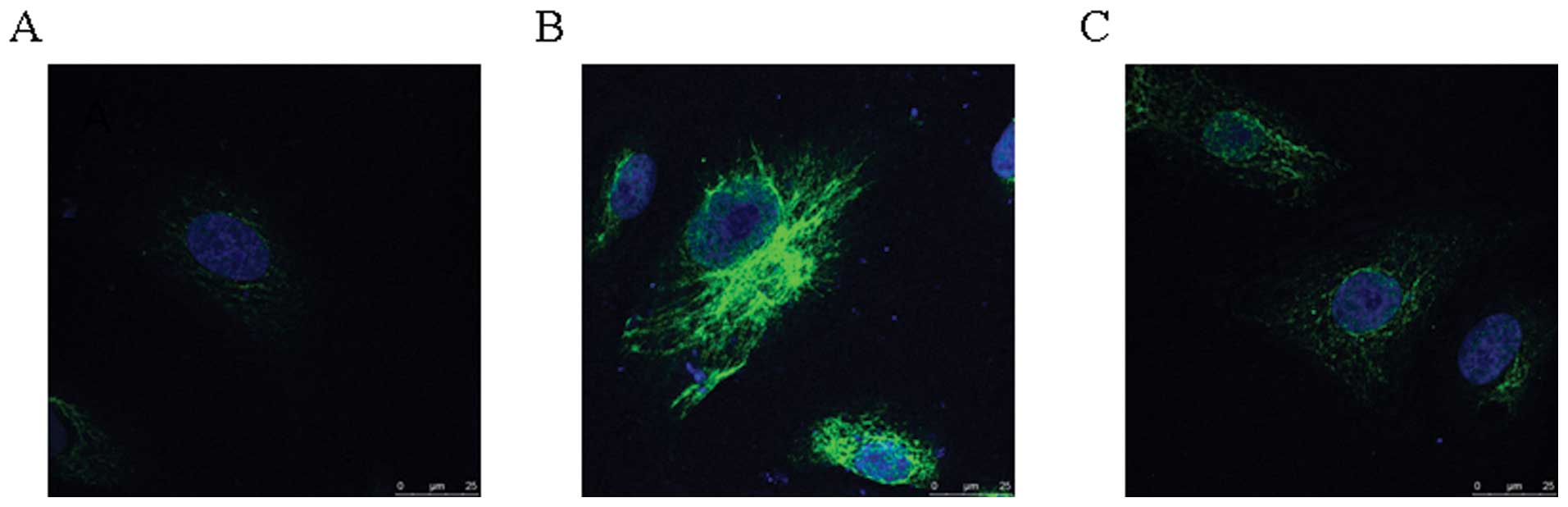
T4 rapidly induced A549 autophagy
Because the IC50 for T4 is 200 nM and
longer incubation periods dramatically increase the number of dead
cells, we chose 200 nM T4 for 24 h treatment as the
sample-conditions of IFA. In the control, the results of IFA showed
that LC3 (microtubule-associated protein 1 light chain 3) was
expressed at low level and distributed in the cytoplasm (Fig. 2). In cells treated with 200 nM T4
for 24 h, the protein expression level of LC3 was dramatically
upregulated, and intense granular staining of LC3 was observed.
However, in the cells pretreated with 3-MA, LC3 protein level was
similar to that in the control. It indicated that T4 can induce
A549 autophagy.
