Introduction
Breast and ovarian cancer are two of the most common
and lethal cancers affecting women worldwide. Estimates rank breast
cancer as being the number one cause of new cancer cases (1,383,000
cases, 22.9%) and cancer related deaths (458,000 deaths, 13.7%) in
women worldwide in 2008. Ovarian cancer was ranked as the seventh
cause of new cancer cases (225,000 cases, 3.7%) and cancer related
deaths (140,000 deaths, 4.2%) in women worldwide in the same year
(1). Among gynecological
malignancies, epithelial ovarian carcinoma (EOC) is the most lethal
to women in the world, accounting for approximately 90% of all
tumors affecting the ovaries (2).
Mortality from breast cancer often results from distant metastatic
spread to lung, bone and lymph node tissue while diagnosis of EOC
typically occurs in the late stages of disease after its spread
into the peritoneal cavity or other distant sites (2), thus making it difficult for effective
treatment to be administered. Although a number of risk factors for
the development of breast and ovarian cancer have been identified
the exact origin and pathogenesis of disease is still poorly
understood (3,4). Increasing our knowledge about the
fundamental biology of these diseases is needed for the development
of improved diagnostic, prognostic and therapeutic interventions at
all stages of disease.
Enzymatic members of the DP4-like gene family, DP4,
FAP, DP8 and DP9, share the rare ability to cleave N-terminal
dipeptides at post-proline bonds in the penultimate position and
play an important role in the biological processing of the
N-termini of peptides such as chemokines, glucagon-like peptide
(GLP)-1, GLP-2 and neuropeptide Y (NPY) (5–8).
Many of these substrates, particularly chemokines, are involved in
cancer remodeling and tumor progression (5–8).
Notably, the chemokine CXC12 [stromal derived factor 1 (SDF-1)], a
known endogenous DP4 substrate (9)
which is also cleaved by DP8 in vitro (7) and its receptor, CXCR4, form a
CXC12/CXCR4 axis that appears to play an important role in tumor
cell proliferation and metastases of numerous cancers including
breast and epithelial ovarian cancer (10–12).
DP4 proteolysis inactivates SDF-1α/β, disrupting the SDF-1/CXCR4
interaction affecting downstream signaling and the likely SDF-1
mediated cell migration and metastasis (13–15).
DP4, DP8 and DP9 have recently been shown to act as survival
factors for Ewing’s sarcoma family of tumors (ESFT) through the
inactivation of NPY-driven cell death within ESFT cells (16). There also appears to be
non-enzymatic functions of DP4, FAP, DP8 and DP9 that are important
to their roles affecting migration, proliferation and apoptosis of
tumor cells (17,18).
Aberrant expression of DP4 and FAP is observed in
many cancers, including breast and ovarian carcinoma, and their
level of expression appears to contribute to their conflicting
roles in these diseases (reviewed in ref. 19). In animal models of breast cancer,
the expression of DP4 at high levels appears to have a tumor
promoter effect (20–22) while overexpression of DP4 in EOC
appears to have a tumor suppressor effect (23,24).
When DP4 is highly expressed on the surface of rat lung capillary
endothelial cells it can function as an adhesion receptor for rat
breast cancer cells, aiding lung metastasis (20,21).
In addition, in Fischer 344/CRJ rats, which express low levels of
an enzymatically inactive DP4 mutant on the surface of rat lung
endothelium, there is a 33% reduction in lung metastasis following
inoculation with rat MTF7 breast cancer cells (22). In contrast, overexpression of DP4
in human EOC results in a decreased migratory and invasive
potential of the cells in vitro and decreased tumor load
with increased survival in vivo (23). Overexpression of DP4 in EOC has
also been associated with enhanced chemosensitivty to paclitaxel
(24), a chemotherapeutic commonly
used in combination with a platinum-based chemotherapy such as
cisplatin for the treatment of EOC and breast cancer. FAP, not
commonly present in normal adult tissues, is highly expressed in
over 90% of human epithelial tumors including breast and ovarian
tumors (25) and enzymatically
active FAP is present in human breast and ovarian carcinoma tissues
(26). In the tumor
microenvironment of transgenic mice, FAP-expressing stromal cells
have a major immunosuppressive effect that helps promote tumor
growth and survival (27).
However, aberrant expression of FAP is associated with differing
tumorigenic properties in breast cancer. Elevated expression of FAP
is observed in invasive ductal carcinoma cells of human breast
cancer tissue, cells from lymph node metastases (28) and has been associated with
pro-longed breast cancer-patient survival (29). Knockdown of FAP in human breast
carcinoma cell lines decreases tumorigenic properties of cells
(30) while overexpression of FAP
has been shown to increase tumor growth and angiogenesis (31). This research is contradictory
suggesting that FAP is functioning as a tumor promoter in breast
cancer while increasing patient survival. Therefore, the exact
levels of FAP expression may be important. Some in vivo
evidence demonstrates that the level of expressed FAP is associated
with its metastatic potential. Inoculation of immunosuppressed
Balb/cnu/nu mice with high, medium and low FAP
expressing SB247 ovarian cancer cell lines revealed a decrease in
metastasis in the low-expressing FAP cells (32). Thus in animal models of ovarian
carcinoma increased FAP expression results in tumor
progression.
DP8 and DP9 are ubiquitously expressed at the mRNA
level in all normal human adult and neonatal tissues (33–35).
However, information on the expression of DP8 and DP9 in
pathophysiological settings and potential roles in cancer is
limited. In contrast to the recently identified NPY-dependent
survival role suggested for DP8/DP9 in Ewings sarcoma (16), the overexpression of DP8 and DP9 in
HEK293T and HepG2 cells has been shown to increase migration,
proliferation and enhance staurosporine-streptomyces induced
apoptosis (17,36) suggesting the effects of DP8/DP9 in
cancer, like DP4 and FAP, may be tumor specific. In chronic
lymphocytic leukemia (CLL) a significant increase in DP8 mRNA has
been observed (37) while
upregulation of DP9 mRNA has been observed in testicular cancer
(38). In human meningiomas, DP8
and DP9 have a higher level of mRNA and protein expression,
relative to DP4 and FAP, and DP8/DP9 were identified as the enzymes
responsible for the majority of DP activity observed (39). In contrast, in human gliomas DP4
was found to be the predominant enzyme and not DP8 or DP9 (40). Altered expression levels of DP8 and
DP9, either alone or in relation to DP4 and FAP, are yet to be
investigated in other cancers including breast and ovarian
cancer.
Expression profiling of the DPs in breast and
ovarian cancer cell lines will help further our understanding of
the contribution of this family and their molecular mechanisms to
breast and ovarian tumorigenesis. DP2 and prolyl endopeptidase
(PEP) are also Clan SC serine proteases but are not members of the
DP4-like gene family. DP2 is an exopeptidase and PEP is an
endopeptidase. Both are capable of cleaving post-prolyl bonds and
as such many DP inhibitors can also inhibit their activity. For
this reason in this study we also investigated the expression of
DP2 and PEP to help delineate the roles of DP8, DP9, DP4 and FAP in
these cell lines. Here we provide the first study into the
expression and activity of DP8 and DP9, in context with DP4 and FAP
in breast and ovarian cancer cell lines.
Materials and methods
Cell culture
Cell lines used in this study are as follows: three
human breast epithelial cancer cell lines, MCF-7 and MDA-MB-231,
both originally derived from pleural effusion (PE) of breast
adenocarcinoma and MDA-MB-453 originally derived from PE of
metastatic breast cancer; three human EOC cell lines, OVCA-432,
OVCA-429 and SKOV3, originally derived from ascites of metastatic
EOC tissue; the human kidney epithelial cell line, 293T and the
human cervical epithelial cancer cell line, HeLa derived from
cervical adenocarcinoma. MCF-7, MDA-MB-231, MDA-MB-453, OVCA-432
and SKOV3 cell lines were maintained in RPMI-1640 supplemented with
10% FBS, 100 units/ml penicillin, and 100 μg/ml streptomycin
(Invitrogen). OVCA-429, 293T and HeLa cell lines were maintained in
DMEM high glucose (4,500 mg/l glucose) supplemented with 10% FBS,
100 units/ml penicillin, and 100 μg/ml streptomycin.
Characteristic features of these cell lines are outlined (Table I).
 | Table ICharacteristics of breast and ovarian
cancer cell lines used in study. |
Table I
Characteristics of breast and ovarian
cancer cell lines used in study.
| Cell type | Name |
Characteristics | References |
|---|
| Breast cancer | MCF-7 | Weakly invasive;
positive for ERcα expression; expresses insulin-like growth factor
binding protein (IGFBP) -2, -4 and -5; expresses WNTB7
oncogene | (ATCC# HTB-22)
(41, 42) |
| MDA-MB-231 | Highly invasive;
negative for ERcα expression; expresses EGFR, TGFα and the WNT7B
oncogene | (ATCC# HTB-26)
(42) |
| MDA-MB-453 | Weakly invasive;
negative for ERcα expression; negative for expression of EGFR;
expresses FGFR receptor | (ATCC# HTB-131)
(42, 43) |
| Ovarian cancer | OVCA-432 | Expresses lower
levels of HER2 receptor compared to SKOV3; positive for ERcα
expression; expresses EGFR | (44–46) |
| OVCA-429 | Expresses lower
levels of HER2 receptor compared to SKOV3; positive for ERcα
expression | (44, 46) |
| SKOV3 | Over-expresses
HER2; positive for ERcα expression but unresponsive to
stimulus; | (ATCC# HTB-77)
(45–47) |
| Other | 293T | Routinely/widely
used cell line; highly transfectable; expresses SV40 T antigen
(temperature sensitive gene) |
(ATCC#CRL-11268) |
| HeLa | Express EGFR; HPV
18 positive; widely considered to be negative for ERcα expression
but positive expression has been detected by some | (ATCC#CCL-2)
(48, 49) |
RNA isolation and real-time quantitative
PCR (qPCR)
Total RNA was isolated from 0.5–1×107
cells by a combination of the TRIzol® reagent
(Invitrogen) reagent method and the RNAqueous® kit
(Qiagen) with on-column DNase I (Invitrogen) digestion.
Complementary DNA (cDNA) was synthesized from 5 μg of total
RNA using 0.5 μg of Oligo(dT)15-primer (Roche
Diagnostics) and SuperScript® III Reverse Transcriptase
(Invitrogen) as per the manufacturer’s instructions. Quantitative
measurements of DPs and PEP mRNA levels were determined by
real-time qPCR using TaqManTM probe technology and
external PCR product standards of known copy number. Primer and
probe sequences used for DP8, DP9, DP4, FAP, DP2 and PEP are the
same as previously published (37). The hypoxanthine ribosyltransferase
1 (HPRT1) house-keeping primer sequences were as previously
published (50). Each PCR reaction
(12.5 μl) contained: 0.5 units Platinum® Taq
(Invitrogen, Carlsbad, CA, USA), 1X High Fidelity Platinum Taq
buffer (Invitrogen), 3 mM MgCl2 for DP8, DP9, DP4, FAP
and HPRT1 amplification or 2 mM MgCl2 for DP2 and PEP
amplification, 0.2 mM dNTPs (Fisher Scientific, WA, Australia) 50
ng each of forward and reverse primers, 200 nM probe and 15.625 ng
of cDNA or 107–102 copies of external PCR
standard. Quantitative real-time PCR reactions were run in a
Rotorgene 3000 (Corbette Research/Qiagen, CA, USA). Cycling
conditions were as follows: Step 1, 1 cycle of 94°C for 2 min; Step
2, 40 cycles of [94°C for 15 sec (denaturation); 60°C for 1 min
(annealing and extension)]; Step 3, 1 cycles of 72°C for 1 min.