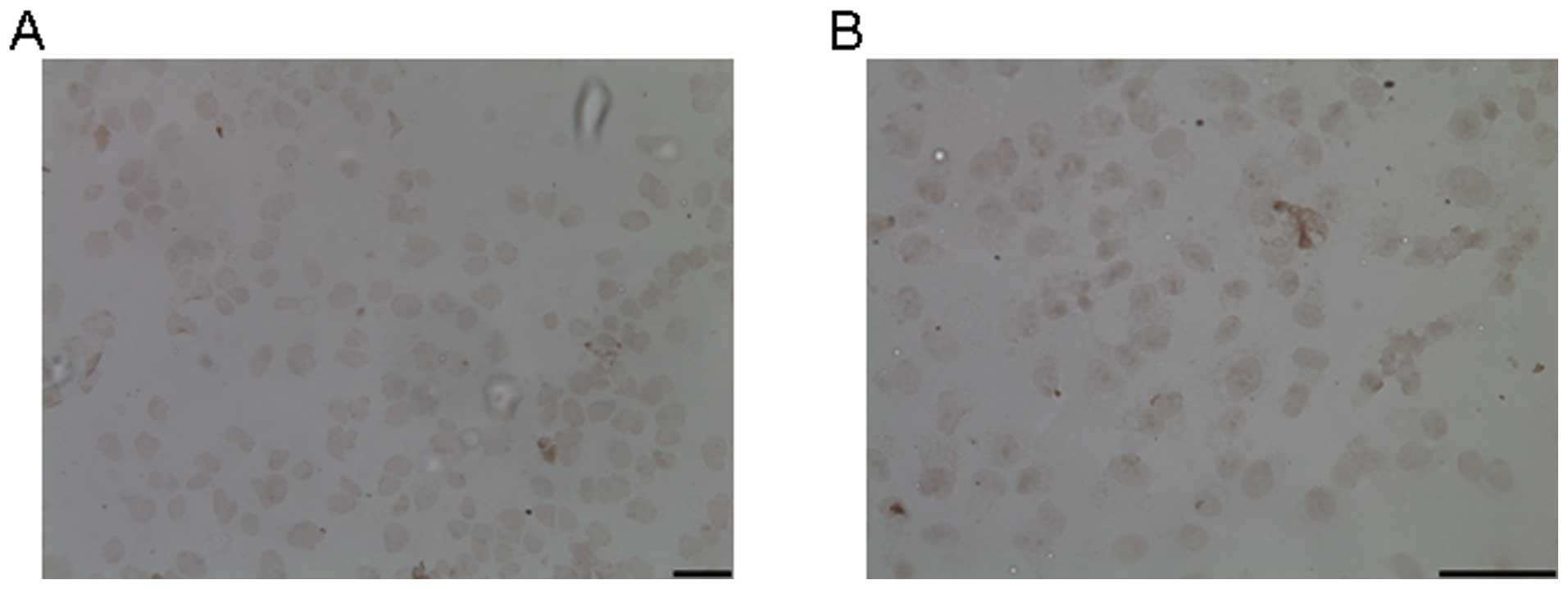
The effect of T4 on apoptosis
To determine whether T4 induced A549 apoptosis or
not, we examined the apoptotic bodies in the cells exposed to 200
nM T4 for 24 h. In the treated cells, no typical apoptotic bodies
were observed, as revealed by the TUNEL assay (Fig. 3). Therefore, A549 death induced by
T4 was not through apoptosis pathway.
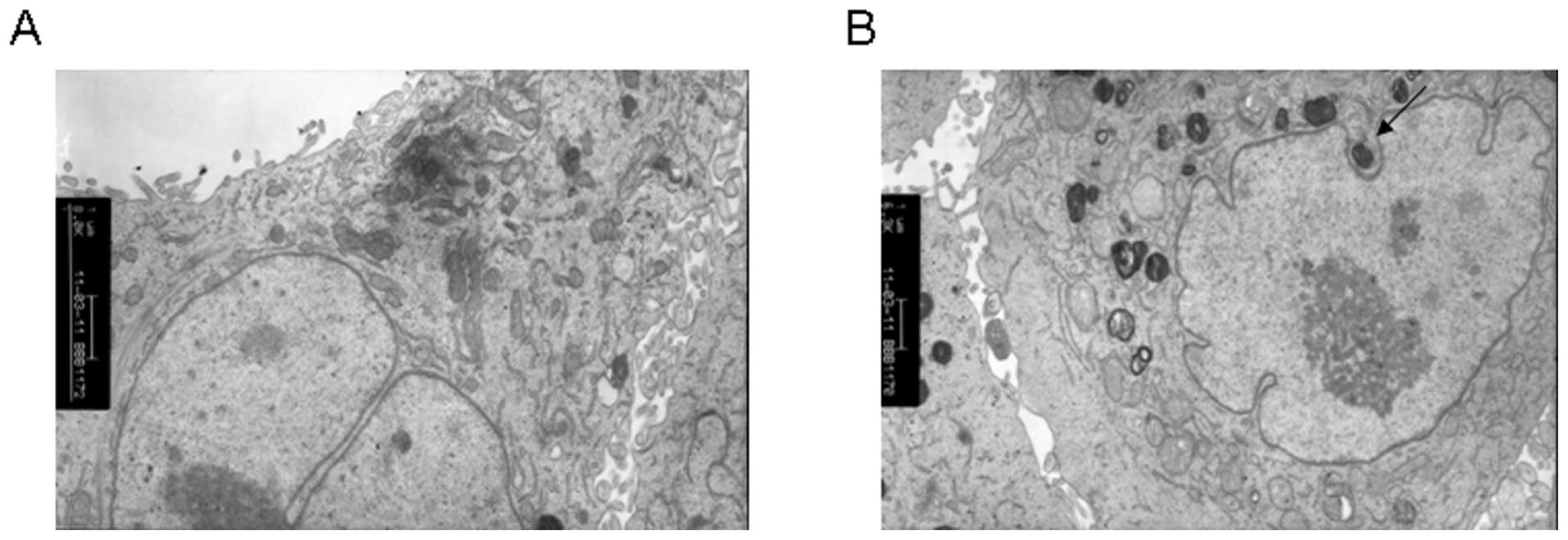
T4 induced autophagosome formation in
A549
To further confirm that T4 can induce A549
autophagy, we checked the formation of autophagosomes in T4 treated
cells using TEM. In the control, many normal vesicles containing
subcellular organelles were observed (Fig. 4). In the T4 treated cells, damaged
organelles were observed, such as swollen mitochondria surrrounded
by double-membrane vacuoles, which further formed autophagosomes.
Autophagosome subsequently fused with a lysozome and the internal
material was degraded. Undegraded debris within the autolysosomes
was also observed.
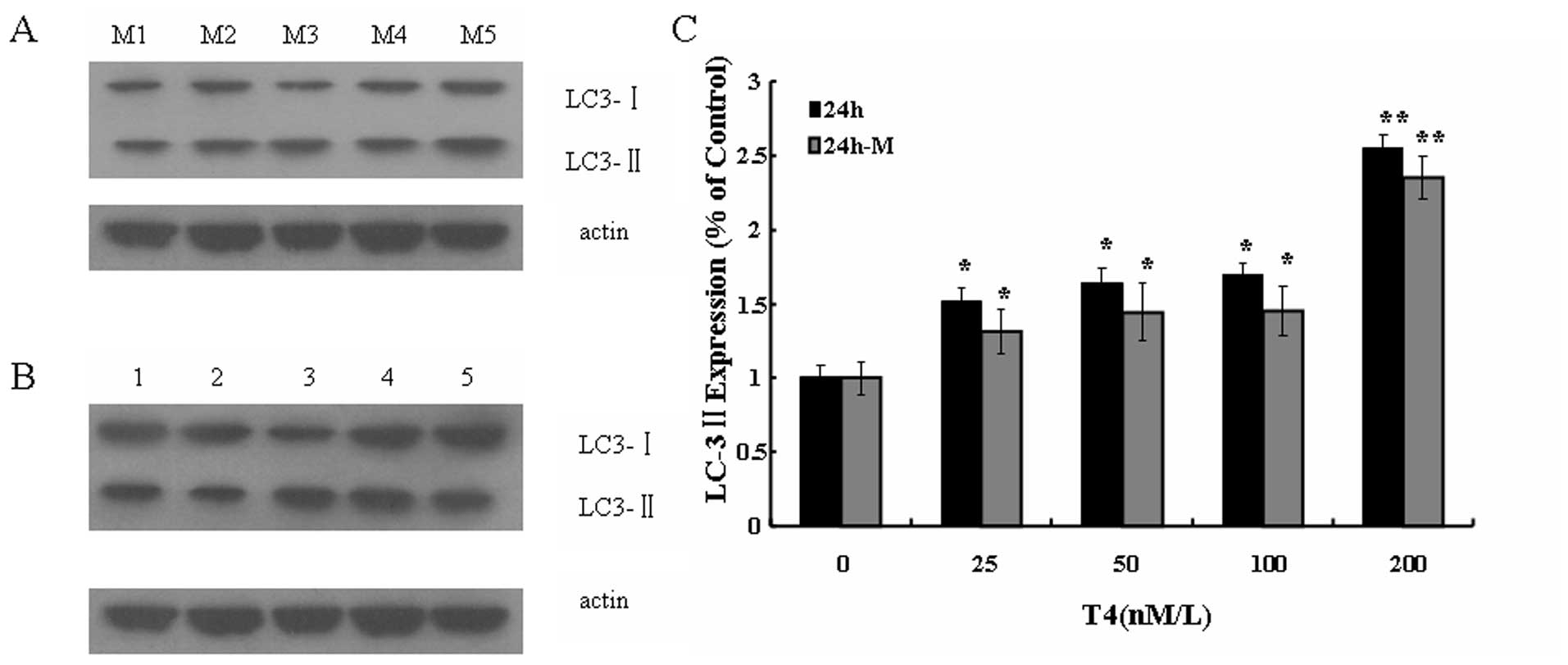
T4 stimulated LC3 expression in A549
cancer cell
LC3 is proposed to serve as a marker of
autophagosome membrane (16). As
determined by Western blotting, the protein level of LC3 II was
upregulated with the increase of T4 concentration, and reached the
peak at 200 nM T4 treatment (Fig.