Fluorescence emitted from the FAM dye upon its release from the
TaqManTM probes was acquired on the FAM/Sybr channel
(470 nm/510 nm filters) of the Rotorgene 3000 at the end of each
extension. Samples were tested in duplicate and repeated twice for
each cDNA sample synthesized. A standard curve ranging from
107–102 copies of each gene of interest and
the control HPRT1 gene was included in each run in
duplicate. The external PCR product standards of known number of
copies of purified PCR product/ml were prepared as previously
described (37). Initial
processing and analysis of the real-time qPCR data were performed
using Rotorgene 6 software (Corbette Research/Qiagen). All qPCR
data are represented as a ratio of the number of gene of interest
copies to the number of HPRT1 copies multiplied by 1000.
Protein extraction and western
blotting
Cell pellets (1–2×107 cells) were
resuspended in 800 μl of buffer [PBS (pH 7.2–7.4) containing
150 mM NaCl, 1 mM EDTA, and protease inhibitors 1 mM
phenylmethylsulfonyl fluoride (PMSF), 10 μM pepstatin A and
10 μM leupeptin] and kept on ice. Cells were then lysed by
sonication for 3×10 sec on ice. Lysates were separated into soluble
and insoluble protein fractions by centrifugation (18,000 g for 30
min at 4°C) and protein concentration determined by Bradford assay
(Bio-Rad) as per manufacturer’s instructions. Soluble proteins (50
μg) were analyzed by SDS-PAGE (8%) and western blotting as
previously described (37).
Membranes were probed with either rabbit polyclonal anti-DP8
(RP1-DP8; Triple Point Biologics Inc., OR, USA), rabbit polyclonal
anti-DP9 (RP1-DP9; Triple Point Biologics Inc.), or rabbit
polyclonal anti-β-actin (ab8227; Abcam), primary antibodies diluted
in blocking buffer (RP1-DP8, 1:1000; RP1-DP9, 1:2500; anti-β-actin,
1:2500). Immunoreactive proteins were visualised using SuperSignal
West Pico Chemiluminescent Substrate (ECL reagent) (Thermo Fisher
Scientific) and captured using a LAS-4000 imager (Fujifilm Life
Science, Tokyo, Japan). All cell lines were harvested on three
separate occasions for analysis by western blotting (n=3).
Quantification of bands was achieved by densitometry using
Multi-Gauge V3.0 software (Fujifilm Life Science).
Immunostaining and flow cytometry
Surface expression of endogenous DP4 and FAP protein
was detected by indirect immunofluorescence staining followed by
flow cytometry analysis. DP4 was detected using the anti-human
CD26, clone 2A6 (mouse IgG1) (14–0269; eBioscience) and FAP
detected using the F19 hybridoma supernatant as previously
described (37). Alexa
Fluor® 488 goat anti-mouse (Molecular
Probes®, Invitrogen) was used as the secondary antibody.
Intracellular protein expression was detected in cells fixed in 4%
(w/v) paraformaldehyde then permeabilized with 0.1% (v/v) Triton
X-100 in PBS. Labeled cells were analysed on a FACScan (BD Bio
Biosciences, San Jose, CA) and analysed using the WinMDI 2.8
(http://facs.scripps.edu) freeware software.
Subcellular fractionation
Cell pellets (>1×107) were resuspended
in 500 μl of homogenization buffer [50 mM Tris, 0.32 M
sucrose, 5 mM EDTA, 0.05% (w/v) NaN3] then incubated for
15 min on ice with brief vortexing every 5 min. Cells were then
homogenized by mechanical disruption with 1–2 strokes of the
Ultra-Turrax® T8 (IKA®-Werke Staufen,
Germany) on speed setting 1–2. The homogenate was centrifuged at
1,500 g for 10 min at 4°C (Eppendorf 5414C centrifuge; Eppendorf
AG, Hamburg, Germany) to remove large debris. Supernatant was
separated into membrane and cytosolic extracts by two steps of
ultra-centrifugation (100,000 g, 30 min, 4°C) in a TL-100
ultracentrifuge (Beckman, USA). After the first round of
centrifugation the supernatant was transferred to a clean
centrifuge tube and the remaining pellet (representing the membrane
protein fraction) was resuspended in 2 ml homogenization buffer for
washing. Following the second step of ultra-centrifugation the
cytosolic extract was transferred to a clean 1.5 ml tube and placed
on ice. Supernatant from the membrane fraction was discarded and
the remaining membrane fraction resuspended in 200 μl of 0.1
M sodium phosphate buffer (Na3PO4, pH 7.6)
and placed on ice. Samples were immediately used for enzyme assays
and determination of protein concentration by Bradford assay
(Bio-Rad) as per manufacturer’s instructions.
Enzyme assays
DP-like, DP2 and PEP enzymatic activity was assayed
colourimetrically using synthetic p-nitroanilide (pNA) containing
substrates (Bachem, Switzerland). Observed DP-like activity
attributable to DP8, DP9, DP4, FAP and DP2 enzymes was measured
using 0.5 mM H-Ala-Pro-pNA in 0.1 M Na3PO4 pH
7.6. For measurement of DP8, DP9, DP4 and FAP activity the
substrate 0.5 mM H-Gly-Pro-pNA in 0.1 M
Na3PO4 pH 7.6 was used; at pH 7.6 the
activity of DP2 towards this substrate is likely to be negligible
to none (63). DP2 activity was
detected using 0.5 mM H-Lys-Ala-pNA in 0.1 M
Na3PO4 pH 5.5. PEP activity was measured
using 0.5 mM Suc-Ala-Pro-pNA in 0.1 M Tris-HCl pH 8.0. Enzymatic
assays were performed using previously described methods (33). Total activity was calculated using
an extinction coefficient of 9.45 mM−1.cm−1
for pNA then corrected for protein concentration. Enzyme activity
is expressed as milli-Units (mU) per mg of protein, where, one unit
of activity is defined as the amount of enzyme that cleaves one
μmol of substrate per minute under the given assay
conditions.
Statistical analysis
Results are expressed as mean ± standard error of
the mean (SEM). For statistical analysis, the SPSS statistical
package (SPSS for Windows, v. 15.0, SPSS, Chicago, IL, USA) was
used. Differences among multiple groups were analysed using one-way
or two-way analysis of variance (ANOVA) followed by Bonferroni
correction. Differences among two separate groups were analysed
using Student’s t-test (assumes equal variance) or two sample
t-test assuming unequal variance. Where needed, data underwent
square-root transformation prior to statistical analysis. A value
of p<0.05 was considered significant. For correlation analysis
between two samples/factors the Pearson coefficient was used to
test for a linear relationship and, in the absence of linearity, a
Spearman’s correlation test was performed to determine if there was
a positive or negative correlation.
Results
DP8, DP9, DP4, FAP, DP2 and PEP mRNA are
differentially expressed in breast and ovarian cancer cell
lines
To help characterize the contribution of individual
DPs and PEP to the enzyme activities measured in this study we
compared the mRNA level expression of all DPs and PEP within each
cell line. DP8 mRNA expression was found to be significantly higher
(p<0.05) in all cell lines examined except the MDA-MB-453 cell
line (large variation in qPCR results) (Fig. 1). Within the breast cancer cell
lines, MCF-7 and MDA-MB-231, and ovarian cancer cell line,
OVCA-429, the expression of DP8 mRNA was significantly higher than
that of all other DPs and PEP (Fig.
1A, B and E). Also within MCF-7 cells the expression of DP2
mRNA was found to be significantly higher than that of FAP
(Fig. 1A) while in the MDA-MB-231
cell line the expression of both DP9 and PEP was found to be
significantly higher than that of DP4 (no mRNA detected) (p=0.022
and p=0.025, respectively) (Fig.
1B). In OVCA-429 cells the expression of DP9 was found to be
significantly higher than that of DP4 and FAP (p=0.042 and p=0.002,
respectively) (Fig. 1E). In the
OVCA-432 cell line DP8 mRNA expression was significantly higher
than that of DP9, DP4, DP2 (no mRNA detected) and PEP (p<0.005)
but not FAP (p=0.420) (Fig. 1D).
In the ovarian cancer cell line, SKOV3, DP8 mRNA was significantly
higher than DP9, FAP, DP2 and PEP (p<0.005) but not DP4
(p=1.000) (Fig. 1D). The
expression of DP4 mRNA in SKOV3 cells was significantly higher than
the levels of FAP and DP2 (no mRNA detected) (p=0.006 and p=0.01,
respectively) (Fig. 1D). In the
non-cancer derived 293T cells, DP8 mRNA expression was
significantly higher than that of DP4, FAP, DP2 and PEP
(p<0.005) but not DP9 (p=0.113) (Fig. 1G). HeLa cells are a commonly used
cervical cancer derived cell line that were included in this study
as a contrast to the breast and ovarian cancer cell lines. In HeLa
cells DP8 mRNA expression was significantly higher than that of
DP9, DP4, FAP, PEP and DP2 (p=1.000) (Fig. 1H). DP2 mRNA expression in HeLa
cells was significantly higher than that of DP4 (no mRNA detected)
(Fig. 1H). Both DP4 and DP2 mRNA
were completely absent in three cell lines; however, dual negative
expression of both was only observed in the breast cancer cell
line, MDA-MB-453 (Fig. 1C).