5). However, in the 3-MA pretreated cells followed by T4
treatment, the level of LC3 II was much lower than that in the
cells only treated by T4, though it was slightly higher than that
in the control. These results suggested that T4 stimulated LC3 II
expression of A549, while 3-MA weakened the effect. Thus, it was
further confirmed that T4 induced cell death in A549 by
autophagy.
Discussion
Autophagy, the type II cell death, is different from
apoptosis (type I cell death). Autophagy is a strictly controlled
process in regulating cellular contents degradation and recycle,
and plays an essential role in organelles metabolism and bio-energy
supplementation. Malfunction of autophagy has been associated with
many human diseases, such as cancer and neurodegene-rative
disorders (12). Cancer is one of
the earliest diseases found to be related to autophagy (13). Autophagy inhibits tumorigenesis
mainly through regulation of the concentration of cellular
peroxide, diminishing the disturbance of protein metabolism and
maintenance of cell homeostasis. The decrease of autophagy activity
promotes oxidation stress and thereby increases accumulation of
cancer-related mutants (14).
Autophagic activity of cancer cells is lower than
that of normal cells (7). The
decrease of autophagic capacity is also found in precancerous cells
of some cancers, such as hepatocarcinoma induced by chemical
carcinogen. Reduced autophagic capacity is in favor of
carcinomagenesis. It is necessary for cells to maintain a normal
acutophgic capacity for cleaning the damaged organelles due to
chemical carcinogens, radioation and oxidation stress, and thus
maintaining genome integrity and reducing tumor occurrence.
Secondly, autophagy degrades organelles (such as endoplasmic
reticulum and Golgi) and long-lived protein, and thereof results in
negative protein balance in premalignant cells, inhibiting
uncontrolled proliferation (15).
In addition, hyperactive autophagy induces tumor cell death
(16–18).
Recently, it has been reported that some herbal
drugs were used for cancer treatment due to their effect on cell
autophagy. For exempt, Aloe-emodin, a herbal anthraquinone
derivative, induced rat C6 glioma autophagic death (19). Reseratrol, a natural phytoalexin
present in grapes, nuts and red wine, induced ovarian cancer cell
death through autophagy (20).
6-Shogaol, an active component from ginger, induced A549 autophagy
by suppressing the AKT/mTOR pathway (21). Triptolide, the precursor of T4,
inhibited the growth of hamster cholangiocarcinoma (22) and human tumors transplanted into
nude mice (23). Several studies
revealed that triptolide suppressed the growth of pancreatic cancer
and neuroblastoma both in vivo and in vitro (24,25).
Nammeeta et al found that triptolide induced pancreatic
cancer cell death through apoptosis and autophagy (26). Furthermore, triptolide in
combination with TRAIL significantly decreased pancreatic cancer
cell viability (27). However, the
role of triptolide in regulating lung cancer cells has not been
reported. The clinical application of the crude extract from
Tripterygium wilfordii is restricted due to its high
toxicity.
In the present study, we studied the role of T4 in
A549 lung cancer cells. MTT results showed that T4 induced cytoxity
in A549 cell death in a dose- and time-dependent manner.