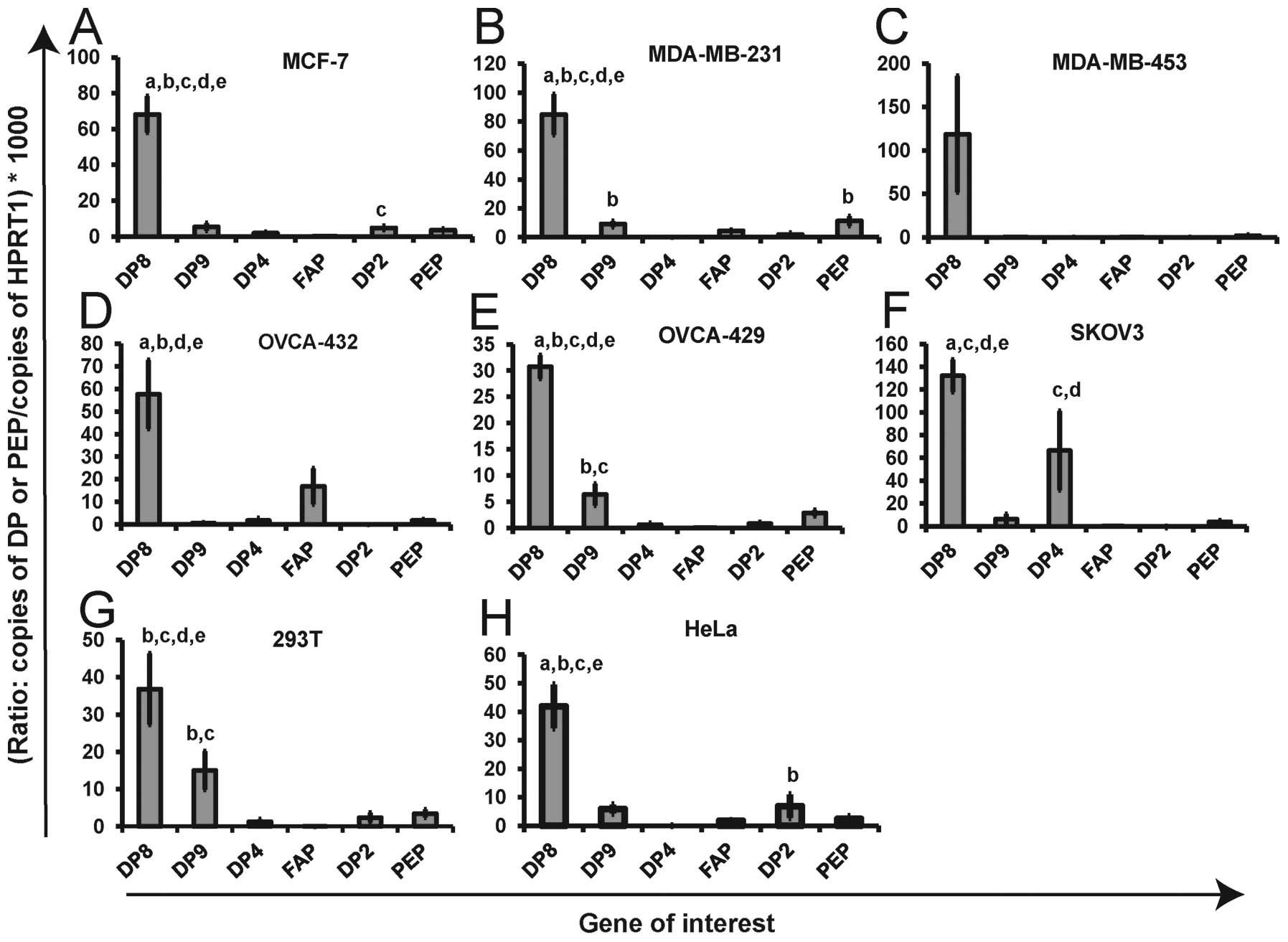 | Figure 1DP and PEP mRNA expression levels
within individual cell lines. DP8, DP9, DP4, FAP, DP2 and PEP mRNA
expression is displayed for three breast cancer [MCF-7 (A),
MDA-MB-231 (B), MDA-MB-453 (C)], three ovarian cancer, [OVCA-432
(D), OVCA-429 (E), SKOV3 (F)] and two alternate [293T (G) and HeLa
(H)] cell lines. Shown on each graph is the ratio of the copies of
each gene of interest/copies of HPRT1 (housekeeping gene)
multiplied by 1000 for ease of visual comparison. The expression of
each gene was determined in duplicate for two sets of cDNA
synthesized from each cell line (four replicates in total). Values
displayed as mean ± SEM (n=4). Statistically higher expression
(p<0.05) of a gene compared to other genes within a given cell
line is indicated by (a=DP9; b=DP4; c=FAP; d=DP2, e=PEP). DP8 was
expressed at significantly higher levels in all cell lines
(p<0.05). Statistical significance was determined by two-way
ANOVA using Bonferonni Correction. |
No correlation between mRNA level
expression of any DPs or PEP
Correlation testing was performed to examine whether
any relationship amongst the expression of DPs and PEP existed at
the mRNA level. A moderate positive non-linear correlation was
observed between DP9 and PEP mRNA expression however the finding
was non-significant (r=0.643, p=0.083, Spearman’s correlation
test). Apart from this, no linear relationship or non-linear
correlation was observed between the mRNA level expression of the
DPs and PEP (data not shown).
Ubiquitous DP8 and DP9 protein expression
across all cell lines
Endogenous DP8 and DP9 protein levels were assessed
in the breast and ovarian cancer cell lines. In agreement with mRNA
expression data, both DP8 and DP9 were found to be ubiquitously but
differentially expressed across all cell lines at the protein level
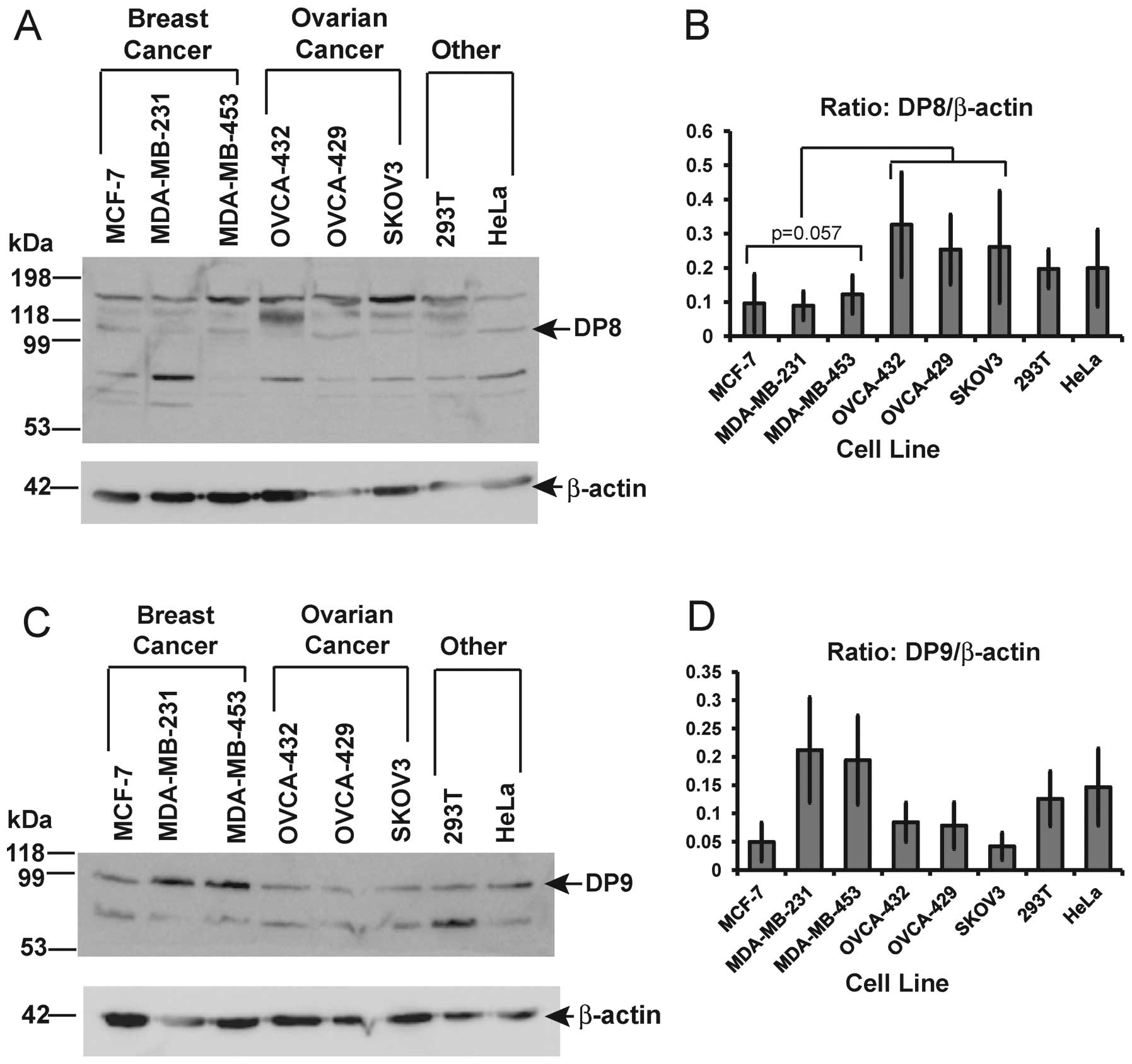
(Fig. 2A and C). Three prominent
DP8 immunoreactive bands of mobility around ∼100 kDa, 135 kDa and
180 kDa were detected with the anti-DP8 (RP1-DP8) antibody. Smaller
molecular mass bands were also detected between the 53 kDa and 99
kDa size markers (Fig. 2A). Two
prominent DP9 immunoreactive bands of monomeric molecular mass ∼100
kDa and a smaller molecular mass between the 53 kDa and 99 kDa size
markers were detected in all cell lines (Fig. 2C). Quantitative densitometry
analysis of the combined 100 kDa, 135 kDa and 180 kDa bands of DP8
(Fig. 2B), revealed a
non-significant decrease in DP8 protein levels in all three breast
cancer cell lines compared to the three ovarian cancer cell lines
(p= 0.057; two sample t-test). A slight decrease in expression was
also observed in all three breast cancer cell lines compared to the
293T and HeLa cells however this was also not significant.
Quantitative densitometry analysis of the ∼100 kDa monomeric band
of DP9 protein revealed no significant differences in DP9 protein
levels in MCF-7, OVCA-432, OVCA-429 and SKOV3 cell lines (Fig. 2D). The highest level of DP9 protein
was detected in the two estrogen receptor negative cell lines,
MDA-MB-231 and MDA-MB-453 (Fig.
2D). Interestingly, these were the two breast cancer cell lines
found to be negative for DP4 mRNA (Fig. 2B and C).
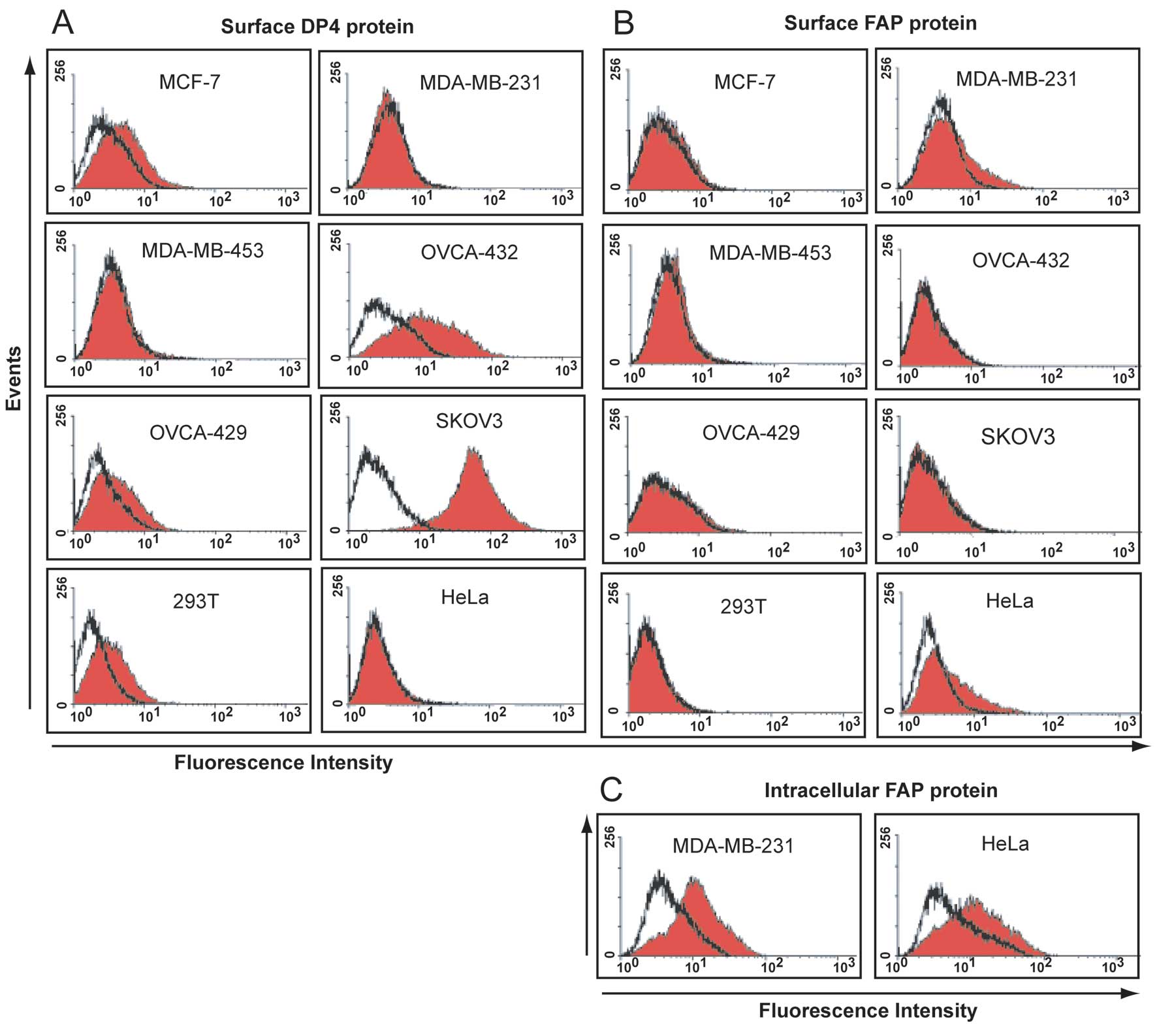
DP4 and FAP are differentially expressed
on the surface of cell lines
DP4 protein was detected on the surface of all three
ovarian cancer cell lines, the MCF-7 breast cancer cell line and
293T cells (Fig. 3A).
Corresponding with high levels of mRNA, the highest level of
surface DP4 protein was detected on SKOV3 and OVCA-432 cells
(Fig. 3A). No DP4 protein was
detected on the surface of the DP4 mRNA negative cell lines,
MDA-MB-231, MDA-MB-453 and HeLa (Fig.
3A). Surface staining revealed the expression of FAP as a weak
antigen on MDA-MB-231 and HeLa cells (Fig. 3B). Little to no FAP protein was
detected in the other cell lines (Fig.
3B) including OVCA-432 cells in which the highest levels of FAP
mRNA were detected.
Intracellular expression of FAP
protein
The weak surface antigen expression of FAP on
MDA-MB-231 and HeLa cell lines was indicative of the FAP protein
being expressed as an intracellular antigen. Intracellular
expression of FAP in MDA-MB-231 and HeLa cells was confirmed
following cell permeabilization (Fig.
3C).
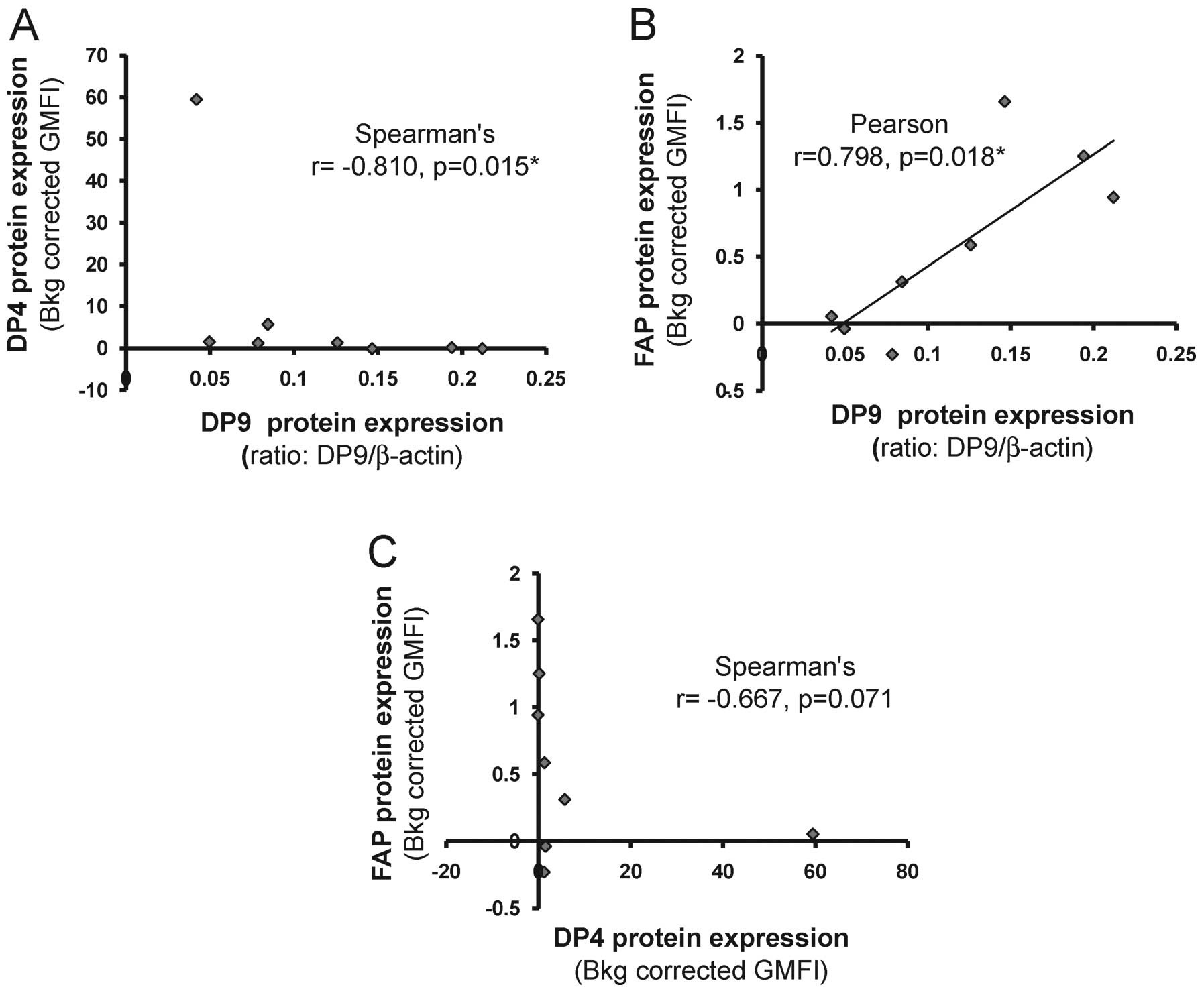
DP9 and DP4 protein levels correlate
To investigate whether there was any potential
relationships/co-regulation of DP8, DP9, DP4 and FAP at the protein
level, correlation analysis was performed. No correlation was
observed between DP8 protein and the DP9, DP4 or FAP proteins (data
not shown). However, a strong negative, non-linear, correlation was
observed between DP9 and DP4 protein level expression (r=−0.810,
p=0.015, Spearman’s correlation test; Fig. 4A). Conversely, a positive,
non-linear, correlation was observed between DP9 protein and
surface FAP protein (r=0.798, p=0.18, Spearman’s correlation test;
Fig. 4B). A moderate negative
correlation was observed between DP4 and FAP protein expression but
this finding was non-significant (r=0.667, p=0.071, Spearman’s
correlation test; Fig. 4C).
Correlation analysis of DP8, DP9, DP4 and
FAP protein expression with DPs and PEP mRNA expression
It may be possible that protein levels of the DPs
may affect their own gene expression via regulatory feedback loops
or, may affect the gene expression of other DPs and PEP. To address
this, the levels of DP8, DP9, DP4 and FAP protein expression were
examined across all cell lines in comparison with mRNA levels of
the DPs and PEP. Although not directly related, due to overlap in
enzymatic activity and specificity, DP2 and PEP were included in
this analysis. A strong positive linear correlation was observed
between the level of DP4 protein and DP4 mRNA (r=0.998, p=0.000,
Pearson coefficient; data not shown). DP4 was the only DP
displaying a correlation between the level of DP mRNA expression
and DP protein expression. No correlation was found between the
expression of DP8 protein and mRNA expression of any of the DPs or
PEP (data not shown). Interestingly, a strong negative correlation
was observed between the level of DP9 protein expression and DP4
mRNA expression (r=−0.903, p= 0.002, Spearman’s correlation test;
data not shown). However, no correlation was observed between DP4
protein and DP9 mRNA (data not shown), suggesting a non-reciprocal
relationship between DP4 expression and DP9 protein levels. A
moderate negative correlation was observed between the level of
expressed FAP protein and DP4 mRNA expression (r=−0.708, p=0.050,
Spearman’s correlation test; data not shown) but no correlation was
observed between the level of DP4 protein and FAP mRNA (data not
shown).
Enzyme activity of DPs, DP2 and PEP in
membrane and soluble fractions of breast and ovarian cancer
cells
To characterize the level of DP-like, DP2 and PEP
activity within each cell line statistical analysis was performed
to examine the difference in activity between fractions (membrane
and soluble) for each substrate within each cell line and the
difference in activity between substrates within each fraction
within each cell line (Fig.
5).