Autophagosomes were observed by TEM in A549 treated by T4. When
A549 was pretreated with 3-MA and followed by T4 treatment, the
mortality of A549 was much lower than that in A549 only exposed to
T4. Although apoptosis adduced by triptolide was found in various
cancer cells (26,28–30),
almost no apoptosis was observed in A549 treated T4. Therefore, T4
suppressed the proliferation of A549 principally through activating
autophagy pathway, but not apoptosis pathway. A recent study showed
that triptolide promotes lung cancer apoptosis dependent on TRAIL
(31). Further studies are
required to investigate the effect of T4 in combination with TRAIL
on lung cancer cells.
Autophagosome formation is known to have a central
role in autophagy. Two ubiquitin-like conjugation systems are
essential for the formation. One is Atg12-Atg5 conjugation system,
and the other is Atg8 regulated lipidation system (15). LC3, the mammalian homologue of
Atg8, has three subtypes: pre-LC3, LC3-I and LC3-II. pre-LC3 is
processed by the cleavage of a portion (22 amino acids) of its
C-terminal to form LC3-I, which is distributed in the cytoplasm.
LC3-I is converted to LC3-II by further cleavage and conjugation to
phosphatidylethanolamine. LC3-II is recruited on the membrane of
pre-autophagosome by Atg5 and still located on autophagosome
membrane after autophagosome formation. Therefore, LC3-II is
proposed to serve as a marker of cell autophagy (15). IFA results showed that the number
of LC3-positive granules was significantly increased in A549
treated by T4. However, in the 3-MA pretreated cells, followed by
T4 treatment, autophagosomes were dramatically reduced compared to
the control. Moreover, the protein level of LC3-II was upregulated
after T4 treatment in a time- and concentration-dependent manner,
and reached the peak at 200 nM. 3-MA pretreatment dramatically
reduced the protein level of LC3-II in A549, though it was slightly
higher than that of the control. These results indicate that T4
kills lung cancer cells by promoting autophagy, which is different
from the mechanism of anti-cancer effect of triptolide for other
cancers (26,28–30).
In conclusion, our study confirmed that T4
suppressed the proliferation of A549 cells by activating autophagy.
The study provides new evidence for T4 clinical application in lung
cancer treatment.
Acknowledgements
We thank H.J. Ren for critical reading of the
manuscript. This study was supported by Education Department of
Fujian Province (JA10142).
References
|
1
|
Gilligan D, Nicolson M, Smith I, et al:
Preoperative chemotherapy in patients with resectable non-small
cell lung cancer: results of the MRC LU22/NVALT 2/EORTC 08012
multicentre randomised trial and update of systematic review.
Lancet. 369:1929–1937. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Melet A, Song K, Bucur O, Jagani Z,
Grassian AR and Khosravi-Far R: Apoptotic pathways in tumor
progression and therapy. Adv Exp Med Biol. 615:47–79. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gozuacik D and Kimchi A: Autophagy as a
cell death and tumor suppressor mechanism. Oncogene. 23:2891–2906.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Edinger AL and Thompson CB: Death by
design: apoptosis, necrosis and autophagy. Curr Opin Cell Biol.
16:663–669. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Boya P, Gonzalez-Polo RA, Casares N, et
al: Inhibition of macroautophagy triggers apoptosis. Mol Cell Biol.
25:1025–1040. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cuervo AM: Autophagy: in sickness and in
health. Trends Cell Biol. 14:70–77. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cuervo AM and Dice JF: A receptor for the
selective uptake and degradation of proteins by lysosomes. Science.
273:501–503. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ogier-Denis E and Codogno P: Autophagy: a
barrier or an adaptive response to cancer. Biochim Biophys Acta.
1603:113–128. 2003.PubMed/NCBI
|
|
9
|
Messina ME Jr and Halaby R: Does
triptolide induce lysosomal-mediated apoptosis in human breast
cancer cells? Med Hypotheses. 91932011.PubMed/NCBI
|
|
10
|
Yang GZ and Chen Y: The study development
of ester ethanol of common threewingnut root. J Chin Med Mater.
29:200–203. 2006.
|
|
11
|
Pan XD, Chen XC, Zhu YG, et al:
Tripchlorolide protects neuronal cells from microglia-mediated
beta-amyloid neurotoxicity through inhibiting NF-kappaB and JNK
signaling. Glia. 57:1227–1238. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Klionsky DJ and Emr SD: Autophagy as a
regulated pathway of cellular degradation. Science. 290:1717–1721.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Holm E, Hildebrandt W, Kinscherf R and
Droge W: Low post-absorptive net protein degradation in male cancer
patients: lack of sensitivity to regulatory amino acids? Oncol Rep.