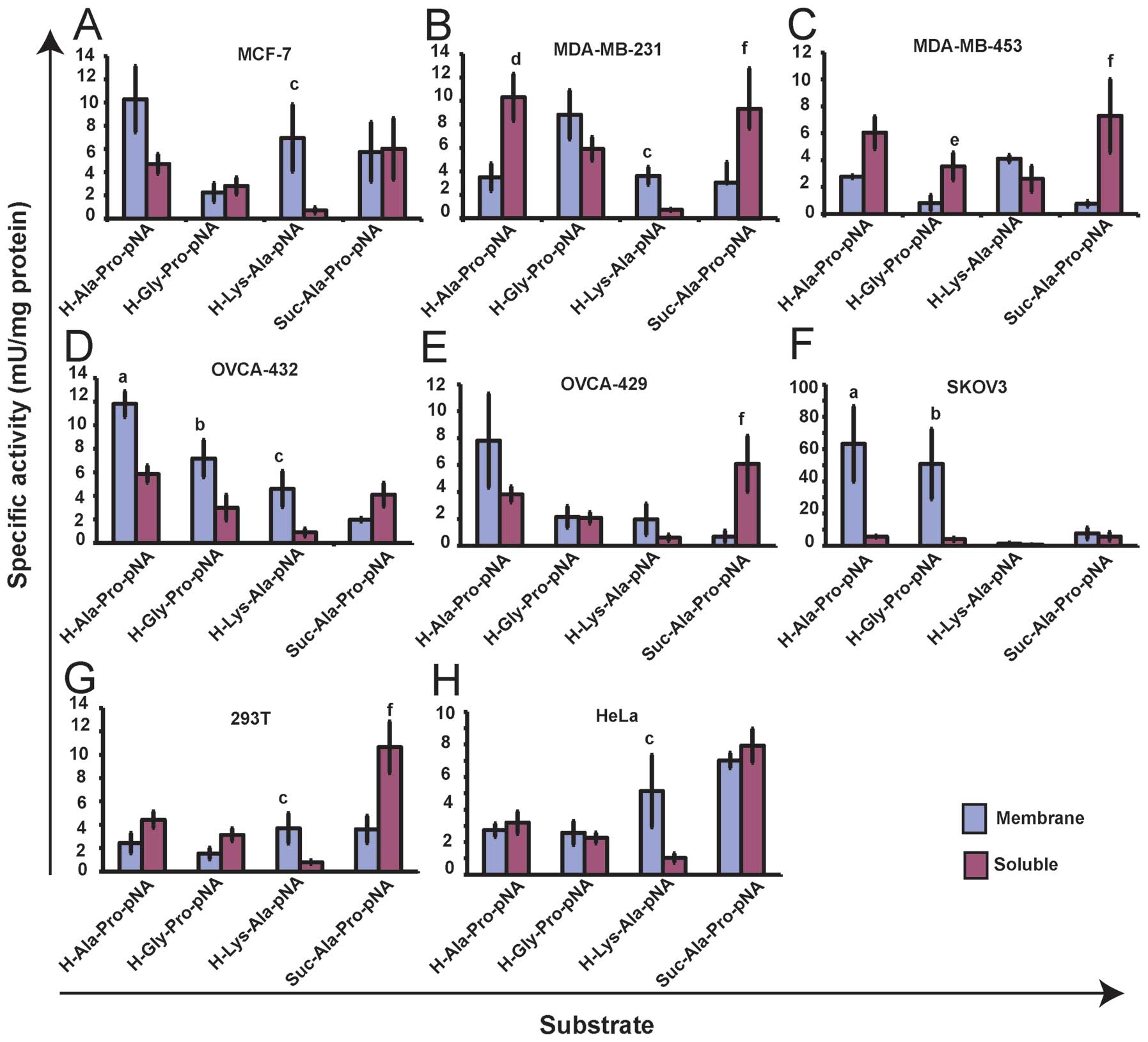 | Figure 5DP-like, DP2 and PEP enzyme activity
within each cell line. Enzyme activity against the DP-like
synthetic substrates H-Ala-Pro-pNA and H-Gly-Pro-pNA, the DP2
synthetic substrate, H-Lys-Ala-pNA and the PEP synthetic substrate,
Suc-Ala-Pro-pNA is displayed for three breast cancer cell lines,
MCF-7 (A), MDA-MB-231 (B), MDA-MB-453 (C), three ovarian cancer
cell lines, OVCA-432 (D), OVCA-429 (E), SKOV3 (F) and two alternate
cell lines, 293T (G) and HeLa (H). Values are expressed as the mean
± SEM (n=3). Statistical significance (p<0.05) was determined by
one-way and two-way ANOVA using Bonferonni Correction. Significance
was tested between fractions for each substrate within each cell
line and between substrates within each fraction within each cell
line. Significance is represented on the Figure as follows: (a)
indicates higher activity against H-Ala-Pro-pNA in membrane
fraction compared to soluble fraction; (b) indicates higher
activity against H-Gly-Pro-pNA in membrane fraction compared to
soluble fraction; (c) indicates higher activity against
H-Lys-Ala-pNA in membrane fraction compared to soluble fraction;
(d) indicates higher activity against H-Ala-Pro-pNA in soluble
fraction compared to membrane fraction; (e) indicates higher
activity against H-Gly-Pro-pNA in soluble fraction compared to
membrane fraction; (f) indicates higher activity against
Suc-Ala-Pro-pNA in soluble fraction compared to membrane
fraction. |
Comparison of activities between substrates within
each fraction revealed no significant difference between activities
in membrane fractions of MCF-7, MDA-MB-231 and MDA-MB-453 cells and
in soluble fractions of MCF-7 and MDA-MB-453 cells. In the soluble
fraction of MDA-MB-231 cells the level of DP-like and PEP
activities were significantly higher than the level of DP2 like
activity (p<0.01) (Fig. 5B). In
the DP4 positive ovarian cancer cell lines, OVCA-432 and SKOV3, the
level of DP-like activity against both H-Ala-Pro-pNA and
H-Gly-Pro-pNA, and the level of DP2 like activity against
H-Lys-Ala-pNA (OVCA-432 cells only), was significantly higher in
the membrane fraction compared to the soluble (p<0.05) (Fig. 5D and F). In OVCA-429 cells the
level of PEP like activity against Suc-Ala-Pro-pNA was
significantly higher in the soluble fraction compared to membrane
(p<0.01) (Fig. 5E). Comparison
of activities between substrates within fractions revealed that in
membrane fractions of OVCA-432 and SKOV3 cells the level of DP-like
activity was significantly higher than the level of DP2 and PEP
like activity (p<0.01) (Fig. 5D and
F) and, in OVCA-429 membrane fractions the level of DP-like
activity against H-Ala-Pro-pNA was significantly higher than the
level of PEP like activity (p=0.01) (Fig. 5E). In soluble fractions of OVCA-432
and OVCA-429 cells the level of PEP like activity and the level of
DP-like activity against H-Ala-Pro-pNA (OVCA-432 cells only), was
significantly higher than the level of DP2 like activity (Fig. 5D and E). No significant difference
was observed between substrates in the soluble fraction of SKOV3
cells.
In both 293T and HeLa cells the level of membrane
DP2 like activity against H-Lys-Ala-pNA was significantly higher
than soluble activity (p<0.05) and, in 293T cells the level of
soluble PEP like activity against Suc-Ala-Pro-pNA was significantly
higher than membrane activity (p=0.01) (Fig. 5G and F). In soluble fractions of
293T and HeLa cells the level of PEP like activity was
significantly higher than the level of DP-like activity against
H-Gly-Pro-pNA and H-Ala-Pro-pNA (293T cells only) and, in HeLa
cells only, the level of DP2 like activity (Fig. 5F and G). In addition the level of
soluble DP-like activity against H-Ala-Pro-pNA was significantly
higher than the level of soluble DP2 activity in 293T cells
(Fig. 5G). No significant
difference was observed between substrates in the membrane fraction
of 293T and HeLa cells.
Correlation analysis between DP8, DP9,
DP4 and FAP protein with observed DP-like, DP2 and PEP like enzyme
activities
To further characterize the enzymes responsible for
the observed activities statistical analysis was performed to test
for correlation between protein levels of DP8, DP9, DP4 and FAP and
the level of DP-like activity in membrane and soluble fractions of
each cell line. Correlation tests were also performed to determine
whether there was any possible relationship (e.g. co-regulation of
enzymes) between the level of DP8, DP9, DP4 and FAP protein and
observed level of activities against the DP2- and PEP-like
substrates, H-Lys-Ala-pNA and Suc-Ala-Pro-pNA. No correlation
between DP8 protein and soluble DP-like, DP2- and PEP-like
activities or between the level of DP9 protein and soluble DP-like
and PEP-like activity was observed (data not shown). A strong
positive linear relationship was observed between the level of
expressed DP4 protein and the level of membrane activity against
the DP-like substrates, H-Ala-Pro-pNA (r=0.993, p=0.000, Pearson
coefficient) and H-Gly-Pro-pNA (r=0.987, p=0.000, Pearson
coefficient; data not shown). A moderate negative, but not
significant, non-linear correlation was observed between the level
of FAP protein and membrane activity against H-Ala-Pro-pNA
(r=−0.643, p=0.086, Spearman’s correlation test; data not
shown).
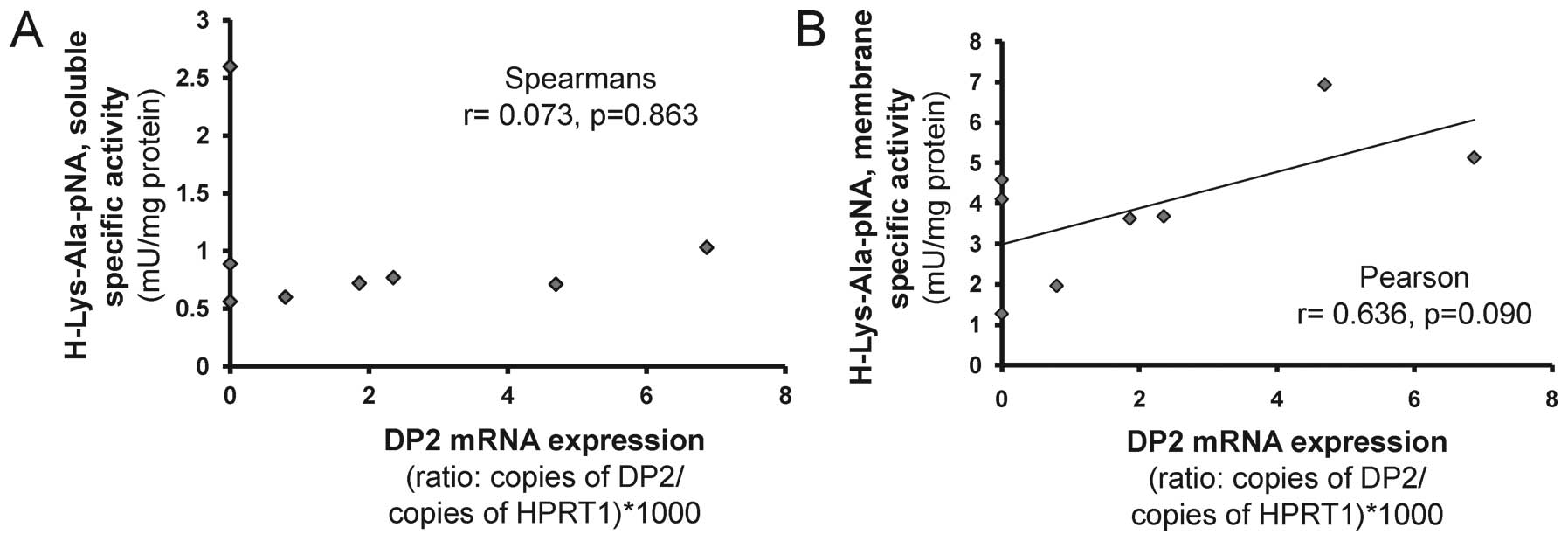
Correlation analysis between DP2 and PEP
mRNA with observed DP2- and PEP-like enzyme activities
Correlation tests were performed to test whether
there was any relationship between the level of DP2 and PEP mRNA
expression and the level of activities against the H-Lys-Ala-pNA
and Suc-Ala-Pro-pNA substrates. No correlation was observed between
the level of DP2 mRNA and the level of soluble activity against
H-Lys-Ala-pNA (Fig. 6A). In
contrast, a moderate positive, but non-significant, linear
relationship was observed between the level of DP2 mRNA and
membrane activity against the substrate H-Lys-Ala-pNA (r= 0.636,
p=0.90, Pearson coefficient; Fig.
6B).
Discussion
To date no one has investigated DP8 and DP9
expression in human breast and ovarian cancer or their expression
in context with that of DP4 and FAP in cancer. This study
demonstrates the ubiquitous but differential expression of DP8 and
DP9 protein in breast and ovarian cancer cell lines, which may
implicate each of these proteases in cancer pathogenesis. The lack
of correlation between transcript and protein levels for both DP8
and DP9 suggests that post-transcriptional regulation plays an
important role in regulating DP8 and DP9 protein levels. DP8
protein levels were lower in the breast cancer cell lines compared
to the ovarian cancer, 293T and HeLa cells, suggesting that a
breast cancer specific factor is contributing to its aberrant
regulation, leading to increased protein turnover or decreased
protein synthesis. For the first time we have identified a negative
non-reciprocal relationship between the expression of DP9 protein
and DP4 mRNA and protein expression, strongly suggesting that the
post-transcriptional regulation of DP9 is influenced/under the
control of a negative DP4 regulator. The relationship between DP9
and DP4 is an important avenue for further studies due to the
conflicting roles of DP4 in cancer and the potential involvement of
DP9 in these cancers.