17:695–700. 2007.PubMed/NCBI
|
|
15
|
Hait WN, Jin S and Yang JM: A matter of
life or death (or both): understanding autophagy in cancer. Clin
Cancer Res. 12:1961–1965. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qu X, Yu J, Bhagat G, et al: Promotion of
tumorigenesis by heterozygous disruption of the beclin 1 autophagy
gene. J Clin Invest. 112:1809–1820. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qian W, Liu J, Jin J, Ni W and Xu W:
Arsenic trioxide induces not only apoptosis but also autophagic
cell death in leukemia cell lines via up-regulation of Beclin-1.
Leuk Res. 31:329–339. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim KW, Mutter RW, Cao C, et al: Autophagy
for cancer therapy through inhibition of pro-apoptotic proteins and
mammalian target of rapamycin signaling. J Biol Chem.
281:36883–36890. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mijatovic S, Maksimovic-Ivanic D, Radovic
J, et al: Anti-glioma action of aloe emodin: the role of ERK
inhibition. Cell Mol Life Sci. 62:589–598. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Opipari AW Jr, Tan L, Boitano AE, Sorenson
DR, Aurora A and Liu JR: Resveratrol-induced autophagocytosis in
ovarian cancer cells. Cancer Res. 64:696–703. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hung JY, Hsu YL, Li CT, et al: 6-Shogaol,
an active constituent of dietary ginger, induces autophagy by
inhibiting the AKT/mTOR pathway in human non-small cell lung cancer
A549 cells. J Agric Food Chem. 57:9809–9816. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tengchaisri T, Chawengkirttikul R,
Rachaphaew N, Reutrakul V, Sangsuwan R and Sirisinha S: Antitumor
activity of triptolide against cholangiocarcinoma growth in vitro
and in hamsters. Cancer Lett. 133:169–175. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang S, Chen J, Guo Z, et al: Triptolide
inhibits the growth and metastasis of solid tumors. Mol Cancer
Ther. 2:65–72. 2003.PubMed/NCBI
|
|
24
|
Phillips PA, Dudeja V, McCarroll JA, et
al: Triptolide induces pancreatic cancer cell death via inhibition
of heat shock protein 70. Cancer Res. 67:9407–9416. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Antonoff MB, Chugh R, Borja-Cacho D, et
al: Triptolide therapy for neuroblastoma decreases cell viability
in vitro and inhibits tumor growth in vivo. Surgery. 146:282–290.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mujumdar N, Mackenzie TN, Dudeja V, et al:
Triptolide induces cell death in pancreatic cancer cells by
apoptotic and autophagic pathways. Gastroenterology. 139:598–608.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Borja-Cacho D, Yokoyama Y, Chugh RK, et
al: TRAIL and triptolide: an effective combination that induces
apoptosis in pancreatic cancer cells. J Gastrointest Surg.
14:252–260. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Meng HT, Zhu L, Ni WM, You LS, Jin J and
Qian WB: Triptolide inhibits the proliferation of cells from
lymphocytic leukemic cell lines in association with downregulation
of NF-kappaB activity and miR-16-1*. Acta Pharmacol Sin.
32:503–511. 2011.PubMed/NCBI
|
|
29
|
Li J, Zhu W, Leng T, et al:
Triptolide-induced cell cycle arrest and apoptosis in human renal
cell carcinoma cells. Oncol Rep. 25:979–987. 2011.PubMed/NCBI
|
|
30
|
Wu PP, Liu KC, Huang WW, et al: Triptolide
induces apoptosis in human adrenal cancer NCI-H295 cells through a
mitochondrial-dependent pathway. Oncol Rep. 25:551–557.
2011.PubMed/NCBI
|
|
31
|
Lee KY, Park JS, Jee YK and Rosen GD:
Triptolide sensitizes lung cancer cells to TNF-related
apoptosis-inducing ligand (TRAIL)-induced apoptosis by inhibition
of NF-kappaB activation. Exp Mol Med. 34:462–468. 2002. View Article : Google Scholar : PubMed/NCBI
|