Using DP4, DP2 and DP8/DP9 selective inhibitors,
DP8/DP9-like activity has been isolated in human leukocytes
(51) non-malignant human brain
tissue (40), human meningiomas
(39) and bovine and rat testes
(35). DP8 and DP9 mRNA expression
has been measured by qPCR in non-malignant human brain tissue
(40) and human meningiomas
(39). In human meningiomas, DP8
and DP9 protein have been detected by the use of specific DP8/DP9
antibodies for immunohistochemistry (39). To date no one has examined the
levels of DP8 and DP9 in breast and ovarian cancer cell lines or
patient samples. In patient samples of human meningiomas, a
non-glial brain tumor, DP8 and DP9 mRNA and proteins are more
abundantly expressed than DP4 and FAP (39). In these samples, the expression of
DP4 mRNA was found to positively correlate with the expression of
FAP mRNA (39). However,
quantitative analysis of DP8/DP9 protein levels in context with DP4
and FAP was not determined in these samples (39). Interestingly, we identified a
moderate negative, non-reciprocal correlation between the level of
expressed FAP protein and DP4 mRNA in breast and ovarian cancer
cell lines but found no correlation between FAP and DP4 mRNA. Both
DP4 and FAP are located adjacent to each other on the long arm of
chromosome 2 (52) and their
co-transcriptional regulation has been previously suggested
(53). The identification of a
negative correlation between FAP protein and DP4 mRNA and protein
may suggest that, similar to DP9, DP4-dependent
post-transcriptional regulation of FAP is occurring in breast and
ovarian cancer cell lines. A significant increase in DP8 mRNA has
been observed in human CLL compared to normal tonsil B-lymphocytes
however, this finding did not correlate with an increase in DP8
protein (37). In CLL, the
expression of DP9 mRNA was also higher in normal tonsil
B-lymphocytes compared to CLL (37). In tissues from testicular cancer
patients, a significant increase in DP9 mRNA was observed compared
to controls, however, the level of DP9 protein was not determined
in this study (38). All cell
lines used in our study were originally of epithelial origin
(Table I). Thus, the high level of
ubiquitously expressed DP8 mRNA suggests the importance of this
enzyme in epithelial cells. In each of these cell lines ubiquitous
but differential expression of DP8 and DP9 mRNA and protein was
detected but no correlation was observed between the levels of mRNA
and protein. This highlights the importance in following gene
expression measurements with protein-based investigations.
While low, but ubiquitous expression of FAP mRNA was
detected in all cell lines, its translated protein product was only
detected in MDA-MB-231 and HeLa cells. The identification of FAP as
an intracellular antigen suggests that FAP may be stored in
intracellular compartments of MDA-MB-231 and HeLa cells where it
remains until external stimuli results in translocation, and
presentation of FAP on the surface of these cells. Expression of
DP4 protein was strongly correlated, in a linear manner, with its
mRNA expression. In contrast, the level of ubiquitously expressed
DP8 and DP9 mRNA did not correlate with their level of ubiquitously
expressed protein, suggesting that post-transcriptional regulation
plays an important role in controlling translation of these
proteins in breast and ovarian cancer and/or that
post-translational turnover is high.
The size of immunoreactive DP8 and DP9 protein bands
detected by western blotting are consistent with previous studies
(33,36,54).
The detection of a smaller molecular mass immunoreactive band
between 53 kDa and 99 kDa for both DP8 and DP9 may be the result of
specific degradation products, suggesting high lability of DP8 and
DP9, or may be an alternative translation product/splice variant.
High lability has also been observed for recombinant DP8/DP9
proteins (6,55). Interestingly, for the first time a
strong negative non-reciprocal relationship was identified between
DP9 and DP4. It is likely that this relationship is cell line
and/or tissue dependent in the context of pathogenic
conditions.
The positive linear relationship observed between
expression of DP9 and FAP proteins is most likely due to the
post-transcriptional level effects of DP4 expression on both these
proteins and not an independent relationship between DP9 and FAP.
Increase in DP4 mRNA and protein decreases the level of DP9 protein
but not mRNA and correlates with the absence of FAP protein
expression but not mRNA. No reciprocal relationship was observed
between DP9 mRNA or FAP mRNA and DP4 protein suggesting that DP4 is
the key player in this negative relationship. It can be speculated
that despite the primarily cytoplasmic localization of DP9, in the
event of decreased or lost DP4 expression, DP9 protein levels may
increase and eventuate in its translocation to the plasma membrane
were it compensates for some of the loss of DP4 activity. Evidence
supporting the presence of DP9 and/or DP8 on the surface of DP4
negative lymphoid cells, including flow cytometry detection of DP9
protein on the surface of cells, has recently been presented
(56). Furthermore, it has been
suggested that DP9 may play a compensatory role in cell growth
regulation in DP4−/− mice (57).
Due to the absence of FAP protein on the surface of
SKOV3 cells and the complete absence of DP2, the significantly
higher level of DP-like activity in the membrane fraction of SKOV3
cells, compared to all other cell lines, can be attributed to DP4.
In this study we detected the highest level of expressed DP4
protein on the surface of SKOV3 cells. This is in contrast to other
studies were SKOV3 cells were found to have low DP4 expression and
were hence used for gain-of-function studies involving
overexpression of DP4 (23,58).
Previously, MDA-MB-231 cells were found to be FAP mRNA negative
(30) as determined by northern
blot analysis. Heterologous overexpression of FAP in MDA-MB-231 has
been shown to provide a gain-of-function leading to more rapid
tumor growth compared to controls in mouse models (31,59).
The difference in expression of DP4 and FAP in the present study
suggests that morphological changes have likely occurred due to
differences in routine cell culture and handling between
international research groups and the importance of re-phenotyping
cell lines before beginning these types of over and under
expression studies.
In MDA-MB-231 cells, the significant increase in
activity against H-Ala-Pro-pNA, but not H-Gly-Pro-pNA is likely due
to the intracellular expression of FAP which kinetically favours
the hydrolysis of H-Ala-Pro-substrates over H-Gly-Pro-substrates
(8,60). With the exception of MDA-MB-231
cells, the level of soluble DP-like activity was fairly constant
across all cell lines (no significant differences). Apart from the
contribution of intra-cellular FAP in MDA-MB-231 cells, this study
suggest that DP8 and/or DP9 are the major enzymes responsible for
the observed DP-like activity in soluble fractions of all cell
lines with the contribution of DP2 to these activities being
negligible. This is supported by the detection of little to no
enzyme activity against the DP2 substrate of H-Lys-Ala-pNA in
soluble fractions of cell lines and the complete absence of DP2 at
the mRNA level in MDA-MB-453, OVCA-432 and SKOV3 cells, all of
which were positive for DP8 and DP9 at the mRNA and protein
level.
Although DP2 is a soluble enzyme, it is primarily
localized to vesicular structures (61) making it highly probable that its
activity will be observed in membrane fractions of the cell lines
isolated in this study. Indeed, as displayed in Fig. 5, a higher level of DP2 activity was
observed in the membrane fractions of cell lines. In addition, a
moderately positive but non-significant correlation was detected
between DP2 mRNA and membrane H-Lys-Ala-pNA activity. However,
membrane activity against H-Lys-Ala-pNA was detected in all cell
lines, including in the three cell lines negative for DP2 mRNA,
MDA-MB-453, OVCA-432 and SKOV3 cells. Therefore, we cannot exclude
the likelihood of another enzyme being responsible for some of the
observed DP2-like activity in the cell lines that are DP2 negative
or positive at the mRNA level. Likewise, in soluble fractions the
level of activity against H-Lys-Ala-pNA was fairly constant and may
be due to the expression of some DP2 protein, in the DP2 positive
cell lines, and another enzyme. Of most interest was the
identification of a significantly positive correlation between the
level of DP9 protein and soluble activity against H-Lys-Ala-pNA.
Purified recombinant DP9 retains a low level of residual activity
against H-Ala-Pro-pNA at acidic pH 5.5 (62) thus we can speculate that DP9 may be
responsible for some of the soluble activity against H-Lys-Ala-pNA
observed in the present study. Although DP4 is capable of cleaving
H-Lys-Ala-pNA, its activity at acidic pH 5.5 is negligible
(63) thus it is not likely to be
the enzyme responsible for the observed DP2-like activity. In
addition, significantly higher levels of activity were detected in
the membrane fraction compared to soluble fraction of the DP4 and
DP2 negative MDA-MB-453 cells. These findings suggest that an
alternate membrane-associated enzyme, or enzymes, is responsible
for hydrolysis of H-Lys-Ala-pNA at an acidic pH. This is further
supported by the detection of residual activity against
H-Lys-Ala-pNA in intact cells following treatment with selective
DP2 inhibitors (64,65). As a number of aminopeptidases are
capable of cleaving N-terminal Lys and Ala residues (66–69)
it is reasonable to suggest that one or more aminopeptidases may be
responsible for the cleavage of Lys, then Ala, from the synthetic
substrate and thus release of pNA for colorimetric detection in the
DP2 enzyme assay.
The higher level of endopeptidase activity against
Suc-Ala-Pro-pNA in soluble fractions compared to membrane fractions
is consistent with the cytosolic localization of PEP. Forms of
membrane bound PEP have been identified (70) and may be responsible for some of
the detected membrane endopeptidase activity against
Suc-Ala-Pro-pNA. Unique to the DP4-like gene family, FAP also has
endopeptidase activity (60),
displaying a strong specificity for
NH2-Gly-Pro-substrates (71), poorly cleaving synthetic
Suc-Ala-Pro containing substrates (8). Thus, its contribution to the activity
against Suc-Ala-Pro-pNA in FAP positive MDA-MB-231 and HeLa is
likely to be minimal, if any.
Recent data suggest that high levels of endogenous
DP4, DP8 and DP9 can provide a survival mechanism for ESFT,
protecting ESFT cells from endogenous NPY-induced cell death
(16) while heterologous
overexpression of DP8 and DP9 has been shown to enhance
staurosporine-induced apoptosis and spontaneous apoptosis in the
case of DP9 (17,36). These differences highlight the
importance of investigating endogenous expression of DPs in the
context of differing pathological settings. Furthermore, it
suggests that the roles of DP8 and DP9 may be tumor specific. As we
have found in the present study, the expression of DP4 in relation
to DP9 is likely to play an important role in breast and ovarian
cancer pathogenesis. Lu et al investigated the roles of DPs
in ESFT, individual siRNA knockdown of DP4, DP8, DP9 and FAP was
performed in human ESFT cell lines, however no increase in DP9 mRNA
levels was detected following siRNA decrease in DP4 mRNA (16). Unfortunately, the Lu et al
study did not measure protein level expression. In all cell lines
analyzed in our study, we observed no correlation between the
endogenous expression of DP8 and DP9 mRNA and protein. This is
consistent with the Yao et al observation of no change in
DP8 protein expression following heterologous overexpression of DP9
in HEK293T and HepG2 cells (36).
Yao et al showed that DP9 overexpression
inactivated phosphoinositide 3-kinase (PI3K)/Akt signaling in an
epidermal growth factor (EGF)-dependent manner, attenuating cell
proliferation and enhancing apoptosis (36). Akt is a key activator of cell
growth and survival mechanisms (72). Dysregulation of the PI3K/Akt
pathway resulting in enhanced Akt signaling is a common feature of
many cancers, including breast and ovarian cancer (73,74),
that helps drive tumorigenesis (72). Thus drugs inhibiting the PI3K/Akt
molecular pathway are attractive for the therapeutic treatment of
breast and ovarian cancer (72,73,75).
Aberrant expression of the EGF receptor (EGFR) is also observed on
the surface of numerous cancers; thus, a complex interplay between
levels of EGFR, DP9, and members of the PI3K/Akt pathway are likely
to contribute to context specific roles of DP9 in cancer.
Inhibition of the Akt pathway may be a potential mechanism by which
overexpression of DP9 enhances apoptosis in HepG2 and HEK293T cell
lines and implicates a potential therapeutic role for DP9 in breast
and ovarian carcinoma.
Breast cancer is a largely hormone-dependent cancer
with the status of steroid hormones, including estrogen,
progesterone and androgen, and the expression of human epidermal
growth factor receptor 2 (HER2) playing a prominent role in the
development and progression of tumors (76–79).
MCF-7 cells used in this study are one of the most widely utilized
cell models for estrogen receptor (ERc) positive breast cancer
(60). In contrast, the other two
breast cancer cell lines, MDA-MB-231 and MDA-MB-453, are ERc
negative. MDA-MB-231 are known to be highly invasive and are
positive for EGFR expression while MDA-MB-453 cells are weakly
invasive, and lack EGFR expression (42). All three ovarian cancer cell lines,
OVCA-429, OVCA-432 and SKOV3 are ERc+ (44); however, despite having a functional
estrogen receptor SKOV3 cells are unresponsive to estrogen or
anti-estrogen treatment (47).
SKOV3 cells express high levels (over-express) HER2 while both
OVCA-432 and OVCA-429 cells express lower levels of HER2 in
relation to SKOV3 cells (46).
Interestingly, the absence of DP4 expression in breast cancer cell
lines, MDA-MB-231, MDA-MB-453 and HeLa cells coincides with the
reported ERc negative status of these lines. An increase in DP4
expression upon increasing ERc positive nature of ovarian cancer
patient samples has been previously reported; however, no
correlation was found between DP4 protein and steroid hormone
receptor expression or severity of disease in patients (80). The findings in our current study
suggest that this should be revisited and investigated in the
context of DP9 and DP4.
In summary, this study demonstrates for the first
time that DP8 and DP9 are expressed in breast and ovarian carcinoma
cell lines and suggests future investigations are required into
understanding the complex interplay between the levels of DP9, DP4
and steroid hormone receptor status in breast and ovarian cancer.
In addition, this study highlights the need for elucidating the
mechanisms controlling transcription and post-transcriptional
regulation of DP8, DP9, DP4 and FAP. Confirmation of our findings
in clinical cancer samples is needed. Further investigations may
identify novel DP specific pathways that can be targeted for the
development of new treatments for breast and ovarian cancer.
Abbreviations:
|
CLL
|
chronic lymphocytic leukemia
|
|
DP
|
dipeptidyl peptidase
|
|
EOC
|
epithelial ovarian carcinoma
|
|
ERc
|
estrogen receptor
|
|
ESFT
|
Ewing’s sarcoma family of tumors
|
|
FAP
|
fibroblast activation protein
|
|
GLP
|
glucagon-like peptide
|
|
HER2
|
human epidermal growth factor receptor
2
|
|
NPY
|
neuropeptide Y
|
|
PEP
|
prolyl endopeptidase
|
|
SDF-1
|
stromal derived factor-1
|
Acknowledgements
We would like to thank Ms. Maressa
Bruhn and Dr Karen Shephard (Peter Macallum Cancer Centre,
Melbourne, Vic., Australia) for kindly providing us with the cells
used in this study. Authors would also like to thank Dr Mark
Gorrell (AW Morrow Gastroenteroogy and Liver Center, University of
Sydney, NSW, Australia) for kindly providing hybridoma cells
secreting the F19 monoclonal antibody.
References
|
1.
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Cho KR and Shih Ie M: Ovarian cancer. Annu
Rev Pathol. 4:287–313. 2009. View Article : Google Scholar
|
|
3.
|
Schnitt SJ: Classification and prognosis
of invasive breast cancer: from morphology to molecular taxonomy.
Mod Pathol. 23(Suppl 2): S60–S64. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Kurman RJ and Shih Ie M: Pathogenesis of
ovarian cancer: lessons from morphology and molecular biology and
their clinical implications. Int J Gynecol Pathol. 27:151–160.
2008.PubMed/NCBI
|
|
5.
|
Lambeir AM, Durinx C, Scharpe S and De
Meester I: Dipeptidyl-peptidase IV from bench to bedside: an update
on structural properties, functions, and clinical aspects of the
enzyme DPP IV. Crit Rev Clin Lab Sci. 40:209–294. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Bjelke JR, Christensen J, Nielsen PF,
Branner S, Kanstrup AB, Wagtmann N and Rasmussen HB: Dipeptidyl
peptidases 8 and 9: specificity and molecular characterization
compared with dipeptidyl peptidase IV. Biochem J. 396:391–399.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Ajami K, Pitman MR, Wilson CH, Park J,
Menz RI, Starr AE, Cox JH, Abbott CA, Overall CM and Gorrell MD:
Stromal cell-derived factors 1alpha and 1beta, inflammatory
protein-10 and interferon-inducible T cell chemo-attractant are
novel substrates of dipeptidyl peptidase 8. FEBS Lett. 582:819–825.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Keane FM, Nadvi NA, Yao TW and Gorrell MD:
Neuropeptide Y, B-type natriuretic peptide, substance P and peptide
YY are novel substrates of fibroblast activation protein-alpha.
FEBS J. 278:1316–1332. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Busso N, Wagtmann N, Herling C,
Chobaz-Peclat V, Bischof-Delaloye A, So A and Grouzmann E:
Circulating CD26 is negatively associated with inflammation in
human and experimental arthritis. Am J Pathol. 166:433–442. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Barbieri F, Bajetto A and Florio T: Role
of chemokine network in the development and progression of ovarian
cancer: a potential novel pharmacological target. J Oncol.
2010:4269562010. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Ali S and Lazennec G: Chemokines: novel
targets for breast cancer metastasis. Cancer Metastasis Rev.
26:401–420. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Balkwill F: Cancer and the chemokine
network. Nat Rev Cancer. 4:540–550. 2004. View Article : Google Scholar
|
|
13.
|
Mentlein R: Dipeptidyl-peptidase IV
(CD26): role in the inactivation of regulatory peptides. Regul
Pept. 85:9–24. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Sun YX, Pedersen EA, Shiozawa Y, Havens
AM, Jung Y, Wang J, Pienta KJ and Taichman RS: CD26/dipeptidyl
peptidase IV regulates prostate cancer metastasis by degrading
SDF-1/CXCL12. Clin Exp Metastasis. 25:765–776. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Arscott WT, La Bauve AE, May V and Wesley
UV: Suppression of neuroblastoma growth by dipeptidyl peptidase IV:
relevance of chemokine regulation and caspase activation. Oncogene.
28:479–491. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Lu C, Tilan JU, Everhart L, Czarnecka M,
Soldin SJ, Mendu DR, Jeha D, Hanafy J, Lee CK, Sun J,
Izycka-Swiezczewska E, Toretsky JA and Kitlinska J: Dipeptidyl
peptidases as survival factors in Ewing sarcoma family of tumors. J
Biol Chem. 286:27494–27505. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Yu DM, Wang XM, McCaughan GW and Gorrell
MD: Extra-enzymatic functions of the dipeptidyl peptidase
IV-related proteins DP8 and DP9 in cell adhesion, migration and
apoptosis. FEBS J. 273:2447–2460. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Wang XM, Yu DM, McCaughan GW and Gorrell
MD: Fibroblast activation protein increases apoptosis, cell
adhesion, and migration by the LX-2 human stellate cell line.
Hepatology. 42:935–945. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Sulda ML, Abbott CA and Hildebrandt M:
DPIV/CD26 and FAP in cancer: a tale of contradictions. Adv Exp Med
Biol. 575:197–206. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Johnson RC, Zhu D, Augustin-Voss HG and
Pauli BU: Lung endothelial dipeptidyl peptidase IV is an adhesion
molecule for lung-metastatic rat breast and prostate carcinoma
cells. J Cell Biol. 121:1423–1432. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Cheng HC, Abdel-Ghany M, Elble RC and
Pauli BU: Lung endothelial dipeptidyl peptidase IV promotes
adhesion and metastasis of rat breast cancer cells via tumor cell
surface-associated fibronectin. J Biol Chem. 273:24207–24215. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Cheng HC, Abdel-Ghany M, Zhang S and Pauli
BU: Is the Fischer 344/CRJ rat a protein-knock-out model for
dipeptidyl peptidase IV-mediated lung metastasis of breast cancer?
Clin Exp Metastasis. 17:609–615. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Kajiyama H, Kikkawa F, Suzuki T, Shibata
K, Ino K and Mizutani S: Prolonged survival and decreased invasive
activity attributable to dipeptidyl peptidase IV overexpression in
ovarian carcinoma. Cancer Res. 62:2753–2757. 2002.PubMed/NCBI
|
|
24.
|
Kajiyama H, Shibata K, Ino K, Mizutani S,
Nawa A and Kikkawa F: The expression of dipeptidyl peptidase IV
(DPPIV/CD26) is associated with enhanced chemosensitivity to
paclitaxel in epithelial ovarian carcinoma cells. Cancer Sci.
101:347–354. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Garin-Chesa P, Old LJ and Rettig WJ: Cell
surface glycoprotein of reactive stromal fibroblasts as a potential
antibody target in human epithelial cancers. Proc Natl Acad Sci
USA. 87:7235–7239. 1990. View Article : Google Scholar
|
|
26.
|
Chen D, Kennedy A, Wang JY, Zeng W, Zhao
Q, Pearl M, Zhang M, Suo Z, Nesland JM, Qiao Y, Ng AK, Hirashima N,
Yamane T, Mori Y, Mitsumata M, Ghersi G and Chen WT: Activation of
EDTA-resistant gelatinases in malignant human tumors. Cancer Res.
66:9977–9985. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Kraman M, Bambrough PJ, Arnold JN, Roberts
EW, Magiera L, Jones JO, Gopinathan A, Tuveson DA and Fearon DT:
Suppression of antitumor immunity by stromal cells expressing
fibroblast activation protein-alpha. Science. 330:827–830. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Kelly T, Kechelava S, Rozypal TL, West KW
and Korourian S: Seprase, a membrane-bound protease, is
overexpressed by invasive ductal carcinoma cells of human breast
cancers. Mod Pathol. 11:855–863. 1998.PubMed/NCBI
|
|
29.
|
Ariga N, Sato E, Ohuchi N, Nagura H and
Ohtani H: Stromal expression of fibroblast activation
protein/seprase, a cell membrane serine proteinase and gelatinase,
is associated with longer survival in patients with invasive ductal
carcinoma of breast. Int J Cancer. 95:67–72. 2001. View Article : Google Scholar
|
|
30.
|
Goodman JD, Rozypal TL and Kelly T:
Seprase, a membrane-bound protease, alleviates the serum growth
requirement of human breast cancer cells. Clin Exp Metastasis.
20:459–470. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Huang Y, Wang S and Kelly T: Seprase
promotes rapid tumor growth and increased microvessel density in a
mouse model of human breast cancer. Cancer Res. 64:2712–2716. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Kennedy A, Dong H, Chen D and Chen WT:
Elevation of seprase expression and promotion of an invasive
phenotype by collagenous matrices in ovarian tumor cells. Int J
Cancer. 124:27–35. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Abbott CA, Yu DM, Woollatt E, Sutherland
GR, McCaughan GW and Gorrell MD: Cloning, expression and
chromosomal localization of a novel human dipeptidyl peptidase
(DPP) IV homolog, DPP8. Eur J Biochem. 267:6140–6150. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Yu DM, Yao TW, Chowdhury S, Nadvi NA,
Osborne B, Church WB, McCaughan GW and Gorrell MD: The dipeptidyl
peptidase IV family in cancer and cell biology. FEBS J.
277:1126–1144. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Dubois V, Van Ginneken C, De Cock H,
Lambeir AM, Van der Veken P, Augustyns K, Chen X, Scharpe S and De
Meester I: Enzyme activity and immunohistochemical localization of
dipeptidyl Peptidase 8 and 9 in male reproductive tissues. J
Histochem Cytochem. 57:531–541. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Yao TW, Kim WS, Yu DM, Sharbeen G,
McCaughan GW, Choi KY, Xia P and Gorrell MD: A novel role of
dipeptidyl peptidase 9 in epidermal growth factor signaling. Mol
Cancer Res. 9:948–959. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Sulda ML, Abbott CA, Macardle PJ, Hall RK
and Kuss BJ: Expression and prognostic assessment of dipeptidyl
peptidase IV and related enzymes in B-cell chronic lymphocytic
leukemia. Cancer Biol Ther. 10:180–189. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Yu DM, Ajami K, Gall MG, Park J, Lee CS,
Evans KA, McLaughlin EA, Pitman MR, Abbott CA, McCaughan GW and
Gorrell MD: The in vivo expression of dipeptidyl peptidases 8 and
9. J Histochem Cytochem. 57:1025–1040. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Stremenova J, Mares V, Lisa V, Hilser M,
Krepela E, Vanickova Z, Syrucek M, Soula O and Sedo A: Expression
of dipeptidyl peptidase-IV activity and/or structure homologs in
human meningiomas. Int J Oncol. 36:351–358. 2010.PubMed/NCBI
|
|
40.
|
Stremenova J, Krepela E, Mares V, Trim J,
Dbaly V, Marek J, Vanickova Z, Lisa V, Yea C and Sedo A: Expression
and enzymatic activity of dipeptidyl peptidase-IV in human
astrocytic tumours are associated with tumour grade. Int J Oncol.
31:785–792. 2007.PubMed/NCBI
|
|
41.
|
Soule HD, Vazguez J, Long A, Albert S and
Brennan M: A human cell line from a pleural effusion derived from a
breast carcinoma. J Natl Cancer Inst. 51:1409–1416. 1973.PubMed/NCBI
|
|
42.
|
Zajchowski DA, Bartholdi MF, Gong Y,
Webster L, Liu HL, Munishkin A, Beauheim C, Harvey S, Ethier SP and
Johnson PH: Identification of gene expression profiles that predict
the aggressive behavior of breast cancer cells. Cancer Res.
61:5168–5178. 2001.PubMed/NCBI
|
|
43.
|
Crow MJ, Grant G, Provenzale JM and Wax A:
Molecular imaging and quantitative measurement of epidermal growth
factor receptor expression in live cancer cells using immunolabeled
gold nanoparticles. AJR Am J Roentgenol. 192:1021–1028. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Lau KM, Mok SC and Ho SM: Expression of
human estrogen receptor-alpha and -beta, progesterone receptor, and
androgen receptor mRNA in normal and malignant ovarian epithelial
cells. Proc Natl Acad Sci USA. 96:5722–5727. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Takai N, Jain A, Kawamata N, Popoviciu LM,
Said JW, Whittaker S, Miyakawa I, Agus DB and Koeffler HP: 2C4, a
monoclonal antibody against HER2, disrupts the HER kinase signaling
pathway and inhibits ovarian carcinoma cell growth. Cancer.
104:2701–2708. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46.
|
Xu F, Yu Y, Le XF, Boyer C, Mills GB and
Bast RC: The outcome of heregulin-induced activation of ovarian
cancer cells depends on the relative levels of HER-2 and HER-3
expression. Clin Cancer Res. 5:3653–3660. 1999.PubMed/NCBI
|
|
47.
|
Hua W, Christianson T, Rougeot C,
Rochefort H and Clinton GM: SKOV3 ovarian carcinoma cells have
functional estrogen receptor but are growth-resistant to estrogen
and antiestrogens. J Steroid Biochem Mol Biol. 55:279–289. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
48.
|
Hu G, Liu W, Mendelsohn J, Ellis LM,
Radinsky R, Andreeff M and Deisseroth AB: Expression of epidermal
growth factor receptor and human papillomavirus E6/E7 proteins in
cervical carcinoma cells. J Natl Cancer Inst. 89:1271–1276. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
49.
|
Monje P and Boland R: Expression and
cellular localization of naturally occurring beta estrogen
receptors in uterine and mammary cell lines. J Cell Biochem.
86:136–144. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50.
|
Lossos IS, Czerwinski DK, Wechser MA and
Levy R: Optimization of quantitative real-time RT-PCR parameters
for the study of lymphoid malignancies. Leukemia. 17:789–795. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
51.
|
Maes MB, Dubois V, Brandt I, Lambeir AM,
Van der Veken P, Augustyns K, Cheng JD, Chen X, Scharpe S and De
Meester I: Dipeptidyl peptidase 8/9-like activity in human
leukocytes. J Leukocyte Biol. 81:1252–1257. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52.
|
Abbott CA and Gorrell MD: The family of
CD26/DPIV related ectopeptidases. Ectopeptidases:
CD13/Aminopeptidase N and CD26/Dipeptidylpeptidase IV in Medicine
and Biology. Langner J and Ansorge S: Kluwer/Plenum; New York: pp.
171–195. 2002, View Article : Google Scholar
|
|
53.
|
Balaziova E, Busek P, Stremenova J,
Sromova L, Krepela E, Lizcova L and Sedo A: Coupled expression of
dipeptidyl peptidase-IV and fibroblast activation protein-alpha in
transformed astrocytic cells. Mol Cell Biochem. 354:283–289. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
54.
|
Ajami K, Abbott CA, McCaughan GW and
Gorrell MD: Dipeptidyl peptidase 9 has two forms, a broad tissue
distribution, cytoplasmic localization and DPIV-like peptidase
activity. Biochim Biophys Acta. 1679:18–28. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
55.
|
Pitman MR, Menz RI and Abbott CA:
Hydrophilic residues surrounding the S1 and S2 pockets contribute
to dimerisation and catalysis in human dipeptidyl peptidase 8
(DP8). Biol Chem. 391:959–972. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
56.
|
Bank U, Heimburg A, Wohlfarth A, Koch G,
Nordhoff K, Julius H, Helmuth M, Breyer D, Reinhold D, Tager M and
Ansorge S: Outside or inside: role of the subcellular localization
of DP4-like enzymes for substrate conversion and inhibitor effects.
Biol Chem. 392:169–187. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
57.
|
Ansorge S, Nordhoff K, Bank U, Heimburg A,
Julius H, Breyer D, Thielitz A, Reinhold D and Tager M: Novel
aspects of cellular action of dipeptidyl peptidase IV/CD26. Biol
Chem. 392:153–168. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
58.
|
Kikkawa F, Kajiyama H, Ino K, Shibata K
and Mizutani S: Increased adhesion potency of ovarian carcinoma
cells to mesothelial cells by overexpression of dipeptidyl
peptidase IV. Int J Cancer. 105:779–783. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
59.
|
Huang Y, Simms AE, Mazur A, Wang S, Leon
NR, Jones B, Aziz N and Kelly T: Fibroblast activation
protein-alpha promotes tumor growth and invasion of breast cancer
cells through non-enzymatic functions. Clin Exp Metastasis.
28:567–579. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60.
|
Aertgeerts K, Levin I, Shi L, Snell GP,
Jennings A, Prasad GS, Zhang Y, Kraus ML, Salakian S, Sridhar V,
Wijnands R and Tennant MG: Structural and kinetic analysis of the
substrate specificity of human fibroblast activation protein alpha.
J Biol Chem. 280:19441–19444. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
61.
|
Chiravuri M, Agarraberes F, Mathieu SL,
Lee H and Huber BT: Vesicular localization and characterization of
a novel post-proline-cleaving aminodipeptidase, quiescent cell
proline dipeptidase. J Immunol. 165:5695–5702. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
62.
|
Tang HK, Tang HY, Hsu SC, Chu YR, Chien
CH, Shu CH and Chen X: Biochemical properties and expression
profile of human prolyl dipeptidase DPP9. Arch Biochem Biophys.
485:120–127. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
63.
|
Leiting B, Pryor KD, Wu JK, Marsilio F,
Patel RA, Craik CS, Ellman JA, Cummings RT and Thornberry NA:
Catalytic properties and inhibition of proline-specific dipeptidyl
peptidases II, IV and VII. Biochem J. 371:525–532. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
64.
|
Maes MB, Martinet W, Schrijvers DM, Van
der Veken P, De Meyer GR, Augustyns K, Lambeir AM, Scharpe S and De
Meester I: Dipeptidyl peptidase II and leukocyte cell death.
Biochem Pharmacol. 72:70–79. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65.
|
Danilova O, Li B, Szardenings AK, Huber BT
and Rosenblum JS: Synthesis and activity of a potent, specific
azabicyclo[3.3.0]-octane-based DPP II inhibitor. Bioorg Med Chem
Lett. 17:507–510. 2007.
|
|
66.
|
Hui M and Hui KS: A novel aminopeptidase
with highest preference for lysine. Neurochem Res. 31:95–102. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
67.
|
Claperon C, Banegas-Font I, Iturrioz X,
Rozenfeld R, Maigret B and Llorens-Cortes C: Identification of
threonine 348 as a residue involved in aminopeptidase A substrate
specificity. J Biol Chem. 284:10618–10626. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
68.
|
Fukasawa KM, Hirose J, Hata T and Ono Y:
Aspartic acid 405 contributes to the substrate specificity of
aminopeptidase B. Biochemistry. 45:11425–11431. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
69.
|
Drag M, Bogyo M, Ellman JA and Salvesen
GS: Aminopeptidase fingerprints, an integrated approach for
identification of good substrates and optimal inhibitors. J Biol
Chem. 285:3310–3318. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
70.
|
Tenorio-Laranga J, Venalainen JI, Mannisto
PT and Garcia-Horsman JA: Characterization of membrane-bound prolyl
endopeptidase from brain. FEBS J. 275:4415–4427. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
71.
|
Edosada CY, Quan C, Tran T, Pham V,
Wiesmann C, Fairbrother W and Wolf BB: Peptide substrate profiling
defines fibroblast activation protein as an endopeptidase of strict
Gly(2)-Pro(1)-cleaving specificity. FEBS Lett. 580:1581–1586. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
72.
|
Cheng JQ, Lindsley CW, Cheng GZ, Yang H
and Nicosia SV: The Akt/PKB pathway: molecular target for cancer
drug discovery. Oncogene. 24:7482–7492. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
73.
|
Hernandez-Aya LF and Gonzalez-Angulo AM:
Targeting the phosphatidylinositol 3-kinase signaling pathway in
breast cancer. Oncologist. 16:404–414. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
74.
|
Zagouri F, Dimopoulos MA, Bournakis E and
Papadimitriou CA: Molecular markers in epithelial ovarian cancer:
their role in prognosis and therapy. Eur J Gynaecol Oncol.
31:268–277. 2010.PubMed/NCBI
|
|
75.
|
Mazzoletti M and Broggini M: PI3K/AKT/mTOR
inhibitors in ovarian cancer. Curr Med Chem. 17:4433–4447. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
76.
|
Dickson RB, Thompson EW and Lippman ME:
Regulation of proliferation, invasion and growth factor synthesis
in breast cancer by steroids. J Steroid Biochem Mol Biol.
37:305–316. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
77.
|
Kim HJ, Cui X, Hilsenbeck SG and Lee AV:
Progesterone receptor loss correlates with human epidermal growth
factor receptor 2 overexpression in estrogen receptor-positive
breast cancer. Clin Cancer Res. 12:S1013–S1018. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
78.
|
Yager JD and Davidson NE: Estrogen
carcinogenesis in breast cancer. N Engl J Med. 354:270–282. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
79.
|
Oakman C, Viale G and Di Leo A: Management
of triple negative breast cancer. Breast. 19:312–321. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
80.
|
Rao BR, Slotman BJ, Geldof AA and Dinjens
WN: Correlation between tumor histology, steroid receptor status,
and adenosine deaminase complexing protein immunoreactivity in
ovarian cancer. Int J Gynecol Pathol. 9:47–54. 1990. View Article : Google Scholar
|